Line × Tester Analysis for Morphological and Fruit Biochemical Traits in Eggplant (Solanum melongena L.) Using Wild Relatives as Testers


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growing Conditions
2.2. Characterisation and Data Analysis
3. Results
3.1. Analysis of Variance for Line, Tester, and L × T Effects and GCA and SCA Estimates
3.2. Contribution to Total Variance
3.3. GCA and SCA
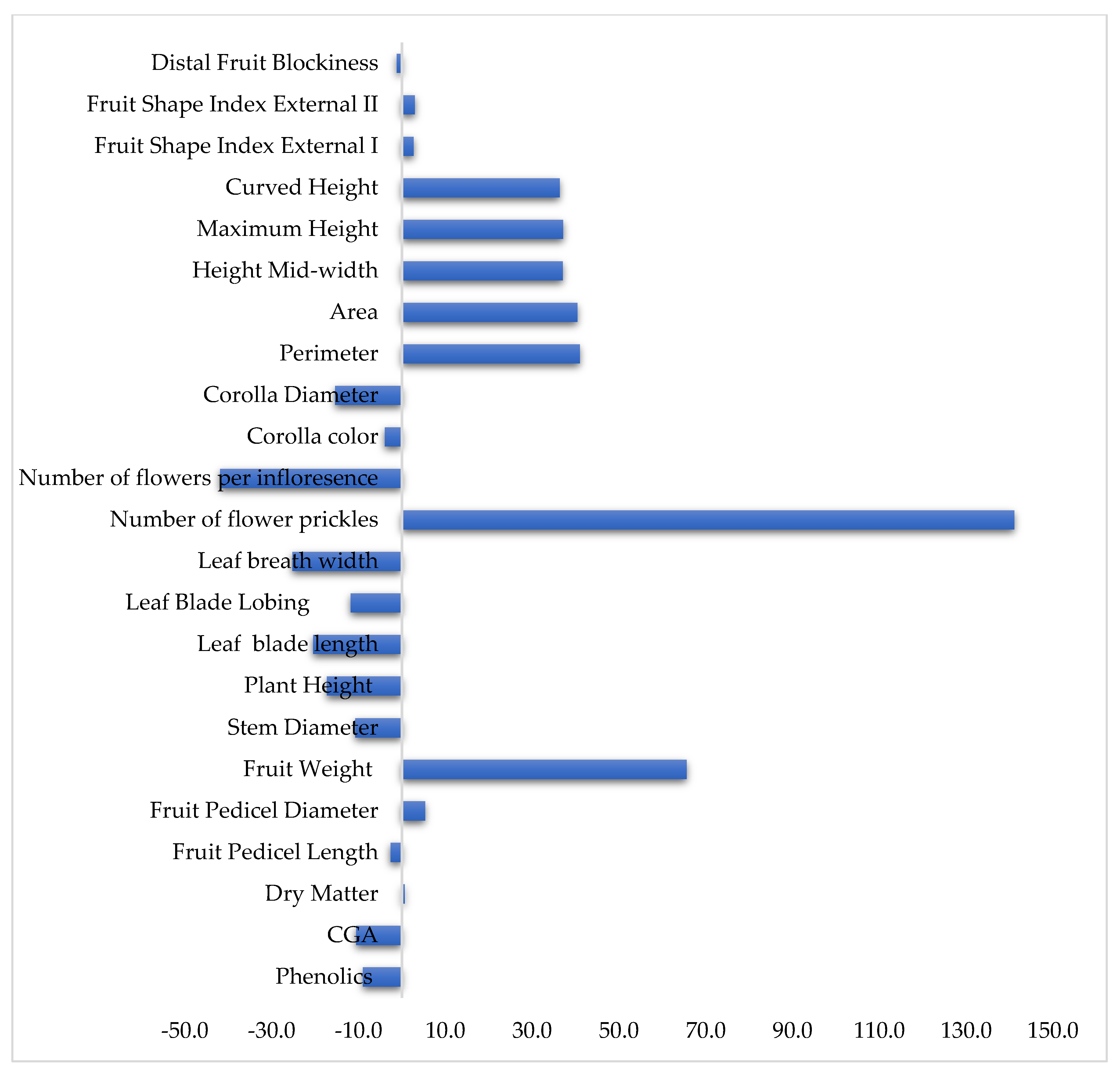
3.4. Heterosis
4. Discussion
Supplementary Materials
Funding
Acknowledgments
Conflicts of Interest
References
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food Security: The Challenge of Feeding 9 Billion People. Science 2010. [Google Scholar] [CrossRef] [PubMed]
- Bisbis, M.B.; Gruda, N.; Blanke, M. Potential impacts of climate change on vegetable production and product quality—A review. J. Clean. Product. 2018, 170, 1602–1620. [Google Scholar] [CrossRef]
- Rotino, G.L.; Sala, T.; Toppino, L. Eggplant. In Alien Gene Transfer in Crop Plants, Volume 2; Springer: New York, NY, USA, 2014; pp. 381–409. ISBN 978-1-4614-9571-0. [Google Scholar]
- Plazas, M.; Vilanova, S.; Gramazio, P.; Rodriguez-Burruezo, A.; Fita, A.; Herraiz, F.; Ranil, R.; Fonseka, R.; Niran, L.; Fonseka, H.; et al. Interspecific Hybridization between Eggplant and Wild Relatives from Different Genepools. J. Am. Soc. Hortic. Sci. 2016, 141, 34–44. [Google Scholar] [CrossRef]
- Meyer, R.S.; Whitaker, B.D.; Little, D.P.; Wu, S.-B.; Kennelly, E.J.; Long, C.-L.; Litt, A. Parallel reductions in phenolic constituents resulting from the domestication of eggplant. Phytochemistry 2015, 115, 194–206. [Google Scholar] [CrossRef] [PubMed]
- Knapp, S.; Vorontsova, M.S.; Prohens, J. Wild relatives of the eggplant (Solanum melongena L.: Solanaceae): New understanding of species names in a complex group. PLoS ONE 2013, 8, e57039. [Google Scholar] [CrossRef]
- Zhang, H.; Mittal, N.; Leamy, L.J.; Barazani, O.; Song, B. Back into the wild—Apply untapped genetic diversity of wild relatives for crop improvement. Evol. Appl. 2016, 10, 5–24. [Google Scholar] [CrossRef]
- Gramazio, P. Genetics and genomics of cultivated eggplants and wild relatives. 2018. [Google Scholar] [CrossRef]
- Plazas, M.; Andújar, I.; Vilanova, S.; Hurtado, M.; Gramazio, P.; Herraiz, F.J.; Prohens, J. Breeding for Chlorogenic Acid Content in Eggplant: Interest and Prospects. Not. Bot. Horti. Agrobo. 2013, 41, 26–35. [Google Scholar] [CrossRef]
- Kaushik, P.; Andújar, I.; Vilanova, S.; Plazas, M.; Gramazio, P.; Herraiz, F.J.; Brar, N.S.; Prohens, J. Breeding Vegetables with Increased Content in Bioactive Phenolic Acids. Molecules 2015, 20, 18464–18481. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, P.; Gramazio, P.; Vilanova, S.; Raigón, M.D.; Prohens, J.; Plazas, M. Phenolics content, fruit flesh colour and browning in cultivated eggplant, wild relatives and interspecific hybrids and implications for fruit quality breeding. Food Res. Int. 2017, 102, 392–401. [Google Scholar] [CrossRef]
- Ranil, R.H.G.; Prohens, J.; Aubriot, X.; Niran, H.M.L.; Plazas, M.; Fonseka, R.M.; Vilanova, S.; Fonseka, H.H.; Gramazio, P.; Knapp, S. Solanum insanum L. (subgenus Leptostemonum Bitter, Solanaceae), the neglected wild progenitor of eggplant (S. melongena L.): A review of taxonomy, characteristics and uses aimed at its enhancement for improved eggplant breeding. Genet. Resour. Crop. Evol. 2017, 64, 1707–1722. [Google Scholar] [CrossRef]
- Kaushik, P.; Prohens, J.; Vilanova, S.; Gramazio, P.; Plazas, M. Phenotyping of Eggplant Wild Relatives and Interspecific Hybrids with Conventional and Phenomics Descriptors Provides Insight for Their Potential Utilization in Breeding. Front. Plant Sci. 2016, 7, 677. [Google Scholar] [CrossRef] [PubMed]
- Hurtado, M.; Vilanova, S.; Plazas, M.; Gramazio, P.; Prohens, J. Development of breeding programmes in eggplant with different objectives and approaches: Three examples of use of primary genepool diversity. Acta Hortic. 2015. [Google Scholar] [CrossRef]
- Goleniowski, M.; Bonfill, M.; Cusido, R.; Palazón, J. Phenolic Acids. In Natural Products; Springer: Berlin, Heidelberg, 2013; pp. 1951–1973. ISBN 978-3-642-22143-9. [Google Scholar]
- Kempthorne, O. An Introduction to Genetic Statistics; John Wiley and Sons, Inc.: New York, NY, USA, 1957. [Google Scholar]
- Mumtaz, A.; Zafar, F.; Saifulmalook; Shehzad, A. A Review on Mating Designs. Nat. Sci. 2015, 13, 98–105. [Google Scholar]
- Singh, R.K.; Pooni, H.S.; Singh, M.; Bandopadhyaya, A. Mating Designs and Their Implications for Plant Breeding. In Plant Breeding; Springer: Dordrecht, The Netherlands, 2004; pp. 523–534. ISBN 978-94-010-3773-0. [Google Scholar]
- Zhu, J.; Weir, B.S. Analysis of cytoplasmic and maternal effects. II. Genetic models for triploid endosperms. Theoret. Appl. Genet. 1994, 89, 160–166. [Google Scholar] [CrossRef]
- Fasahat, P.; Rajabi, A.; Rad, J.M.; Derera, J. Principles and Utilization of Combining Ability in Plant Breeding. Biomet. Biostat. Int. J. 2016, 4, 1–24. [Google Scholar] [CrossRef]
- Kaushik, P.; Plazas, M.; Prohens, J.; Vilanova, S.; Gramazio, P. Diallel genetic analysis for multiple traits in eggplant and assessment of genetic distances for predicting hybrids performance. PLoS ONE 2018, 13, e0199943. [Google Scholar] [CrossRef]
- Kaushik, P.; Dhaliwal, M.; Kaushik, P.; Dhaliwal, M.S. Diallel Analysis for Morphological and Biochemical Traits in Tomato Cultivated under the Influence of Tomato Leaf Curl Virus. Agronomy 2018, 8, 153. [Google Scholar] [CrossRef]
- Sambandam, C.N. Heterosis in Eggplant (Solanum melongena Linn.): Prospects and Problems in Commercial Production of Hybrid Seeds. Econom. Bot. 1962, 16, 71–76. [Google Scholar] [CrossRef]
- Tyagi, V.; Dhillon, S.; Kaushik, P.; Kaur, G. Characterization for Drought Tolerance and Physiological Efficiency in Novel Cytoplasmic Male Sterile Sources of Sunflower (Helianthus annuus L.). Agronomy 2018, 8, 232. [Google Scholar] [CrossRef]
- Birchler, J.A.; Yao, H.; Chudalayandi, S.; Vaiman, D.; Veitia, R.A. Heterosis. Plant. Cell 2010. [Google Scholar] [CrossRef] [PubMed]
- Holland, J.B. Genetic architecture of complex traits in plants. Curr. Opin. Plant Biol. 2007, 10, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Dhaka, S.K.; Kaushik, R.A.; Jat, L.; Choudhary, R. Heterosis Breeding in Eggplant: A Review. J. Pharm. Phytochem. 2017, 6, 181–185. [Google Scholar]
- Vilanova, S.; Manzur, J.P.; Prohens, J. Development and characterization of genomic simple sequence repeat markers in eggplant and their application to the study of diversity and relationships in a collection of different cultivar types and origins. Mol. Breeding 2012, 30, 647–660. [Google Scholar] [CrossRef]
- Van der Weerden, G.M.; Barendse, G.W.M. A Web-Based Searchable Database Developed for the Eggnet Project and Applied to the Radboud University Solanaceae Database. Acta Hort. 2007. [Google Scholar] [CrossRef]
- Descriptors for eggplant/Descripteurs pour l’aubergine. Available online: https://www.bioversityinternational.org/e-library/publications/detail/descriptors-for-eggplantdescripteurs-pour-laubergine/ (accessed on 23 June 2018).
- Rodríguez, G.R.; Moyseenko, J.B.; Robbins, M.D.; Morejón, N.H.; Francis, D.M.; van der Knaap, E. Tomato Analyzer: A useful software application to collect accurate and detailed morphological and colorimetric data from two-dimensional objects. J. Vis. Exp. 2010. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Rodríguez, F.; Alvarado, G.; Pacheco, Á.; Crossa, J.; Burgueño, J. AGD-R (Analysis of Genetic Designs with R for Windows) Version 4.0. Available online: https://data.cimmyt.org/dataset.xhtml?persistentId=hdl: 11529/10202 (accessed on 15 July 2018).
- Xu, Y.; Li, P.; Zou, C.; Lu, Y.; Xie, C.; Zhang, X.; Prasanna, B.M.; Olsen, M.S. Enhancing genetic gain in the era of molecular breeding. J. Exp. Bot. 2017, 68, 2641–2666. [Google Scholar] [CrossRef]
- Govindaraj, M.; Vetriventhan, M.; Srinivasan, M. Importance of Genetic Diversity Assessment in Crop Plants and Its Recent Advances: An Overview of Its Analytical Perspectives. Available online: https://www.hindawi.com/journals/gri/2015/431487/ (accessed on 20 October 2018).
- Yadav, S.K.; Singh, B.K.; Baranwal, D.K.; Solankey, S.S. Genetic study of heterosis for yield and quality components in tomato (Solanum lycopersicum). AJAR 2013, 8, 5585–5591. [Google Scholar] [CrossRef]
- Sood, S.; Kumar, N. Heterotic expression for fruit yield and yield components in intervarietal hybrids of sweet pepper (Capsicum annuum L. var. grossum Sendt.). SABRAO J. Breed. Genet. 2010, 42, 106–116. [Google Scholar]
- Chattopadhyay, A.; Seth, T.; Dutta, S.; Ghosh, P.P.; Chattopadhyay, S.B.; Majumder, D.; Hazra, P. Breeding eggplant for higher productivity and bacterial wilt tolerance. Int. J. Veg. Sci. 2012, 18, 376–392. [Google Scholar] [CrossRef]
- Evans, D.M.; Gillespie, N.A.; Martin, N.G. Biometrical genetics. Biol. Psychol. 2002, 61, 33–51. [Google Scholar] [CrossRef]
- Guimarães, L.J.M.; Miranda, G.V.; DeLima, R.O.; Maia, C.; de Oliveira, L.R.; de Souza, L.V. Performance of testers with different genetic structure for evaluation of maize inbred lines. Ciência Rural 2012, 42, 770–776. [Google Scholar] [CrossRef]
- Relative Values of Three Different Testers in Evaluating Combining Ability of New Maize Inbred Lines—SciAlert Responsive Version. Available online: https://scialert.net/fulltextmobile/?doi=ijpbg.2014.57.65 (accessed on 20 October 2018).
- Taher, D.; Solberg, S.Ø.; Prohens, J.; Chou, Y.; Rakha, M.; Wu, T. World Vegetable Center Eggplant Collection: Origin, Composition, Seed Dissemination and Utilization in Breeding. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef]
- Liu, J.; Yang, Y.; Zhou, X.; Bao, S.; Zhuang, Y. Genetic diversity and population structure of worldwide eggplant (Solanum melongena L.) germplasm using SSR markers. Genet. Resour. Crop Evol 2018, 65, 1663–1670. [Google Scholar] [CrossRef]
- Naegele, R.P.; Boyle, S.; Quesada-Ocampo, L.M.; Hausbeck, M.K. Genetic Diversity, Population Structure, and Resistance to Phytophthora capsici of a Worldwide Collection of Eggplant Germplasm. PLoS ONE 2014, 9, e95930. [Google Scholar] [CrossRef] [PubMed]
- Mistry, C.B.; Kathiria, K.; Sabolu, S.; Kumar, S. Heritability and gene effects for yield related quantitative traits in eggplant. Ann. Agr. Sci. 2016, 61. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Species | Accession Code | Germplasm Collection Code | Country of Origin | Interspecific Hybrids | |
|---|---|---|---|---|---|
| With MEL3 | With MEL4 | ||||
| Cultivated Eggplant | |||||
| S. melongena | MEL3 | BBS-175 | Ivory Coast | ||
| MEL4 | 7145 | Sri Lanka | |||
| Wild primary genepool (GP1) | |||||
| S. insanum | INS1 | SLKINS-1 | Sri Lanka | MEL3 × INS1 | MEL4 × INS1 |
| INS2 | SLKINS-1 | Sri Lanka | MEL3 × INS2 | MEL4 × INS2 | |
| Wild secondary genepool (GP2) | |||||
| S. anguivi | ANG1 | BBS119 | Ivory Coast | MEL3 × ANG1 | MEL4 × ANG1 |
| S. lichtensteinii | LIC2 | MM677 | Iran | MEL3 × LIC2 | MEL4 × LIC2 |
| Traits/Descriptors | Scale |
|---|---|
| Biochemical Traits | |
| Phenolics | mg/g |
| CGA | mg/g |
| Dry Matter | % |
| Morphological Traits | |
| Fruit Pedicel Length | mm |
| Fruit Pedicel Diameter | mm |
| Fruit Weight | g |
| Stem Diameter | mm |
| Plant Height | cm |
| Leaf blade length | cm |
| Leaf Blade Lobing | 1 = Very weak (none); 9 = Very Strong |
| Leaf Blade Width | cm |
| Number of Flower Prickles | 0 = None; 9 = Very many (>20) |
| Number of Flowers Per Inflorescence | - |
| Corolla Color | 1 = Greenish white; 9 = Bluish violet |
| Corolla Diameter | mm |
| Tomato Analyzer-Based Descriptors | |
| Perimeter | cm |
| Area | cm2 |
| Height Mid-Width | cm |
| Maximum Height | cm |
| Curved Height | cm |
| Fruit Shape Index External I | The ratio of maximum height to maximum width. |
| Fruit Shape Index External II | The ratio of height mid-width to width mid-height. |
| Distal Fruit Blockiness | The ratio of the width at the lower blockiness position to width mid-height. |
| Source of Variation | Replicates | Treatments | Parents | Lines | Testers | Lines vs Testers | Parents vs Hybrids | Hybrids | Lines | Testers | Lines X Testers | Error | s ² GCA | s ² SCA | GCA/SCA |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| d.f | 2 | 13 | 5 | 1 | 3 | 1 | 1 | 7 | 1 | 3 | 3 | 26 | |||
| Phenolics | 10.86 | 46.95 *** | 54.31 *** | 4.58 | 50.43*** | 11 5.67 | 15.15 | 46.25 *** | 1.69 | 0.96 | 106.40 *** | 4.84 | 0.07 | 53.20 | 0.001 |
| CGA | 0.06 | 1.56 *** | 0.59 | 0.07 | 0.27 | 2.11 ** | 0.97 | 2.35 *** | 1.15 | 1.29 | 3.81 *** | 0.24 | 0.05 | 1.91 | 0.025 |
| Dry Matter | 0.04 | 46.35 *** | 7.21 | 19.3 | 3.7 | 5.67 | 0.09 | 80.92 *** | 99.35 | 8.09 | 147.60 *** | 5.01 | 4.14 | 73.80 | 0.056 |
| Fruit Pedicel Length | 2.34 | 488.86 *** | 477.05 *** | 60.17 ** | 118.64 *** | 1969.14 *** | 5.98 | 566.28 *** | 598 | 3.5 | 1118.50 *** | 5.89 | 24.91 | 559.24 | 0.045 |
| Fruit Pedicel Diameter | 0.09 | 28.82 *** | 25.81 *** | 1.5 | 14.81 *** | 83.11 *** | 1.04 | 34.94 *** | 23.21 | 16.96 | 56.83 *** | 0.39 | 0.96 | 28.41 | 0.034 |
| Fruit Weight | 152.26 | 29,905.50 *** | 20,384.54 *** | 411.35 | 154.91 | 10,1046.60 *** | 16,454.45 *** | 38,627.77 *** | 56,326.5 | 15,950.7 | 55,405.25 *** | 558.62 | 2346.93 | 27,702.63 | 0.085 |
| Stem Diameter | 5.22 | 51.15 *** | 18.55 | 10.67 | 13.46 | 41.71 * | 78.77 ** | 70.49 *** | 93.02 | 61.89 | 71.57 ** | 9.82 | 3.87 | 35.78 | 0.108 |
| Plant Height | 16.67 | 1879.98 *** | 1715.83 *** | 32.67 | 948.75 *** | 5700.25 *** | 6396.44 *** | 1352.02 *** | 3762.52 | 742.32 | 1158.23 *** | 83.22 | 156.77 | 579.11 | 0.271 |
| Leaf Blade Length | 0.23 | 77.67 *** | 26.58 *** | 21.09 *** | 37.24 *** | 0.07 | 157.58 *** | 102.76 *** | 13.28 | 10.37 | 224.97 *** | 1.47 | 0.55 | 112.48 | 0.005 |
| Leaf Blade Lobing | 0.01 | 5.27 *** | 3.20 *** | 6.00 *** | 3.00 *** | 1.00 *** | 4.57 *** | 6.86 *** | 6 | 12 | 2.00 *** | 0.01 | 0.25 | 1.01 | 0.248 |
| Leaf Blade Width | 0.56 | 48.60 *** | 21.45 *** | 0.98 | 35.36 *** | 0.18 | 140.26 *** | 54.91 *** | 13.37 | 6.93 | 116.74 *** | 0.65 | 0.55 | 58.37 | 0.009 |
| Number of Flower Prickles | 0.87 | 16.17 *** | 11.30 *** | 1.5 | 17.00 *** | 4.00 * | 27.86 *** | 17.99 *** | 21.09 | 21.84 | 13.10 *** | 0.88 | 0.87 | 6.54 | 0.133 |
| Number of Flowers per Infloresence | 0.39 | 22.41 *** | 23.68 *** | 0.02 | 14.43 *** | 75.10 *** | 84.26 *** | 12.67 *** | 50.85 ** | 12.03* | 0.57 | 0.34 | 2.11 | 0.28 | 7.536 |
| Corolla Color | 0.07 | 8.40 *** | 11.60 *** | 6.00 *** | 16.00 *** | 4.00 *** | 0.45 | 7.23 *** | 0.38 | 10.38 | 6.38 *** | 0.07 | 0.02 | 3.18 | 0.005 |
| Corolla Diameter | 15.98 | 328.41 *** | 96.97 *** | 7.48 | 81.68 *** | 232.31 *** | 301.63 *** | 497.56 *** | 106.26 | 57.77 | 1067.77 *** | 6 | 4.42 | 533.88 | 0.008 |
| Perimeter | 0.45 | 277.04 *** | 335.48 *** | 5.66 | 2.33 | 1664.76 *** | 209.84 *** | 244.90 *** | 43.63 | 84.2 | 472.70 *** | 6.75 | 1.81 | 236.35 | 0.008 |
| Area | 23.02 | 877.33 *** | 1166.98 *** | 15.06 | 0.93 | 5817.05 *** | 328.38 * | 748.87 *** | 150.87 | 229.38 | 1467.70 *** | 56.89 | 6.28 | 733.84 | 0.009 |
| Height Mid-Width | 0.12 | 30.77 *** | 37.84 *** | 4.24 * | 0.19 | 184.40 *** | 17.70 *** | 27.60 ** | 1.01 | 4.52 | 59.52 *** | 0.8 | 0.04 | 29.75 | 0.001 |
| Maximum Height | 0.11 | 31.76 *** | 38.96 *** | 3.60 * | 0.2 | 190.61 *** | 18.42 *** | 28.52 *** | 1.16 | 4.92 | 61.25 *** | 0.8 | 0.05 | 30.62 | 0.002 |
| Curved Height | 0.05 | 31.23 *** | 36.90 *** | 3.39 | 0.19 | 180.55 *** | 19.47 *** | 28.88 *** | 1.66 | 5.19 | 61.63 *** | 0.83 | 0.07 | 30.81 | 0.002 |
| Fruit Shape Index External I | 0.01 | 0.30 *** | 0.14 *** | 0.19 *** | 0.05 * | 0.36 *** | 0.01 | 0.46 *** | 0.01 | 0.24 | 0.83 *** | 0.01 | 0.01 | 0.41 | 0.024 |
| Fruit Shape Index External II | 0.01 | 0.34 *** | 0.16 *** | 0.27 *** | 0.05 * | 0.39 *** | 0.01 | 0.52 *** | 0 | 0.28 | 0.93 *** | 0.01 | 0.01 | 0.46 | 0.022 |
| Distal Fruit Blockiness | 0.01 | 0.02 *** | 0.01 *** | 0.01 ** | 0.01 | 0.03 *** | 0 | 0.03 *** | 0.02 | 0.01 | 0.05 *** | 0.01 | 0.01 | 0.02 | 0.500 |
| Traits | Lines | Testers | L × T |
|---|---|---|---|
| Phenolics | 0.52 | 0.89 | 98.59 |
| CGA | 7.02 | 23.45 | 69.54 |
| Dry Matter | 17.54 | 4.29 | 78.17 |
| Fruit Pedicel Length | 15.09 | 0.26 | 84.65 |
| Fruit Pedicel Diameter | 9.49 | 20.8 | 69.71 |
| Fruit Weight | 20.83 | 17.7 | 61.47 |
| Stem Diameter | 18.85 | 37.63 | 43.52 |
| Plant Height | 39.76 | 23.53 | 36.71 |
| Leaf Blade Length | 1.85 | 4.32 | 93.83 |
| Leaf Blade Lobbing | 12.5 | 75.02 | 12.5 |
| Leaf Blade Width | 3.48 | 5.41 | 91.11 |
| Number of Flower Prickles | 16.75 | 52.05 | 31.2 |
| Number of Flowers Per Inflorescence | 57.36 | 40.71 | 1.94 |
| Corolla Color | 0.74 | 61.48 | 37.78 |
| Corolla Diameter | 3.05 | 4.98 | 91.97 |
| Perimeter | 2.54 | 14.73 | 82.72 |
| Area | 2.88 | 13.13 | 83.99 |
| Height Mid-Width | 0.52 | 7.02 | 92.46 |
| Maximum Height | 0.58 | 7.39 | 92.03 |
| Curved Height | 0.82 | 7.71 | 91.47 |
| Fruit Shape Index External I | 0.18 | 22.41 | 77.4 |
| Fruit Shape Index External II | 0.02 | 23.00 | 76.98 |
| Distal Fruit Blockiness | 8.26 | 15.84 | 75.9 |
| Lines | Testers | |||||
|---|---|---|---|---|---|---|
| Traits/Characters | MEL3 | MEL4 | INS1 | INS2 | ANG1 | LIC2 |
| Phenolics | −0.27 | 0.27 | −0.28 | 0.24 | 0.43 * | −0.4 |
| CGA | 0.22 | −0.22 | −0.44 | −0.23 | 0.63 * | 0.05 |
| Dry Matter | −2.03 ** | 2.03 ** | −0.46 | 1.45 | 0.3 | −1.29 |
| Fruit Pedicel Length | 4.99 *** | −4.99 *** | −0.53 | 0.71 | −0.78 | 0.6 |
| Fruit Pedicel Diameter | 0.98 *** | −0.98 *** | −0.82 *** | 2.39 *** | −1.45 *** | −0.12 |
| Fruit Weight | 48.45 *** | −48.45 *** | 26.96 * | 57.12 *** | −57.06 *** | −27.02 * |
| Stem Diameter | −1.97 | 1.97 | 4.24 * | −3.14 * | −1.68 | 0.57 |
| Plant Height | −12.52 *** | 12.52 *** | 16.6 *** | −6.9 * | −4.23 | −5.48 |
| Leaf Blade Length | −0.74 | 0.74 | 0.84 | 0.79 | −1.94 ** | 0.31 |
| Leaf Blade Lobing | −0.50 *** | 0.50 *** | −2.00 *** | 0.01 *** | 1.00 *** | 1.00 *** |
| Leaf Blade Width | −0.75 * | 0.75 * | −0.46 | 0.22 | −1.14 ** | 1.38 ** |
| Number of Flower Prickles | −0.94 * | 0.94 * | −2.81 *** | 1.19 * | 1.19 * | 0.44 |
| Number of Flowers Per Inflorescence | −1.46 *** | 1.46 *** | 0.47 | 1.38 *** | −1.97 *** | 0.12 |
| Corolla Color | 0.13 | −0.13 | −1.13 *** | −1.13 *** | 0.88 *** | 1.38 *** |
| Corolla Diameter | 2.1 * | −2.1 * | −3.54 * | 0.4 | −0.8 | 3.95 ** |
| Perimeter | 1.35 | −1.35 | −2.82 | 2.97 * | −3.63 * | 3.48 * |
| Area | 2.51 | −2.51 | −3.44 | 5.84 | −6.94 | 4.53 |
| Height Mid-Width | 0.21 | −0.21 | −0.32 | 0.03 | −0.88 | 1.17 * |
| Maximum Height | 0.22 | −0.22 | −0.35 | 0.04 | −0.91 | 1.22 * |
| Curved Height | 0.26 | −0.26 | −0.36 | 0.2 | −1.02 * | 1.18 * |
| Fruit Shape Index External I | −0.02 | 0.02 | 0.16 *** | −0.29 *** | 0.06 | 0.07 |
| Fruit Shape Index External II | −0.01 | 0.01 | 0.20 *** | −0.3 *** | 0.05 | 0.05 |
| Distal Fruit Blockiness | 0.03 | −0.03 | 0.04 | −0.02 | −0.05 * | 0.04 |
| Traits | Minimum | Maximum | Mid-Parent Heterosis |
|---|---|---|---|
| Phenolics | −49.35 | 49.35 | −8.96 |
| CGA | −44.73 | 44.73 | −10.52 |
| Dry Matter | −34.69 | 34.69 | 0.67 |
| Fruit Pedicel Length | −45.08 | 45.08 | −2.63 |
| Fruit Pedicel Diameter | −43.38 | 43.38 | 5.34 |
| Fruit Weight | −86.33 | 86.33 | 65.59 |
| Stem Diameter | −17.02 | 17.02 | −10.72 |
| Plant Height | −9.77 | 9.77 | −17.30 |
| Leaf Blade Length | −40.49 | 40.49 | −20.43 |
| Leaf Blade Lobing | −10.00 | 10 | −11.76 |
| Leaf Blade Width | −43.55 | 43.55 | −25.24 |
| Number of Flower Prickles | −73.36 | 73.36 | 141.07 |
| Number of Flowers Per Inflorescence | −11.52 | 11.52 | −41.90 |
| Corolla Color | −21.97 | 21.97 | −3.91 |
| Corolla Diameter | −55.35 | 55.35 | −15.34 |
| Perimeter | −60.53 | 60.53 | 40.99 |
| Area | −70.71 | 70.71 | 40.39 |
| Height Mid-Width | −78.14 | 78.14 | 37.05 |
| Maximum Height | −77.37 | 77.37 | 37.16 |
| Curved Height | −71.51 | 71.51 | 36.33 |
| Fruit Shape Index External I | −43.33 | 43.33 | 2.71 |
| Fruit Shape Index External II | −47.00 | 47 | 2.97 |
| Distal Fruit Blockiness | −17.57 | 17.57 | −1.18 |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaushik, P. Line × Tester Analysis for Morphological and Fruit Biochemical Traits in Eggplant (Solanum melongena L.) Using Wild Relatives as Testers. Agronomy 2019, 9, 185. https://doi.org/10.3390/agronomy9040185
Kaushik P. Line × Tester Analysis for Morphological and Fruit Biochemical Traits in Eggplant (Solanum melongena L.) Using Wild Relatives as Testers. Agronomy. 2019; 9(4):185. https://doi.org/10.3390/agronomy9040185
Chicago/Turabian StyleKaushik, Prashant. 2019. "Line × Tester Analysis for Morphological and Fruit Biochemical Traits in Eggplant (Solanum melongena L.) Using Wild Relatives as Testers" Agronomy 9, no. 4: 185. https://doi.org/10.3390/agronomy9040185
APA StyleKaushik, P. (2019). Line × Tester Analysis for Morphological and Fruit Biochemical Traits in Eggplant (Solanum melongena L.) Using Wild Relatives as Testers. Agronomy, 9(4), 185. https://doi.org/10.3390/agronomy9040185