Association of Physiological Responses and Root Distribution Patterns of Ratooning Ability and Yield of the Second Ratoon Cane in Sugarcane Elite Clones



Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Plant Materials
2.2. Data Collection
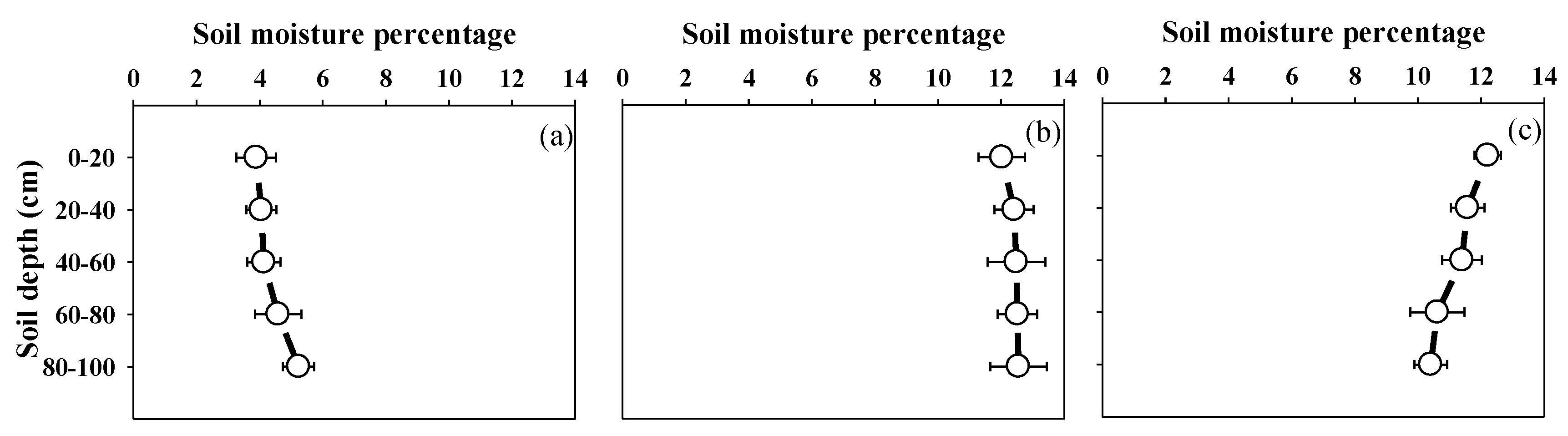
2.2.1. Meteorological and Soil Data
2.2.2. Germination Percentage
2.2.3. Physiological Traits
2.2.4. Root Traits
2.2.5. Yield
2.3. Statistical Analysis
3. Results
3.1. Germination and Yield
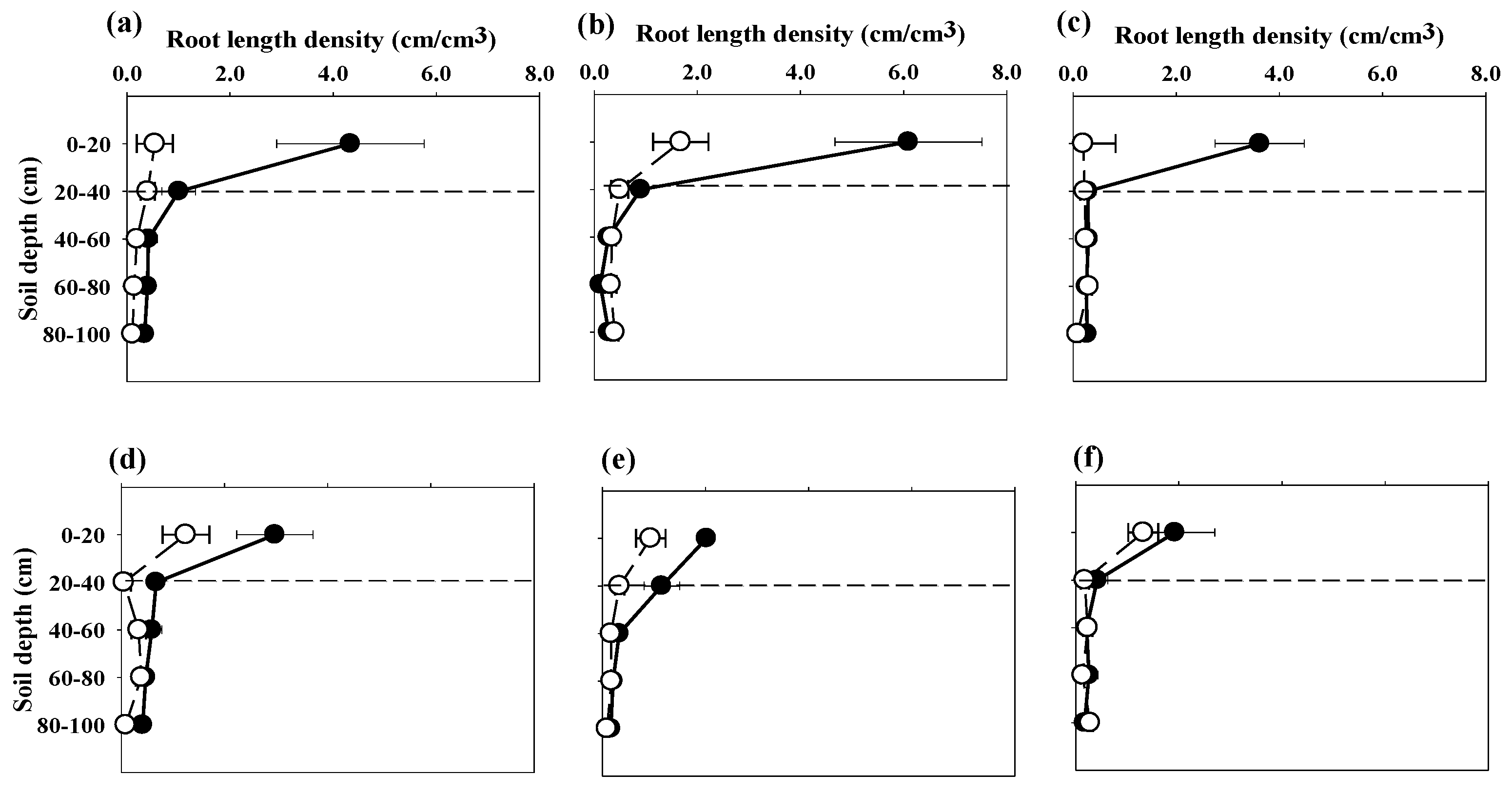
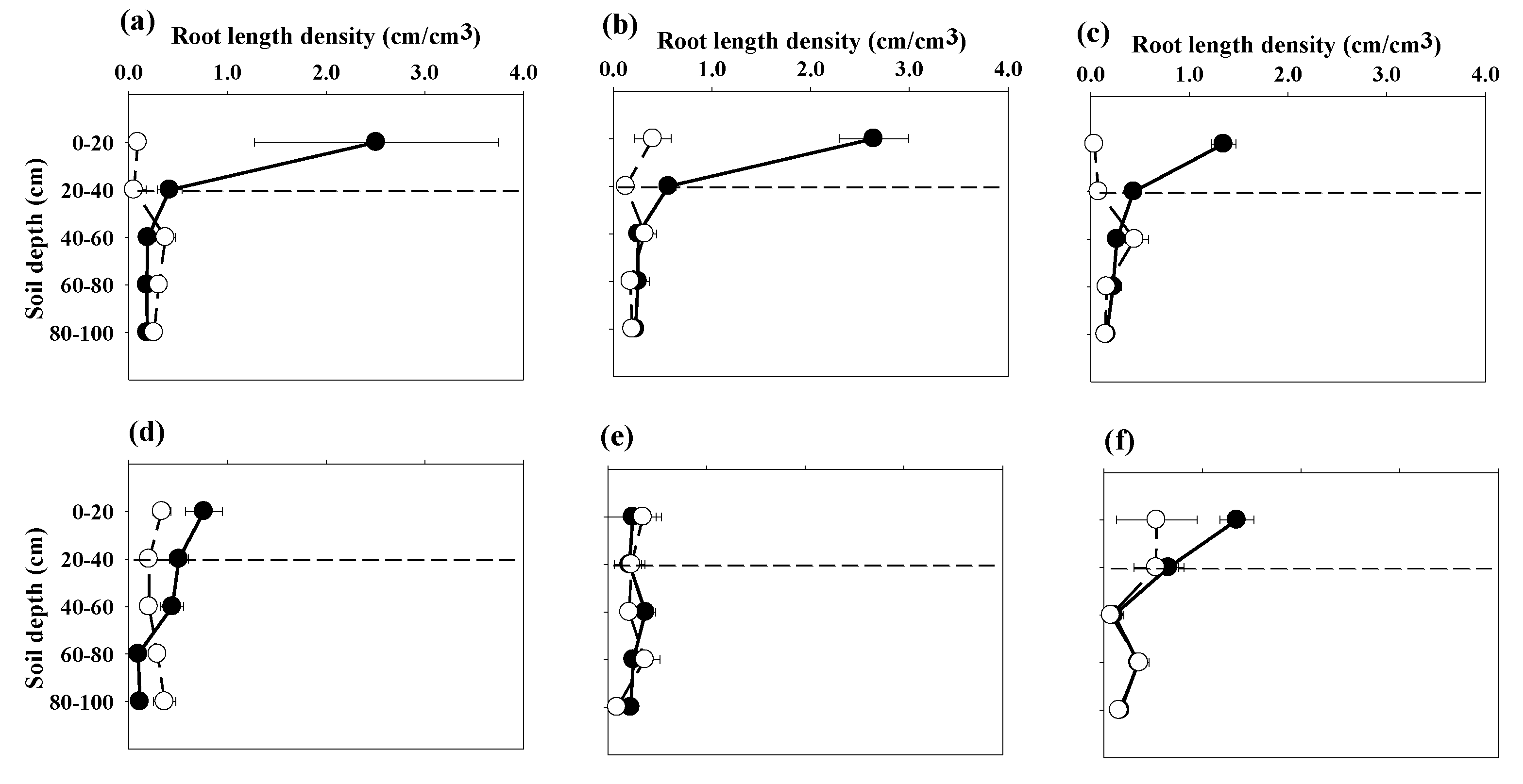
3.2. Root Length Density
3.3. Root Distribution Patterns
3.4. Physiological Traits Related to Drought Resistance
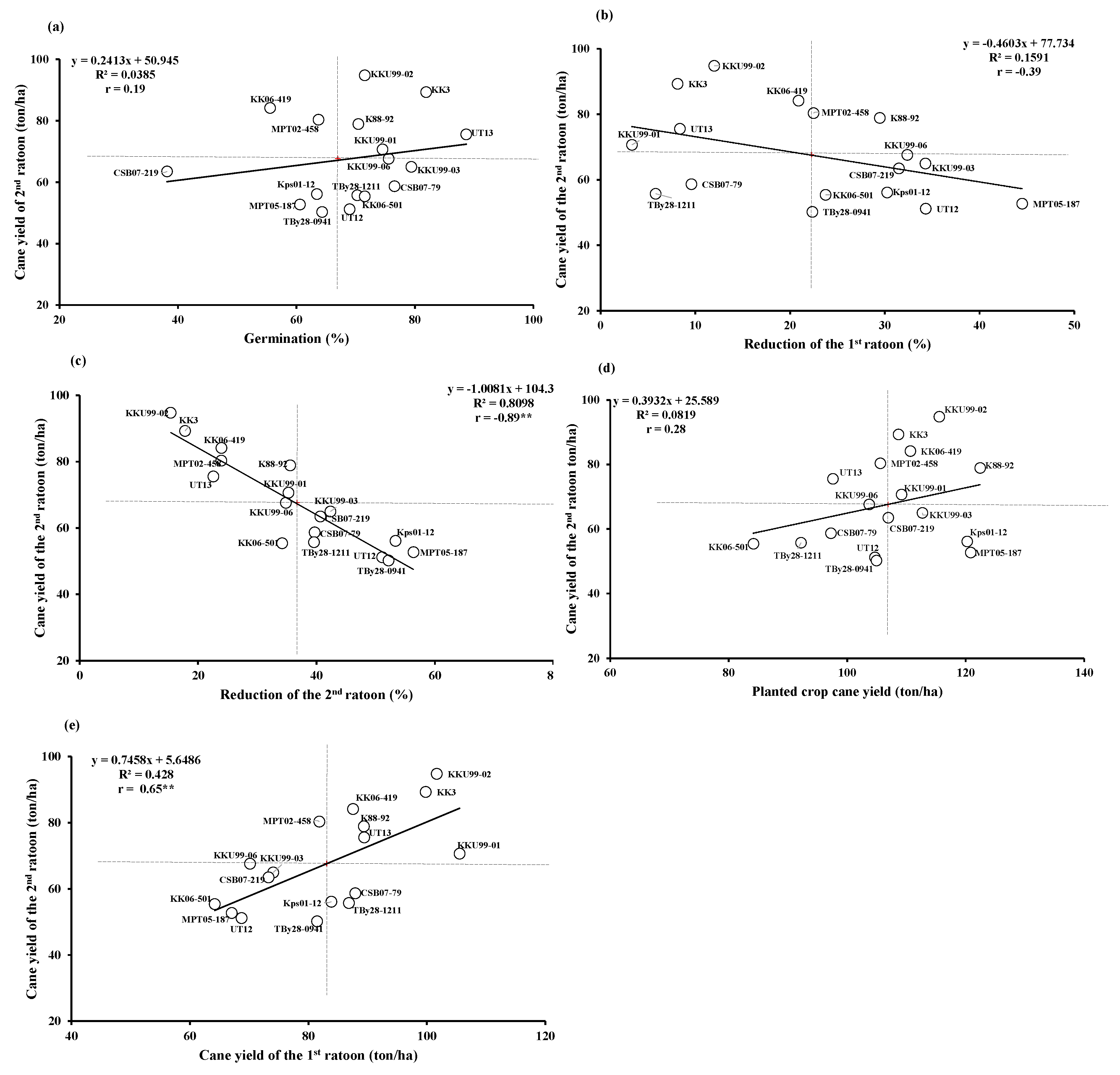
3.5. Correlations Between Traits
4. Discussion
4.1. Germination and Yield
4.2. Root Length Density
4.3. Root Distribution Patterns
4.4. Physiological Traits Related to Drought Resistance
4.5. Correlations among Traits
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| 1R | First ratoon crop |
| 2R | Second ratoon crop |
| DAH | Days after harvest |
| LSD | Least significant difference |
| PC | Plant crop |
| RLD | Root length density |
| RWC | Relative water content |
| SCMR | SPAD chlorophyll meter reading |
References
- Robertson, M.J.; Inman-Bamber, N.G.; Muchow, R.C.; Wood, A.W. Physiology and productivity of sugarcane with early and mid-season water deficit. Field Crop Res. 1999, 64, 211–227. [Google Scholar] [CrossRef]
- Bashir, S.; Hassan, M.; Fiaz, N.; Khan, Z.; Ali, Z. Ratooning potential of different promising sugarcane genotypes at varying harvesting dates. J. Agric. Biol. Sci. 2013, 8, 437–440. [Google Scholar]
- Bashir, S.; Fiaz, N.; Ghaffar, A.; Khalid, F. Ratooning ability of sugarcane genotypes at different harvesting dates. Int. Sugar J. 2012, 114, 273–276. [Google Scholar]
- Smith, D.M.; Inman-Bamber, N.G.; Thorburn, P.J. Growth and function of the sugarcane root system. Field Crop Res. 2005, 92, 169–183. [Google Scholar] [CrossRef]
- Laclau, P.B.; Laclau, J.P. Growth of the whole root system for a plant crop of sugarcane under rainfed and irrigated environments in Brazil. Field Crop Res. 2009, 114, 351–360. [Google Scholar] [CrossRef]
- Gregory, P.J. Plant Roots: Growth, Activity and Interaction with Soil; Blackwell Publishing: Oxford, UK, 2006; pp. 1–44. [Google Scholar]
- Silva, M.D.A.; Jifon, J.L.; Da Silva, J.A.; Sharma, V. Use of physiological parameters as fast tools to screen for drought tolerance in sugarcane. Braz. J. Plant Physiol. 2007, 19, 193–201. [Google Scholar] [CrossRef]
- Silva, M.A.; Jifon, J.L.; Sharma, V.; Silva, J.A.G.; Caputo, M.M.; Damaj, M.B.; Guimara, E.R. Use of physiological parameters in screening drought tolerance in sugarcane genotypes. Sugar Tech 2011, 13, 191–197. [Google Scholar] [CrossRef]
- Santos, C.M.; Almeida Silva, M.; Lima, G.P.; Almeida Prado Bortolheiro, F.P.; Brunelli, M.C.; Holanda, L.A.; Oliver, R. Physiological changes associated with antioxidant enzymes in response to sugarcane tolerance to water deficit and rehydration. Sugar Tech 2015, 17, 291–304. [Google Scholar] [CrossRef]
- Welbank, P.J.; Gibb, M.J.; Taylor, P.J.; Williams, E.D. Root growth of cereal crops. In Rothamsted Experimental Station Annual Report 1973, Part 2. Rothamsted Experimental Station; Lawes Agricultural Trust: Harpenden, UK, 1974; pp. 26–66. [Google Scholar]
- Ford, K.E.; Gregory, P.J.; Gooding, M.J.; Pepler, S. Genotype and fungicide effects on late-season root growth of winter wheat. Plant Soil 2006, 284, 33–44. [Google Scholar] [CrossRef]
- Watanabe, K.; Saensupo, S.; Na-iam, Y.; Klomsa-ard, P.; Sriroth, K. Effects of superabsorbent polymer on soil water content and sugarcane germination and early growth in sandy soil conditions. Sugar Tech. 2018. [CrossRef]
- Gomathi, R.; Rao, P.N.G.; Rakkiyappan, P.; Sundara, B.P.; Shiyamala, S. Physiological studies on ratoonability of sugarcane varieties under tropical Indian condition. Am. J. Plant Sci. 2013, 4, 274. [Google Scholar] [CrossRef]
- Zhou, M.M.; Shoko, M.D. Simultaneous selection for yield and ratooning ability in sugarcane genotypes using analysis of covariance. S. Afr. J. Plant. Soil. 2012, 29, 93–100. [Google Scholar] [CrossRef]
- Bull, T.A.; Glasziou, K.T. Sugarcane. In Crop Physiology; Evans, L.T., Ed.; Cambridge University Press: Cambridge, UK, 1975; pp. 51–72. [Google Scholar]
- Abayomi, Y.A.; Etejere, E.O.; Fadayomi, O. Effect of stalk section, coverage depth and date of first irrigation on seedcane germination of two commercial sugarcane cultivars in Nigeria. Turrialba 1990, 40, 58–62. [Google Scholar]
- Mauri, R.; Coelho, R.D.; Junior, F.; Eusímio, F.; Barbosa, F.D.S.; Leal, D.P. Water relations at the initial sugarcane growth phase under variable water deficit. Eng. Agríc. 2017, 37, 268–276. [Google Scholar] [CrossRef]
- Mu, Z.; Guo, L.; Ru-Kai, C.H.E.N. Photosynthesis characteristics in eleven cultivars of sugarcane and their responses to water stress during the elongation stage. Proc. Int. Soc. Sugar Cane Technol. 2001, 24, 642–643. [Google Scholar]
- Gascho, G.J.; Shih, S.F. Sugarcane. In Crop-Water Relations; Teare, I.D., Peet, M.M., Eds.; John Wiley and Sons: New York, NY, USA, 1983; pp. 445–479. [Google Scholar]
- Kang, M.S.; Sosa, O.; Miller, J.D. Path analyses for percent fiber and cane and sugar yield in sugarcane. Crop Sci. 1989, 29, 1481–1483. [Google Scholar] [CrossRef]
- Milligan, S.B.; Gravois, K.A.; Bischoff, K.P.; Martin, F.A. Crop effects on genetic relationships among sugarcane traits. Crop Sci. 1990, 30, 927–931. [Google Scholar] [CrossRef]
- Ilyas, M.K.; Khan, F.A. Determining sucrose recovery in Saccarrum officinarum L. using regression and correlation analyses. Afr. J. Biotechnol. 2010, 9, 2985–2988. [Google Scholar]
- Samson, B.K.; Sinclair, T.R. Soil core and minirhizotron comparison for the determination of root length density. Plant Soil 1994, 161, 225–232. [Google Scholar] [CrossRef]
- Lynch, J. Root architecture and plant productivity1. Plant Physiol. 1995, 109, 7–13. [Google Scholar] [CrossRef]
- Jongrungklanga, N.; Toomsana, B.; Vorasoota, N.; Jogloya, S.; Booteb, K.J.; Hoogenboomc, G.; Patanothaia, A. Drought tolerance mechanisms for yield responses to pre-flowering drought stress of peanut genotypes with different drought tolerant levels. Field Crops Res. 2013, 144, 34–42. [Google Scholar] [CrossRef]
- Araki, H.; Morita, S.; Tatsumi, J.; Iijima, M. Physiol-morphological analysis on axile root growth in upland rice. Plant Prod. Sci. 2002, 5, 286–293. [Google Scholar] [CrossRef]
- Uga, Y.; Sugimoto, K.; Ogawa, S.; Rane, J.; Ishitani, M.; Hara, N.; Kitomi, Y.; Inukai, Y.; Ono, K.; Kanno, N.; et al. Control of root system architecture by deeper rooting 1 increases rice yield under drought conditions. Nat. Genet. 2013, 45, 1097. [Google Scholar] [CrossRef]
- An, H.; Dong, H.; Wu, T.; Wang, Y.; Xu, X.; Zhang, X.; Han, Z. Root growth angle: An important trait that influences the deep rooting of apple rootstocks. Sci. Hort. 2017, 216, 256–263. [Google Scholar] [CrossRef]
- Kato, Y.; Okami, M. Root morphology, hydraulic conductivity and plant water relations of high-yielding rice grown under aerobic conditions. Ann. Bot. 2011, 108, 575–583. [Google Scholar] [CrossRef] [Green Version]
- Moreira, D.R.; Cardoso, V.J.M. Effect of soil moisture content and the irrigation frequency on the sugarcane germination. Pesqui. Agropecu. Bras. 1998, 721–729. [Google Scholar]
- Jangpromma, N.; Thammasirirak, S.; Jaisil, P.; Songsri, P. Effects of drought and recovery from drought stress on above ground and root growth, and water use efficiency in sugarcane (Saccharum officinarum L.). Aust. J. Crop Sci. 2012, 6, 1298–1304. [Google Scholar]
- Jangpromma, N.; Songsri, P.; Thammasirirak, S.; Jaisil, P. Rapid assessment of chlorophyll content in sugarcane using a SPAD chlorophyll meter across different water stress conditions. Asian J. Plant Sci. 2010, 9, 368–374. [Google Scholar] [CrossRef]
- Richardson, A.D.; Duigan, S.P.; Berlyn, G.P. An evaluation of noninvasive methods to estimate foliar chlorophyll content. New Phytol. 2002, 153, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Akkasaeng, C.; Vorasoot, N.; Jogloy, S.; Patanotai, A. Relationship between SPAD readings and chlorophyll contents in leaves of peanut (Arachis hypogaea L.). Thai J. Agric. Sci. 2003, 36, 279–284. [Google Scholar]
- Arunyanark, A.; Jogloy, S.; Akkasaeng, C.; Vorasoot, N.; Kesmala, T.; Nageswara Rao, R.C.; Patanothai, A. Chlorophyll stability is an indicator of drought tolerance in peanut. J. Agron. Crop Sci. 2008, 194, 113–125. [Google Scholar] [CrossRef]
- Silva, M.D.A.; Silva, J.A.G.D.; Enciso, J.; Sharma, V.; Jifon, J. Yield components as indicators of drought tolerance of sugarcane. Sci. Agric. 2008, 65, 620–627. [Google Scholar] [CrossRef]
- Steudle, E. Water uptake by roots: Effects of water deficit. J. Exp. Bot. 2000, 51, 1531–1542. [Google Scholar] [CrossRef]
- Liberato, M.A.R.; Gonçalves, J.F.D.C.; Chevreuil, L.R.; Junior, N.; da Rocha, A.; Fernandes, A.V.; Santos Junior, U.M.D. Leaf water potential, gas exchange and chlorophyll a fluorescence in acariquara seedlings (Minquartia guianensis Aubl.) under water stress and recovery. Braz. J. Plant Physiol. 2006, 18, 315–323. [Google Scholar] [CrossRef]
- Lobato, A.K.S.; Oliveria Neto, C.F.; Costa, R.C.L.; Santos Filho, B.G.; Cruz, F.J.R.; Laughinghouse, I.V. Biochemical and physiological behavior of Vigna unguiculata (L.) Walp. under water stress during the vegetative stage. Asian J. Plant Sci. 2008, 7, 44–49. [Google Scholar]
- Zhao, D.; Glaz, B.; Comstock, J.C. Sugarcane response to water-deficit stress during early growth on organic and sandy soils. Am. J. Agric. Biol. Sci. 2010, 5, 403–414. [Google Scholar] [CrossRef]
- Davies, W.J.; Wilkinson, S.; Loveys, B. Stomatal control by chemical signaling and the exploitation of this mechanism to increase water use efficiency in agriculture. New Phytol. 2002, 153, 449–460. [Google Scholar] [CrossRef]
- Azevedo Neto, A.D.D.; Prisco, J.T.; Enéas-Filho, J.; Lacerda, C.F.D.; Silva, J.V.; Costa, P.H.A.D.; Gomes-Filho, E. Effects of salt stress on plant growth, stomatal response and solute accumulation of different maize genotypes. Braz. J. Plant Physiol. 2004, 16, 31–38. [Google Scholar] [CrossRef]
- Smit, M.A.; Singels, A. The response of sugarcane canopy development to water stress. Field Crops Res. 2006, 98, 91–97. [Google Scholar] [CrossRef]
- Turner, N.C. Adaptation to water deficits: A changing perspective. Funct. Plant Biol. 1986, 13, 175–190. [Google Scholar] [CrossRef]
- Smith, J.P.; Lawn, R.J.; Nable, R.O. Investigations into the root: Shoot relationship of sugarcane, and some implications for crop productivity in the presence of sub-optimal soil conditions. Proc. Aust. Soc. Sugar Cane Technol. 1999, 21, 108–113. [Google Scholar]
- Boutraa, T.; Akhkha, A.; Al-Shoaibi, A.A.; Alhejeli, A.M. Effect of water stress on growth and water use efficiency (WUE) of some wheat cultivars (Triticum durum) grown in Saudi Arabia. J. Taibah Univ. Sci. 2010, 3, 39–48. [Google Scholar] [CrossRef]
- Graça, J.P.D.; Rodrigues, F.A.; Farias, J.R.B.; Oliveira, M.C.N.D.; Hoffmann-Campo, C.B.; Zingaretti, S.M. Physiological parameters in sugarcane cultivars submitted to water deficit. Braz. J. Plant Physiol. 2010, 22, 189–197. [Google Scholar] [CrossRef] [Green Version]
- Songsri, P.; Jogloy, S.; Kesmala, T.; Vorasoot, N.; Akkasaeng, C.; Patanothai, A.; Holbrook, C.C. Response of reproductive characters of drought resistant peanut genotypes to drought. Asian J. Plant Sci. 2008, 7, 427–439. [Google Scholar]
- Derner, J.D.; Polley, H.W.; Johnson, H.B.; Tischler, C.R. Root system response of C4 grass seedlings to CO2 and soil water. Plant Soil 2001, 231, 97–104. [Google Scholar] [CrossRef]
- Khonghintaisong, J.; Songsri, P.; Toomsan, B.; Jongrungklang, N. Rooting and physiological trait responses to early drought stress of sugarcane cultivars. Sugar Tech 2018, 20, 396–406. [Google Scholar] [CrossRef]
- Glover, J. The behavior of the root-system of sugarcane at and after harvest. Proc. S. Afr. Sugar Technol. Assoc. 1968, 42, 133–135. [Google Scholar]
- Azevedo, M.C.B.D.; Chopart, J.L.; Medina, C.D.C. Sugarcane root length density and distribution from root intersection counting on a trench-profile. Sci. Agric. 2011, 68, 94–101. [Google Scholar] [CrossRef] [Green Version]
- Atta, B.M.; Mahmood, T.; Trethowan, T.M. Relationship between root morphology and grain yield of wheat in north-western NSW, Australia. Aust. J. Crop Sci. 2013, 7, 2108. [Google Scholar]
- Reynolds, M.; Dreccer, F.; Trethowan, R. Drought-adaptive traits derived from wheat wild relatives and landraces. J. Exp. Bot. 2006, 58, 177–186. [Google Scholar] [CrossRef] [Green Version]
- Basnayake, J.; Jackson, P.A.; Inman-Bamber, N.G.; Lakshmanan, P. Sugarcane for water-limited environments. Sugarcane for water-limited environments: Genetic variation in cane yield and sugar content in response to water stress. J. Exp. Bot. 2012, 63, 6023–6033. [Google Scholar] [CrossRef]
- Krause, G.H.; Weis, E. Chlorophyll fluorescence and photosynthesis: The basics. Annu. Rev. Plant Biol. 1991, 42, 313–349. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Source |
|---|---|
| 1.) TBy28-1211 | Kasetsart University, Kamphaeng Saen Campus |
| 2.) TBy28-0941 | Kasetsart University, Kamphaeng Saen Campus |
| 3.) MPT02-458 | Mitr Phol Innovation & Research Center |
| 4.) MPT05-187 | Mitr Phol Innovation & Research Center |
| 5.) UT12 | Suphan Buri Field Crops Research Center |
| 6.) UT13 | Suphan Buri Field Crops Research Center |
| 7.) CSB07-79 | Office of the Cane and Sugar Board |
| 8.) CSB07-219 | Office of the Cane and Sugar Board |
| 9.) KK06-419 | Khon Kaen Field Crops Research Center |
| 10.) KK06-501 | Khon Kaen Field Crops Research Center |
| 11.) KKU99-01 | Khon Kaen University |
| 12.) KKU99-02 | Khon Kaen University |
| 13.) KKU99-03 | Khon Kaen University |
| 14.) KKU99-06 | Khon Kaen University |
| 15.) Kps01-12 | Kasetsart University |
| 16.) KK3 | Khon Kaen Field Crops Research Center |
| 17.) K88-92 | Office of the Cane and Sugar Board |
| Source | Cane Yield | Germination Percentage | ||
|---|---|---|---|---|
| PC | 1R | 2R | ||
| Replication | 9.81 | 27.44 | 0.88 | 351.07 |
| Genotypes | 293.42 ** | 579.35 ** | 593.02 ** | 718.86 ** |
| Error | 16.74 | 22.83 | 68.43 | 191.79 |
| CV. (%) | 3.83 | 5.27 | 12.23 | 21.27 |
| Overall mean | 106.98 | 90.72 | 67.63 | 65.12 |
| Source | RLD (cm/cm3) between Plants/at 90 DAH | RLD (cm/cm3) between Rows/at 90 DAH | RLD (cm/cm3) between Plants/at 270 DAH | RLD (cm/cm3) between Rows/at 270 DAH | ||||
|---|---|---|---|---|---|---|---|---|
| Upper (0–40 cm) | Lower (40–100 cm) | Upper (0–40 cm) | Lower (40–100 cm) | Upper (0–40 cm) | Lower (40–100 cm) | Upper (0–40 cm) | Lower (40–100 cm) | |
| Replication | 0.00 | 0.00 | 0.00 | 0.00 | 0.03 | 0.00 | 0.01 | 0.00 |
| Genotypes | 0.70 ** | 0.02 ** | 0.06 ** | 0.01 ** | 1.42 ** | 0.03 ** | 0.30 ** | 0.02 ** |
| Error | 0.02 | 0.00 | 0.00 | 0.00 | 0.02 | 0.00 | 0.00 | 0.00 |
| CV. (%) | 12.55 | 10.10 | 12.67 | 14.80 | 7.15 | 13.97 | 14.50 | 18.19 |
| Overall mean | 1.16 | 0.23 | 0.18 | 0.20 | 2.29 | 0.28 | 0.46 | 0.21 |
| Source | SCMR | Chlorophyll Fluorescence (Fv/Fm) | RWC (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 90 DAH | 180 DAH | 270 DAH | 90 DAH | 180 DAH | 270 DAH | 90 DAH | 180 DAH | 270 DAH | |
| Replication | 15.800 | 2.340 | 7.520 | 0.001 | 0.001 | 0.004 | 1.460 | 3.750 | 52.310 |
| Genotypes | 6.420 * | 8.710 ** | 8.420 ** | 0.001 ** | 0.000 ** | 0.000ns | 10.120ns | 2.690ns | 56.980ns |
| Error | 2.880 | 2.710 | 2.600 | 0.000 | 0.000 | 0.000 | 5.230 | 2.210 | 49.520 |
| CV. (%) | 4.540 | 4.020 | 3.940 | 2.400 | 0.900 | 1.420 | 2.380 | 1.510 | 7.280 |
| Overall mean | 37.350 | 41.060 | 40.950 | 0.789 | 0.804 | 0.808 | 96.200 | 98.250 | 96.610 |
| Source | Specific Leaf Area (cm2/g) | Stomatal Conductance (µm2/s) | ||
|---|---|---|---|---|
| 180 DAH | 270 DAH | 180 DAH | 270 DAH | |
| Replication | 67.90 | 193.40 | 1235.43 | 12445.30 |
| Genotypes | 708.35 ** | 180.78 ** | 4599.38 ** | 3934.80 * |
| Error | 70.08 | 28.06 | 689.23 | 1692.70 |
| CV. (%) | 10.09 | 7.97 | 8.63 | 11.76 |
| Overall mean | 82.99 | 66.46 | 304.32 | 349.86 |
| Cane Yield | Germination Percentage | |
|---|---|---|
| Root length density | ||
| Upper soil layer (0–40 cm) | ||
| RLD between plants at 90 DAH | −0.12 | 0.36 ** |
| RLD between rows at 90 DAH | 0.22 | 0.14 |
| RLD between plants at 270 DAH | 0.17 | −0.14 |
| RLD between rows at 270 DAH | −0.09 | −0.01 |
| Lower soil layer (40–100 cm) | ||
| RLD between plants at 90 DAH | 0.12 | −0.00 |
| RLD between rows at 90 DAH | 0.35 * | 0.12 |
| RLD between plants at 270 DAH | −0.00 | −0.32 * |
| RLD between rows at 270 DAH | 0.51 ** | −0.12 |
| Cane Yield | Germination Percentage | |
|---|---|---|
| Physiological traits | ||
| At 90 DAH. | ||
| Chlorophyll fluorescence | −0.01 | 0.05 |
| RWC | 0.13 | 0.27 * |
| SCMR | 0.21 | 0.21 |
| At 180 DAH. | ||
| Chlorophyll fluorescence | 0.44 ** | 0.27 |
| RWC | 0.06 | −0.07 |
| SCMR | 0.04 | −0.23 |
| SLA | 0.18 | 0.14 |
| Stomatal conductance | 0.36 ** | 0.45 ** |
| At 270 DAH. | ||
| Chlorophyll fluorescence | 0.19 | −0.03 |
| RWC | 0.10 | 0.21 |
| SCMR | −0.07 | 0.01 |
| SLA | −0.26 | −0.05 |
| Stomatal conductance | 0.08 | −0.16 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chumphu, S.; Jongrungklang, N.; Songsri, P. Association of Physiological Responses and Root Distribution Patterns of Ratooning Ability and Yield of the Second Ratoon Cane in Sugarcane Elite Clones. Agronomy 2019, 9, 200. https://doi.org/10.3390/agronomy9040200
Chumphu S, Jongrungklang N, Songsri P. Association of Physiological Responses and Root Distribution Patterns of Ratooning Ability and Yield of the Second Ratoon Cane in Sugarcane Elite Clones. Agronomy. 2019; 9(4):200. https://doi.org/10.3390/agronomy9040200
Chicago/Turabian StyleChumphu, Saranya, Nuntawoot Jongrungklang, and Patcharin Songsri. 2019. "Association of Physiological Responses and Root Distribution Patterns of Ratooning Ability and Yield of the Second Ratoon Cane in Sugarcane Elite Clones" Agronomy 9, no. 4: 200. https://doi.org/10.3390/agronomy9040200
APA StyleChumphu, S., Jongrungklang, N., & Songsri, P. (2019). Association of Physiological Responses and Root Distribution Patterns of Ratooning Ability and Yield of the Second Ratoon Cane in Sugarcane Elite Clones. Agronomy, 9(4), 200. https://doi.org/10.3390/agronomy9040200