Stocking Methods and Soil Macronutrient Distributions in Southern Tallgrass Paddocks: Are There Linkages?


Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
3.1. Physical Properties
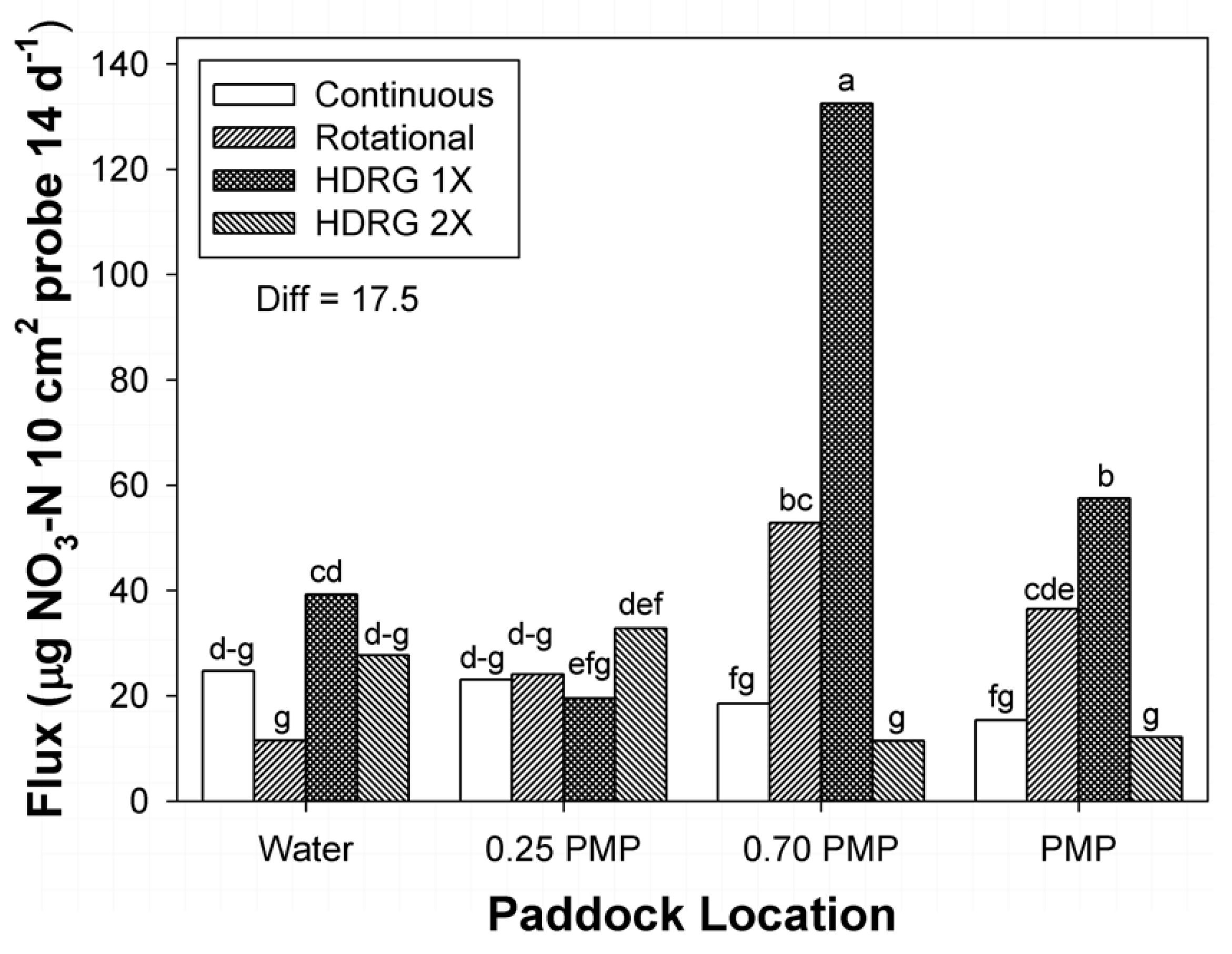
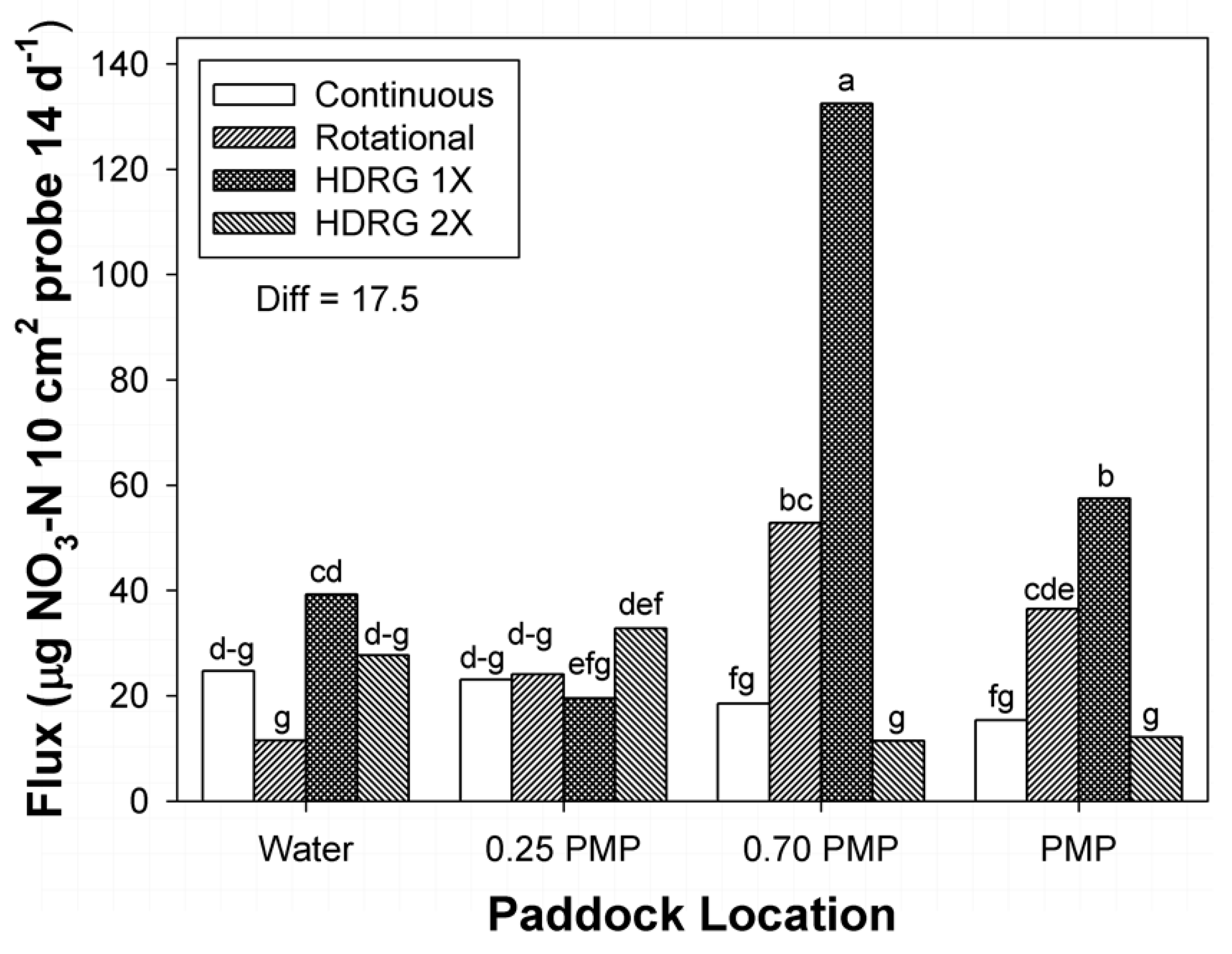
3.2. Mineral N
3.3. Phosphorus
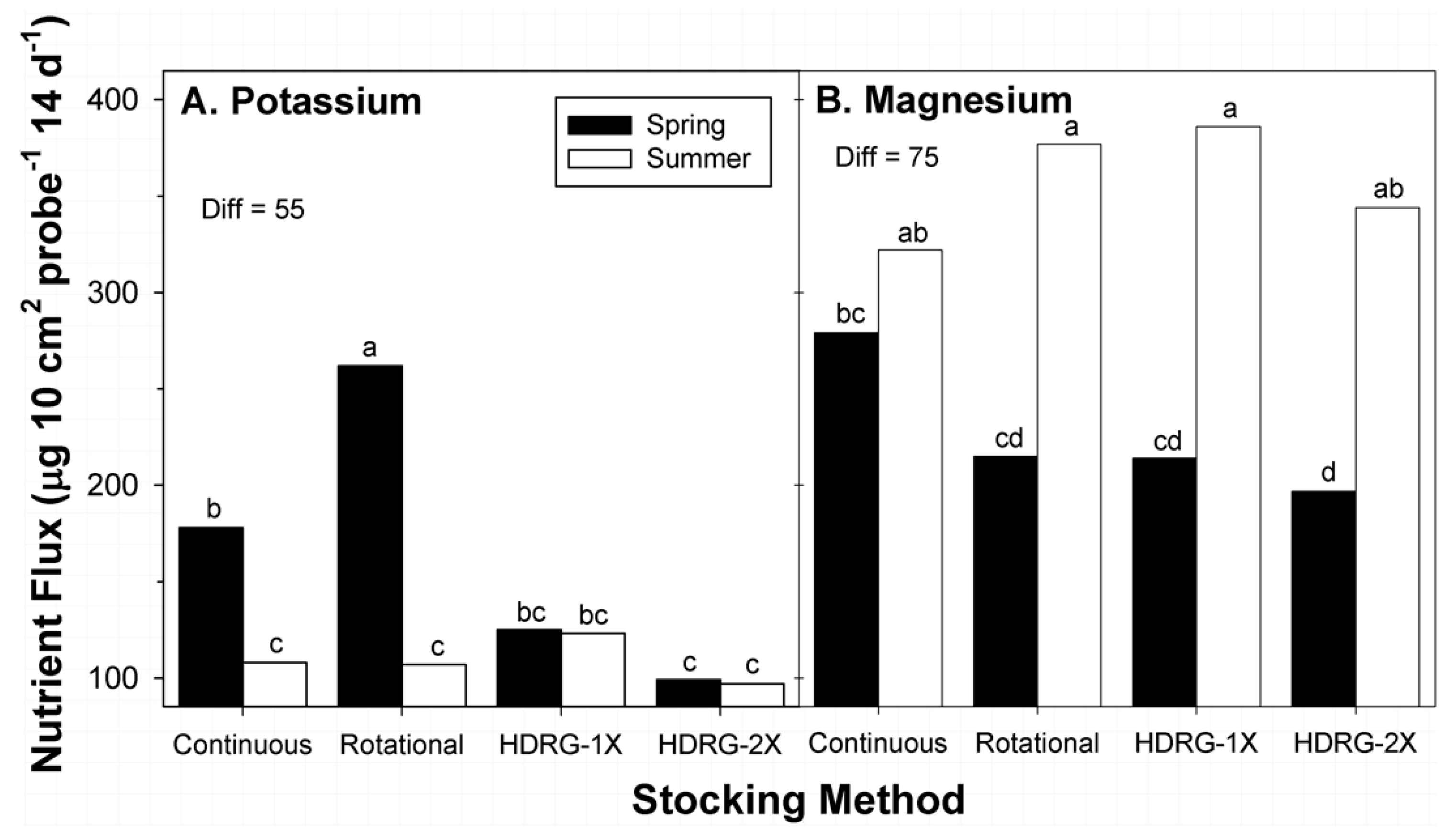
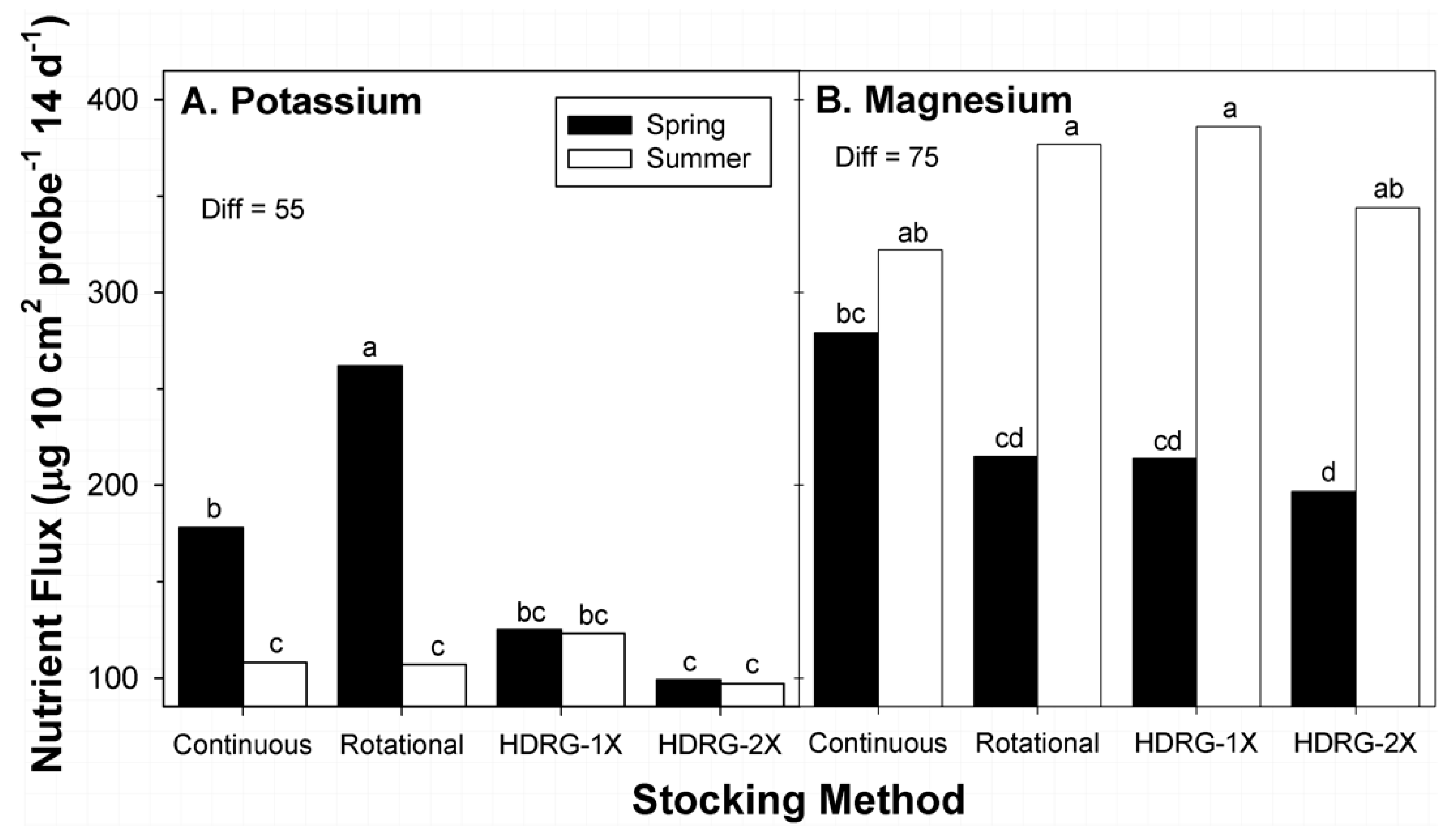
3.4. Potassium
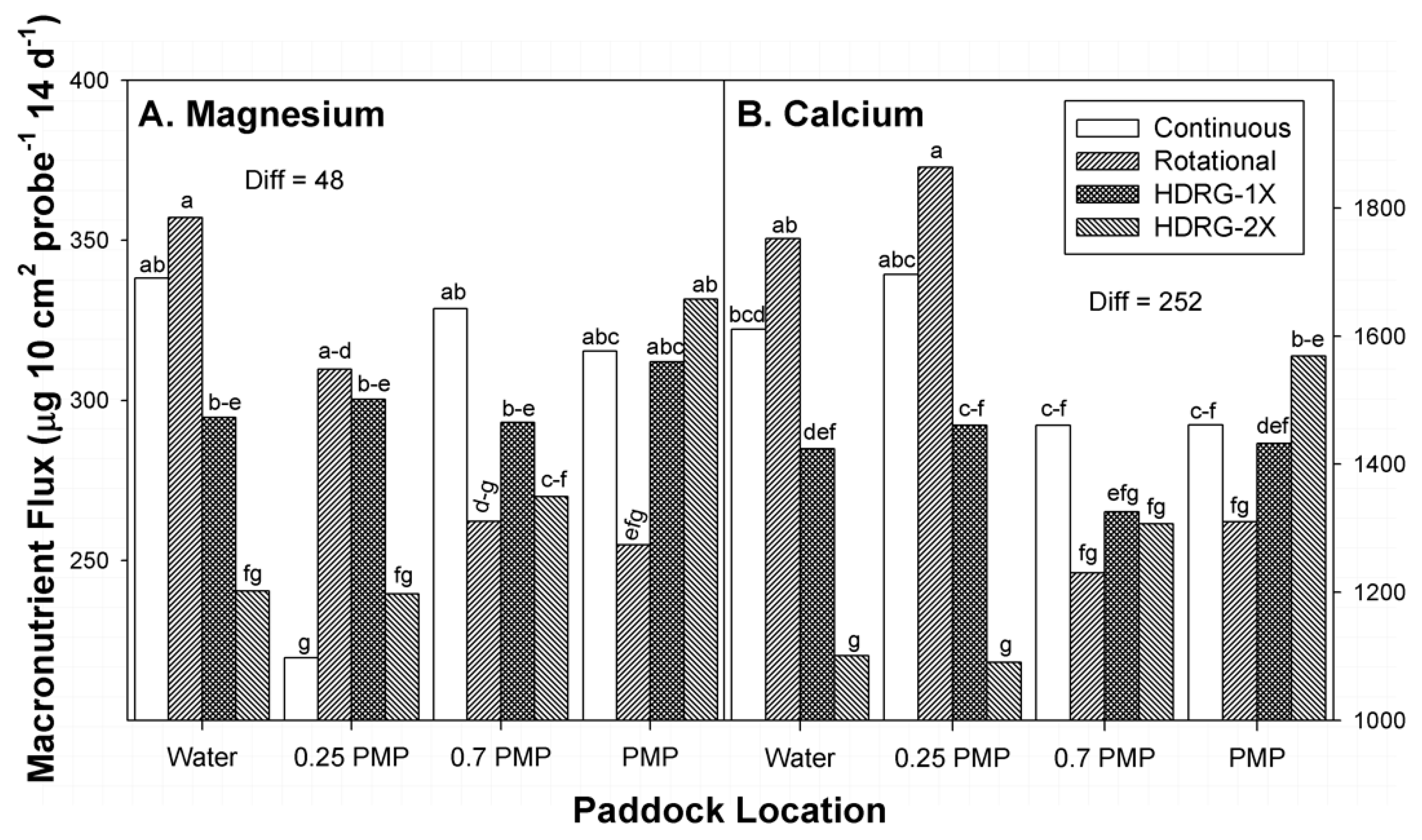
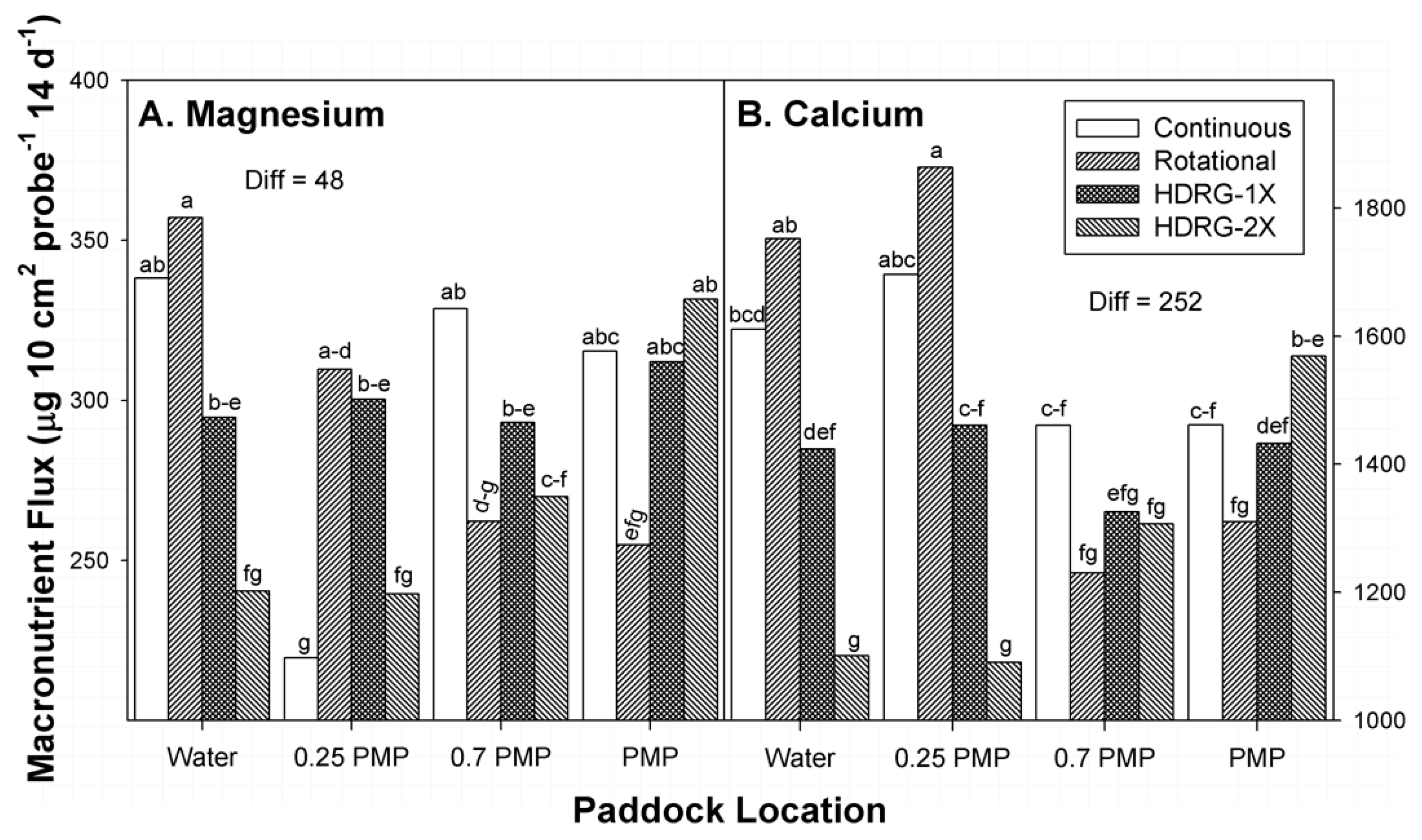
3.5. Magnesium
3.6. Calcium
3.7. Sulfur
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Archer, S.; Smeins, F.E. Ecosystem-level processes. In Grazing Management: An Ecological Perspective; Heitschmidt, R.K., Stuth, J.W., Eds.; Timber Press: Portland OR, USA, 1991; pp. 109–139. [Google Scholar]
- Jenny, H. Factors of Soil Formation: A System for Quantitative Pedology; Dover Publications: Mineola, NY, USA, 1994. [Google Scholar]
- Whitehead, D.C. Nutrient Elements in Grassland: Soil-Plant-Animal Relationships; CABI Publishing: New York, NY, USA, 2000. [Google Scholar]
- Oades, J.M. The role of biology in the formation, stabilization, and degradation of soil structure. Geoderma 1993, 56, 377–400. [Google Scholar] [CrossRef]
- Burke, I.; Lauenroth, W.; Vinton, M.; Hook, P.; Kelly, R.; Epstein, H.; Agular, M.; Robles, M.; Aguilera, M.; Murphy, M.; et al. Plant-soil interactions in temperate grasslands. Biogeochemistry 1998, 42, 121–143. [Google Scholar] [CrossRef]
- Hook, P.; Burke, I.C. Biogeochemistry in a shortgrass landscape: Control by topography, soil texture and microclimate. Ecology 2000, 81, 2686–2703. [Google Scholar] [CrossRef]
- Norman, M.J.T.; Green, J.O. The local influence of cattle dung and urine upon yield and botanical composition of permanent pasture. J. British Grassl. Soc. 1958, 13, 39–45. [Google Scholar] [CrossRef]
- Sheldrick, W.; Syers, J.K.; Lindgard, J. Contribution of livestock excreta to nutrient balances. Nutr. Cycl. Agroecosyst. 2003, 66, 119–131. [Google Scholar] [CrossRef]
- Orwin, K.H.; Bertram, J.E.; Clough, T.J.; Condron, L.M.; Sherlock, R.R.; O’Callaghan, M. Short-term consequences of spatial heterogeneity in soil nitrogen concentrations caused by urine patches of different sizes. Appl. Soil Ecol. 2009, 42, 271–278. [Google Scholar] [CrossRef]
- Bailey, D.W.; Gross, J.E.; Laca, E.A.; Rittenhouse, L.R.; Coughenor, M.B.; Swift, D.M.; Sims, P.L. Mechanisms that result in large herbivore grazing patterns. J. Range Manag. 1996, 49, 386–400. [Google Scholar] [CrossRef]
- Ganskopp, D.C.; Bohnert, D.W. Landscape nutritional patterns and cattle distribution in rangeland pastures. Appl. Anim. Behav. Sci. 2009, 116, 110–119. [Google Scholar] [CrossRef]
- Schnyder, H.; Locher, F.; Auerswald, K. Nutrient redistribution by grazing cattle drives patterns of topsoil N and P stocks in a low-input pasture ecosystem. Nutr. Cycl. Agroecosyst. 2010, 88, 183–195. [Google Scholar] [CrossRef]
- Augustine, D.J.; Milchunas, D.G.; Derner, J.D. Spatial redistribution of nitrogen by cattle in semiarid rangeland. Rangeland Ecol. Manag. 2013, 66, 56–62. [Google Scholar] [CrossRef]
- Saunders, W.H.M. Effects of cow urine and its major constituents on pasture properties. N. Z. J. Agric. Res. 1982, 25, 61–68. [Google Scholar] [CrossRef]
- Mathews, B.W.; Sollenberger, L.E.; Nair, V.D.; Staples, C.R. Impact of grazing management on soil nitrogen, phosphorus, potassium, and sulfur distribution. J. Environ. Qual. 1994, 23, 1006–1013. [Google Scholar] [CrossRef]
- Barnes, M.K.; Norton, B.E.; Maeno, M.; Malachek, J.C. Paddock size and stocking density affect spatial heterogeneity of grazing. Rangeland Ecol. Manag. 2008, 61, 380–388. [Google Scholar] [CrossRef]
- USDA-NRCS. Soil Survey of Canadian County, Oklahoma. Supplement Manuscript; USDA-NRCS and Okla. Agric. Exp. Stn.: Stillwater, OK, USA, 1999.
- Northup, B.K.; Phillips, W.A.; Daniel, J.A.; Mayeux, H.S. Managing southern tallgrass prairie: Case studies on grazing and climatic effects. In Proceedings of the 2nd National Conference on Grazing Lands, Nashville, TN, USA, 8–10 December 2003; Theurer, M., Peterson, J., Golla, M., Eds.; Omnipress Inc.: Madison, WI, USA, 2003; pp. 834–890. [Google Scholar]
- Phillips, W.A.; Coleman, S.A. Productivity and economic return of three warm-season grass stocker systems of the southern Great Plains. J. Prod. Agric. 1995, 8, 334–339. [Google Scholar] [CrossRef]
- Phillips, W.A.; Northup, B.K.; Mayeux, H.S.; Daniel, J.A. Performance and economic returns of stocker cattle on tallgrass prairie under different grazing management strategies. Prof. Anim. Sci. 2003, 19, 416–423. [Google Scholar] [CrossRef]
- Goodman, J.M. Physical environments of Oklahoma. In Geography of Oklahoma; Morris, J.W., Ed.; Oklahoma Historical Society: Oklahoma City, OK, USA, 1977; pp. 9–25. [Google Scholar]
- Briske, D.D.; Derner, J.D.; Brown, J.R.; Fuhlendorf, S.D.; Teague, W.R.; Havstad, K.M.; Gillen, R.L.; Ash, A.J.; Willms, W.D. Rotational grazing on rangelands: Reconciliation of perception and experimental evidence. Rangeland Ecol. Manag. 2008, 61, 3–17. [Google Scholar] [CrossRef]
- Arnold, G.W.; Dudzinski, M.L. Ethology of Free-Ranging Domestic Animals; Elsevier Scientific Publ.: New York, NY, USA, 1979. [Google Scholar]
- Grossman, R.B.; Reinsch, T.G. Bulk density and linear extensibility. In Methods of Soil Analysis. Part. 4. Physical Methods; Dane, J.H., Topp, G.C., Eds.; SSSA: Madison, WI, USA, 2002; pp. 201–228. [Google Scholar]
- Day, P.R. Particle fractionation and particle size analysis. In Methods of Soil Analysis, Part. I; Black, C.A., Evans, D.D., White, J.L., Ensminger, L.E., Clark, F.E., Eds.; Agronomy Monograph 9; ASA and SSSA: Madison, WI, USA, 1965; pp. 545–567. [Google Scholar]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon, and organic matter. In Methods of Soil Analysis: Part. 3. Chemical Methods; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Eds.; SSSA: Madison, WI, USA, 1996; pp. 961–1010. [Google Scholar]
- Steel, R.G.D.; Torrie, J.H. Principles and Procedures of Statistics: A Biometrical Approach, 2nd ed.; McGraw-Hill: New York, NY, USA, 1980. [Google Scholar]
- Littel, R.C.; Milliken, G.A.; Stroup, W.W.; Wolfinger, R.D. SAS Systems for Mixed Models; SAS Institute Inc.: Cary, NC, USA, 1996. [Google Scholar]
- Patetta, M. Longitudinal Data Analysis with Discrete and Continuous Responses: Course Notes for Instructor-Based Training; SAS Institute Inc.: Cary, NC, USA, 2005. [Google Scholar]
- Buol, S.W.; Hole, F.D.; McCracken, R.J. Soil Genesis and Classification, 2nd ed.; Iowa State University Press: Ames, IA, USA, 1980. [Google Scholar]
- Daniel, J.A.; Potter, K.N.; Altom, W.; Aljoe, H.; Stevens, R. Long-term grazing density impacts on soil compaction. Trans. ASAE. 2002, 45, 1911–1915. [Google Scholar] [CrossRef]
- Northup, B.K.; Daniel, J.A. Distribution of soil bulk density and organic matter along an elevation gradient in central Oklahoma. Trans. ASABE. 2010, 53, 1749–1757. [Google Scholar] [CrossRef]
- Williams, R.D.; Ahuja, L.R.; Naney, J.W.; Ross, J.D.; Barnes, B.B. Spatial trends and variability of soil properties and crop yield in a small watershed. Trans. ASAE. 1987, 30, 1653–1660. [Google Scholar] [CrossRef]
- Dubeux, J.C.B., Jr.; Sollenberger, L.E.; Vendramini, J.M.B.; Interrante, S.M.; Lira, M.A., Jr. Stocking methods, animal behavior, and soil nutrient redistribution: How are they linked? Crop. Sci. 2014, 54, 2341–2350. [Google Scholar] [CrossRef]
- Jones, G.B.; Tracy, B.J. Pasture soil and herbage nutrient dynamics through five years of rotational stocking. Crop. Sci. 2014, 54, 2351–2361. [Google Scholar] [CrossRef]
- Haynes, R.J.; Williams, P.H. Nutrient cycling and soil fertility in the grazed pasture ecosystem. Adv. Agron. 1993, 49, 119–199. [Google Scholar]
- Dickinson, C.H.; Craig, J. Effects of water on the decomposition and release of nutrients from cow pats. New Phytol. 1990, 115, 139–147. [Google Scholar] [CrossRef]
- Underhay, V.H.S.; Dickinson, C.H. Water, mineral and energy fluctuations in decomposing cow pats. J. Br. Grassl. Soc. 1978, 33, 189–196. [Google Scholar] [CrossRef]
- Nguyen, M.L.; Goh, K.M. Sulphur cycling and its implications on sulphur fertilizer requirements of grazed grassland ecosystems. Agric. Ecosys. Environ. 1994, 49, 173–206. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Paddock | Paddock Locations | ||||
|---|---|---|---|---|---|
| Stocking Method | Size | Water | 0.25 PMP | 0.7 PMP | PMP |
| (ha) | --------------------- (m) --------------------- | ||||
| Continuous | 61.0 (±2) | 2(±1) | 109(±7) | 301(±9) | 435(±8) |
| Rotational | 6.7 (±2) | 2(±1) | 43(±17) | 119(±33) | 174(±29) |
| HDRG-1× | 0.4 | 2(±1) | 10(±2) | 26(±2) | 37(±2) |
| HDRG-2× | 0.2 | 2(±1) | 9(±1) | 23(±2) | 35(±2) |
| Macronutrients | |||||||
|---|---|---|---|---|---|---|---|
| Mineral N | NO3−-N | P | S | K | Mg | Ca | |
| ------(µg 10 cm−2 probe 7.5 cm soil depth−1 14 d−1) ------ | |||||||
| Statistics | |||||||
| Mean | 57 | 53 | 4.2 | 93.9 | 203 | 1444 | 292 |
| Std. deviation | 78 | 74 | 6.4 | 191.6 | 142 | 602 | 111 |
| Median | 32 | 30 | 2.1 | 27.4 | 190 | 1465 | 307 |
| Minimum | 2 | 1 | 0.2 | 0.7 | 10 | 113 | 26 |
| Maximum | 481 | 426 | 44.8 | 1124.1 | 781 | 2770 | 563 |
| Distribution | |||||||
| Skewness | 3.4 | 3.1 | 4.0 | 3.7 | 1.2 | −0.1 | −0.2 |
| Kurtosis | 13.9 | 11.2 | 19.5 | 14.4 | 2.3 | −0.6 | −0.5 |
| K-S normality | 0.3 | 0.3 | 0.3 | 0.3 | 0.1 | 0.1 | 0.1 |
| K-S P | <0.01 | <0.01 | <0.01 | <0.01 | >0.2 | >0.2 | <0.2 |
| Ln Transform | Y | Y | Y | Y | Y | N | N |
| Soil Properties | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Particle Fractions | ||||||||||
| Stocking | Clay | Silt | Sand | Bulk Density | Organic matter | |||||
| Method † | 0–7.5 | 7.5–15 | 0–7.5 | 7.5–15 | 0–7.5 | 7.5–15 | 0–7.5 | 7.5–15 | 0–7.5 | 7.5–15 |
| ----------------------- (%) ----------------------- | ------ (Mg m−3) ------ | ----- (g kg−1) ----- | ||||||||
| Continuous | 20(5) | 26(6) | 34(14) | 36(10) | 46(9) | 38(6) | 0.99(0.15) | 1.35(0.15) | 42(10) | 34(9) |
| Rotational | 20(2) | 23(2) | 45(5) | 46(2) | 35(5) | 31(2) | 0.79(0.17) | 1.39(0.15) | 43(13) | 43(12) |
| HDRG-1× | 17(2) | 21(2) | 41(4) | 37(4) | 42(4) | 42(4) | 0.93(0.12) | 1.29(0.15) | 33(8) | 35(7) |
| HDRG-2× | 18(2) | 22(2) | 55(6) | 53(3) | 27(6) | 25(2) | 0.92(0.19) | 1.37(0.10) | 35(2) | 35(4) |
| Location† | ||||
|---|---|---|---|---|
| Stocking Method ‡ | Water | 0.25 PMP | 0.7 PMP | PMP |
| ---(µg 10 cm−2 probe 7.5 cm soil−1 14 d−1) --- | ||||
| Continuous | 31.7 cde | 27.2 def | 23.1 efg | 25.0 ef |
| Rotational | 19.1 fg | 32.4 cde | 61.0 b | 42.5 c |
| HDRG-1× | 42.3 c | 22.2 efg | 141.1 a | 61.2 b |
| HDRG-2× | 37.5 cd | 33.9 cde | 11.9 g | 12.2 g |
| Diff | 11.9 | |||
| Time of Year | |||
|---|---|---|---|
| Main Effect | Level of Effect | Spring | Summer |
| Stocking Method † | Continuous | 3.2 bc | 5.1 ab |
| Rotational | 6.7 a | 2.5 c | |
| HDRG-1׆ | 2.4 c | 2.4 c | |
| HDRG-2׆ | 2.3 c | 2.3 c | |
| Diff | 2.5 | ||
| Location ‡ | Water | 3.2 b | 2.5 b |
| 0.25 PMP | 6.3 a | 3.3 b | |
| 0.7 PMP | 2.3 b | 3.6 ab | |
| PMP | 2.5 b | 2.3 b | |
| Diff | 2.7 | ||
| Location † | ||||
|---|---|---|---|---|
| Stocking Method ‡ | Water | 0.25 PMP | 0.7 PMP | PMP |
| (µg P 10 cm−2 probe 7.5 cm soil depth−1 14 d−1) | ||||
| Continuous | 3.2 cd | 5.5 a | 1.6 d | 1.3 d |
| Rotational | 1.5 d | 4.1 ab | 3.5 bc | 1.7 d |
| HDRG-1× | 1.3 d | 2.1 c | 1.1 d | 1.4 d |
| HDRG-2× | 1.4 d | 1.2 d | 1.5 d | 1.5 d |
| Diff | 1.9 | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Northup, B.K.; Starks, P.J.; Turner, K.E. Stocking Methods and Soil Macronutrient Distributions in Southern Tallgrass Paddocks: Are There Linkages? Agronomy 2019, 9, 281. https://doi.org/10.3390/agronomy9060281
Northup BK, Starks PJ, Turner KE. Stocking Methods and Soil Macronutrient Distributions in Southern Tallgrass Paddocks: Are There Linkages? Agronomy. 2019; 9(6):281. https://doi.org/10.3390/agronomy9060281
Chicago/Turabian StyleNorthup, Brian K., Patrick J. Starks, and Kenneth E. Turner. 2019. "Stocking Methods and Soil Macronutrient Distributions in Southern Tallgrass Paddocks: Are There Linkages?" Agronomy 9, no. 6: 281. https://doi.org/10.3390/agronomy9060281
APA StyleNorthup, B. K., Starks, P. J., & Turner, K. E. (2019). Stocking Methods and Soil Macronutrient Distributions in Southern Tallgrass Paddocks: Are There Linkages? Agronomy, 9(6), 281. https://doi.org/10.3390/agronomy9060281