Temporal and Organ-specific Responses in NUE Traits to N Fertilization, Fungicide Intensity and Early Sowing in Winter Wheat Cultivars


Abstract
:1. Introduction
2. Materials and Methods
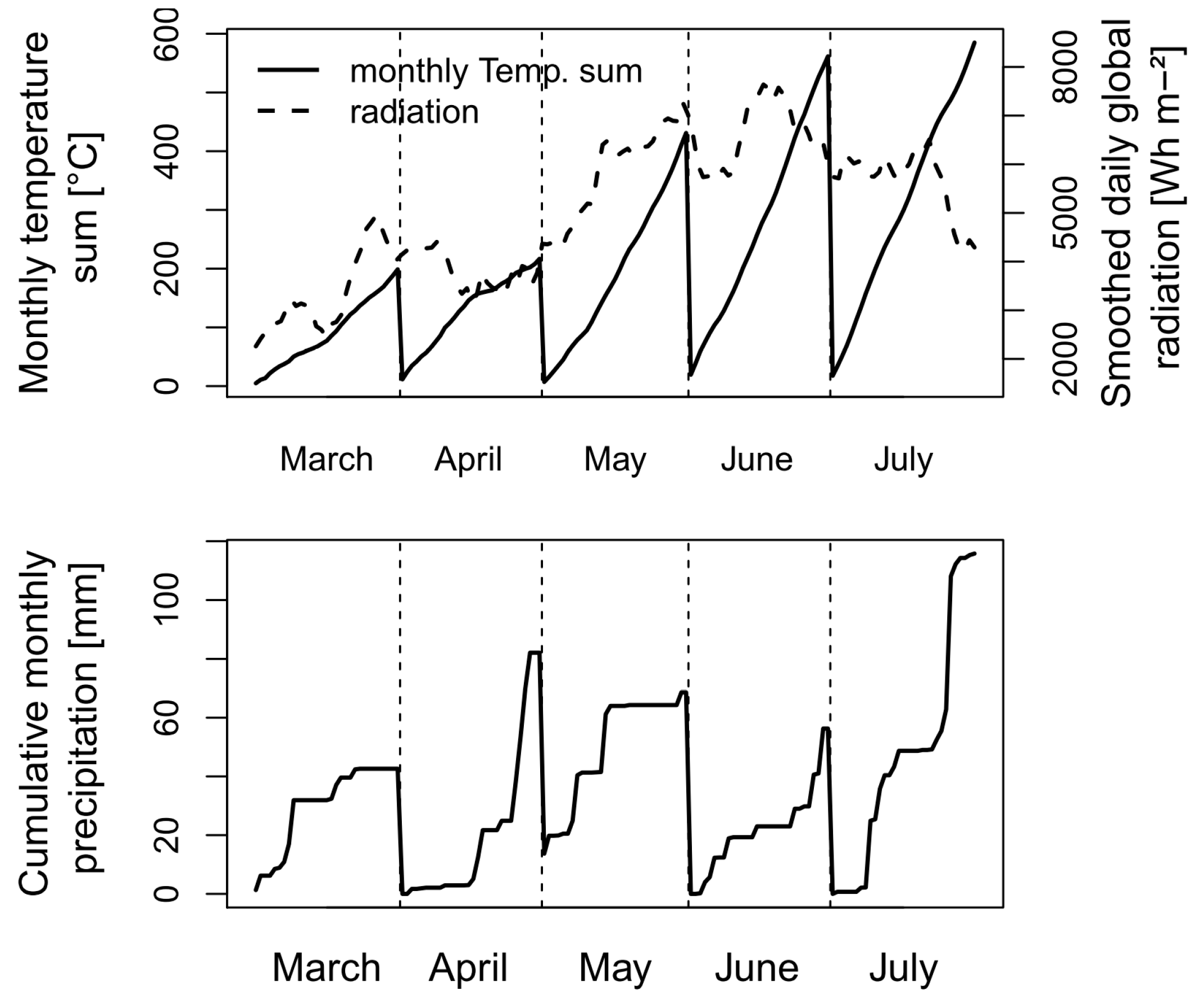
2.1. Study Site and Experiment
2.2. Plant Sampling and Sample Analysis
2.3. Calculation of Derived Plant Traits
2.4. Statistical Analysis
3. Results
3.1. Treatment Effects
3.2. Comparison of Trait Values and Trait Relationships
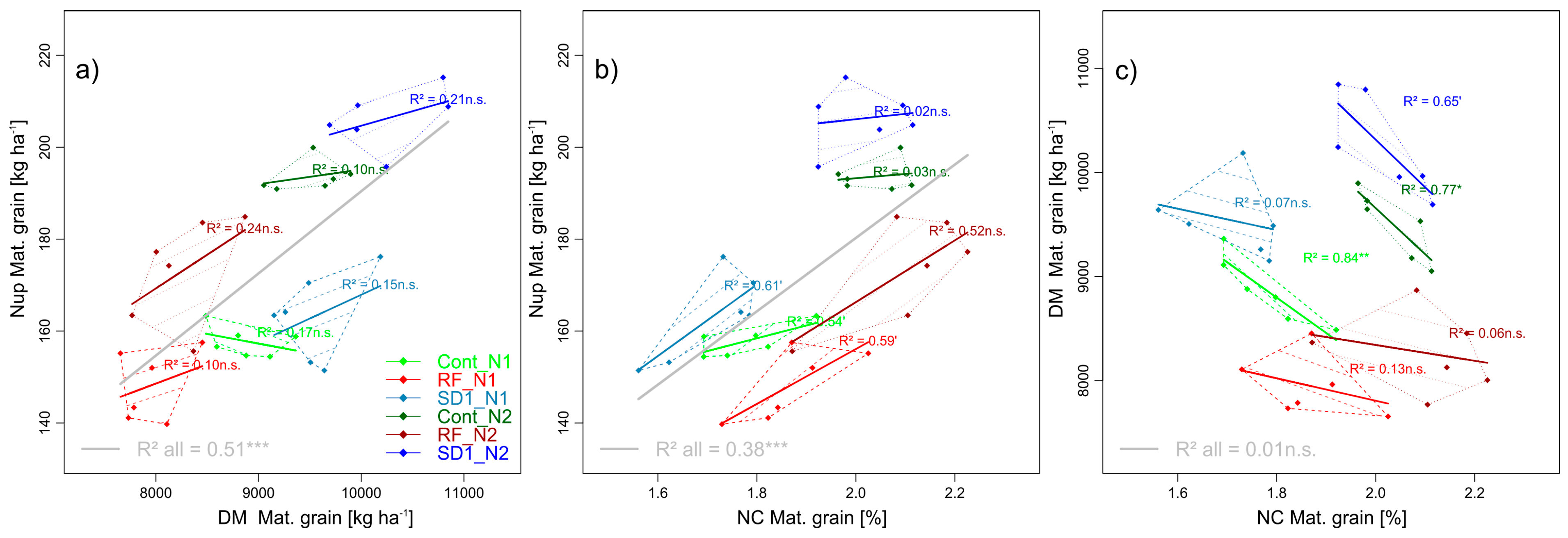
3.2.1. Trait Complex a: GNup = GY × GNC
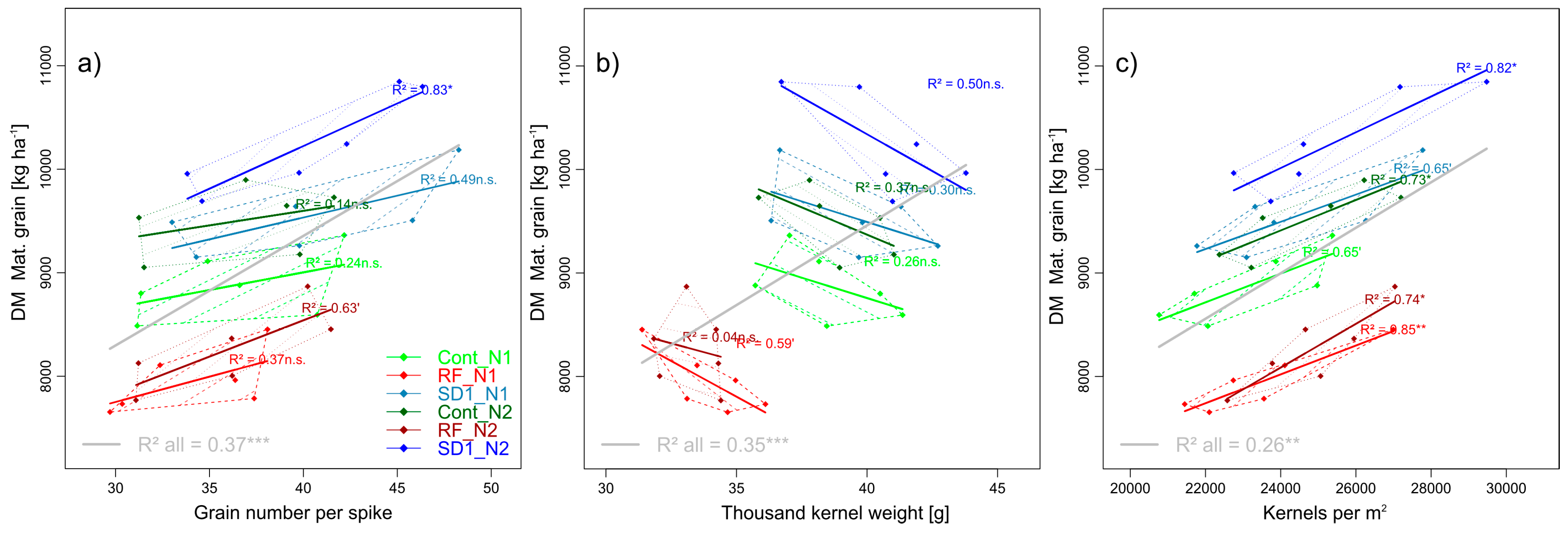
3.2.2. Trait Complex b: GY = SD × GNS × TKW
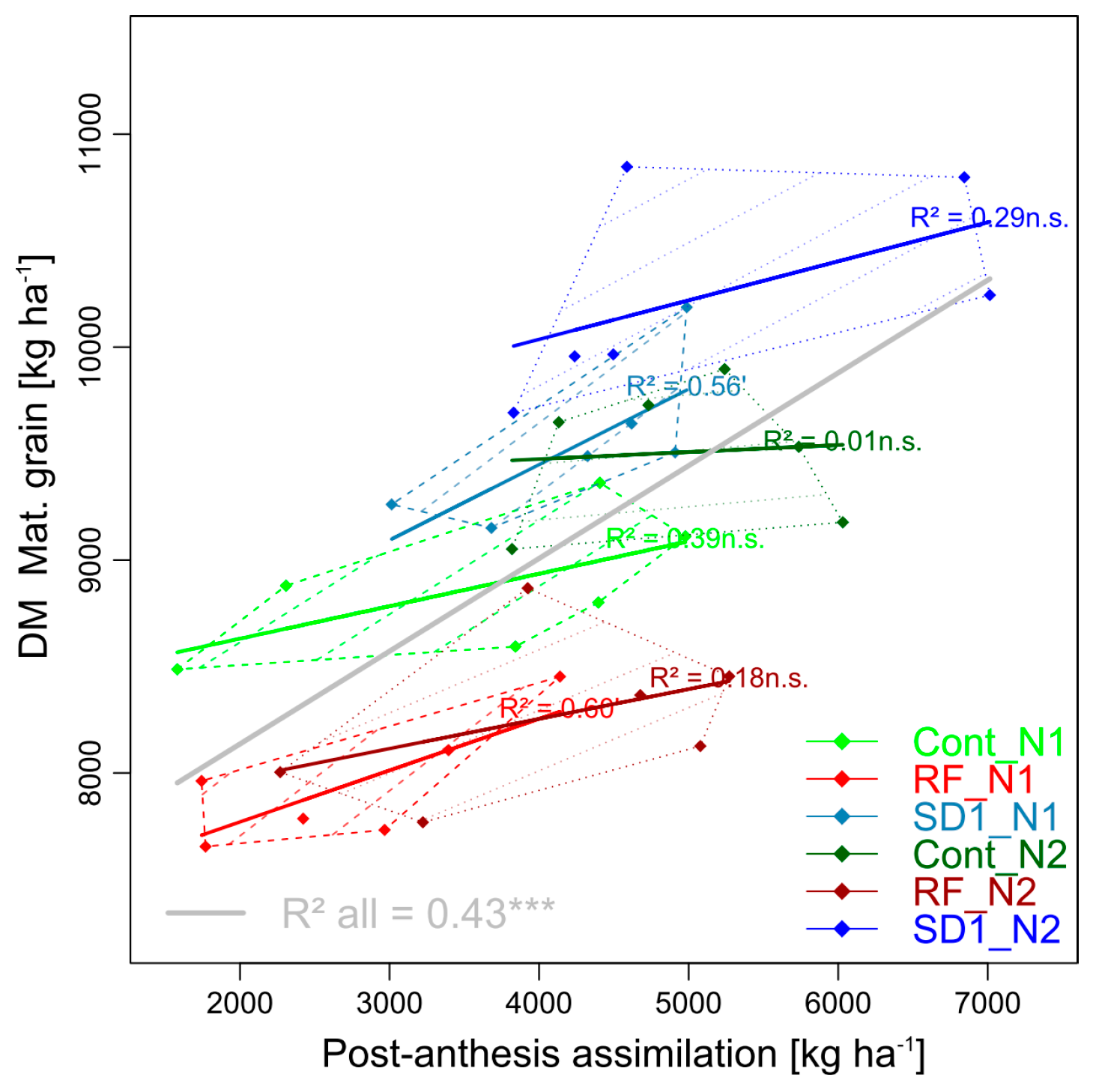
3.2.3. Trait Complex c: GY = DMT + PAA
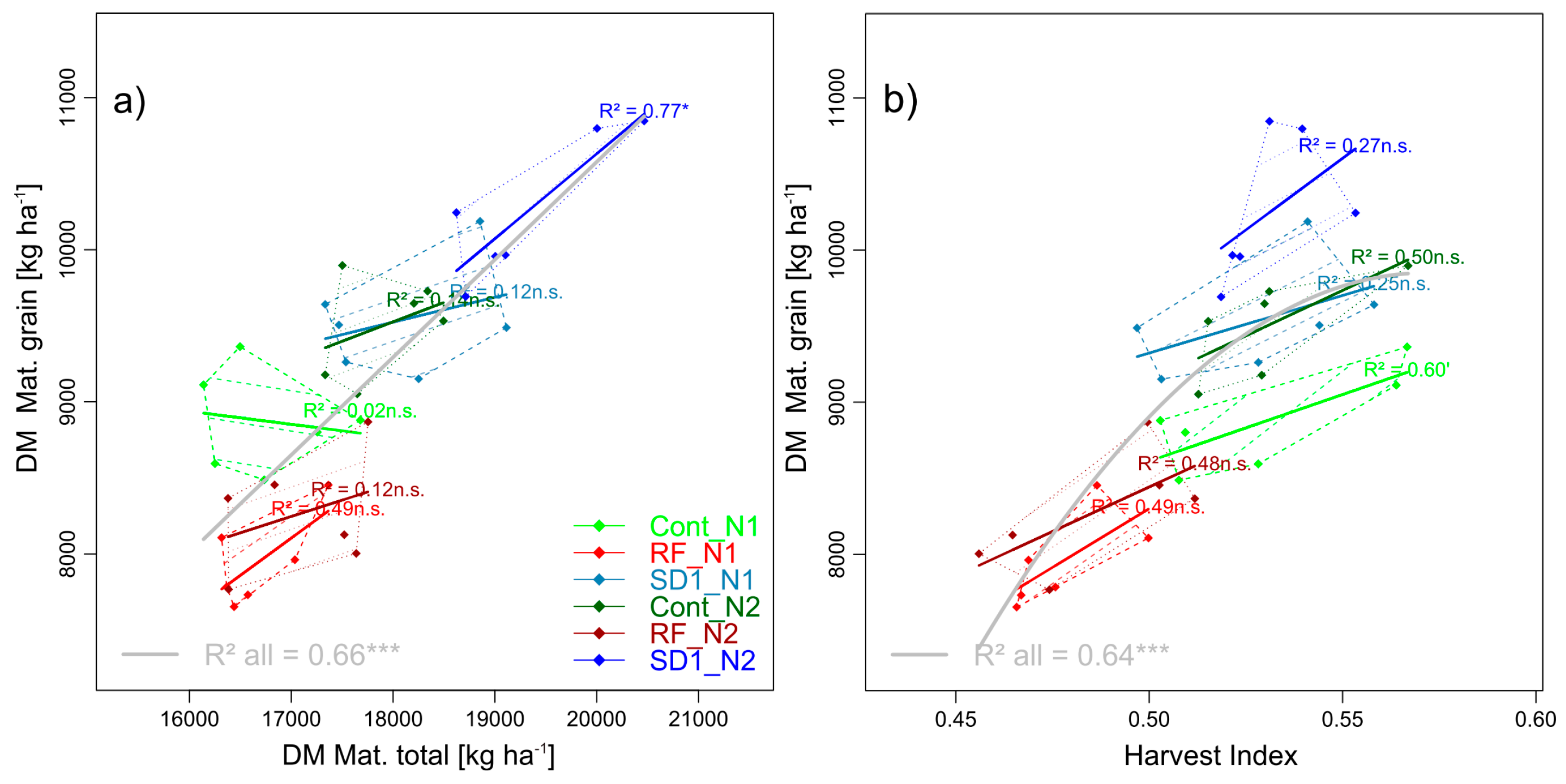
3.2.4. Trait Complex d: GY = DM × HI
3.2.5. Trait Complex e: NUE_total = NupEff xNutEff_total and Trait Complex f: NUE_grain = NupEff × NutEff_grain
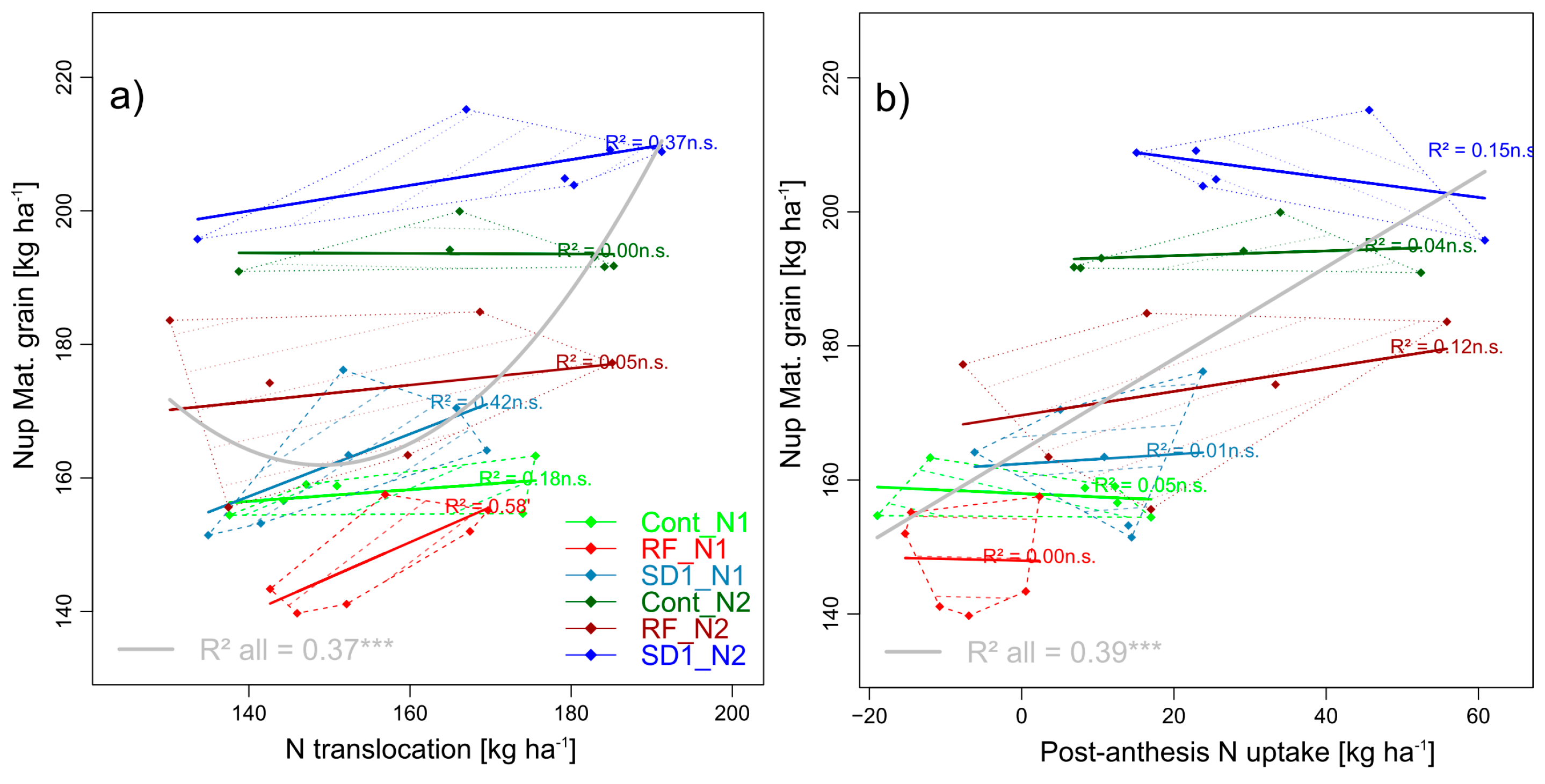
3.2.6. Trait COMPLEX g: GNup = NT + PANup
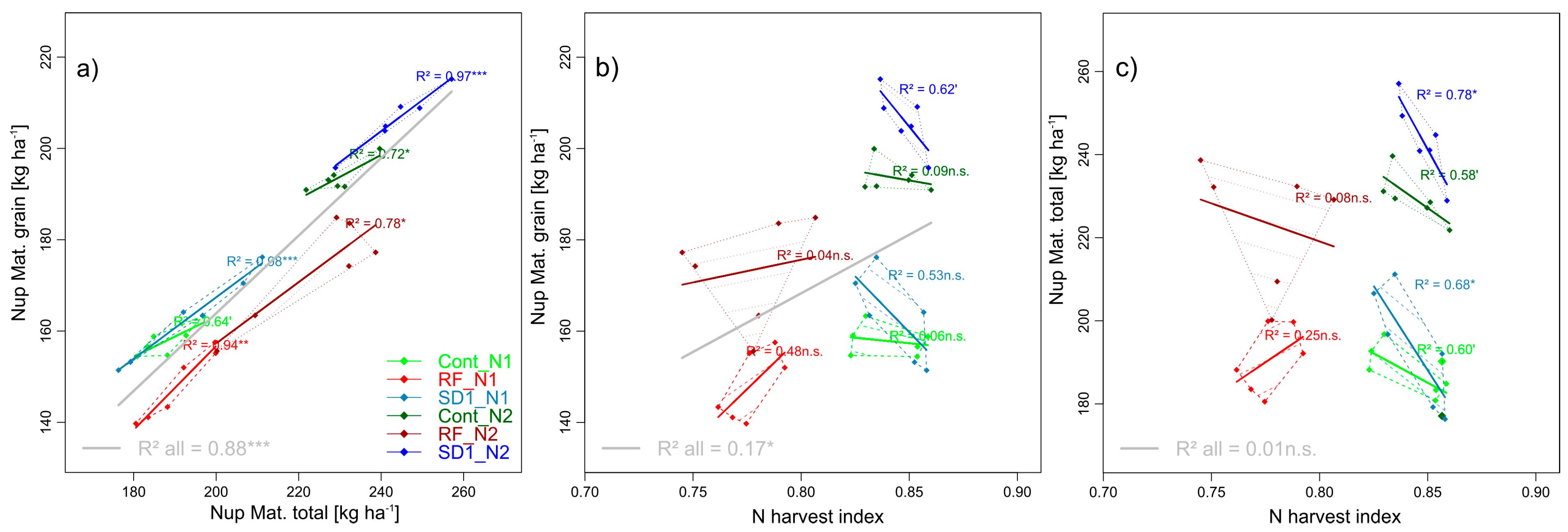
3.2.7. Trait Complex h: GNup = Nup × NHI
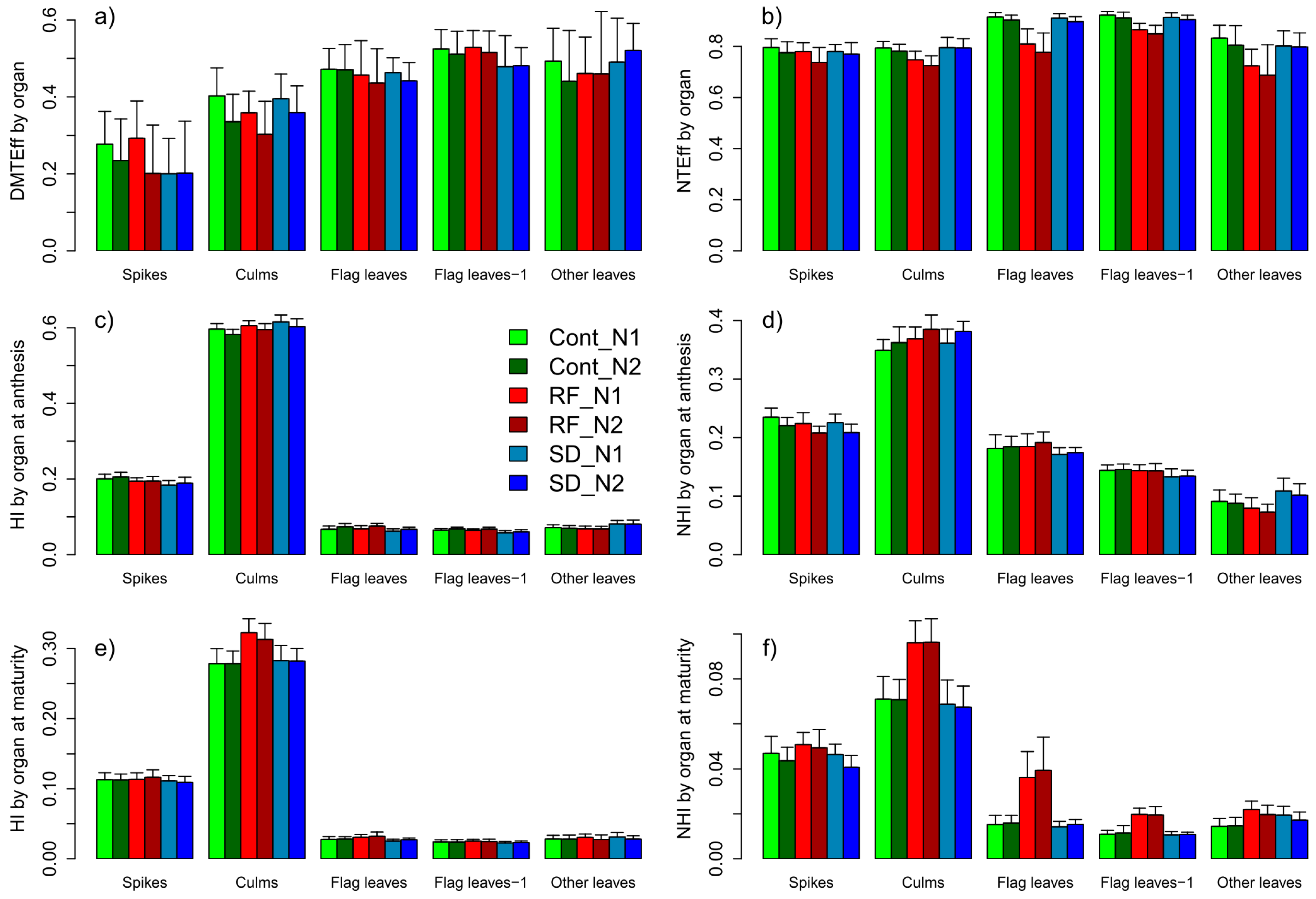
3.2.8. Trait Complex i: DMT = DMAnt × DMTEff
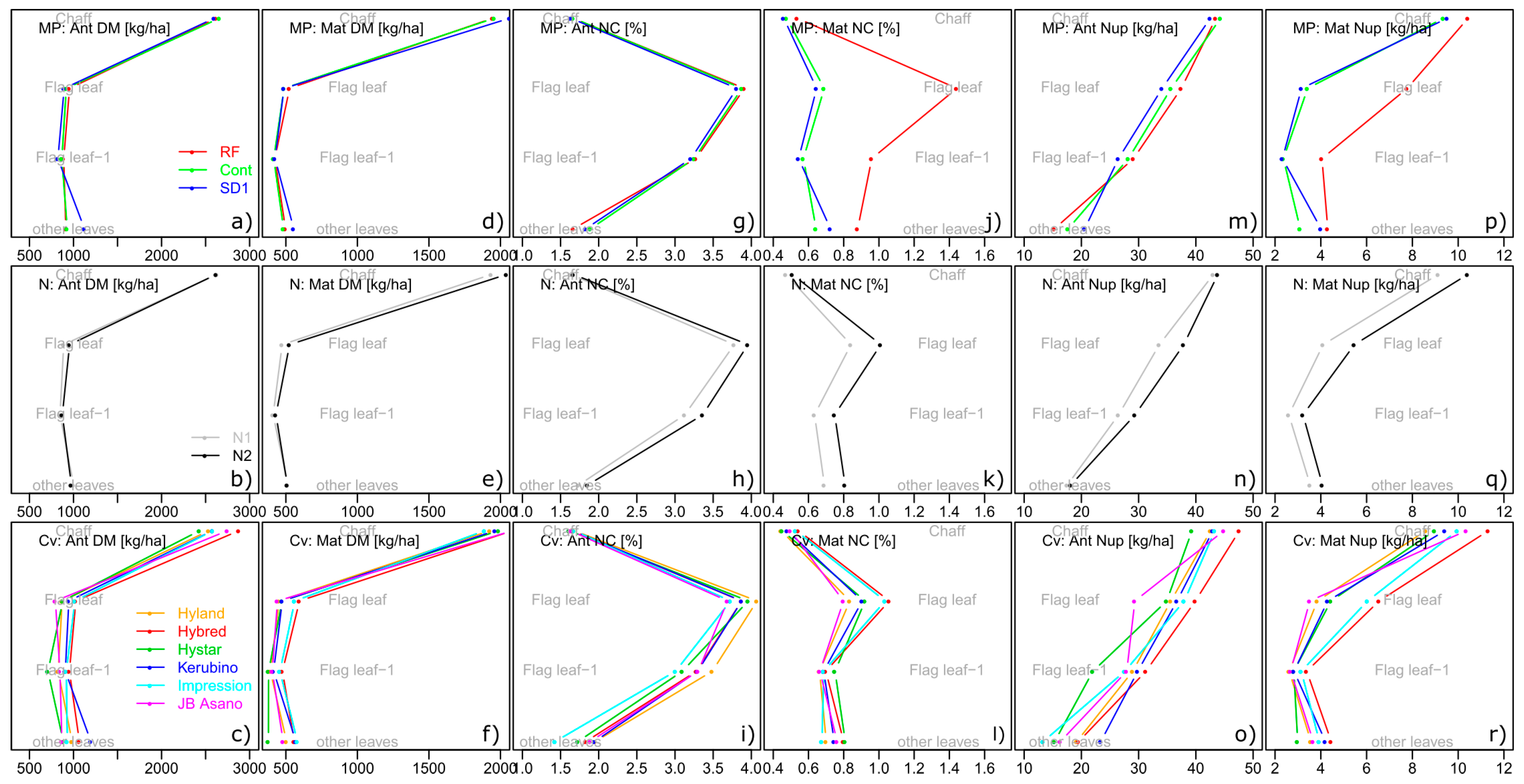
3.2.9. Trait Complex j: NupAnt = Nupspikes + Nupculm + Nupflag leaf + Nupflag leaf-1 + Nupother leaves
3.2.10. Trait Complex k: Nuptotal = Nupgrain + Nupspike + Nupculm + Nupflag leaf + Nupflag leaf-1 + Nupother leaves
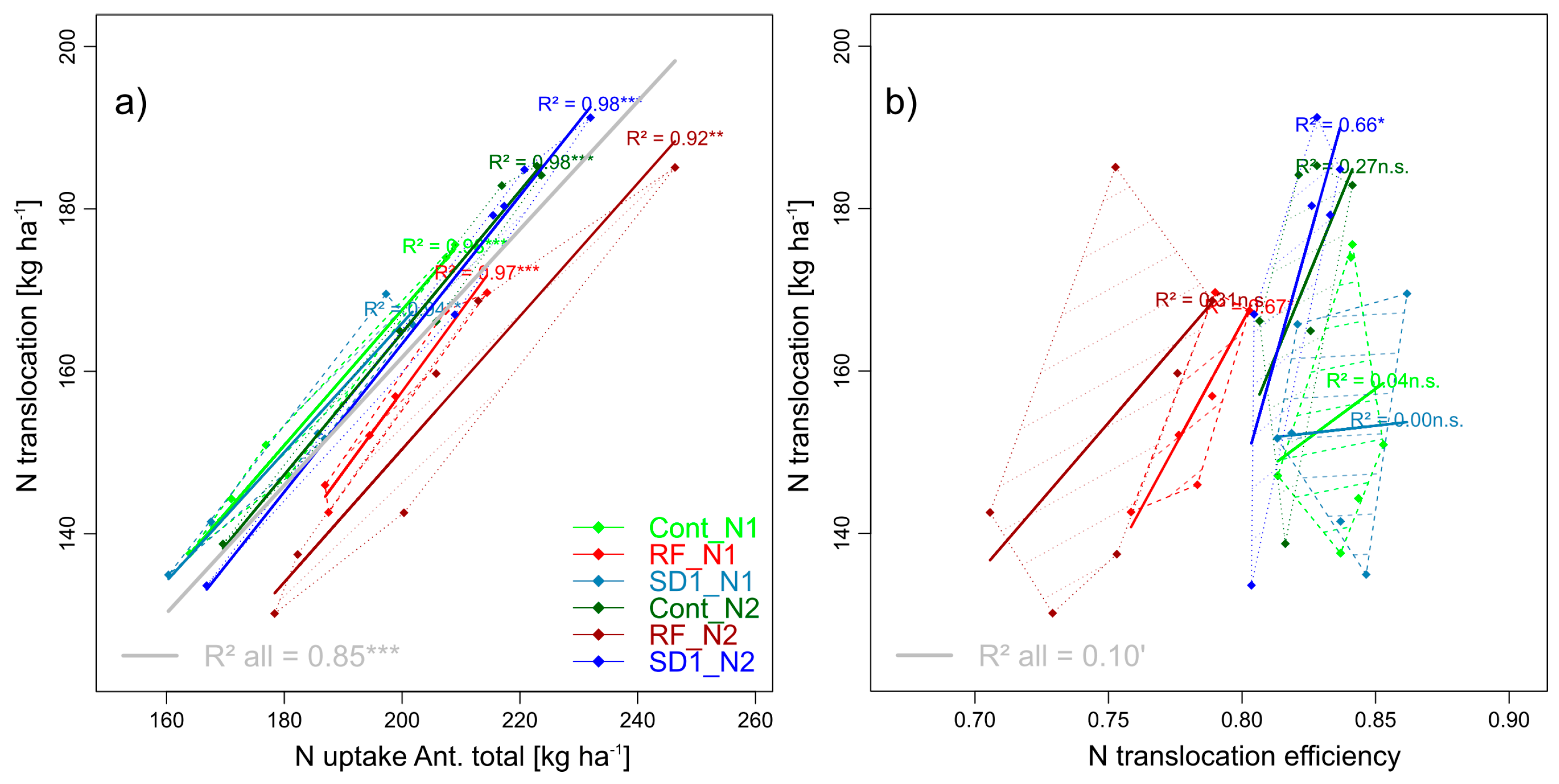
3.2.11. Trait Complex l: NT = NTspike + NTculm + NTflag leaf + NTflag leaf-1 + NTother leaves and trait complex m: NT = NupAnt × NTEff
4. Discussion
4.1. Trait Level of Dry Matter and N Traits
4.2. Dissecting the Grain Yield Response to Fungicide
4.3. Dissecting the Grain Yield Response to Early Sowing
4.4. Dissecting the Response in Grain N Uptake to Reduced Fungicide
4.5. Dissecting the Response in Grain N Uptake to Early Sowing
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mueller, N.D.; Gerber, J.S.; Johnston, M.; Ray, D.K.; Ramankutty, N.; Foley, J.A. Closing yield gaps through nutrient and water management. Nature 2012, 490, 254–257. [Google Scholar] [CrossRef] [PubMed]
- Galloway, J.N.; Cowling, E.B. Reactive nitrogen and the world: 200 years of change. AMBIO A J. Hum. Environ. 2002, 31, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Hirel, B.; Lemaire, G. From agronomy and ecophysiology to molecular genetics for improving nitrogen use efficiency in crops. J. Crop Improv. 2006, 15, 213–257. [Google Scholar] [CrossRef]
- Parry, M.A.J.; Reynolds, M.; Salvucci, M.E.; Raines, C.; Andralojc, P.J.; Zhu, X.G.; Price, G.D.; Condon, A.G.; Furbank, R.T. Raising yield potential of wheat. II. Increasing photosynthetic capacity and efficiency. J. Exp. Bot. 2011, 62, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Cormier, F.; Faure, S.; Dubreuil, P.; Heumez, E.; Beauchêne, K.; Lafarge, S.; Praud, S.; Le Gouis, J. A multi-environmental study of recent breeding progress on nitrogen use efficiency in wheat (Triticum aestivum L.). Theor. Appl. Genet. 2013, 126, 3035–3048. [Google Scholar] [CrossRef]
- Guttieri, M.J.; Frels, K.; Regassa, T.; Waters, B.M.; Baenziger, P.S. Variation for nitrogen use efficiency traits in current and historical great plains hard winter wheat. Euphytica 2017, 213, 1–18. [Google Scholar] [CrossRef]
- Latshaw, S.P.; Vigil, M.F.; Haley, S.D. Genotypic Differences for Nitrogen Use Efficiency and Grain Protein Deviation in Hard Winter Wheat. Agron. J. 2016, 108, 2201. [Google Scholar] [CrossRef]
- Zecevic, V.; Boskovic, J.; Knezevic, D.; Micanovic, D. Effect of seeding rate on grain quality of winter wheat. Chil. J. Agric. Res. 2014, 74, 23–28. [Google Scholar] [CrossRef] [Green Version]
- Ehdaie, B.; Waines, J.G. Sowing date and nitrogen rate effects on dry matter and nitrogen partitioning in bread and durum wheat. Field Crop. Res. 2001, 73, 47–61. [Google Scholar] [CrossRef]
- Ferrise, R.; Triossi, A.; Stratonovitch, P.; Bindi, M.; Martre, P. Sowing date and nitrogen fertilisation effects on dry matter and nitrogen dynamics for durum wheat: An experimental and simulation study. Field Crop. Res. 2010, 117, 245–257. [Google Scholar] [CrossRef]
- Efretuei, A.; Gooding, M.; White, E.; Spink, J.; Hackett, R. Effect of nitrogen fertilizer application timing on nitrogen use efficiency and grain yield of winter wheat in Ireland. Ir. J. Agric. Food Res. 2016, 55, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, G.N.; Kant, S. Improving nitrogen use efficiency in plants: Effective phenotyping in conjunction with agronomic and genetic approaches. Funct. Plant Biol. 2018. [Google Scholar] [CrossRef]
- Gooding, M.J.; Gregory, P.J.; Ford, K.E.; Pepler, S. Fungicide and cultivar affect post-anthesis patterns of nitrogen uptake, remobilization and utilization efficiency in wheat. J. Agric. Sci. 2005, 143, 503. [Google Scholar] [CrossRef]
- Ruske, R.E.; Gooding, M.J.; Jones, S.A. The effects of triazole and strobilurin fungicide programmes on nitrogen uptake, partitioning, remobilization and grain N accumulation in winter wheat cultivars. J. Agric. Sci. 2003, 140, 395–407. [Google Scholar] [CrossRef]
- Munns, R.; James, R.A.; Sirault, X.R.R.; Furbank, R.T.; Jones, H.G. New phenotyping methods for screening wheat and barley for beneficial responses to water deficit. J. Exp. Bot. 2010, 61, 3499–3507. [Google Scholar] [CrossRef] [Green Version]
- Cammarano, D.; Basso, B.; Stefanova, L.; Grace, P. Adapting wheat sowing dates to projected climate change in the Australian subtropics: Analysis of crop water use and yield. Crop Pasture Sci. 2012, 63, 974. [Google Scholar] [CrossRef]
- Kipp, S.; Mistele, B.; Baresel, P.; Schmidhalter, U. High-throughput phenotyping early plant vigour of winter wheat. Eur. J. Agron. 2014, 52, 271–278. [Google Scholar] [CrossRef]
- Subedi, K.D.; Ma, B.L.; Xue, A.G. Planting date and nitrogen effects on grain yield and protein content of spring wheat. Crop Sci. 2007, 47, 36–44. [Google Scholar] [CrossRef]
- Rasmussen, I.S.; Thorup-Kristensen, K. Does earlier sowing of winter wheat improve root growth and N uptake? Field Crop. Res. 2016, 196, 10–21. [Google Scholar] [CrossRef]
- Kiss, T.; Balla, K.; Bányai, J.; Veisz, O.; Karsai, I. Effect of different sowing times on the plant developmental parameters of wheat (Triticum aestivum L.). Cereal Res. Commun. 2014, 42, 239–251. [Google Scholar] [CrossRef]
- Semenov, M.A.; Stratonovitch, P.; Alghabari, F.; Gooding, M.J. Adapting wheat in Europe for climate change. J. Cereal Sci. 2014, 59, 245–256. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, I.S.; Dresbøll, D.B.; Thorup-Kristensen, K. Winter wheat cultivars and nitrogen (N) fertilization—Effects on root growth, N uptake efficiency and N use efficiency. Eur. J. Agron. 2015, 68, 38–49. [Google Scholar] [CrossRef]
- Joergensen, L.N.; Secher, B.J.M.; Olesen, J.E.; Mortensen, J. Need for fungicide treatments when varying agricultural parameters. Asp. Appl. Biol. 1997, 50, 285–292. [Google Scholar]
- Kelley, K.W. Planting date and foliar fungicide effects on yield components and grain traits of winter wheat. Agron. J. 2001, 93, 380–389. [Google Scholar] [CrossRef]
- Milford, G.F.J.; Penny, A.; Prew, R.D.; Darby, R.J.; Todd, A.D. Effects of previous crop, sowing date, and winter and spring applications of nitrogen on the growth, nitrogen uptake and yield of winter wheat. J. Agric. Sci. 1993, 121, 1–12. [Google Scholar] [CrossRef]
- Donaldson, E.; Schillinger, W.F.; Dofing, S.M. Straw Production and Grain Yield Relationships in Winter Wheat. Crop Sci. 2001, 41, 100. [Google Scholar] [CrossRef]
- Schwarte, A.J.; Gibson, L.R.; Karlen, D.L.; Liebman, M.; Jannink, J. Planting Date Effects on Winter Triticale Dry Matter and Nitrogen Accumulation. Agron. J. 2005, 97, 1333–1341. [Google Scholar] [CrossRef] [Green Version]
- Christensen, B.T.; Jensen, J.L.; Thomsen, I.K. Impact of Early Sowing on Winter Wheat Receiving Manure or Mineral Fertilizers. Agron. J. 2017, 109, 1312. [Google Scholar] [CrossRef]
- Suarez-Tapia, A.; Rasmussen, J.; Thomsen, I.K.; Christensen, B.T. Early sowing increases nitrogen uptake and yields of winter wheat grown with cattle slurry or mineral fertilizers. J. Agric. Sci. 2018, 156, 177–187. [Google Scholar] [CrossRef]
- Mirosavljevic, M.; Przulj, N.; Momcilovic, V.; Hristov, N.; Maksimovic, I. Dry matter accumulation and remobilization in winter barley as affected by genotype and sowing date. Genetika 2015, 47, 751–763. [Google Scholar] [CrossRef]
- Ding, D.; Feng, H.; He, J.Q.; Zou, F.; Jin, J.M. Modifying Winter Wheat Sowing Date as an Adaptation to Climate Change on the Loess Plateau. Agron. J. 2016, 108, 53–63. [Google Scholar] [CrossRef] [Green Version]
- Yin, L.; Dai, X.; He, M. Delayed sowing improves nitrogen utilization e ffi ciency in winter wheat without impacting yield. Field Crop. Res. 2018, 221, 90–97. [Google Scholar] [CrossRef]
- Singh, R.P.; Singh, P.K.; Rutkoski, J.; Hodson, D.P.; He, X.; Jørgensen, L.N.; Hovmøller, M.S.; Huerta-Espino, J. Disease Impact on Wheat Yield Potential and Prospects of Genetic Control. Annu. Rev. Phytopathol. 2016, 54, 303–322. [Google Scholar] [CrossRef] [Green Version]
- Figueroa, M.; Hammond-Kosack, K.E.; Solomon, P.S. A review of wheat diseases—A field perspective. Mol. Plant Pathol. 2018, 19, 1523–1536. [Google Scholar] [CrossRef]
- Bertelsen, J.R.; De Neergaard, E.; Smedegaard-Petersen, V. Fungicidal effects of azoxystrobin and epoxiconazole on phyllosphere fungi, senescence and yield of winter wheat. Plant Pathol. 2001, 50, 190–205. [Google Scholar] [CrossRef]
- Wegulo, S.; Stevens, J.; Zwingman, M.; Baenziger, P.S. Yield Response to Foliar Fungicide Application in Winter Wheat. In Fungicides for Plant and Animal Diseases; InTech: Rijeka, Croatia, 2012; pp. 227–244. ISBN 9533078049. [Google Scholar]
- Bhatta, M.; Regassa, T.; Wegulo, S.N.; Baenziger, P.S. Foliar fungicide effects on disease severity, yield, and agronomic characteristics of modern winter wheat genotypes. Agron. J. 2018, 110, 602–610. [Google Scholar] [CrossRef]
- Olesen, J.E.; Jørgensen, L.N.; Mortensen, J.V. Irrigation strategy, nitrogen application and fungicide control in winter wheat on a sandy soil. II. Radiation interception and conversion. J. Agric. Sci. 2000, 134, 13–23. [Google Scholar] [CrossRef]
- Varga, B.; Svečnjak, Z.; Maćešić, D.; Uher, D. Winter wheat cultivar responses to fungicide application are affected by nitrogen fertilization rate. J. Agron. Crop Sci. 2005, 191, 130–137. [Google Scholar] [CrossRef]
- Ishikawa, S.; Hare, M.C.; Kettlewell, P.S. Effects of strobilurin fungicide programmes and fertilizer nitrogen rates on winter wheat: Leaf area, dry matter yield and nitrogen yield. J. Agric. Sci. 2012, 150, 427–441. [Google Scholar] [CrossRef]
- Mourtzinis, S.; Marburger, D.; Gaska, J.; Diallo, T.; Lauer, J.G.; Conley, S. Corn, soybean, and wheat yield response to crop rotation, nitrogen rates, and foliar fungicide application. Crop Sci. 2017, 57, 983–992. [Google Scholar] [CrossRef]
- Gooding, M.J.; Dimmock, J.P.R.E.; France, J.; Jones, S.A. Green leaf area decline of wheat flag leaves: The influence of fungicides and relationships with mean grain weight and grain yield. Ann. Appl. Biol. 2000, 136, 77–84. [Google Scholar] [CrossRef]
- Gooding, M.J. Influence of foliar diseases and their control by fungicides on grain yield and quality in wheat. In Wheat Production in Stressed Environments; Springer: Dordrecht, The Neverlands, 2007; Volume 12, pp. 567–581. [Google Scholar]
- Bancal, M.-O.; Robert, C.; Ney, B. Modelling Wheat Growth and Yield Losses from Late Epidemics of Foliar Diseases using Loss of Green Leaf Area per Layer and Pre-anthesis Reserves. Ann. Bot. 2007, 100, 777–789. [Google Scholar] [CrossRef] [Green Version]
- Bancal, M.-O.; Roche, R.; Bancal, P. Late foliar diseases in wheat crops decrease nitrogen yield through N uptake rather than through variations in N remobilization. Ann. Bot. 2008, 102, 579–590. [Google Scholar] [CrossRef]
- Garnault, M.; Duplaix, C.; Leroux, P.; Couleaud, G.; Carpentier, F.; David, O.; Walker, A.A. Spatiotemporal dynamics of fungicide resistance in the wheat pathogen Zymoseptoria tritici in France. Pest Manag. Sci. 2019. [Google Scholar] [CrossRef]
- Wegulo, S.N.; Zwingman, M.V.; Breathnach, J.A.; Baenziger, P.S. Economic returns from fungicide application to control foliar fungal diseases in winter wheat. Crop Prot. 2011, 30, 685–692. [Google Scholar] [CrossRef] [Green Version]
- Prey, L.; von Bloh, M.; Schmidhalter, U. Evaluating RGB Imaging and Multispectral Active and Hyperspectral Passive Sensing for Assessing Early Plant Vigor in Winter Wheat. Sensors 2018, 18, 2931. [Google Scholar] [CrossRef]
- Bundessortenamt. Beschreibende Sortenliste Getreide, Mais, Öl- und Faserpflanzen, Leguminosen, Rüben, Zwischenfrüchte; Bundessortenamt: Hannover, Germany, 2016. [Google Scholar]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Meier, U. Entwicklungsstadien mono- und dikotyler Pflanzen. Biol. Bundesanst. L. Forstwirtsch. 2001, 2, 1–165. [Google Scholar]
- Prey, L.; Kipp, S.; Hu, Y.; Schmidhalter, U. Nitrogen Use Efficiency and Carbon Traits of High-Yielding European Hybrid vs. Line Winter Wheat Cultivars: Potentials and Limitations. Front. Plant Sci. 2019, 9, 1988. [Google Scholar] [CrossRef] [Green Version]
- Barmeier, G.; Mistele, B.; Schmidhalter, U. Referencing laser and ultrasonic height measurements of barley cultivars by using a herbometre as standard. Crop Pasture Sci. 2017, 67, 1215–1222. [Google Scholar] [CrossRef]
- Papakosta, D.K.; Gagianas, A.A. Nitrogen and Dry Matter Accumulation, Remobilization, and Losses for Mediterranean Wheat during Grain Filling. Agron. J. 1991, 83, 864–870. [Google Scholar] [CrossRef]
- Cox, M.C.; Qualset, C.O.; Rains, D.W. Genetic Variation for Nitrogen Assimilation and Translocation in Wheat. II. Nitrogen Assimilation in Relation to Grain Yield and Protein. Crop Sci. 1985, 25, 435–440. [Google Scholar] [CrossRef]
- Moll, R.H.; Kamprath, E.J.; Jackson, W.A. Analysis and Interpretation of Factors Which Contribute to Efficiency of Nitrogen Utilization1. Agron. J. 1982, 74, 562. [Google Scholar] [CrossRef]
- Brozyna, M.A.; Petersen, S.O.; Chirinda, N.; Olesen, J.E. Effects of grass-clover management and cover crops on nitrogen cycling and nitrous oxide emissions in a stockless organic crop rotation. Agric. Ecosyst. Environ. 2013, 181, 115–126. [Google Scholar] [CrossRef]
- McCartney, C.; Mercer, P.C.; Cooke, L.R.; Fraaije, B.A. Effects of a strobilurin-based spray programme on disease control, green leaf area, yield and development of fungicide-resistance in Mycosphaerella graminicola in Northern Ireland. Crop Prot. 2007, 26, 1272–1280. [Google Scholar] [CrossRef]
- Serrago, R.A.; Carretero, R.; Bancal, M.O.; Miralles, D.J. Foliar diseases affect the eco-physiological attributes linked with yield and biomass in wheat (Triticum aestivum L.). Eur. J. Agron. 2009, 31, 195–203. [Google Scholar] [CrossRef]
- Gebbing, T.; Schnyder, H.; Kühbauch, W. The utilization of pre-anthesis reserves in grain filling of wheat. Assessment by steady-state 13CO2/12CO2 labelling. Plant Cell Environ. 1999, 22, 851–858. [Google Scholar] [CrossRef]
- Ayres, P.G.; Press, M.C.; Spencer-Phillips, P.T.N. Effects of pathogens and parasitic plants on source-sink relationships. In Photoassimilate Distribution in Plants and Crops; Zamski, E., Schaffer, A.A., Eds.; Marcel Dekker: New York, NY, USA, 1996; Volume 8, pp. 479–499. [Google Scholar]
- Fagard, M.; Launay, A.; Clement, G.; Courtial, J.; Dellagi, A.; Farjad, M.; Krapp, A.; Soulie, M.-C.; Masclaux-Daubresse, C. Nitrogen metabolism meets phytopathology. J. Exp. Bot. 2014, 65, 5643–5656. [Google Scholar] [CrossRef] [PubMed]
- Dordas, C. Role of nutrients in controlling plant diseases in sustainable agriculture. A review. Agron. Sustain. Dev. 2008, 28, 33–46. [Google Scholar] [CrossRef] [Green Version]
- Robert, C.; Bancal, M.; Lannou, C. Wheat Leaf Rust Uredospore Production and Carbon and Nitrogen Export in Relation to Lesion Size and Density. Phytopathology 2002, 92, 762–768. [Google Scholar] [CrossRef] [Green Version]
- Howard, D.D.; Chambers, A.Y.; Logan, J. Nitrogen and fungicide effects on yield components and disease severity in wheat. J. Prod. Agric. 1994, 7, 448–454. [Google Scholar] [CrossRef]
- Leitch, M.H.; Jenkins, P.D. Influence of nitrogen on the development of Septoria epidemics in winter wheat. J. Agric. Sci. 1995, 124, 361–368. [Google Scholar] [CrossRef]
- Tapley, M.; Ortiz, B.V.; Van Santen, E.; Balkcom, K.S.; Mask, P.; Weaver, D.B. Location, Seeding Date, and Variety Interactions on Winter Wheat Yield in Southeastern United States. Agron. J. 2001, 105, 509–518. [Google Scholar] [CrossRef]
- Bodner, G.; Nakhforoosh, A.; Kaul, H.-P. Management of crop water under drought: A review. Agron. Sustain. Dev. 2015, 35, 401–442. [Google Scholar] [CrossRef]
- Barbottin, A.; Lecomte, C.; Bouchard, C.; Jeuffroy, M.-H. Nitrogen Remobilization during Grain Filling in Wheat. Crop Sci. 2005, 45, 1141. [Google Scholar] [CrossRef]
- Dimmock, J.; Gooding, M.J. The influence of foliar diseases, and their control by fungicides, on the protein concentration in wheat grain: A review. Wheat Prod. Stress. Environ. 2005, 138, 567–581. [Google Scholar] [CrossRef]
- Bancal, P. Decorrelating source and sink determinism of nitrogen remobilization during grain filling in wheat. Ann. Bot. 2009, 103, 1315–1324. [Google Scholar] [CrossRef] [Green Version]
- Winter, S.R.; Musick, J.T. Wheat planting date effects on soil water extraction and grain yield. Agron. J. 1993, 85, 912–916. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Year of Release | Cultivar Group | Spikes/m2 | GNS | Kernel Weight | Quality Class | Heading | Senesc-ence | Suscep. Septoria | Suscep. Yellow Rust | Suscep. Fusariosis |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Hybred | 2003 | hybrid | 4 | 8 | 5 | B | 5 | 6 | 6 | 4 | |
| Hyland | 2009 | hybrid | 4 | 9 | 4 | B | 4 | 5 | 3 | 2 | 4 |
| Hystar | 2007 | hybrid | 5 | 6 | 5 | B | - | 4 | - | - | - |
| Impression | 2005 | line | 7 | 4 | 5 | A | 5 | 6 | 5 | 3 | 3 |
| JB Asano | 2008 | line | 5 | 5 | 7 | A | 4 | 4 | 6 | 5 | 5 |
| Kerubino | 2004 | line | 7 | 4 | 5 | E | 4 | 5 | 4 | - | 4 |
| Trait Group | Plant Trait | Abbreviation | Description | Trait Complex |
|---|---|---|---|---|
| Dry matter traits [kg ha−1] | organ-specific dry matter (vegetative) | DM [organ-name] | at anthesis (Ant) and maturity (Mat) for flag leaf, flag leaf-1, ‘other leaves’, culms, and summed up for the three leaf layers, at anthesis for spikes and at maturity for chaff | |
| total dry matter | DMAnt, DMMat | sum of organ-specific DM-traits at anthesis and maturity | (d), (i) | |
| grain yield | GY (DM grain) | (a), (b), (c), (d) | ||
| N concentration traits [%] | organ-specific N concentration (vegetative) | NC [organ-name] | at anthesis and maturity for flag leaf, flag leaf-1, ‘other leaves’ and culms, at anthesis for spikes and at maturity for chaff | |
| grain N concentration | GNC | (a) | ||
| N uptake traits [kg ha−1] | organ-specific N uptake (vegetative) | Nup [organ-name] | at anthesis and maturity for flag leaf, flag leaf-1, ‘other leaves’, culms, and summed up for the three leaf layers at anthesis for spikes and at maturity for chaff | (j), (k) |
| total N uptake | NupAnt, NupMat | sum of organ-specific DM-traits at anthesis and maturity | (h), (k) | |
| N uptake straw | Nup straw | sum of organ-specific vegetative N uptake at maturity | ||
| grain N uptake | GNup | (a), (g), (h) | ||
| Derived DM traits | harvest index | HI | relative dry matter partitioning to the grain | (d) |
| organ-specific dry matter harvest index (vegetative) | HI [organ-name] | relative dry matter partitioning to vegetative organs at anthesis and maturity | ||
| post-anthesis assimilation | PAA | (c) | ||
| Table 2 (continued) | ||||
| Derived DM traits | total dry matter translocation efficiency | DMTEff | (i) | |
| organ-specific dry matter translocation efficiency | DMTEff [organ-name] | |||
| contribution of post-anthesis assimilates to grain-filling | CPAA | |||
| total dry matter translocation | DMT | (c) | ||
| organ-specific dry matter translocation | DMT [organ-name] | |||
| grain number per spike | GNS | (b) | ||
| thousand kernel weight | TKW | (b) | ||
| spike density | SD | (b) | ||
| N utilization efficiency for total dry matter at maturity | NutEff_total | |||
| N utilization efficiency for grain yield | NutEff_grain | |||
| N use efficiency for total dry matter at maturity | NUE_total_Mat | |||
| N use efficiency for grain yield | NUE_grain | |||
| kernels per m2 | - | |||
| yield per spike | - | |||
| Derived N traits | post-anthesis N uptake | PANup | (g) | |
| contribution of post-anthesis nitrogen to total nitrogen uptake | CPNup | |||
| total N translocation | NT | (g) | ||
| organ-specific N translocation (vegetative) | NT [organ-name] | |||
| N translocation efficiency | NTEff | (m) | ||
| organ-specific N translocation efficiency | NTEff [organ-name] | |||
| N harvest index | NHI | relative N partitioning to the grain | (h) | |
| organ-specific N harvest index (vegetative) | NHI [organ-name] | relative N partitioning to vegetative organs at anthesis and maturity | ||
| apparent N uptake efficiency at anthesis | NupEffAnt | |||
| apparent N uptake efficiency at maturity | NupEffMat | |||
| Other traits | plant height | - | ||
| anthesis date | - | |||
| Equation | Plant Trait and Description |
|---|---|
| Dry matter translocation (DMT): Absolute amount of translocated pre-anthesis accumulated assimilates from vegetative plant organs into grains between anthesis and maturity, in kg ha−1 [54] | |
| DMT-efficiency (DMTEff): Relative amount of translocated pre-anthesis accumulated assimilates into grains [54]. | |
| Post-anthesis assimilation (PAA) | |
| Contribution of post-anthesis assimilates to grain-filling (CPAA) | |
| Harvest index (HI): Ratio of grain DM to total DM at maturity | |
| Absolute amount of translocated pre-anthesis accumulated nitrogen from vegetative plant organs into grains between anthesis and maturity in kg ha−1 (NT) [55] | |
| NT-efficiency (NTEff): Relative amount of translocated pre-anthesis accumulated nitrogen into grains [55] | |
| Post-anthesis nitrogen uptake (PANup) | |
| Contribution of post-anthesis nitrogen to total nitrogen uptake (CPNup) | |
| N-Harvest index (NHI): Ratio of grain nitrogen uptake (Nup) to total Nup at maturity | |
| Apparent nitrogen uptake efficiency, calculated for anthesis and maturity as ratio of total Nup to total N fertilized [56] | |
| Nitrogen utilization efficiency (NutEff): Efficiency of the internal conversion of N into grain and total DM [56]. | |
| (1) |
| Whole Trial | Cont + RF | Cont + SD1 | Whole Trial | Cont + RF | Cont + SD1 | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MP | N | Cv | MP | N | Cv | MP | N | Cv | MP x N | MP x Cv | N x Cv | MP x | MP x Cv | N x Cv | MP x N | MP x Cv | N x Cv | ||
| DM traits [kg ha−1] | Ant. spikes | *** | *** | *** | * | * | *** | * | |||||||||||
| Ant. culms | * | *** | * | *** | * | *** | * | ** | * | *** | * | ||||||||
| Ant. flag leaf | *** | *** | ** | *** | *** | *** | *** | *** | |||||||||||
| Ant. flag leaf-1 | *** | *** | * | *** | * | *** | |||||||||||||
| Ant. other leaves | *** | *** | * | *** | ** | *** | ** | ** | ** | *** | * | ||||||||
| Ant. leaves | *** | *** | * | *** | * | *** | |||||||||||||
| Mat. grain (GY) | *** | *** | *** | * | *** | *** | *** | *** | |||||||||||
| Mat. chaff | ** | ** | ** | * | ** | * | |||||||||||||
| Mat. culms | * | * | *** | *** | ** | *** | *** | * | |||||||||||
| Mat. flag leaf | *** | *** | ** | *** | *** | *** | ** | ||||||||||||
| Mat. flag leaf-1 | * | *** | *** | * | *** | * | |||||||||||||
| Mat. other leaves | * | *** | *** | * | *** | * | |||||||||||||
| Mat. leaves | * | *** | *** | ** | *** | * | |||||||||||||
| Ant. total | *** | *** | *** | * | * | * | *** | * | |||||||||||
| Mat. total | * | *** | * | *** | *** | ** | |||||||||||||
| NC traits [%] | Ant. spikes | * | *** | *** | *** | *** | * | *** | ** | *** | *** | *** | *** | *** | |||||
| Ant. culms | *** | *** | *** | *** | *** | ** | * | ||||||||||||
| Ant. flag leaf | *** | *** | *** | *** | *** | *** | * | *** | * | ** | ** | ||||||||
| Ant. flag leaf-1 | *** | *** | *** | *** | *** | *** | *** | *** | * | ** | |||||||||
| Ant. other leaves | ** | *** | *** | ** | ** | *** | *** | *** | * | * | |||||||||
| Mat. grain (GNC) | ** | *** | *** | * | *** | *** | *** | *** | * | * | |||||||||
| Mat. chaff | *** | *** | *** | *** | *** | *** | *** | *** | |||||||||||
| Mat. culms | *** | *** | ** | *** | *** | ** | *** | *** | * | * | * | ||||||||
| Mat. flag leaf | *** | *** | *** | *** | *** | *** | *** | *** | ** | *** | ** | *** | * | ||||||
| Mat. flag leaf-1 | *** | *** | *** | *** | *** | * | * | ||||||||||||
| Mat. other leaves | *** | *** | *** | *** | *** | *** | *** | *** | *** | ** | *** | ||||||||
| Nup traits [kg ha−1] | Ant. spikes | *** | ** | ** | *** | ** | * | * | *** | ||||||||||
| Ant. culms | *** | * | *** | *** | *** | ** | |||||||||||||
| Ant. flag leaf | *** | *** | *** | *** | *** | *** | ** | *** | |||||||||||
| Ant. flag leaf-1 | * | *** | *** | *** | *** | *** | *** | * | * | *** | |||||||||
| Ant. other leaves | *** | *** | ** | *** | * | ** | *** | ** | * | *** | * | * | *** | * | |||||
| Ant. leaves | *** | *** | ** | *** | *** | *** | * | *** | |||||||||||
| Mat. grain (GNup) | * | *** | *** | *** | ** | * | |||||||||||||
| Mat. chaff | *** | *** | * | *** | *** | *** | *** | ** | |||||||||||
| Mat. culms | *** | *** | *** | *** | *** | ** | *** | *** | |||||||||||
| Mat. flag leaf | *** | *** | *** | *** | *** | *** | *** | *** | ** | *** | ** | *** | * | ||||||
| Mat. flag leaf-1 | *** | *** | *** | *** | *** | *** | *** | * | |||||||||||
| Mat. other leaves | *** | *** | *** | *** | ** | *** | ** | *** | *** | * | |||||||||
| Table 4 (continued) | |||||||||||||||||||
| Nup traits [kg ha−1] | Mat. leaves | *** | *** | *** | *** | *** | *** | *** | *** | *** | ** | ||||||||
| Ant. total | *** | *** | ** | ** | *** | *** | * | *** | |||||||||||
| Mat. total | *** | ** | *** | *** | * | * | |||||||||||||
| Mat. straw | *** | *** | *** | *** | *** | *** | *** | *** | |||||||||||
| Derived DM traits | HI | *** | *** | *** | *** | *** | * | ** | |||||||||||
| PAA | *** | *** | *** | *** | *** | *** | ** | * | *** | ||||||||||
| DMTEff | *** | * | *** | ** | ** | ** | |||||||||||||
| CPAA | *** | *** | *** | *** | *** | *** | * | * | *** | ||||||||||
| DMT | ** | *** | ** | *** | * | ** | * | *** | |||||||||||
| GNS | *** | * | *** | ** | * | *** | *** | *** | *** | *** | |||||||||
| TKW | *** | *** | *** | *** | *** | * | |||||||||||||
| spike density | * | *** | * | *** | * | *** | * | ||||||||||||
| NutEff_total | * | *** | ** | *** | *** | * | * | * | |||||||||||
| NutEff_grain | *** | *** | *** | *** | *** | *** | *** | *** | * | ||||||||||
| NUE_total | * | *** | *** | *** | ** | ||||||||||||||
| kernels per m2 | *** | *** | *** | *** | *** | *** | ** | * | |||||||||||
| yield per spike | *** | * | *** | *** | *** | ** | *** | *** | *** | ||||||||||
| NUE_grain | *** | *** | *** | * | *** | *** | *** | *** | ** | * | |||||||||
| derived N traits | PANup | *** | * | *** | *** | *** | * | ** | * | * | *** | ||||||||
| CPNup | *** | ** | *** | *** | *** | ** | ** | * | * | *** | |||||||||
| NT | ** | ** | ** | *** | *** | * | * | * | *** | ||||||||||
| NTEff | *** | *** | *** | *** | *** | *** | ** | * | * | ||||||||||
| NHI | *** | *** | *** | *** | *** | ||||||||||||||
| NupEff Ant. | *** | *** | *** | ** | *** | *** | *** | ||||||||||||
| NupEff Mat. | *** | * | *** | *** | * | ||||||||||||||
| other traits | plant height | *** | *** | ** | *** | *** | *** | *** | * | * | |||||||||
| anthesis date | *** | *** | - | *** | - | *** | *** | - | *** | - | - | - | - | *** | - | - | |||
| Treatment | DM Mat. Grain (GY) | DM Ant. total | DM Mat. total | HI | PAA | DMTEff | DMT | GNS | TKW | Spike Density | NutEff Total | NutEff Grain | NUE Mat. Total | NUE Mat. Grain | NC Mat. Grain (GNC) | Nup Ant. Spikes | Nup Ant. Culms | Nup Ant. Flag Leaf | Nup Ant. Flag Leaf-1 | Nup Ant. Other Leaves | Nup Mat. Grain (GNup) | Nup Mat. Chaff | Nup Mat. Culms | Nup Mat. Flag Leaf | Nup Mat. Flag Leaf-1 | Nup Mat. Other Leaves | Nup Ant. Total | Nup Mat. Total | PANup | NT | NTEff | NHI | NupEff Mat. | Plant Height | Anthesis (Days in June) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cont | 9189b | 13091a | 17340ab | 0.53b | 4267ab | 0.37a | 4930a | 36.4a | 38.6b | 663ab | 83.94ab | 44.49b | 214a | 114b | 1.9a | 44a | 70a | 36a | 28ab | 18b | 176ab | 9a | 15a | 3a | 2a | 3a | 196a | 209a | 13a | 163a | 0.83b | 0.84b | 2.52a | 0.65a | 9.5b |
| RF | 8107a | 13477a | 16884a | 0.48a | 3407a | 0.34a | 4717a | 35.1a | 33.6a | 697b | 82.09a | 39.43a | 211a | 101a | 2b | 43a | 76a | 37a | 29b | 15a | 161a | 10a | 20b | 8b | 4b | 4b | 201a | 207a | 6a | 155a | 0.77a | 0.78a | 2.52a | 0.69b | 9.5b |
| SD1 | 9895b | 13956a | 18706b | 0.53b | 4712b | 0.37a | 5154a | 40.2b | 40b | 629a | 86.56b | 45.87b | 231b | 122c | 1.9a | 43a | 73a | 34a | 26a | 21c | 185b | 10a | 15a | 3a | 2a | 4b | 197a | 219a | 21a | 163a | 0.83b | 0.85b | 2.63a | 0.67ab | 6.7a |
| Cont->RF | −12% | 3% | −3% | −9% | −20% | −8% | −4% | −4% | −13% | 5% | −2% | −11% | −2% | −11% | 4% | −2% | 9% | 5% | 3% | −14% | −9% | 11% | 35% | 127% | 71% | 39% | 3% | −1% | −5% | −8% | −8% | 0% | 6% | ||
| Cont->SD1 | 8% | 7% | 8% | 0% | 10% | −2% | 5% | 10% | 4% | −5% | 3% | 3% | 8% | 8% | −2% | −4% | 5% | −4% | −6% | 16% | 5% | 2% | 1% | −8% | −2% | 29% | 1% | 5% | 0% | 0% | 0% | 5% | 3% | ||
| N1 | 8787a | 13658a | 17178a | 0.51a | 3527a | 0.38b | 5264b | 36.8a | 37.3a | 653a | 90.33b | 46.27b | 286b | 146b | 1.8a | 43a | 68a | 34a | 27a | 18a | 156a | 9a | 15a | 4a | 3a | 4a | 189a | 191a | 2a | 154a | 0.82b | 0.82a | 3.18b | 0.65a | 8.3a |
| N2 | 9340b | 13358a | 18108b | 0.52a | 4730b | 0.34a | 4603a | 37.7a | 37.6a | 673a | 78.06a | 40.25a | 151a | 78a | 2.1b | 44a | 78b | 38b | 29b | 18a | 191b | 10b | 18b | 5b | 3b | 4b | 207b | 232b | 25b | 166b | 0.8a | 0.82a | 1.94a | 0.69b | 8.9b |
| cv. Hyland | 9461c | 13546b | 17982a | 0.53cd | 4450b | 0.37ab | 5024abc | 42.2c | 35.1a | 645b | 85.74b | 45.18b | 222a | 117c | 1.9ab | 43ab | 74ab | 36bc | 29bc | 19c | 176a | 9a | 17ab | 4a | 3a | 4ab | 201ab | 212ab | 11ab | 165b | 0.82c | 0.83c | 2.55ab | 0.68bc | 8.7c |
| cv. Hybred | 9217bc | 14281b | 18125a | 0.51ab | 3827ab | 0.37ab | 5379bc | 40.7c | 35.9ab | 639b | 83.58ab | 42.46a | 224a | 114bc | 1.9bc | 48c | 72ab | 40c | 31c | 20c | 176a | 11d | 17ab | 7b | 3b | 4c | 210b | 219b | 9ab | 167b | 0.8ab | 0.81a | 2.64b | 0.66b | 10.5f |
| cv. Hystar | 9228bc | 11989a | 17049a | 0.54d | 4987b | 0.35ab | 4182a | 37b | 37.4bc | 678bc | 86.25b | 46.7b | 212a | 114bc | 1.8a | 39a | 65a | 35b | 22a | 15ab | 165a | 9ab | 15a | 4a | 3ab | 3a | 177a | 199a | 22b | 142a | 0.81abc | 0.83bc | 2.41a | 0.6a | 4.3a |
| cv. Kerubino | 8634a | 14380b | 17362a | 0.5a | 2984a | 0.39b | 5652c | 32.1a | 37.9c | 715c | 82.2a | 40.85a | 216a | 107a | 2d | 43ab | 76b | 37bc | 30bc | 23d | 174a | 9abc | 18b | 4a | 3a | 4bc | 209b | 212ab | 3a | 170b | 0.81bc | 0.82ab | 2.59ab | 0.69bcd | 8.5b |
| cv. Impression | 8939ab | 13572b | 17994a | 0.5a | 4457b | 0.33a | 4517ab | 31.8a | 38.7cd | 728c | 84.34ab | 41.8a | 223a | 111ab | 1.9cd | 43abc | 78b | 38bc | 28b | 13a | 175a | 10bc | 18b | 6b | 3ab | 4bc | 200ab | 216b | 16ab | 159ab | 0.79a | 0.81a | 2.61ab | 0.69cd | 10.3e |
| cv. JB Asano | 8902ab | 13279ab | 17350a | 0.51bc | 4066ab | 0.36ab | 4848abc | 39.7bc | 39.7d | 571a | 83.06ab | 42.58a | 215a | 110ab | 2cd | 45bc | 72ab | 29a | 28b | 16b | 176a | 10cd | 15a | 4a | 3a | 4abc | 191ab | 211ab | 20ab | 156ab | 0.82bc | 0.83c | 2.54ab | 0.71d | 9d |
| Cont N1 | 8872a | 13198a | 16760a | 0.53a | 3585a | 0.4b | 5304b | 36.2a | 38.5a | 645a | 89.64b | 47.47b | 280b | 148b | 1.8a | 43a | 64a | 33a | 27a | 17a | 158a | 9a | 13a | 3a | 2a | 3a | 185a | 188a | 3a | 155a | 0.84b | 0.84a | 3.13b | 0.64a | - |
| Cont N2 | 9505b | 12984a | 17921b | 0.53a | 4948b | 0.35a | 4557a | 36.7a | 38.7a | 680a | 78.24a | 41.5a | 149a | 79a | 2b | 45a | 75b | 38b | 30b | 18a | 194b | 10b | 16b | 4b | 3b | 3b | 206b | 230b | 23b | 170b | 0.82a | 0.84a | 1.91a | 0.67b | - |
| RF N1 | 7949a | 13942b | 16683a | 0.48a | 2739a | 0.37b | 5219b | 34a | 34a | 696a | 87.63b | 41.79b | 278b | 133b | 1.9a | 44a | 73a | 36a | 28a | 16a | 148a | 10a | 18a | 7a | 4a | 4a | 198a | 191a | -7a | 156a | 0.78b | 0.78a | 3.18b | 0.68a | - |
| RF N2 | 8264b | 13011a | 17086a | 0.48a | 4074b | 0.31a | 4215a | 36.1b | 33.3a | 698a | 76.55a | 37.07a | 143a | 69a | 2.1b | 43a | 78a | 39a | 30a | 15a | 173b | 11b | 22b | 9b | 4b | 4a | 204a | 224b | 20b | 154a | 0.75a | 0.78a | 1.87a | 0.7a | - |
| SD1 N1 | 9539a | 13833a | 18093a | 0.53a | 4256a | 0.38a | 5270a | 40.1a | 39.4a | 617a | 93.72b | 49.57b | 301b | 159b | 1.7a | 41a | 66a | 32a | 24a | 20a | 163a | 9a | 13a | 3a | 2a | 4a | 183a | 194a | 10a | 153a | 0.83a | 0.84a | 3.23b | 0.64a | - |
| SD1 N2 | 10250b | 14079a | 19319b | 0.53a | 5168b | 0.35a | 5037a | 40.3a | 40.6b | 641a | 79.39a | 42.18a | 162a | 86a | 2b | 44a | 80b | 37b | 29b | 22b | 206b | 10b | 17b | 4b | 3b | 4b | 210b | 244b | 32b | 173b | 0.82a | 0.85a | 2.04a | 0.69b | - |
| Cont: N1->N2 | 7% | −2% | 7% | 0% | 38% | −13% | −14% | 2% | 0% | 5% | −13% | −13% | −47% | −47% | 14% | 5% | 16% | 15% | 12% | 7% | 23% | 13% | 22% | 24% | 29% | 22% | 12% | 22% | 10% | −2% | 0% | −39% | 5% | ||
| RF: N1->N2 | 4% | −7% | 2% | 2% | 49% | −15% | −19% | 6% | −2% | 0% | −13% | −11% | −48% | −48% | 13% | −4% | 7% | 8% | 4% | −8% | 17% | 16% | 17% | 34% | 19% | 8% | 3% | 17% | −1% | −4% | 0% | −41% | 3% | ||
| SD1: N1->N2 | 7% | 2% | 7% | 1% | 21% | −7% | −4% | 0% | 3% | 4% | −15% | −15% | −46% | −46% | 18% | 6% | 20% | 17% | 17% | 10% | 26% | 12% | 25% | 41% | 28% | 18% | 15% | 26% | 13% | −1% | 1% | −37% | 8% |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prey, L.; Germer, M.; Schmidhalter, U. Temporal and Organ-specific Responses in NUE Traits to N Fertilization, Fungicide Intensity and Early Sowing in Winter Wheat Cultivars. Agronomy 2019, 9, 313. https://doi.org/10.3390/agronomy9060313
Prey L, Germer M, Schmidhalter U. Temporal and Organ-specific Responses in NUE Traits to N Fertilization, Fungicide Intensity and Early Sowing in Winter Wheat Cultivars. Agronomy. 2019; 9(6):313. https://doi.org/10.3390/agronomy9060313
Chicago/Turabian StylePrey, Lukas, Moritz Germer, and Urs Schmidhalter. 2019. "Temporal and Organ-specific Responses in NUE Traits to N Fertilization, Fungicide Intensity and Early Sowing in Winter Wheat Cultivars" Agronomy 9, no. 6: 313. https://doi.org/10.3390/agronomy9060313
APA StylePrey, L., Germer, M., & Schmidhalter, U. (2019). Temporal and Organ-specific Responses in NUE Traits to N Fertilization, Fungicide Intensity and Early Sowing in Winter Wheat Cultivars. Agronomy, 9(6), 313. https://doi.org/10.3390/agronomy9060313