
The Function of Immunoproteasomes—An Immunologists’ Perspective

 , and
, and {kind=link}
{kind=link}
{kind=link}
Abstract
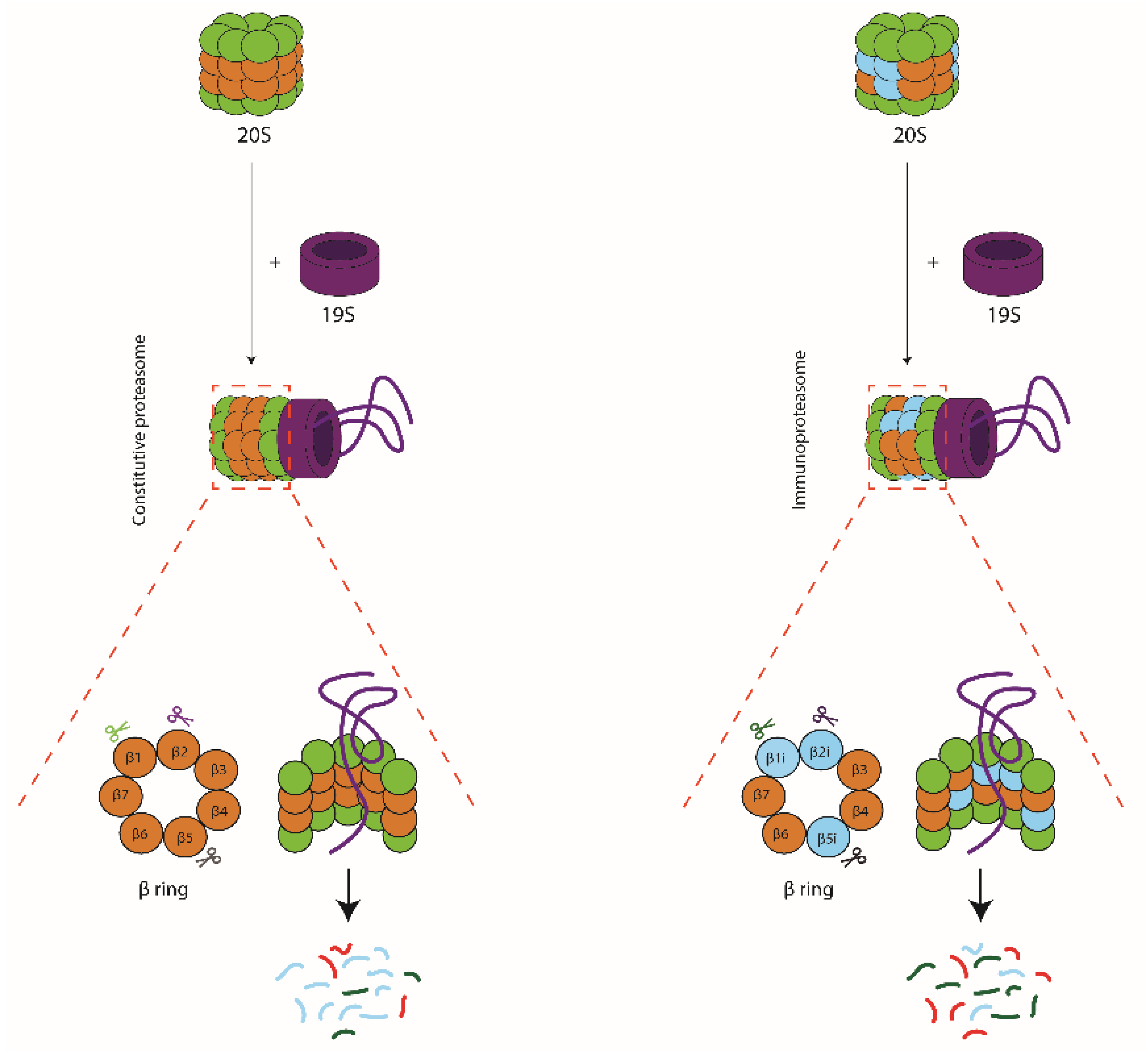
:1. Introduction
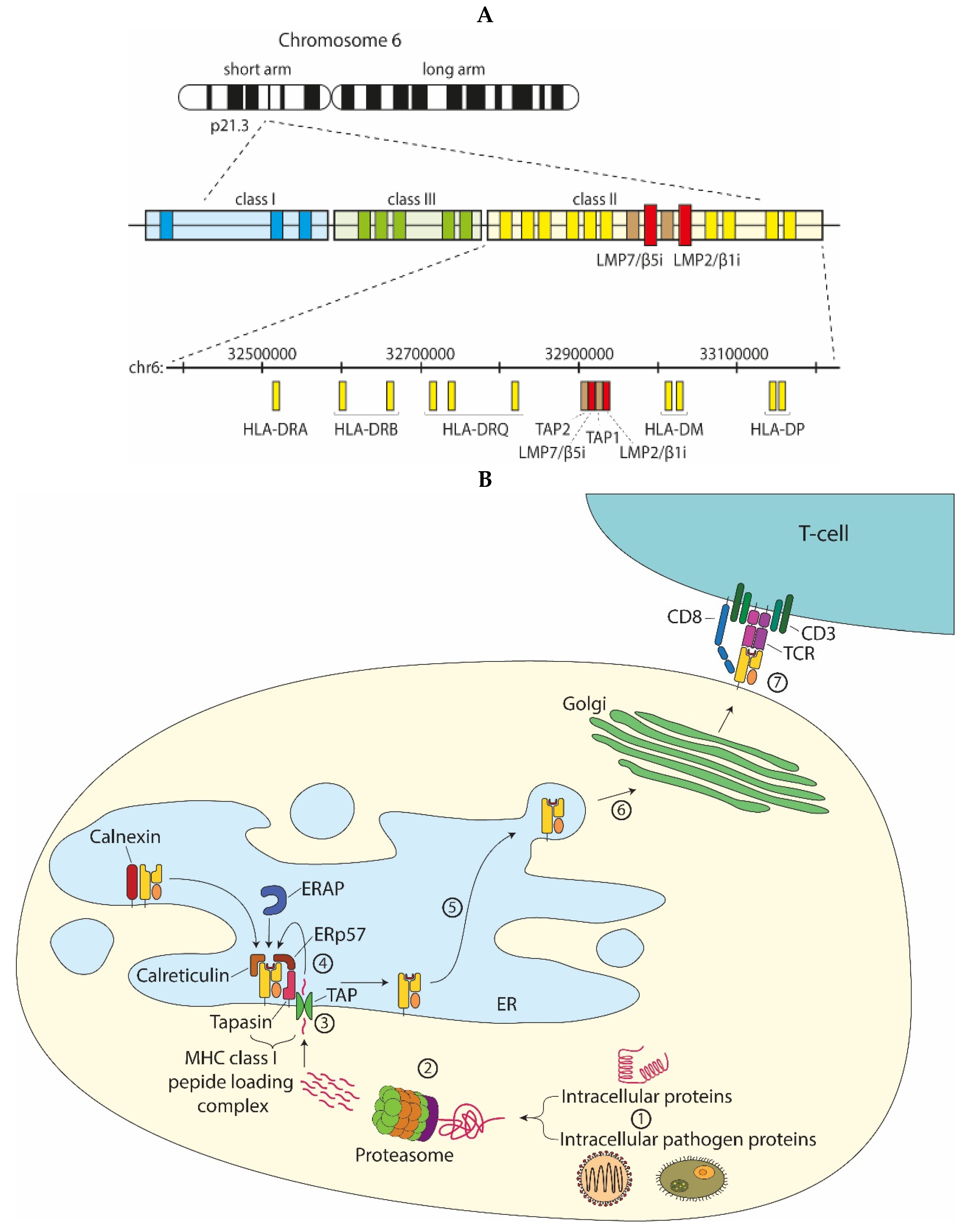
2. Role of Immunoproteasomes in MHC Class I Antigen Processing and CD8+ T-Cell Responses
2.1. Role of the Inducible Proteasome Subunits Based on Expression Patterns
2.2. In Vitro Digestion Analyses to Reveal Immunoproteasome Function
2.3. Immune Responses in the Absence of Immunoproteasome Subunits
2.4. Role of Immunoproteasomes in Immune Protection
2.5. Exploiting Immunoproteasomes to Control Infections
3. Proteasome Subunit Composition and T-Cell Selection
4. Role of Immunoproteasomes in Auto-Immune Responses
4.1. CD8 T Cell-Mediated Early Stage, Multi-Tissue Autoimmune Disease in Immunoproteasome Subunit-Deficient Mice
4.2. Genetic Linkage of Immunoproteasome Subunits with Different Autoinflammatory and Autoimmune Diseases
4.3. Targeting Immunoproteasomes to Dampen Auto-Immune and Inflammatory Disease
5. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hershko, A.; Ciechanover, A. The Ubiquitin System. Annu. Rev. Biochem. 1998, 67, 425–479. [Google Scholar] [CrossRef]
- Wilk, S.; Orlowski, M. Evidence That Pituitary Cation-Sensitive Neutral Endopeptidase Is a Multicatalytic Protease Complex. J. Neurochem. 1983, 40, 842–849. [Google Scholar] [CrossRef] [PubMed]
- Falkenburg, P.E.; Haass, C.; Kloetzel, P.M.; Niedel, B.; Kopp, F.; Kuehn, L.; Dahlmann, B. Drosophila Small Cytoplasmic 19S Ribonucleoprotein Is Homologous to the Rat Multicatalytic Proteinase. Nature 1988, 331, 190–192. [Google Scholar] [CrossRef] [PubMed]
- Arrigo, A.P.; Tanaka, K.; Goldberg, A.L.; Welch, W.J. Identity of the 19S “prosome” Particle with the Large Multifunctional Protease Complex of Mammalian Cells (the Proteasome). Nature 1988, 331, 192–194. [Google Scholar] [CrossRef]
- Kasahara, M.; Flajnik, M.F. Origin and Evolution of the Specialized Forms of Proteasomes Involved in Antigen Presentation. Immunogenetics 2019, 71, 251–261. [Google Scholar] [CrossRef]
- Majumder, P.; Baumeister, W. Proteasomes: Unfoldase-Assisted Protein Degradation Machines. Biol. Chem. 2019, 401, 183–199. [Google Scholar] [CrossRef]
- Bard, J.A.M.; Goodall, E.A.; Greene, E.R.; Jonsson, E.; Dong, K.C.; Martin, A. Structure and Function of the 26S Proteasome. Annu. Rev. Biochem. 2018, 87, 697–724. [Google Scholar] [CrossRef]
- Schweitzer, A.; Aufderheide, A.; Rudack, T.; Beck, F.; Pfeider, G.; Plitzko, J.M.; Sakata, E.; Schuiten, K.; Forster, F.; Baumeister, W. Structure of the human 26S proteasome at a resolution of 3.9 angstrom. Proc. Natl. Acad. Sci. USA 2016, 113, 7816–7821. [Google Scholar] [CrossRef] [Green Version]
- Parham, P. Antigen Processing. Transporters of Delight. Nature 1990, 348, 674–675. [Google Scholar] [CrossRef]
- Robertson, M. Proteasomes in the Pathway. Nature 1991, 353, 300–301. [Google Scholar] [CrossRef]
- Brown, M.G.; Driscoll, J.; Monaco, J.J. Structural and Serological Similarity of MHC-Linked LMP and Proteasome (Multicatalytic Proteinase) Complexes. Nature 1991, 353, 355–357. [Google Scholar] [CrossRef] [PubMed]
- Glynne, R.; Powis, S.H.; Beck, S.; Kelly, A.; Kerr, L.A.; Trowsdale, J. A Proteasome-Related Gene between the Two ABC Transporter Loci in the Class II Region of the Human MHC. Nature 1991, 353, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Navarrete, V.; Seelig, A.; Gernold, M.; Frentzel, S.; Kloetzel, P.M.; Hämmerling, G.J. Subunit of the “20S” Proteasome (Multicatalytic Proteinase) Encoded by the Major Histocompatibility Complex. Nature 1991, 353, 662–664. [Google Scholar] [CrossRef] [PubMed]
- Martinez, C.K.; Monaco, J.J. Homology of Proteasome Subunits to a Major Histocompatibility Complex-Linked LMP Gene. Nature 1991, 353, 664–667. [Google Scholar] [CrossRef]
- Nandi, D.; Jiang, H.; Monaco, J.J. Identification of MECL-1 (LMP-10) as the Third IFN-Gamma-Inducible Proteasome Subunit. J. Immunol. 1996, 156, 2361–2364. [Google Scholar] [PubMed]
- Groettrup, M.; Kraft, R.; Kostka, S.; Standera, S.; Stohwasser, R.; Kloetzel, P.M. A Third Interferon-Gamma-Induced Subunit Exchange in the 20S Proteasome. Eur. J. Immunol. 1996, 26, 863–869. [Google Scholar] [CrossRef]
- Groettrup, M.; Soza, A.; Kuckelkorn, U.; Kloetzel, P.-M. Peptide Antigen Production by the Proteasome: Complexity Provides Efficiency. Immunol. Today 1996, 17, 429–435. [Google Scholar] [CrossRef]
- Rock, K.L.; Gramm, C.; Rothstein, L.; Clark, K.; Stein, R.; Dick, L.; Hwang, D.; Goldberg, A.L. Inhibitors of the Proteasome Block the Degradation of Most Cell Proteins and the Generation of Peptides Presented on MHC Class I Molecules. Cell 1994, 78, 761–771. [Google Scholar] [CrossRef]
- Harding, C.V.; France, J.; Song, R.; Farah, J.M.; Chatterjee, S.; Iqbal, M.; Siman, R. Novel Dipeptide Aldehydes Are Proteasome Inhibitors and Block the MHC-I Antigen-Processing Pathway. J. Immunol. 1995, 155, 1767–1775. [Google Scholar]
- Sijts, A.J.; Standera, S.; Toes, R.E.; Ruppert, T.; Beekman, N.J.; van Veelen, P.A.; Ossendorp, F.A.; Melief, C.J.; Kloetzel, P.M. MHC Class I Antigen Processing of an Adenovirus CTL Epitope Is Linked to the Levels of Immunoproteasomes in Infected Cells. J. Immunol. 2000, 164, 4500–4506. [Google Scholar] [CrossRef] [Green Version]
- Dick, L.R.; Aldrich, C.; Jameson, S.C.; Moomaw, C.R.; Pramanik, B.C.; Doyle, C.K.; DeMartino, G.N.; Bevan, M.J.; Forman, J.M.; Slaughter, C.A. Proteolytic Processing of Ovalbumin and Beta-Galactosidase by the Proteasome to a Yield Antigenic Peptides. J. Immunol. 1994, 152, 3884–3894. [Google Scholar] [PubMed]
- Boes, B.; Hengel, H.; Ruppert, T.; Multhaup, G.; Koszinowski, U.H.; Kloetzel, P.M. Interferon Gamma Stimulation Modulates the Proteolytic Activity and Cleavage Site Preference of 20S Mouse Proteasomes. J. Exp. Med. 1994, 179, 901–909. [Google Scholar] [CrossRef] [Green Version]
- Driscoll, J.; Brown, M.G.; Finley, D.; Monaco, J.J. MHC-Linked LMP Gene Products Specifically Alter Peptidase Activities of the Proteasome. Nature 1993, 365, 262–264. [Google Scholar] [CrossRef] [PubMed]
- Fehling, H.J.; Swat, W.; Laplace, C.; Kühn, R.; Rajewsky, K.; Müller, U.; von Boehmer, H. MHC Class I Expression in Mice Lacking the Proteasome Subunit LMP-7. Science 1994, 265, 1234–1237. [Google Scholar] [CrossRef]
- Van Kaer, L.; Ashton-Rickardt, P.G.; Eichelberger, M.; Gaczynska, M.; Nagashima, K.; Rock, K.L.; Goldberg, A.L.; Doherty, P.C.; Tonegawa, S. Altered Peptidase and Viral-Specific T Cell Response in LMP2 Mutant Mice. Immunity 1994, 1, 533–541. [Google Scholar] [CrossRef]
- Arnold, D.; Driscoll, J.; Androlewicz, M.; Hughes, E.; Cresswell, P.; Spies, T. Proteasome Subunits Encoded in the MHC Are Not Generally Required for the Processing of Peptides Bound by MHC Class I Molecules. Nature 1992, 360, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Yewdell, J.; Lapham, C.; Bacik, I.; Spies, T.; Bennink, J. MHC-Encoded Proteasome Subunits LMP2 and LMP7 Are Not Required for Efficient Antigen Presentation. J. Immunol. 1994, 152, 1163–1170. [Google Scholar] [PubMed]
- Stohwasser, R.; Standera, S.; Peters, I.; Kloetzel, P.M.; Groettrup, M. Molecular Cloning of the Mouse Proteasome Subunits MC14 and MECL-1: Reciprocally Regulated Tissue Expression of Interferon-Gamma-Modulated Proteasome Subunits. Eur. J. Immunol. 1997, 27, 1182–1187. [Google Scholar] [CrossRef]
- Macagno, A.; Kuehn, L.; de Giuli, R.; Groettrup, M. Pronounced Up-Regulation of the PA28alpha/Beta Proteasome Regulator but Little Increase in the Steady-State Content of Immunoproteasome during Dendritic Cell Maturation. Eur. J. Immunol. 2001, 31, 3271–3280. [Google Scholar] [CrossRef]
- Li, J.; Schuler-Thurner, B.; Schuler, G.; Huber, C.; Seliger, B. Bipartite Regulation of Different Components of the MHC Class I Antigen-Processing Machinery during Dendritic Cell Maturation. Int. Immunol. 2001, 13, 1515–1523. [Google Scholar] [CrossRef] [Green Version]
- Hallermalm, K.; Seki, K.; Wei, C.; Castelli, C.; Rivoltini, L.; Kiessling, R.; Levitskaya, J. Tumor Necrosis Factor-α Induces Coordinated Changes in Major Histocompatibility Class I Presentation Pathway, Resulting in Increased Stability of Class I Complexes at the Cell Surface. Blood 2001, 98, 1108–1115. [Google Scholar] [CrossRef]
- Shin, E.-C.; Seifert, U.; Kato, T.; Rice, C.M.; Feinstone, S.M.; Kloetzel, P.-M.; Rehermann, B. Virus-Induced Type I IFN Stimulates Generation of Immunoproteasomes at the Site of Infection. J. Clin. Investig. 2006, 116, 3006–3014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wieland, S.F.; Vega, R.G.; Müller, R.; Evans, C.F.; Hilbush, B.; Guidotti, L.G.; Sutcliffe, J.G.; Schultz, P.G.; Chisari, F.V. Searching for Interferon-Induced Genes That Inhibit Hepatitis B Virus Replication in Transgenic Mouse Hepatocytes. J. Virol. 2003, 77, 1227–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treise, I.; Huber, E.M.; Klein-Rodewald, T.; Heinemeyer, W.; Grassmann, S.A.; Basler, M.; Adler, T.; Rathkolb, B.; Helming, L.; Andres, C.; et al. Defective Immuno- and Thymoproteasome Assembly Causes Severe Immunodeficiency. Sci. Rep. 2018, 8, 5975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boegel, S.; Löwer, M.; Bukur, T.; Sorn, P.; Castle, J.C.; Sahin, U. HLA and Proteasome Expression Body Map. BMC Med. Genom. 2018, 11, 36. [Google Scholar] [CrossRef] [PubMed]
- Sijts, A.J.; Ruppert, T.; Rehermann, B.; Schmidt, M.; Koszinowski, U.; Kloetzel, P.M. Efficient Generation of a Hepatitis B Virus Cytotoxic T Lymphocyte Epitope Requires the Structural Features of Immunoproteasomes. J. Exp. Med. 2000, 191, 503–514. [Google Scholar] [CrossRef] [Green Version]
- Basler, M.; Youhnovski, N.; Van Den Broek, M.; Przybylski, M.; Groettrup, M. Immunoproteasomes Down-Regulate Presentation of a Subdominant T Cell Epitope from Lymphocytic Choriomeningitis Virus. J. Immunol. 2004, 173, 3925–3934. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, K.; van Den Broek, M.; Kostka, S.; Kraft, R.; Soza, A.; Schmidtke, G.; Kloetzel, P.M.; Groettrup, M. Overexpression of the Proteasome Subunits LMP2, LMP7, and MECL-1, but Not PA28 Alpha/Beta, Enhances the Presentation of an Immunodominant Lymphocytic Choriomeningitis Virus T Cell Epitope. J. Immunol. 2000, 165, 768–778. [Google Scholar] [CrossRef] [Green Version]
- Toes, R.E.; Nussbaum, A.K.; Degermann, S.; Schirle, M.; Emmerich, N.P.; Kraft, M.; Laplace, C.; Zwinderman, A.; Dick, T.P.; Müller, J.; et al. Discrete Cleavage Motifs of Constitutive and Immunoproteasomes Revealed by Quantitative Analysis of Cleavage Products. J. Exp. Med. 2001, 194, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Mishto, M.; Liepe, J.; Textoris-Taube, K.; Keller, C.; Henklein, P.; Weberruß, M.; Dahlmann, B.; Enenkel, C.; Voigt, A.; Kuckelkorn, U.; et al. Proteasome Isoforms Exhibit Only Quantitative Differences in Cleavage and Epitope Generation. Eur. J. Immunol. 2014, 44, 3508–3521. [Google Scholar] [CrossRef]
- Winter, M.B.; La Greca, F.; Arastu-Kapur, S.; Caiazza, F.; Cimermancic, P.; Buchholz, T.J.; Anderl, J.L.; Ravalin, M.; Bohn, M.F.; Sali, A.; et al. Immunoproteasome functions explained by divergence in cleavage specificity and regulation. Elife 2017, 6, e27364. [Google Scholar] [CrossRef]
- Basler, M.; Moebius, J.; Elenich, L.; Groettrup, M.; Monaco, J.J. An Altered T Cell Repertoire in MECL-1-Deficient Mice. J. Immunol. 2006, 176, 6665–6672. [Google Scholar] [CrossRef] [Green Version]
- Caudill, C.M.; Jayarapu, K.; Elenich, L.; Monaco, J.J.; Colbert, R.A.; Griffin, T.A. T Cells Lacking Immunoproteasome Subunits MECL-1 and LMP7 Hyperproliferate in Response to Polyclonal Mitogens. J. Immunol. 2006, 176, 4075–4082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kincaid, E.Z.; Che, J.W.; York, I.; Escobar, H.; Reyes-Vargas, E.; Delgado, J.C.; Welsh, R.M.; Karow, M.L.; Murphy, A.J.; Valenzuela, D.M.; et al. Mice Completely Lacking Immunoproteasomes Show Major Changes in Antigen Presentation. Nat. Immunol. 2011, 13, 129–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaiss, D.M.W.; de Graaf, N.; Sijts, A.J.A.M. The Proteasome Immunosubunit Multicatalytic Endopeptidase Complex-like 1 Is a T-Cell-Intrinsic Factor Influencing Homeostatic Expansion. Infect Immun. 2008, 76, 1207–1213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Graaf, N.; van Helden, M.J.G.; Textoris-Taube, K.; Chiba, T.; Topham, D.J.; Kloetzel, P.-M.; Zaiss, D.M.W.; Sijts, A.J.A.M. PA28 and the Proteasome Immunosubunits Play a Central and Independent Role in the Production of MHC Class I-Binding Peptides in Vivo. Eur. J. Immunol. 2011, 41, 926–935. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Norbury, C.C.; Cho, Y.; Yewdell, J.W.; Bennink, J.R. Immunoproteasomes Shape Immunodominance Hierarchies of Antiviral CD8(+) T Cells at the Levels of T Cell Repertoire and Presentation of Viral Antigens. J. Exp. Med. 2001, 193, 1319–1326. [Google Scholar] [CrossRef] [Green Version]
- Deol, P.; Zaiss, D.M.W.; Monaco, J.J.; Sijts, A.J.A.M. Rates of Processing Determine the Immunogenicity of Immunoproteasome-Generated Epitopes. J. Immunol. 2007, 178, 7557–7562. [Google Scholar] [CrossRef] [Green Version]
- Zanker, D.; Waithman, J.; Yewdell, J.W.; Chen, W. Mixed Proteasomes Function to Increase Viral Peptide Diversity and Broaden Antiviral CD8+ T Cell Responses. J. Immunol. 2013, 191, 52–59. [Google Scholar] [CrossRef] [Green Version]
- Yewdell, J.W. Confronting Complexity: Real-World Immunodominance in Antiviral CD8+ T Cell Responses. Immunity 2006, 25, 533–543. [Google Scholar] [CrossRef] [Green Version]
- Kedl, R.M.; Rees, W.A.; Hildeman, D.A.; Schaefer, B.; Mitchell, T.; Kappler, J.; Marrack, P. T Cells Compete for Access to Antigen-Bearing Antigen-Presenting Cells. J. Exp. Med. 2000, 192, 1105–1113. [Google Scholar] [CrossRef]
- Thomas, P.G.; Brown, S.A.; Keating, R.; Yue, W.; Morris, M.Y.; So, J.; Webby, R.J.; Doherty, P.C. Hidden Epitopes Emerge in Secondary Influenza Virus-Specific CD8+ T Cell Responses. J. Immunol. 2007, 178, 3091–3098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, P.S.; Lee, P.P.; Levy, D. A Theory of Immunodominance and Adaptive Regulation. Bull. Math. Biol. 2011, 73, 1645–1665. [Google Scholar] [CrossRef] [PubMed]
- Hensley, S.E.; Zanker, D.; Dolan, B.P.; David, A.; Hickman, H.D.; Embry, A.C.; Skon, C.N.; Grebe, K.M.; Griffin, T.A.; Chen, W.; et al. Unexpected Role for the Immunoproteasome Subunit LMP2 in Antiviral Humoral and Innate Immune Responses. J. Immunol. 2010, 184, 4115–4122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seifert, U.; Bialy, L.P.; Ebstein, F.; Bech-Otschir, D.; Voigt, A.; Schröter, F.; Prozorovski, T.; Lange, N.; Steffen, J.; Rieger, M.; et al. Immunoproteasomes Preserve Protein Homeostasis upon Interferon-Induced Oxidative Stress. Cell 2010, 142, 613–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pickering, A.M.; Koop, A.L.; Teoh, C.Y.; Ermak, G.; Grune, T.; Davies, K.J.A. The Immunoproteasome, the 20S Proteasome and the PA28αβ Proteasome Regulator Are Oxidative-Stress-Adaptive Proteolytic Complexes. Biochem. J. 2010, 432, 585–594. [Google Scholar] [CrossRef] [Green Version]
- Kalim, K.W.; Basler, M.; Kirk, C.J.; Groettrup, M. Immunoproteasome Subunit LMP7 Deficiency and Inhibition Suppresses Th1 and Th17 but Enhances Regulatory T Cell Differentiation. J. Immunol. 2012, 189, 4182–4193. [Google Scholar] [CrossRef]
- Muchamuel, T.; Basler, M.; Aujay, M.A.; Suzuki, E.; Kalim, K.W.; Lauer, C.; Sylvain, C.; Ring, E.R.; Shields, J.; Jiang, J.; et al. A Selective Inhibitor of the Immunoproteasome Subunit LMP7 Blocks Cytokine Production and Attenuates Progression of Experimental Arthritis. Nat. Med. 2009, 15, 781–787. [Google Scholar] [CrossRef] [Green Version]
- Qureshi, N.; Morrison, D.C.; Reis, J. Proteasome Protease Mediated Regulation of Cytokine Induction and Inflammation. Biochim. Biophys. Acta 2012, 1823, 2087–2093. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, N.; Gonzalez, E.; Visekruna, A.; Kühl, A.A.; Loddenkemper, C.; Mollenkopf, H.; Kaufmann, S.H.E.; Steinhoff, U.; Joeris, T. Targeting the Proteasome: Partial Inhibition of the Proteasome by Bortezomib or Deletion of the Immunosubunit LMP7 Attenuates Experimental Colitis. Gut 2010, 59, 896–906. [Google Scholar] [CrossRef]
- Nussbaum, A.K.; Rodriguez-Carreno, M.P.; Benning, N.; Botten, J.; Whitton, J.L. Immunoproteasome-Deficient Mice Mount Largely Normal CD8+ T Cell Responses to Lymphocytic Choriomeningitis Virus Infection and DNA Vaccination. J. Immunol. 2005, 175, 1153–1160. [Google Scholar] [CrossRef] [Green Version]
- Tu, L.; Moriya, C.; Imai, T.; Ishida, H.; Tetsutani, K.; Duan, X.; Murata, S.; Tanaka, K.; Shimokawa, C.; Hisaeda, H.; et al. Critical Role for the Immunoproteasome Subunit LMP7 in the Resistance of Mice to Toxoplasma Gondii Infection. Eur. J. Immunol. 2009, 39, 3385–3394. [Google Scholar] [CrossRef] [PubMed]
- Ersching, J.; Vasconcelos, J.R.; Ferreira, C.P.; Caetano, B.C.; Machado, A.V.; Bruna-Romero, O.; Baron, M.A.; Ferreira, L.R.P.; Cunha-Neto, E.; Rock, K.L.; et al. The Combined Deficiency of Immunoproteasome Subunits Affects Both the Magnitude and Quality of Pathogen- and Genetic Vaccination-Induced CD8+ T Cell Responses to the Human Protozoan Parasite Trypanosoma Cruzi. PLoS Pathog. 2016, 12, e1005593. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, G.; Gomes, M.T.R.; Campos, P.C.; Marinho, F.V.; de Assis, N.R.G.; Silveira, T.N.; Oliveira, S.C. Immunoproteasome Subunits Are Required for CD8+ T Cell Function and Host Resistance to Brucella Abortus Infection in Mice. Infect Immun. 2018, 86, e00615-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwendinger, M.; Thiry, G.; De Vos, B.; Leroux-Roels, G.; Bruhwyler, J.; Huygens, A.; Ganeff, C.; Buchinger, H.; Orlinger, K.K.; Pinschewer, D.D.; et al. A Randomized Dose-Escalating Phase I Trial of a Replication-Deficient Lymphocytic Choriomeningitis Virus Vector-Based Vaccine Against Human Cytomegalovirus. J. Infect Dis. 2020, jiaa121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samy, N.; Reichhardt, D.; Schmidt, D.; Chen, L.M.; Silbernagl, G.; Vidojkovic, S.; Meyer, T.P.; Jordan, E.; Adams, T.; Weidenthaler, H.; et al. Safety and Immunogenicity of Novel Modified Vaccinia Ankara-Vectored RSV Vaccine: A Randomized Phase I Clinical Trial. Vaccine 2020, 38, 2608–2619. [Google Scholar] [CrossRef] [PubMed]
- Sadoff, J.; Gray, G.; Vandebosch, A.; Cárdenas, V.; Shukarev, G.; Grinsztejn, B.; Goepfert, P.A.; Truyers, C.; Fennema, H.; Spiessens, B.; et al. Safety and Efficacy of Single-Dose Ad26.COV2.S Vaccine against Covid-19. N. Engl. J. Med. 2021, 384, 2187–2201. [Google Scholar] [CrossRef]
- Ramasamy, M.N.; Minassian, A.M.; Ewer, K.J.; Flaxman, A.L.; Folegatti, P.M.; Owens, D.R.; Voysey, M.; Aley, P.K.; Angus, B.; Babbage, G.; et al. Safety and Immunogenicity of ChAdOx1 NCoV-19 Vaccine Administered in a Prime-Boost Regimen in Young and Old Adults (COV002): A Single-Blind, Randomised, Controlled, Phase 2/3 Trial. Lancet 2021, 396, 1979–1993. [Google Scholar] [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 MRNA Covid-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef]
- Jackson, L.A.; Anderson, E.J.; Rouphael, N.G.; Roberts, P.C.; Makhene, M.; Coler, R.N.; McCullough, M.P.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; et al. An MRNA Vaccine against SARS-CoV-2—Preliminary Report. N. Engl. J. Med. 2020, 383, 1920–1931. [Google Scholar] [CrossRef]
- Ciotti, M.; Ciccozzi, M.; Pieri, M.; Bernardini, S. The COVID-19 Pandemic: Viral Variants and Vaccine Efficacy. Crit. Rev. Clin. Lab. Sci. 2021, 1–10. [Google Scholar] [CrossRef]
- Wu, T.; Guan, J.; Handel, A.; Tscharke, D.C.; Sidney, J.; Sette, A.; Wakim, L.M.; Sng, X.Y.X.; Thomas, P.G.; Croft, N.P.; et al. Quantification of Epitope Abundance Reveals the Effect of Direct and Cross-Presentation on Influenza CTL Responses. Nat. Commun. 2019, 10, 2846. [Google Scholar] [CrossRef] [Green Version]
- Kedl, R.M.; Schaefer, B.C.; Kappler, J.W.; Marrack, P. T Cells Down-Modulate Peptide-MHC Complexes on APCs in Vivo. Nat. Immunol. 2002, 3, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Bousso, P.; Robey, E. Dynamics of CD8+ T Cell Priming by Dendritic Cells in Intact Lymph Nodes. Nat. Immunol. 2003, 4, 579–585. [Google Scholar] [CrossRef]
- Huang, M.; Xu, R.; Triffon, C.; Mifsud, N.; Chen, W. Broad-Based Influenza-Specific CD8+ T Cell Response without the Typical Immunodominance Hierarchy and Its Potential Implication. Viruses 2021, 13, 1080. [Google Scholar] [CrossRef] [PubMed]
- Murata, S.; Sasaki, K.; Kishimoto, T.; Niwa, S.-I.; Hayashi, H.; Takahama, Y.; Tanaka, K. Regulation of CD8+ T Cell Development by Thymus-Specific Proteasomes. Science 2007, 316, 1349–1353. [Google Scholar] [CrossRef]
- Gaczynska, M.; Goldberg, A.L.; Tanaka, K.; Hendil, K.B.; Rock, K.L. Proteasome Subunits X and Y Alter Peptidase Activities in Opposite Ways to the Interferon-Gamma-Induced Subunits LMP2 and LMP7. J. Biol. Chem. 1996, 271, 17275–17280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, K.; Takada, K.; Ohte, Y.; Kondo, H.; Sorimachi, H.; Tanaka, K.; Takahama, Y.; Murata, S. Thymoproteasomes Produce Unique Peptide Motifs for Positive Selection of CD8(+) T Cells. Nat. Commun. 2015, 6, 7484. [Google Scholar] [CrossRef] [Green Version]
- Nitta, T.; Murata, S.; Sasaki, K.; Fujii, H.; Ripen, A.M.; Ishimaru, N.; Koyasu, S.; Tanaka, K.; Takahama, Y. Thymoproteasome Shapes Immunocompetent Repertoire of CD8+ T Cells. Immunity 2010, 32, 29–40. [Google Scholar] [CrossRef] [Green Version]
- Xing, Y.; Jameson, S.C.; Hogquist, K.A. Thymoproteasome Subunit-Β5T Generates Peptide-MHC Complexes Specialized for Positive Selection. Proc. Natl. Acad. Sci. USA 2013, 110, 6979–6984. [Google Scholar] [CrossRef] [Green Version]
- Kincaid, E.Z.; Murata, S.; Tanaka, K.; Rock, K.L. Specialized Proteasome Subunits Have an Essential Role in the Thymic Selection of CD8(+) T Cells. Nat. Immunol. 2016, 17, 938–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaiss, D.M.W.; Bekker, C.P.J.; Gröne, A.; Lie, B.A.; Sijts, A.J.A.M. Proteasome Immunosubunits Protect against the Development of CD8 T Cell-Mediated Autoimmune Diseases. J. Immunol. 2011, 187, 2302–2309. [Google Scholar] [CrossRef] [Green Version]
- Di Lorenzo, T.P.; Peakman, M.; Roep, B.O. Translational Mini-Review Series on Type 1 Diabetes: Systematic Analysis of T Cell Epitopes in Autoimmune Diabetes. Clin. Exp. Immunol. 2007, 148, 1–16. [Google Scholar] [CrossRef]
- Khilji, M.S.; Verstappen, D.; Dahlby, T.; Burstein Prause, M.C.; Pihl, C.; Bresson, S.E.; Bryde, T.H.; Keller Andersen, P.A.; Klindt, K.; Zivkovic, D.; et al. The Intermediate Proteasome Is Constitutively Expressed in Pancreatic Beta Cells and Upregulated by Stimulatory, Low Concentrations of Interleukin 1 β. PLoS ONE 2020, 15, e0222432. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.V. Genetics of Proteasome Diseases. Sci. Cairo 2013, 2013, 637629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torrelo, A.; Patel, S.; Colmenero, I.; Gurbindo, D.; Lendínez, F.; Hernández, A.; López-Robledillo, J.C.; Dadban, A.; Requena, L.; Paller, A.S. Chronic atypical neutrophilic dermatosis with lipodystrophy and elevated temperature (CANDLE) syndrome. J. Am. Acad. Dermatol. 2010, 62, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.K.; Xing, C.; DeMartino, G.N.; Mizrachi, D.; Hernandez, H.D.; Sousa, A.B.; Martínez de Villarreal, L.; dos Santos, H.G.; Garg, A. PSMB8 encoding the β5i proteasome subunit is mutated in joint contractures, muscle atrophy, microcytic anemia, and panniculitis-induced lipodystrophy syndrome. Am. J. Hum. Genet 2010, 87, 866–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arima, K.; Kinoshita, A.; Mishima, H.; Kanazawa, N.; Kaneko, T.; Mizushima, T.; Ichinose, K.; Nakamura, H.; Tsujino, A.; Kawakami, A.; et al. Proteasome assembly defect due to a proteasome subunit beta type 8 (PSMB8) mutation causes the autoinflammatory disorder, Nakajo-Nishimura syndrome. Proc. Natl. Acad. Sci. USA 2011, 108, 14914–14919. [Google Scholar] [CrossRef] [Green Version]
- Kitamura, A.; Maekawa, Y.; Uehara, H.; Izumi, K.; Kawachi, I.; Nishizawa, M.; Toyoshima, Y.; Takahashi, H.; Standley, D.M.; Tanaka, K.; et al. A mutation in the immunoproteasome subunit PSMB8 causes autoinflammation and lipodystrophy in humans. J. Clin. Investig. 2011, 121, 4150–4160. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Ramot, Y.; Torrelo, A.; Paller, A.S.; Si, N.; Babay, S.; Kim, P.W.; Sheikh, A.; Lee, C.C.; Chen, Y.; et al. Mutations in proteasome subunit beta type 8 cause chronic atypical neutrophilic dermatosis with lipodystrophy and elevated temperature with evidence of genetic and phenotypic heterogeneity. Arthritis Rheum. 2012, 64, 895–907. [Google Scholar] [CrossRef] [Green Version]
- Brehm, A.; Liu, Y.; Sheikh, A.; Marrero, B.; Omoyinmi, E.; Zhou, Q.; Montealegre, G.; Biancotto, A.; Reinhardt, A.; de Jesus, A.A.; et al. Additive loss-of-function proteasome subunit mutations in CANDLE/PRAAS patients promote type I IFN production. J. Clin. Investig. 2015, 125, 4196–4211. [Google Scholar] [CrossRef]
- Ebstein, F.; Poli Harlowe, M.C.; Studencka-Turski, M.; Krüger, E. Contribution of the Unfolded Protein Response (UPR) to the Pathogenesis of Proteasome-Associated Autoinflammatory Syndromes (PRAAS). Front. Immunol. 2019, 10, 2756. [Google Scholar] [CrossRef]
- Xu, Y.; Liu, G.; Zhou, Y.; Lu, Z.; Shi, Z.; Wang, J. The Genetic Association between LMP2 and LMP7 Polymorphisms and Susceptibility of Insulin Dependent Diabetes Mellitus: A Meta-Analysis. Med. Baltim. 2020, 99, e19482. [Google Scholar] [CrossRef] [PubMed]
- Safari-Alighiarloo, N.; Taghizadeh, M.; Tabatabaei, S.M.; Shahsavari, S.; Namaki, S.; Khodakarim, S.; Rezaei-Tavirani, M. Identification of New Key Genes for Type 1 Diabetes through Construction and Analysis of Protein-Protein Interaction Networks Based on Blood and Pancreatic Islet Transcriptomes. J. Diabetes 2017, 9, 764–777. [Google Scholar] [CrossRef]
- Safari-Alighiarloo, N.; Taghizadeh, M.; Mohammad Tabatabaei, S.; Namaki, S.; Rezaei-Tavirani, M. Identification of Common Key Genes and Pathways between Type 1 Diabetes and Multiple Sclerosis Using Transcriptome and Interactome Analysis. Endocrine 2020, 68, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Sjakste, T.; Paramonova, N.; Osina, K.; Dokane, K.; Sokolovska, J.; Sjakste, N. Genetic Variations in the PSMA3, PSMA6 and PSMC6 Genes Are Associated with Type 1 Diabetes in Latvians and with Expression Level of Number of UPS-Related and T1DM-Susceptible Genes in HapMap Individuals. Mol. Genet. Genom. 2016, 291, 891–903. [Google Scholar] [CrossRef]
- Kimura, H.; Caturegli, P.; Takahashi, M.; Suzuki, K. New Insights into the Function of the Immunoproteasome in Immune and Nonimmune Cells. J. Immunol. Res. 2015, 2015, 541984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angeles, A.; Fung, G.; Luo, H. Immune and Non-Immune Functions of the Immunoproteasome. Front. Biosci. Landmark Ed. 2012, 17, 1904–1916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nathan, J.A.; Spinnenhirn, V.; Schmidtke, G.; Basler, M.; Groettrup, M.; Goldberg, A.L. Immuno- and constitutive proteasomes do not differ in their abilities to degrade ubiquitinated proteins. Cell 2013, 152, 1184–1194. [Google Scholar] [CrossRef] [Green Version]
- Ebstein, F.; Voigt, A.; Lange, N.; Warnatsch, A.; Schröter, F.; Prozorovski, T.; Kuckelkorn, U.; Aktas, O.; Seifert, U.; Kloetzel, P.M.; et al. Immunoproteasomes are important for proteostasis in immune responses. Cell 2013, 152, 935–937. [Google Scholar] [CrossRef] [Green Version]
- Basler, M.; Christ, M.; Goebel, H.; Groettrup, M. Immunoproteasome Upregulation Is Not Required to Control Protein Homeostasis during Viral Infection. J. Immunol. 2021, 206, 1697–1708. [Google Scholar] [CrossRef]
- Rasid, O.; Meulenbroeks, C.; Gröne, A.; Zaiss, D.; Sijts, A. Enhanced Inflammatory Potential of CD4+ T-Cells That Lack Proteasome Immunosubunit Expression, in a T-Cell Transfer-Based Colitis Model. PLoS ONE 2014, 9, e95378. [Google Scholar] [CrossRef] [PubMed]
- Basler, M.; Mundt, S.; Bitzer, A.; Schmidt, C.; Groettrup, M. The Immunoproteasome: A Novel Drug Target for Autoimmune Diseases. Clin. Exp. Rheumatol. 2015, 33, S74–S79. [Google Scholar] [PubMed]
- Basler, M.; Groettrup, M. Recent Insights How Combined Inhibition of Immuno/Proteasome Subunits Enables Therapeutic Efficacy. Genes Immun. 2020, 21, 273–287. [Google Scholar] [CrossRef]
- Huber, E.M.; Groll, M. A Nut for Every Bolt: Subunit-Selective Inhibitors of the Immunoproteasome and Their Therapeutic Potential. Cells 2021, 10, 1929. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, T.W.; Downey-Kopyscinski, S.L.; Fields, J.L.; Rahme, G.J.; Colley, W.C.; Israel, M.A.; Maksimenko, A.V.; Fiering, S.N.; Kisselev, A.F. Activity of Immunoproteasome Inhibitor ONX-0914 in Acute Lymphoblastic Leukemia Expressing MLL-AF4 Fusion Protein. Sci. Rep. 2021, 11, 10883. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van den Eshof, B.L.; Medfai, L.; Nolfi, E.; Wawrzyniuk, M.; Sijts, A.J.A.M. The Function of Immunoproteasomes—An Immunologists’ Perspective. Cells 2021, 10, 3360. https://doi.org/10.3390/cells10123360
van den Eshof BL, Medfai L, Nolfi E, Wawrzyniuk M, Sijts AJAM. The Function of Immunoproteasomes—An Immunologists’ Perspective. Cells. 2021; 10(12):3360. https://doi.org/10.3390/cells10123360
Chicago/Turabian Stylevan den Eshof, Bart L., Lobna Medfai, Emanuele Nolfi, Magdalena Wawrzyniuk, and Alice J. A. M. Sijts. 2021. "The Function of Immunoproteasomes—An Immunologists’ Perspective" Cells 10, no. 12: 3360. https://doi.org/10.3390/cells10123360
APA Stylevan den Eshof, B. L., Medfai, L., Nolfi, E., Wawrzyniuk, M., & Sijts, A. J. A. M. (2021). The Function of Immunoproteasomes—An Immunologists’ Perspective. Cells, 10(12), 3360. https://doi.org/10.3390/cells10123360