Drought Stress Tolerance Screening of Elite American Breeding Rice Genotypes Using Low-Cost Pre-Fabricated Mini-Hoop Modules

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
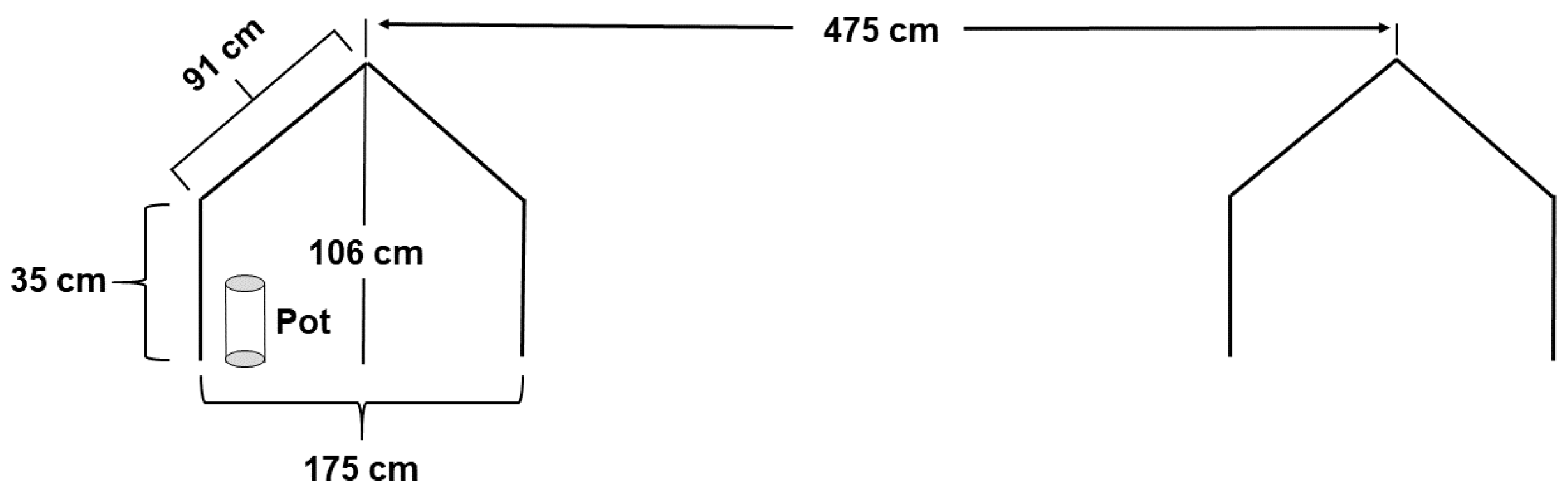
2.1. Germplasm Used and Experimental Setup
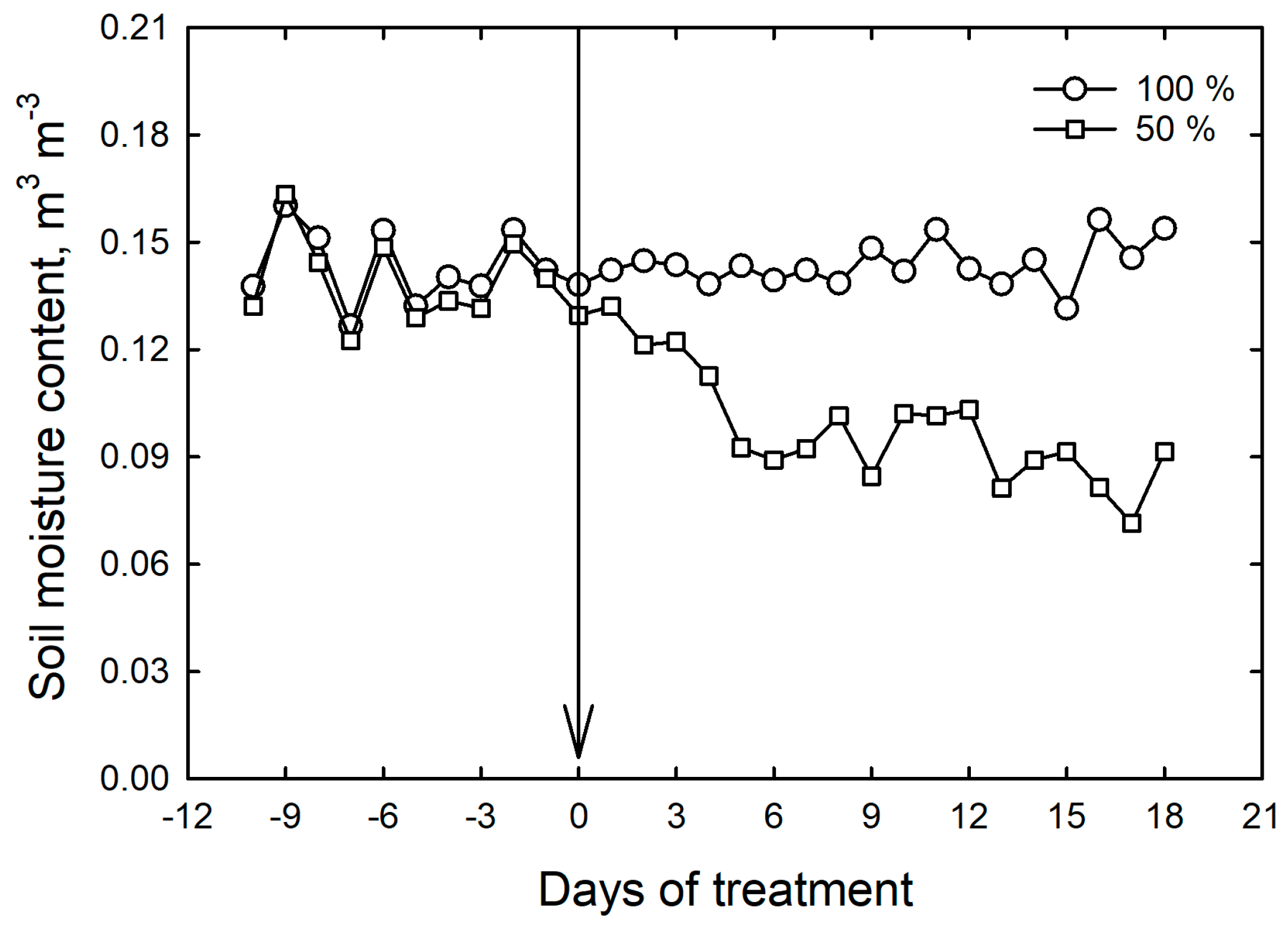
2.2. Drought Treatments
2.3. Measurements
2.3.1. Developmental Parameters
2.3.2. Physiological Parameters
2.3.3. Root Image Capture and Analysis
2.4. Data Analysis, Terminology and Drought Tolerance Indices
2.5. Drought Response Characterization
3. Results
3.1. Performance of Rice Genotypes and Interaction with Drought
3.2. Developmental Traits
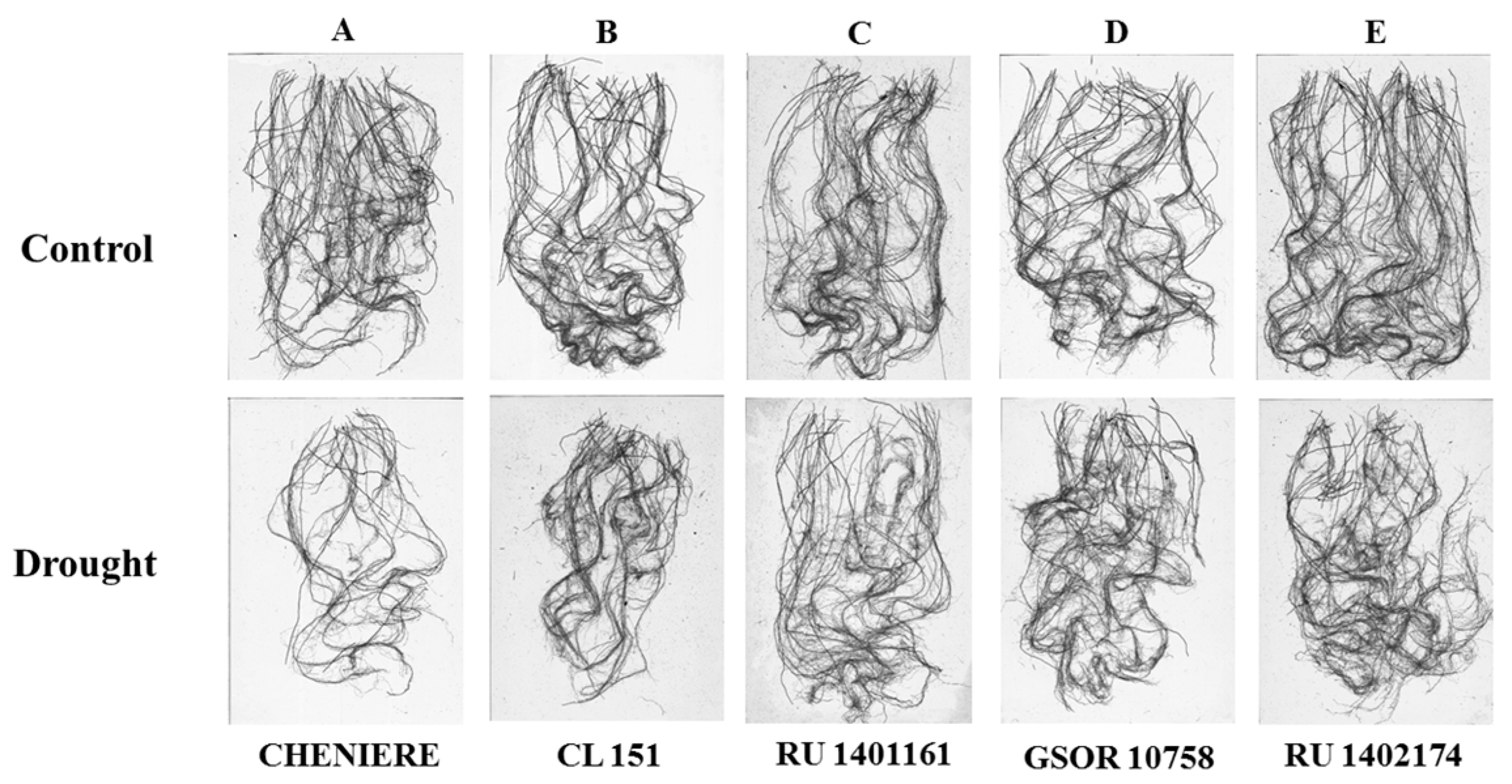
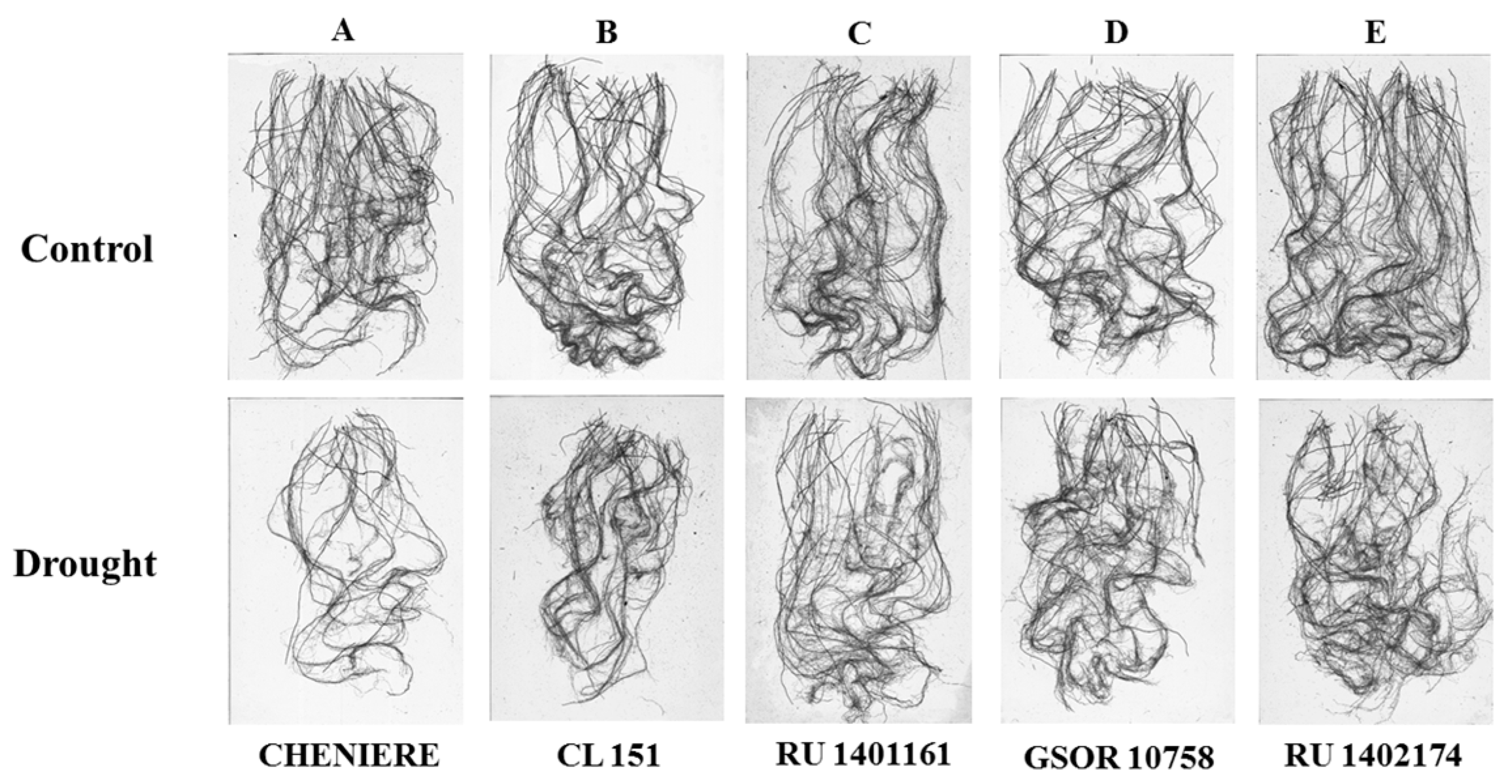
3.3. Root Traits
3.4. Physiological Traits
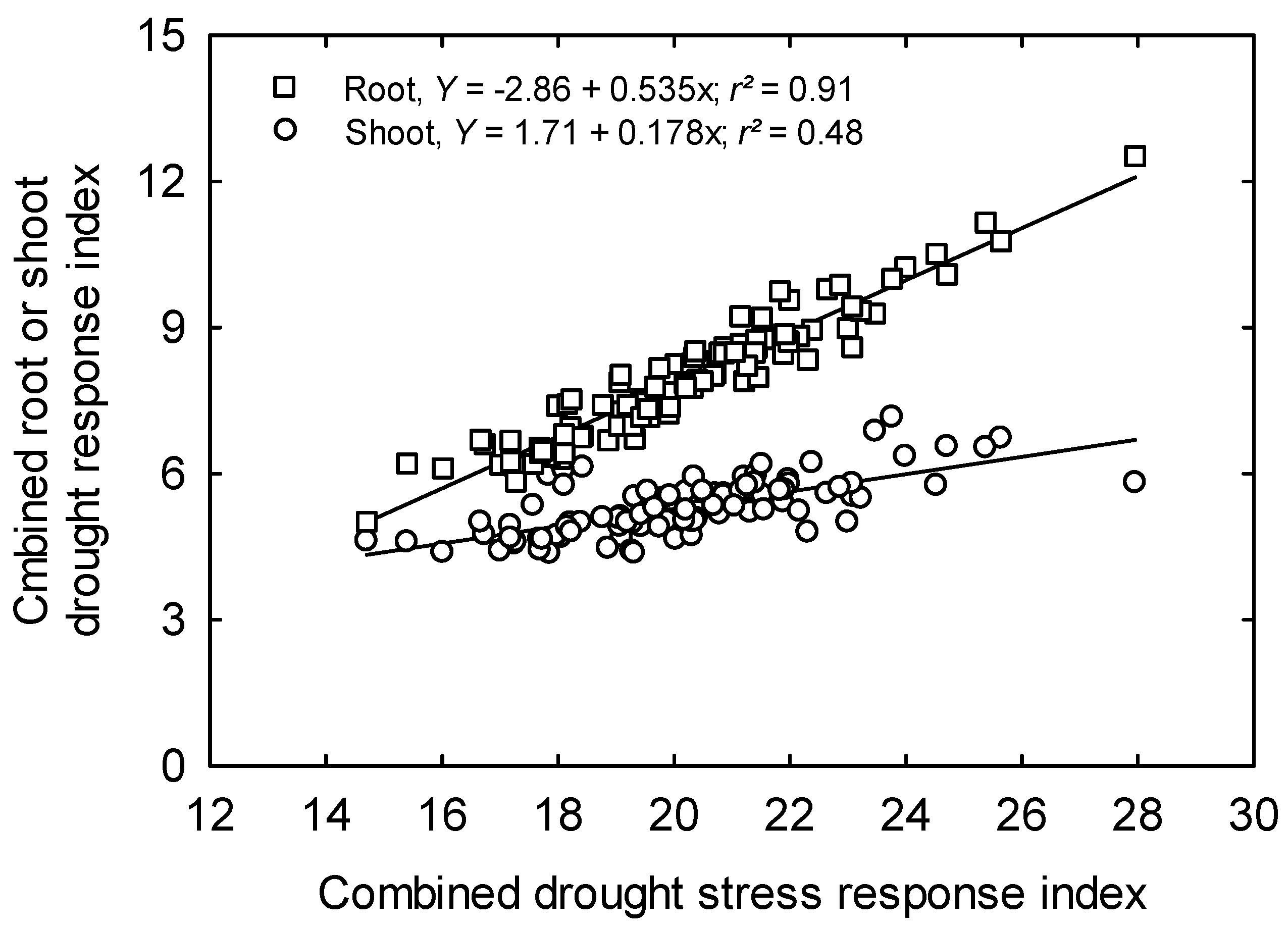
3.5. Root Traits Using Image Analysis
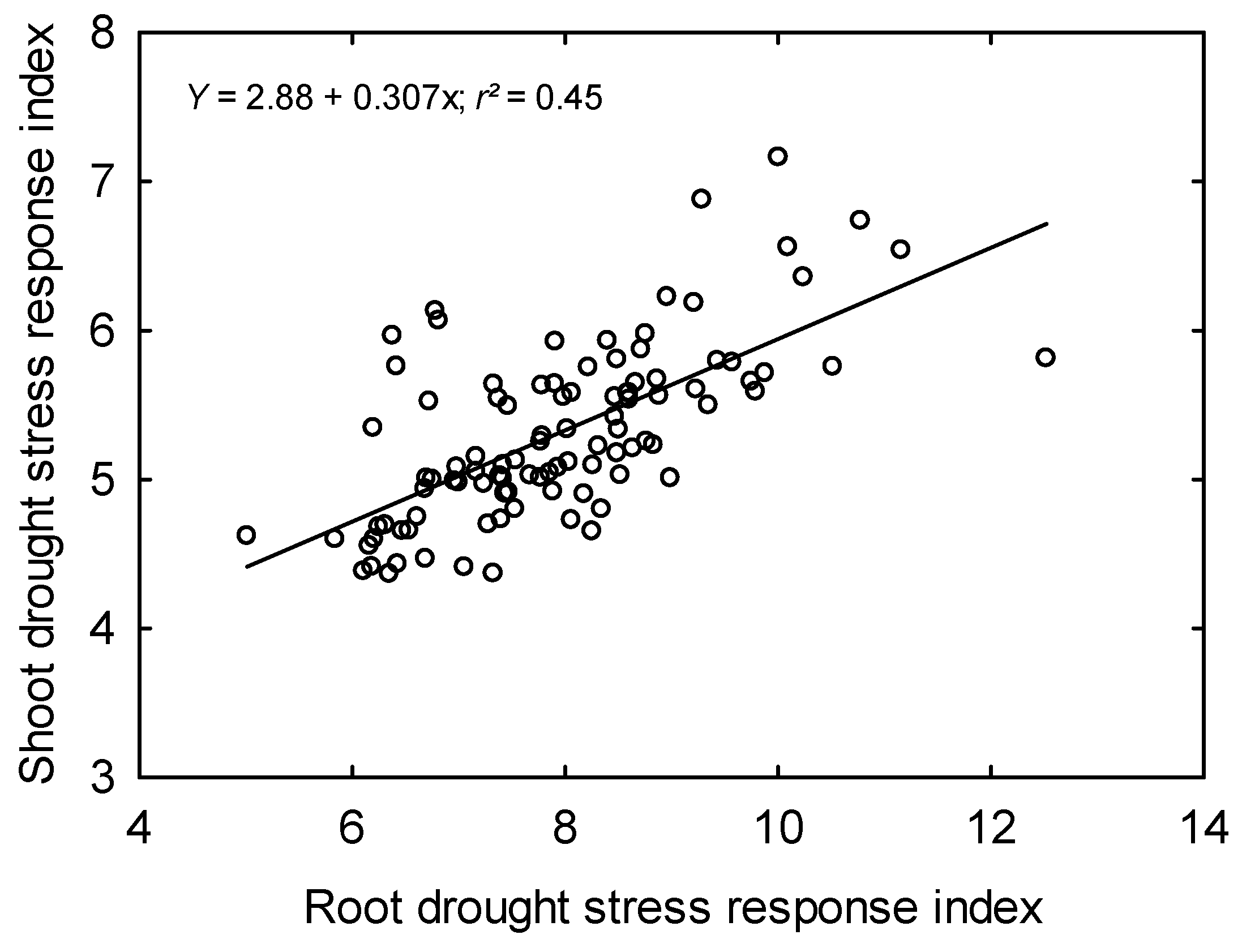
3.6. Classification of Rice Genotypes Based on Drought Response
4. Discussion
4.1. Performance of Rice Genotypes and Interaction with Drought
4.2. Developmental Traits
4.3. Root Traits
4.4. Physiological Traits
4.5. Classification of Rice Genotypes Based on Drought Response
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Song, Z.P.; Lu, B.R.; Zhu, Y.G.; Chen, J.K. Gene flow from cultivated rice to the wild species Oryza rufipogon under experimental field conditions. New Phytol. 2003, 157, 657–665. [Google Scholar] [CrossRef] [Green Version]
- Muthayya, S.; Sugimoto, J.D.; Montgomery, S.; Maberly, G.F. An overview of global rice production, supply, trade, and consumption. Ann. N. Y. Acad. Sci. 2014, 1324, 7–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gleick, P.H. Water in Crisis. A Guide to the World’s Fresh Water Resources; Oxford University Press: New York, NY, USA, 1993. [Google Scholar]
- Narciso, J.; Hossain, M. World Rice Statistics. IRRI, 2002. Available online: http://ricestat.irri.org:8080/wrs/ (accessed on 19 July 2017).
- Lafitte, H.R.; Yongsheng, G.; Yan, S.; Li, Z. Whole plant responses, key processes, and adaptation to drought stress: The case of rice. J. Exp. Bot. 2007, 58, 169–175. [Google Scholar] [CrossRef]
- Wu, N.; Guan, Y.; Shi, Y. Effect of water stress on physiological traits and yield in rice backcross lines after anthesis. Energy Procedia 2011, 5, 255–260. [Google Scholar] [CrossRef]
- Singh, S.; Prasad, S.; Yadav, V.; Kumar, A.; Jaiswal, B. Effect of drought stress on yield and yield components of rice (Oryza sativa L.) genotypes. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 2752–2759. [Google Scholar]
- Singh, B.; Reddy, K.R.; Redona, E.D.; Walker, T. Screening of rice cultivars for morpho-physiological responses to early-season soil moisture stress. Rice Sci. 2017, 24, 322–335. [Google Scholar] [CrossRef]
- Finch-Savage, W.E.; Clay, H.A.; Lynn, J.R.; Morris, K. Towards a genetic understanding of seed vigour in small-seeded crops using natural variation in (Brassica oleracea). Plant Sci. 2010, 179, 582–589. [Google Scholar] [CrossRef]
- Wopereis, M.C.S.; Kropff, M.J.; Maligaya, A.R.; Tuong, T.P. Drought-stress responses of two lowland rice cultivars to soil water status. Field Crops Res. 1996, 46, 21–39. [Google Scholar] [CrossRef]
- Shipley, B. Net assimilation rate, specific leaf area and leaf mass ratio: Which is most closely correlated with relative growth rate? A meta-analysis. Funct. Econ. 2006, 20, 565–574. [Google Scholar] [CrossRef]
- Zhang, Z.; Qu, X.; Wan, S.; Chen, L.; Shu, Y. Comparison of QTL controlling seedling vigour under different temperature conditions using recombinant inbred lines in rice (Oryza sativa). Ann. Bot. 2005, 95, 423–429. [Google Scholar] [CrossRef]
- Wijewardana, C.; Hock, M.; Henry, B.; Reddy, K.R. Screening corn hybrids for cold tolerance using morphological traits for early-season seeding. Crop Sci. 2015, 55, 851–867. [Google Scholar] [CrossRef]
- Serraj, R.; Atlin, G. Drought-resistant rice for increased rainfed production and poverty alleviation: A concept note. In Drought Frontiers in Rice: Crop Improvement for Increased Rainfed Production; Serraj, R., Bennett, J., Hardy, D., Eds.; IRRI: Los Banos, Philippines, 2008; pp. 385–400. [Google Scholar]
- Tuberosa, R.; Salvi, S.; Sanguineti, M.C.; Landi, P.; Maccaferri, M.; Conti, S. Mapping QTLs regulating morpho-physiological traits and yield: Case studies, shortcomings, and perspectives in drought-stressed maize. Ann. Bot. 2002, 89, 941–963. [Google Scholar] [CrossRef]
- Hewitt, E.J. Sand and Water Culture Methods Used in the Study of Plant Nutrition; Tech. Comm, No. 22; Commonwealth Bureau of Horticulture and Plantation Crops, Commonwealth Agriculture Bureau Farnham Royal: Buckinghamshire, UK, 1952.
- Bunnag, S.; Pongthai, P. Selection of rice (Oryza sativa L.) cultivars tolerant to drought stress at the vegetative stage under field conditions. Am. J. Plant Sci. 2013, 4, 1701–1708. [Google Scholar] [CrossRef]
- Brand, D.; Wijewardana, C.; Gao, W.; Reddy, K.R. Interactive effects of carbon dioxide, low temperature, and ultraviolet-B radiation on cotton seedling root and shoot morphology and growth. Front. Earth Sci. 2016, 10, 607–620. [Google Scholar] [CrossRef]
- Reddy, K.R.; Brand, D.; Wijewardana, C.; Gao, W. Temperature effects on cotton seedling emergence, growth, and development. Agron. J. 2017, 109, 1379–1387. [Google Scholar] [CrossRef]
- SAS Institute. SAS Guide to Macro Processing; SAS Institute: Cary, NC, USA, 2011; Volume 11. [Google Scholar]
- Raman, A.; Verulkar, S.B.; Mandal, N.P.; Variar, M.; Shukla, V.; Dwivedi, J.; Singh, B.N.; Singh, N.O.; Pamini, S.; Mall, K.A.; et al. Drought yield index to select high yielding rice lines under different drought stress severities. Rice 2012, 5, 31. [Google Scholar] [CrossRef] [Green Version]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress effects, mechanisms, and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef]
- Luciano, L.B.G.; Su, S.; Wu, C.; Hsieh, C. Establishment of a rapid screening method for drought tolerance of rice genotypes at seedling stage. J. Int. Coop. 2012, 7, 107–122. [Google Scholar]
- Rebolledo, M.C.; Dingkuhn, M.; Pere, P.; Mcnally, K.L.; Luquet, D. Developmental dynamics and early growth vigour in rice. I. relationship between development rate (1/Phyllochron) and growth. J. Agron. Crop Sci. 2012, 198, 374–384. [Google Scholar] [CrossRef]
- Yadav, R.K.; Kalia, P.; Choudhary, H.; Brihama, Z.H. Low-cost polyhouse technologies for higher income and nutritional security. Int. J. Agric. Food Sci. Technol. 2014, 5, 191–196. [Google Scholar]
- Redoña, E.D.; Golden, B.R.; Dickey, Z.D.; Smith, W.E.; Glenn, J.; Lanford, L.S. Mississippi Rice: Variety Trials (2016); Information Bulletin No. 517; Mississippi Agricultural and Forestry Experiment Station, Mississippi State University: Starkville, MS, USA, 2017; 18p. [Google Scholar]
- O’Neill, P.M.; Shanahan, J.F.; Schepers, J.S. Use of chlorophyll fluorescence assessments to differentiate corn hybrid response to variable water conditions. Crop Sci. 2006, 46, 681–687. [Google Scholar] [CrossRef]
- Kamoshita, A.; Rodriguez, R.; Yamauchi, A.; Wade, L. Genotypic variation in response of rainfed lowland rice to prolonged drought and rewatering. Plant Prod. Sci. 2004, 7, 406–420. [Google Scholar] [CrossRef]
- Serraj, R.; McNally, K.L.; Slamet-Loedin, I.; Kohli, A.; Haefele, S.M.; Atlin, G.; Kumar, A. Drought resistance improvement in rice: An integrated genetic and resource management strategy. Plant Prod. Sci. 2011, 14, 1–14. [Google Scholar] [CrossRef]
- Kamoshita, A.; Babu, R.C.; Boopathi, N.M.; Fukai, S. Phenotypic and genotypic analysis of drought-resistance traits for development of rice cultivars adapted to rainfed environments. Field Crops Res. 2008, 109, 1–23. [Google Scholar] [CrossRef]
- Chutia, J.; Borah, S.P. Water stress effects on leaf growth and chlorophyll content but not the grain yield in traditional rice (Oryza sativa Linn.) genotypes of Assam, India II. Protein and proline status in seedlings under PEG induced water stress. Am. J. Plant Sci. 2012, 3, 971–980. [Google Scholar] [CrossRef]
- Zain, N.A.M.; Ismail, M.R.; Mahmood, M.; Puteh, A.; Ibrahim, M.H. Alleviation of water stress effects on mr220 rice by application of periodical water stress and potassium fertilization. Molecules 2014, 19, 1795–1819. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.T. Plant Water Relation Studies in Diverse Rice Cultivars under Bangladesh Climatic Conditions. Ph.D. Thesis, Institute of Agronomy, University of Agricultural Sciences, Vienna, Austria, 1999. [Google Scholar]
- Bota, J.; Medrano, H.; Flexas, J. Is photosynthesis limited by decreased Rubisco activity and RuBP content under progressive water stress? New Phytol. 2004, 162, 671–681. [Google Scholar] [CrossRef] [Green Version]
- Loreto, F.; Tricoli, D.; Di, M.G. On the relationship between electron transport rate and photosynthesis in leaves of the C4 plant (Sorghum bicolor) exposed to water stress, temperature changes and carbon metabolism inhibition. Aust. J. Plant Physiol. 1995, 22, 885–892. [Google Scholar] [CrossRef]
- Borrell, A.K.; Hammer, G.L.; Douglas, A.C.L. Does maintaining green leaf area in sorghum improve yield under drought? I. Leaf growth and senescence. Crop Sci. 2000, 40, 1026–1037. [Google Scholar] [CrossRef]
- Murty, P.S.S.; Murty, K.S. Spikelet sterility in relation to nitrogen and carbohydrate contents in rice. Ind. J. Plant Physiol. 1982, 25, 40–48. [Google Scholar]
- Farooq, M.; Kobayashi, N.; Ito, O.; Wahid, A.; Serraj, R. Broader leaves result in better performance of indica rice under drought stress. J. Plant Physiol. 2010, 167, 1066–1075. [Google Scholar] [CrossRef] [PubMed]
- Mostajeran, A.; Rahimi-Eichi, V. Effects of drought stress on growth and yield of rice (Oryza sativa L.) cultivars and accumulation of proline and soluble sugars in sheath and blades of their different ages leaves. Am. Eurasian J. Agric. Environ. Sci. 2009, 5, 264–272. [Google Scholar]
- Xu, W.; Cui, K.; Xu, A.; Nie, L.; Huang, J.; Peng, S. Drought stress condition increases root to shoot ratio via alteration of carbohydrate partitioning and enzymatic activity in rice seedlings. Acta Physiol. Plant. 2015, 37, 9. [Google Scholar] [CrossRef]
- Paez-Garcia, A.; Motes, C.; Scheible, W.R.; Chen, R.; Blancaflor, E.; Monteros, M. Root traits and phenotyping strategies for plant improvement. Plants 2015, 4, 334–355. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.T.; Kano-Nakata, M.; Suralta, R.R.; Menge, D.; Mitsuya, S.; Inukai, Y.; Yamauchi, A. Root plasticity and its functional roles were triggered by water deficit but not by the resulting changes in the forms of soil N in rice. Plant Soil 2014, 386, 65–76. [Google Scholar] [CrossRef]
- Kano-Nakata, M.; Inukai, Y.; Wade, L.; Siopongco, J.D.; Yamauchi, A. Root development, water uptake, and shoot dry matter production under water deficit conditions in two CSSLs of rice: Functional roles of root plasticity. Plant Prod. Sci. 2011, 14, 307–317. [Google Scholar] [CrossRef]
- Abe, J.; Morita, S. Growth direction of nodal roots in rice: Its variation and contribution to root system formation. Plant Soil 1994, 165, 333–337. [Google Scholar] [CrossRef]
- Yoshida, S.; Hasegawa, S. The rice root system: Its development and function. In Drought Resistance in Crops with Emphasis on Rice; International Rice Research Institute: Manila, Philippines, 1982; pp. 97–114. [Google Scholar]
- Parker, J.S.; Alison, C.C.; Liam, D.; Keith, R.; Claire, S.G. Genetic interactions during root hair morphogenesis in Arabidopsis. Plant Cell Online 2000, 12, 1961–1974. [Google Scholar] [CrossRef]
- Samson, B.; Wade, L. Soil physical constraints affecting root growth, water extraction, and nutrient uptake in rainfed lowland rice. In Rainfed Lowland Rice: Advances in Nutrient Management Research; Ladha, J.K., Wade, L.J., Dobermann, A., Reinhardt, W., Kirk, W., Piggin, C., Eds.; International Rice Research Institute: Manila, Philippines, 1998; pp. 231–244. [Google Scholar]
- Clark, L.J.; Price, A.H.; Steele, K.A.; Whalley, W.R. Evidence from near-isogenic lines that root penetration increases with root diameter and bending stiffness in rice. Fun. Plant Biol. 2008, 35, 1163–1171. [Google Scholar] [CrossRef]
- Fu, J.; Huang, B. Involvement of antioxidants and lipid peroxidation in the adaptation of two cool-season grasses to localized drought stress. Environ. Exp. Bot. 2001, 45, 105–114. [Google Scholar] [CrossRef]
- Hura, T.; Grzesiak, S.; Hura, K.; Thiemt, E.; Tokarz, K.; Wȩdzony, M. Physiological and biochemical tools useful in drought-tolerance detection in genotypes of winter triticale: Accumulation of ferulic acid correlates with drought tolerance. Ann. Bot. 2007, 100, 767–775. [Google Scholar] [CrossRef] [PubMed]
- Long, S.P.; Humphries, S.; Falkowski, P.G. Photoinhibition of photosynthesis in nature. Ann. Rev. Plant Physio. Biol. 1994, 45, 633–662. [Google Scholar] [CrossRef]
- Ambavaram, M.M.R.; Basu, S.; Krishnan, A.; Ramegowda, V.; Batlang, U.L.; Rahman, A.P. Coordinated regulation of photosynthesis in rice increases yield and tolerance to environmental stress. Nat. Commun. 2014, 5, 5302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maxwell, K.; Johanson, G.N. Chlorophyll fluorescence-a practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Colom, M.R.; Vazzana, C. Photosynthesis and PSII functionality of drought-resistant and drought-sensitive weeping lovegrass plants. Environ. Exp. Bot. 2003, 49, 135–144. [Google Scholar] [CrossRef]
- IRRI. Standard Evaluation System for Rice, 5th ed.; International Rice Research Institute: Los Baños, the Philippines, 2014. [Google Scholar]
- Krishnan, P.; Ramakrishnan, B.; Reddy, K.R.; Reddy, V.R. High temperature effects on rice growth, yield, and grain quality. Adv. Agron. 2011, 111, 87–206. [Google Scholar]
- Massey, J.H.; Walker, T.W.; Anders, M.M.; Smith, M.C.; Avila, L.A. Farmer adaptation of intermittent flooding using multiple inlet rice irrigation in Mississippi. Agric. Water Manag. 2014, 146, 297–304. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | PH | TN | LN | LA | RCL | ARD | RV | NRT | NRF | NRC | LRL | RN | LDW | SDW | RDW | SHDW | RSR | TDW | SPAD | Fo | Fm | Fv | Fv/Fm |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Genotypes | *** | *** | NS | *** | *** | ** | *** | NS | *** | *** | * | *** | *** | *** | *** | *** | ** | *** | *** | ** | *** | *** | * |
| Drought | *** | * | NS | *** | *** | NS | ** | NS | *** | ** | NS | * | * | *** | *** | *** | ** | *** | *** | NS | * | *** | NS |
| Control | *** | *** | NS | *** | * | * | *** | * | *** | *** | *** | * | *** | NS | *** | ** | NS | *** | NS | *** | *** | *** | * |
| Control × Drought | NS | * | NS | *** | * | NS | * | NS | *** | *** | NS | NS | ** | NS | * | * | * | * | * | *** | *** | NS |
| Highly Sensitive | Sensitive | Moderate | Tolerant | Highly Tolerant |
|---|---|---|---|---|
| (14.701–16.989) | (16.990–19.278) | (19.279–21.567) | (21.568–23.855) | (>23.856) |
| CHENIERE (14.701), CL Jazzman (15.397), LA 2008 (16.012), RU1504157 (16.662), RU1403126 (16.731) | JUPITER(17.001), RU0603075 (17.178), RU1504114 (17.181), 14CVPYT144 (17.251), RU1401164 (17.273), RU1402065 (17.573), RU1303138 (17.679), RU1404191 (17.690), RU1504194 (17.730), 14CLPYT033 (17.836), RoyJ (17.854), NIPONBARE (17.980), CL271 (18.054), RU1403107 (18.103), RU1504083 (18.106), RU1404154 (18.157), RU1401090 (18.210), RU1504198 (18.228), RU1402115 (18.390), RU1301102 (18.427), RU1402031 (18.771), ANTONIO (18.865), RU1304114 (19.046), Bowman (19.062), IRGA409 (19.079), 14CVPYT094 (19.100), RU1504100 (19.195), CL151 (19.269) | RU1305001 (19.293), CL152 (19.314), LA 2134 (19.319) RU1302192 (19.425), RU1304122 (19.431), RU1504122 (19.434), RU1504154 (19.549), CL172 (19.573), RU1504197 (19.671), CL111 (19.742), RU1504191 (19.748), RU1401070 (19.829), RU1402005 (19.884), CAFFEY (19.909), RU1402131 (19.930), El Paso 144 (20.026), RU1504196 (20.181), Thad (20.205), CL142-AR(20.211), RU1401099 (20.293), RU1201136 (20.310), RU1304156 (20.319), RU1401102 (20.347), RU1404156 (20.365), RU1402134 (20.399), RU1301084 (20.427), RU1404194 (20.495), RU1304154 (20.685), GSOR100417 (20.714), Rex (20.791), RU1201047 (20.863), MERMENTAU (20.868), Taggart (21.050), COLORADO (21.147), RU1303181 (21.153), RU1104122 (21.216), Sabine (21.266), RU1204156 (21.314), RU1404193 (21.395), RU1402189 (21.422), 14CLPYT108 (21.427), CLJZMN (21.457), RU1404122 (21.518), RU1201024 (21.526), RU1401161 (21.557) | RU1504193 (21.831), RU1401067 (21.884), RU1504156 (21.910), RU1404157 (21.978), RU1404198 (21.990), JES (22.160), GSOR100390 (22.307), RU1204197 (22.381), RU1301093 (22.639), RU1504186 (22.864), Cocodrie (22.998), RU1402149 (23.074), LAKAST (23.076), Presidio (23.225), GSOR101758 (23.472), RU1404196 (23.763) | INIA Tacuari (23.989), RU1402195 (24.527), CL163 (24.709), RU1401145 (25.376), N-22 (25.638), RU1402174 (27.955) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lone, A.A.; Jumaa, S.H.; Wijewardana, C.; Taduri, S.; Redona, E.D.; Reddy, K.R. Drought Stress Tolerance Screening of Elite American Breeding Rice Genotypes Using Low-Cost Pre-Fabricated Mini-Hoop Modules. Agronomy 2019, 9, 199. https://doi.org/10.3390/agronomy9040199
Lone AA, Jumaa SH, Wijewardana C, Taduri S, Redona ED, Reddy KR. Drought Stress Tolerance Screening of Elite American Breeding Rice Genotypes Using Low-Cost Pre-Fabricated Mini-Hoop Modules. Agronomy. 2019; 9(4):199. https://doi.org/10.3390/agronomy9040199
Chicago/Turabian StyleLone, Ajaz A., Salah H. Jumaa, Chathurika Wijewardana, Shasthree Taduri, Edilberto D. Redona, and K. Raja Reddy. 2019. "Drought Stress Tolerance Screening of Elite American Breeding Rice Genotypes Using Low-Cost Pre-Fabricated Mini-Hoop Modules" Agronomy 9, no. 4: 199. https://doi.org/10.3390/agronomy9040199
APA StyleLone, A. A., Jumaa, S. H., Wijewardana, C., Taduri, S., Redona, E. D., & Reddy, K. R. (2019). Drought Stress Tolerance Screening of Elite American Breeding Rice Genotypes Using Low-Cost Pre-Fabricated Mini-Hoop Modules. Agronomy, 9(4), 199. https://doi.org/10.3390/agronomy9040199