

Hepatic Polarized Differentiation Promoted the Maturity and Liver Function of Human Embryonic Stem Cell-Derived Hepatocytes via Activating Hippo and AMPK Signaling Pathways


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Establishment of the Polarized hEHs
2.2. Isolation and Culture of Primary Human Hepatocytes
2.3. Flow Cytometry Analysis
2.4. Assays for Albumin Secretion, Urea Nitrogen and Total Bile Acids
2.5. Determination of Cytochrome p450 Activity (CYP3A4)
2.6. Assay for Paracellular Permeability
2.7. Quantitative Real-Time Polymerase Chain Reaction
2.8. Western Blot Analysis
2.9. Immunofluorescence Staining
2.10. RNA Sequencing
2.11. Statistics
3. Results
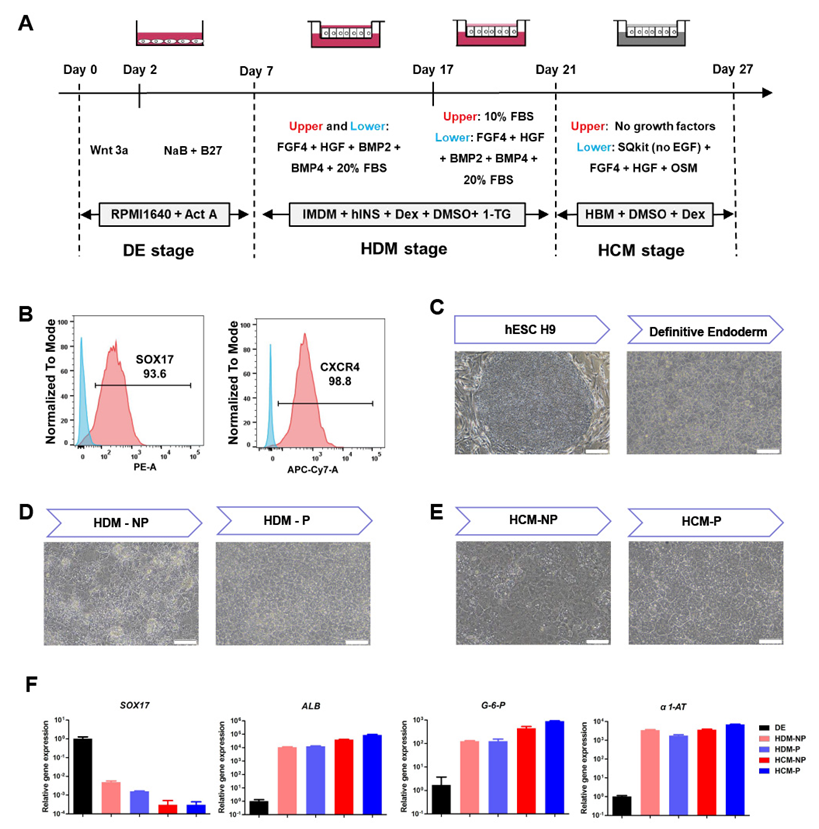
3.1. Generation of the Polarized hEHs
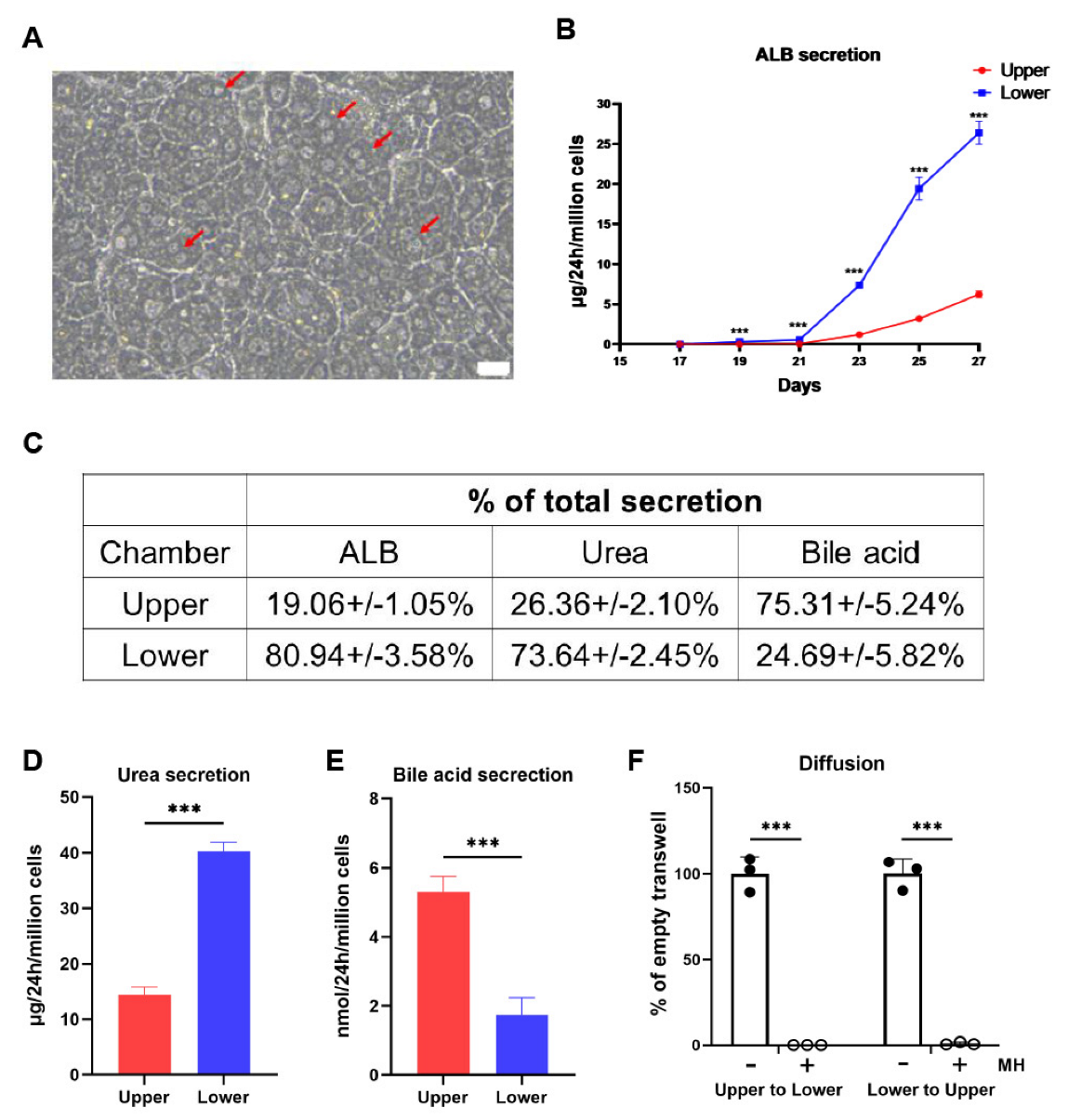
3.2. The Polarized hEHs Directionally Secreted the Plasma Proteins, Urea and Bile Acids
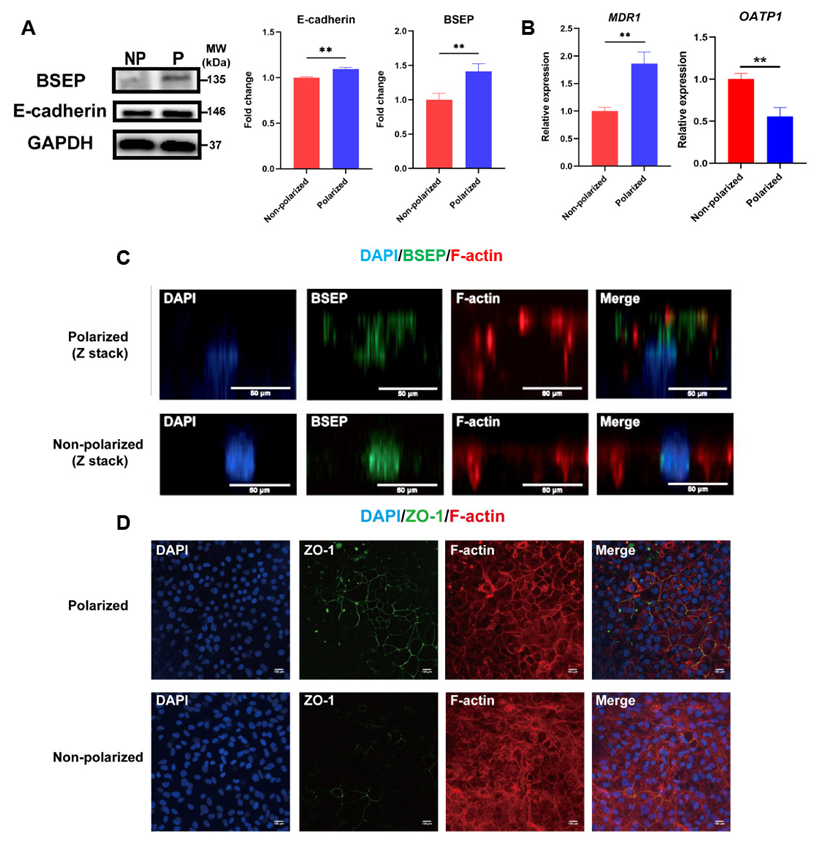
3.3. Identification of Apical Membrane Proteins and BBIB in the Polarized hEHs
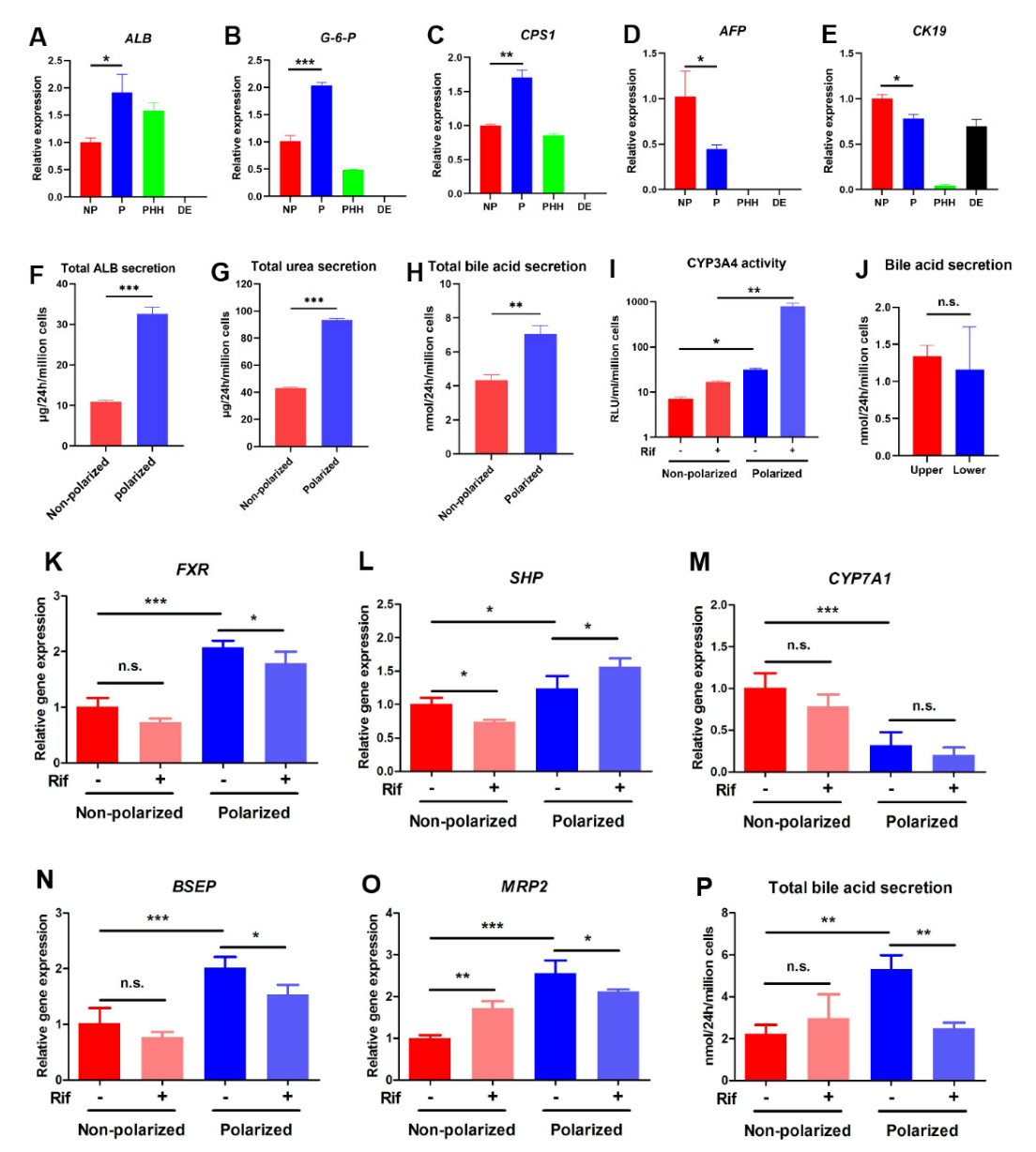
3.4. The Polarized Differentiation Improved the Maturation and Liver Function of hEHs
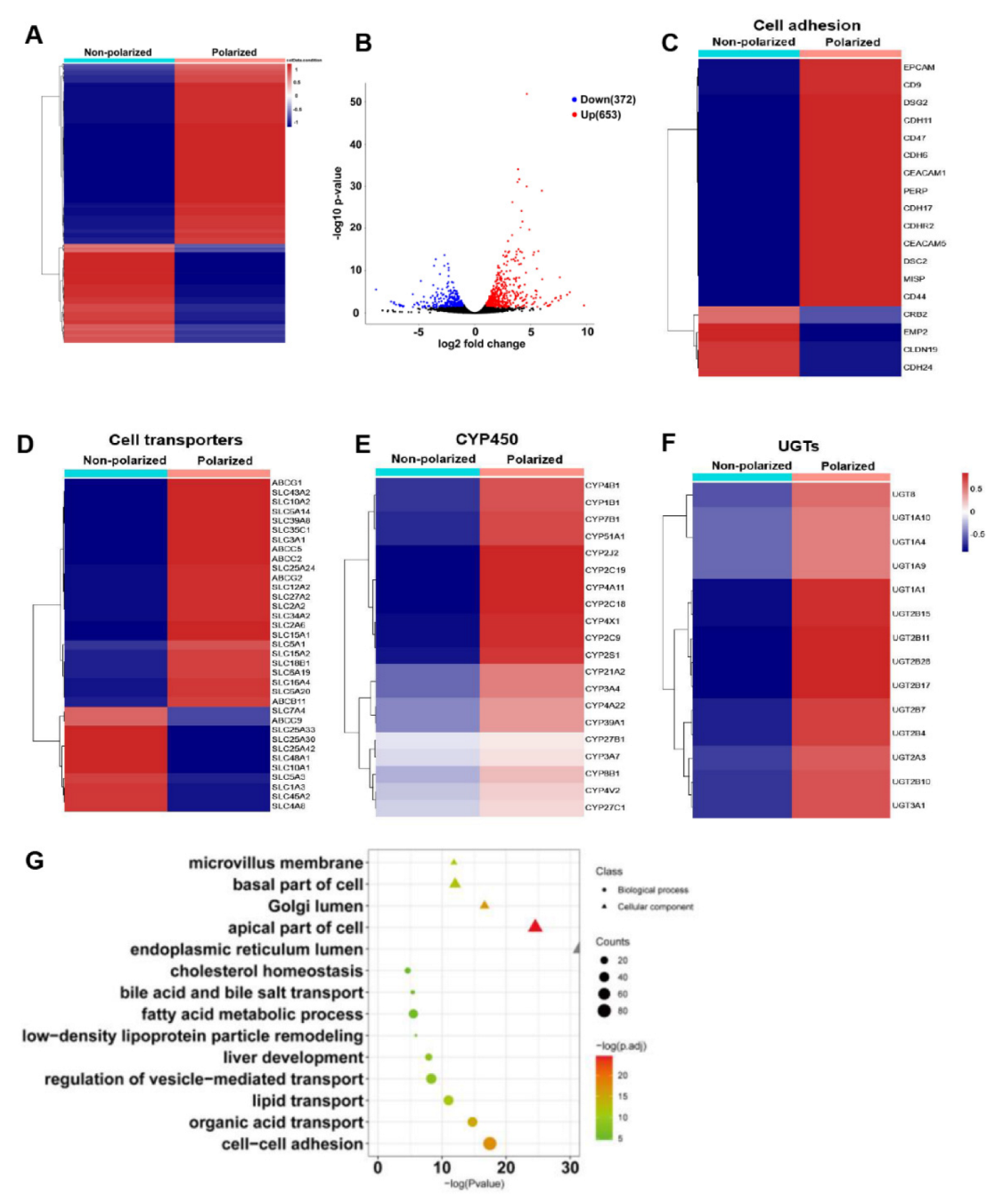
3.5. The Polarized Differentiation Altered the Gene Expression Profiles of hEHs
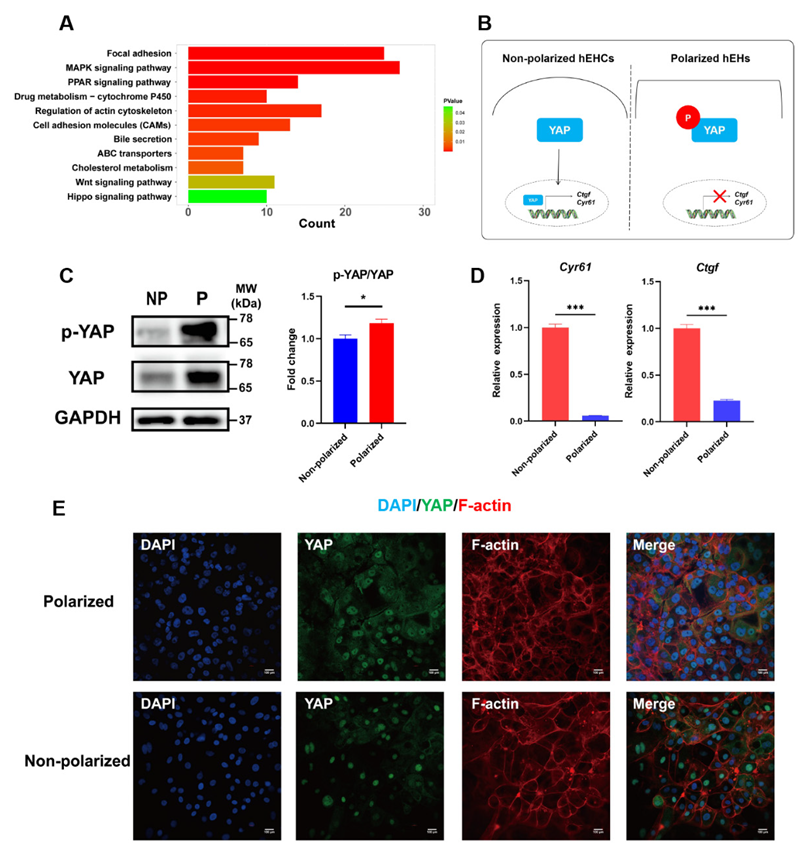
3.6. Hippo Signaling Pathway Was Activated in the Polarized hEHs
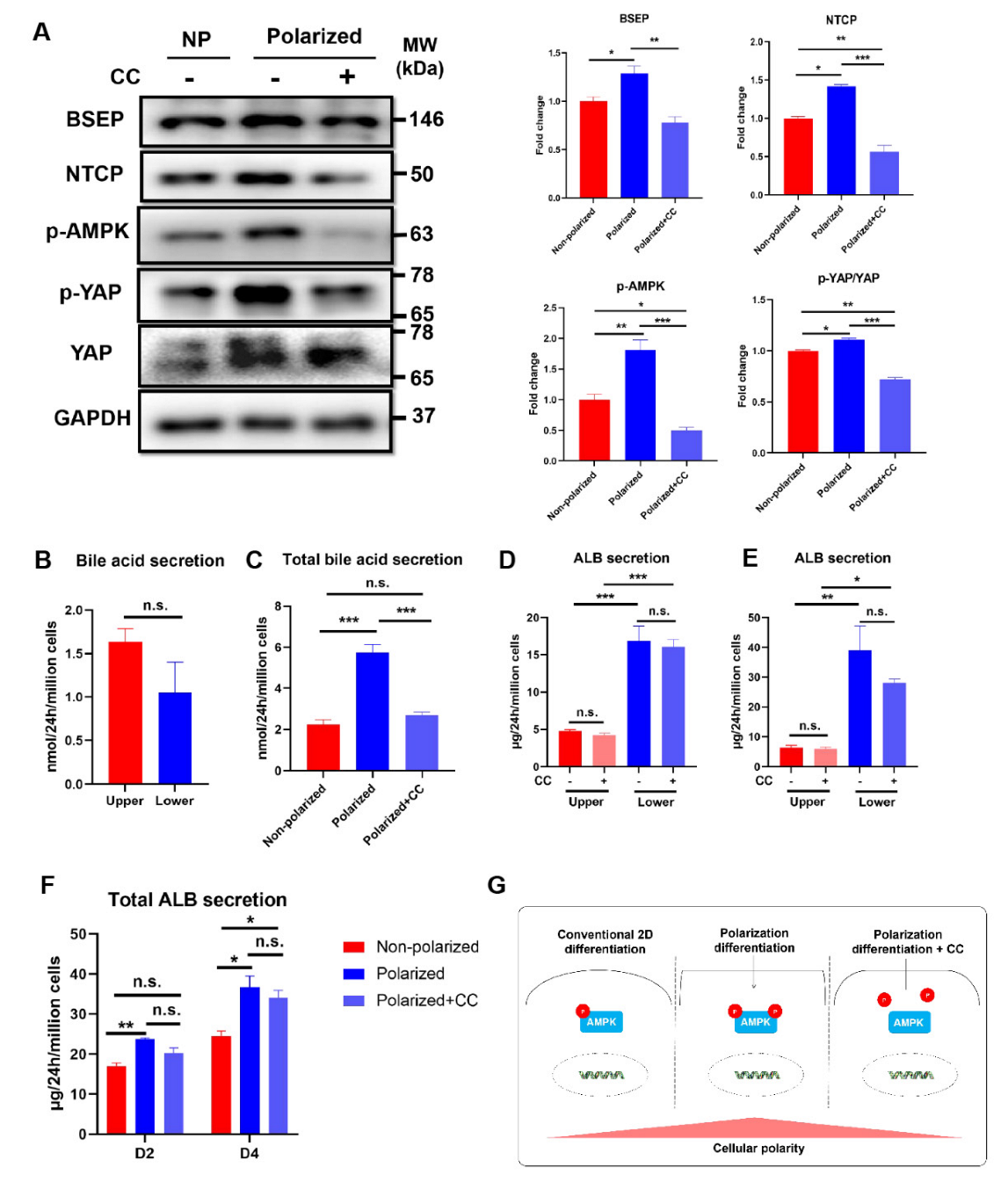
3.7. Activation of AMPK Signaling Pathway Was Involved in the Maintenance of the Polarity in the Polarized hEHs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Luce, E.; Messina, A.; Duclos-Vallée, J. Dubart-Kupperschmitt A. Advanced Techniques and Awaited Clinical Applications for Human Pluripotent Stem Cell Differentiation into Hepatocytes. Hepatology 2021, 74, 1101–1116. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Wu, J.; Yang, L.; Deng, X.; Qiu, W.L.; Zhou, Y.; Chen, Y.; Li, X.; Yu, L.; Li, H.; et al. Large-scale Generation of Functional and Transplantable Hepatocytes and Cholangiocytes from Human Endoderm Stem Cells. Cell Rep. 2020, 33, 108455. [Google Scholar] [CrossRef] [PubMed]
- Pasqua, M.; Gesù, R.D.; Chinnici, C.M.; Conaldi, P.G.; Francipane, M.G. Generation of Hepatobiliary Cell Lineages from Human Induced Pluripotent Stem Cells: Applications in Disease Modeling and Drug Screening. Int. J. Mol. Sci. 2021, 22, 8227. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, R.; Togo, S.; Kimura, M.; Shinozawa, T.; Koido, M.; Koike, H.; Thompson, W.; Karns, R.A.; Mayhew, C.N.; McGrath, P.S.; et al. Modeling Steatohepatitis in Humans with Pluripotent Stem Cell-derived Organoids. Cell Metab. 2019, 30, 374–384.e6. [Google Scholar] [CrossRef]
- Xiang, C.; Du, Y.; Meng, G.; Yi, L.S.; Sun, S.C.; Song, N.; Zhang, X.; Xiao, Y.; Wang, J.; Yi, Z.; et al. Long-term Functional Maintenance of Primary Human Hepatocytes in vitro. Science 2019, 364, 399–402. [Google Scholar] [CrossRef]
- Hu, H.; Gehart, H.; Artegiani, B.; Lpez-Iglesias, C.; Dekkers, F.; Basak, O.; van Es, J.; Chuva de Sousa Lopes, S.M.; Begthel, H.; Korving, J.; et al. Long-term Expansion of Functional Mouse and Human Hepatocytes as 3D Organoids. Cell 2018, 175, 1591–1606. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Tang, X.; Wang, Y.; Wang, N.; Chen, Q.; Xie, J.; Liu, S.; Zhong, Z.; Qiu, Y.; Situ, P.; et al. Dextran Sulfate Prevents Excess Aggregation of Human Pluripotent Stem Cells in 3D Culture by Inhibiting ICAM1 Expression Coupled with Down-regulating E-cadherin through Activating the Wnt Signaling Pathway. Stem Cell Res. Ther. 2022, 13, 218. [Google Scholar] [CrossRef]
- Si-Tayeb, K.; Noto, F.K.; Nagaoka, M.; Li, J.X.; Battle, M.A.; Duris, C.; North, P.E.; Dalton, S.; Duncan, S.A. Highly Efficient Generation of Human Hepatocyte-like Cells from Induced Pluripotent Stem Cells. Hepatology 2010, 51, 297–305. [Google Scholar] [CrossRef] [Green Version]
- Lynch, S.; Pridgeon, C.S.; Duckworth, C.A.; Sharma, P.; Park, B.K.; Goldring, C.E. Stem Cell Models as An in vitro Model for Predictive Toxicology. Biochem. J. 2019, 476, 1149–1158. [Google Scholar] [CrossRef] [Green Version]
- Shinozawa, T.; Kimura, M.; Cai, Y.; Saiki, N.; Yoneyama, Y.; Ouchi, R.; Koike, H.; Maezawa, M.; Zhang, R.R.; Dunn, A.; et al. High-fidelity Drug-induced Liver Injury Screen Using Human Pluripotent Stem Cell-derived Organoids. Gastroenterology 2021, 160, 831–846. [Google Scholar] [CrossRef]
- Wang, J.; Ren, H.; Liu, Y.; Sun, L.; Zhang, Z.; Zhao, Y.; Shi, X. Bioinspired Artificial Liver System with Hipsc-derived Hepatocytes for Acute Liver Failure Treatment. Adv. Healthc. Mater. 2021, 10, 2101580. [Google Scholar] [CrossRef] [PubMed]
- Zabulica, M.; Srinivasan, R.C.; Vosough, M.; Hammarstedt, C.; Wu, T.; Gramignoli, R.; Ellis, E.; Kannisto, K.; Collin de l’Hortet, A.; Takeishi, K.; et al. Guide to The Assessment of Mature Liver Gene Expression in Stem Cell-derived Hepatocytes. Stem Cells Dev. 2019, 28, 907–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treyer, A.; Müsch, A. Hepatocyte Polarity. Compr. Physiol. 2013, 3, 243–287. [Google Scholar] [PubMed] [Green Version]
- Gissen, P.; Arias, I.M. Structural and Functional Hepatocyte Polarity and Liver Disease. J. Hepatol. 2015, 63, 1023–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeigerer, A.; Wuttke, A.; Marsico, G.; Seifert, S.; Kalaidzidis, Y.; Zerial, M. Functional Properties of Hepatocytes in vitro are Correlated with Cell Polarity Maintenance. Exp. Cell Res. 2017, 350, 242–252. [Google Scholar] [CrossRef]
- Palakkan, A.A.; Drummond, R.; Anderson, R.A.; Greenhough, S.; Tv, K.; Hay, D.C.; Ross, J.M. Polarisation and Functional Characterisation of Hepatocytes Derived from Human Embryonic and Mesenchymal Stem Cells. Biomed. Rep. 2015, 3, 626–636. [Google Scholar] [CrossRef] [Green Version]
- Dao Thi, V.L.; Wu, X.; Belote, R.L.; Andreo, U.; Takacs, C.N.; Fernandez, J.P.; Vale-Silva, L.; Prallet, S.; Decker, C.; Fu, R.; et al. Stem Cell-derived Polarized Hepatocytes. Nat Commun. 2020, 11, 1677. [Google Scholar] [CrossRef] [Green Version]
- Duan, Y.; Ma, X.; Zou, W.; Wang, C.; Bahbahan, I.S.; Ahuja, T.P.; Tolstikov, V.; Zern, M.A. Differentiation and Characterization of Metabolically Functioning Hepatocytes from Human Embryonic Stem Cells. Stem Cells 2010, 28, 674–686. [Google Scholar] [CrossRef]
- Bushweller, L.; Zhao, Y.; Zhang, F.; Wu, X. Generation of Human Pluripotent Stem Cell-derived Polarized Hepatocytes. Curr. Protoc. 2022, 2, e345. [Google Scholar] [CrossRef]
- Fu, G.; Huang, W.; Zeng, M.; Zhou, X.; Wu, H.; Liu, C.; Wu, H.; Weng, J.; Zhang, H.; Cai, Y.; et al. Expansion and Differentiation of Human Hepatocyte-derived Liver Progenitor-like Cells and Their Use for The Study of Hepatotropic Pathogens. Cell Res. 2019, 29, 8–22. [Google Scholar] [CrossRef]
- Broutier, L.; Andersson-Rolf, A.; Hindley, C.J.; Boj, S.F.; Clevers, H.; Koo, B.K.; Huch, M. Culture and establishment of self-renewing human and mouse adult liver and pancreas 3D organoids and their genetic manipulation. Nat. Protoc. 2016, 11, 1724–1743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulze, R.J.; Schott, M.B.; Casey, C.A.; Tuma, P.L.; Mcniven, M.A. The Cell Biology of the Hepatocyte: A Membrane Trafficking Machine. J. Cell Biol. 2019, 218, 2096–2112. [Google Scholar] [CrossRef] [PubMed]
- Pradhan-Sundd, T.; Monga, S.P. Blood-bile Barrier: Morphology, Regulation, and Pathophysiology. Gene Expr. 2019, 19, 69–87. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; De Mets, R.; Monzel, C.; Acharya, V.; Toh, P.; Chin, J.F.L.; Van Hul, N.; Chuan Ng, I.; Yu, H.; Seng Ng, S.; et al. Biomimetic Niches Reveal the Minimal Cues to Trigger Apical Lumen Formation in Single Hepatocytes. Nat. Mater. 2020, 19, 1026–1035. [Google Scholar] [CrossRef]
- Wang, P.; Yang, Y.; Pang, G.; Zhang, C.; Wei, C.S.; Tao, X.F.; Liu, J.; Xu, J.; Zhang, W.; Shen, Y. Hepatocyte-derived MANF is Protective for Rifampicin-induced Cholestatic Hepatic Injury via Inhibiting ATF4-CHOP Signal Activation. Free Radic. Biol. Med. 2020, 162, 283–297. [Google Scholar] [CrossRef]
- Xu, B.Y.; Tang, X.D.; Chen, J.; Wu, H.B.; Chen, W.S.; Chen, L. Rifampicin Induces Clathrin-dependent Endocytosis and Ubiquitin–proteasome Degradation of MRP2 via Oxidative Stress-activated PKC-ERK/JNK/p38 and PI3K Signaling Pathways in HepG2 Cells. Acta Pharmacol. Sin. 2020, 41, 56–64. [Google Scholar] [CrossRef]
- Yang, Y.J.; Liu, L.; Zhang, X.Q.; Jiang, X.H.; Wang, L. Tanshinone IIA Prevents Rifampicin-induced Liver Injury by Regulating BSEP/NTCP Expression via Epigenetic Activation of NRF2. Liver Int. 2020, 40, 141–154. [Google Scholar] [CrossRef]
- Yamamoto, J.; Udono, M.; Miura, S.; Sekiya, S.; Suzuki, A. Cell Aggregation Culture Induces Functional Differentiation of Induced Hepatocyte-like Cells through Activation of Hippo Signaling. Cell Rep. 2018, 25, 183–198. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.W.S.; Fu, D. Bioenergetics for Hepatocyte Polarization: Coordination of Multiple Cellular Organelles and The Master Tegulator AMPK. Crit. Rev. Eukar. Gene 2019, 29, 483–497. [Google Scholar] [CrossRef]
- Decaens, C.; Durand, M.; Grosse, B.; Cassio, D. Which in vitro Models Could Be Best Used to Study Hepatocyte Polarity? Biol. Cell 2012, 100, 387–398. [Google Scholar] [CrossRef]
- Shen, T.; Liu, Y.; Shang, J.; Xie, Q.; Li, J.; Yan, M.; Xu, J.; Niu, J.; Liu, J.; Watkins, P.; et al. Incidence and Etiology of Drug-induced Liver Injury in Mainland China. Gastroenterology 2019, 156, 2230–2241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gijbels, E.; Vilas-Boas, V.; Deferm, N.; Devisscher, L.; Jaeschke, H.; Annaert, P.; Vinken, M. Mechanisms and in vitro Models of Drug-induced Cholestasis. Arch. Toxicol. 2019, 93, 1169–1186. [Google Scholar] [CrossRef] [PubMed]
- Natale, A.; Boeckmans, J.; Desmae, T.; De Boe, V.; De Kock, J.; Vanhaecke, T.; Rogiers, V.; Rodrigues, R. Hepatic Cells Derived from Human Skin Progenitors Show a Typical Phospholipidotic Response Upon Exposure to Amiodarone. Toxicol. Lett. 2018, 284, 184–194. [Google Scholar] [CrossRef]
- Raggi, C.; M’callum, M.A.; Pham, Q.T.; Gaub, P.; Selleri, S.; Baratang, N.V.; Mangahas, C.; Cagnone, G.; Reversade, B.; Joyal, J.S.; et al. Leveraging Interacting Signaling Pathways to Robustly Improve the Quality and Yield of Human Pluripotent Stem Cell-derived Hepatoblasts and Hepatocytes. Stem Cell Rep. 2022, 17, 584–598. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Tschudy-Seney, B.; Ma, X.; Zern, M.A.; Liu, P.; Duan, Y. Salvianolic Acid B Enhances Hepatic Differentiation of Human Embryonic Stem Cells through Upregulation of WNT Pathway and Inhibition of Notch Pathway. Stem Cells Dev. 2018, 27, 252–261. [Google Scholar] [CrossRef]
- Chen, J.; Gao, W.; Zhou, P.; Ma, X.; Tschudy-Seney, B.; Liu, C.; Zern, M.; Liu, P.; Duan, Y. Enhancement of Hepatocyte Differentiation from Human Embryonic Stem Cells by Chinese Medicine Fuzhenghuayu. Sci. Rep. 2016, 6, 18841. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Zhou, P.; Ma, X.; Benjamin, T.S.; Chen, J.; Magner, N.L.; Alexander, R.; Nolta, J.A.; Zern, M.A.; Duan, Y. Ethanol Negatively Regulates Hepatic Differentiation of hESC by Inhibition of the MAPK/ERK Signaling Pathway In Vitro. PLoS ONE 2014, 9, e112698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, F.X.; Zhao, B.; Guan, K.L. Hippo Pathway in Organ Size Control, Tissue Homeostasis, and Cancer. Cell 2015, 163, 811–828. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.C.; Graves, H.K.; Moya, I.M.; Tao, C.; Hamaratoglu, F.; Gladden, A.B.; Halder, G. Differential Regulation of The Hippo Pathway by Adherens Junctions and Apical-Basal Cell Polarity Modules. Proc. Natl. Acad. Sci. USA 2015, 112, 1785–1790. [Google Scholar] [CrossRef] [Green Version]
- Borreguero-Muoz, N.; Fletcher, G.C.; Aguilar-Aragon, M.; Elbediwy, A.; Vincent-Mistiaen, Z.I.; Thompson, B.J. The Hippo Pathway Integrates PI3K-Akt Signals with Mechanical and Polarity Cues to Control Tissue Growth. PLoS Biol. 2019, 17, e3000509. [Google Scholar]
- Alder, O.; Cullum, R.; Lee, S.; Kan, A.C.; Wei, W.; Yi, Y.; Garside, V.; Bilenky, M.; Griffith, M.; Morrissy, A.S.; et al. Hippo Signaling Influences HNF4A and FOXA2 Enhancer Switching During Hepatocyte Differentiation. Cell Rep. 2014, 9, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Genevet, A.; Tapon, N. The Hippo Pathway and Apico-basal Cell Polarity. Biochem. J. 2011, 436, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Foretz, M.; Viollet, B. Regulation of Hepatic Metabolism by AMPK. J. Hepatol. 2011, 54, 827–829. [Google Scholar] [CrossRef] [PubMed]
- Fu, D.; Wakabayashi, Y.; Ido, Y.; Lippincott-Schwartz, J.; Arias, I.M. Regulation of Bile Canalicular Network Formation and Maintenance by AMP-activated Protein Kinase and LKB1. J. Cell Sci. 2010, 123, 3294–3302. [Google Scholar] [CrossRef] [Green Version]
- Fu, D.; Wakabayashi, Y.; Lippincott-Schwartz, J.; Arias, I.M. Bile Acid Stimulates Hepatocyte Polarization through a cAMP-Epac-MEK-LKB1-AMPK Pathway. Proc. Natl. Acad. Sci. USA 2011, 108, 1403–1408. [Google Scholar] [CrossRef] [Green Version]
- Peng, L.; Li, Z.R.; Green, R.S.; Holzman, I.R.; Lin, J. Butyrate Enhances the Intestinal Barrier by Facilitating Tight Junction Assembly via Activation of AMP-activated Protein Kinase in Caco-2 Cell Monolayers. J. Nutr. 2009, 139, 1619–1625. [Google Scholar] [CrossRef] [Green Version]
- Zheng, B.; Cantley, L.C. Regulation of Epithelial Tight Junction Assembly and Disassembly by AMP-activated Protein Kinase. Proc. Natl. Acad. Sci. USA 2007, 104, 819–822. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Situ, P.; Chen, S.; Wu, H.; Zhang, X.; Liu, S.; Wang, Y.; Xie, J.; Chen, H.; Duan, Y. Hepatic Polarized Differentiation Promoted the Maturity and Liver Function of Human Embryonic Stem Cell-Derived Hepatocytes via Activating Hippo and AMPK Signaling Pathways. Cells 2022, 11, 4117. https://doi.org/10.3390/cells11244117
Wang J, Situ P, Chen S, Wu H, Zhang X, Liu S, Wang Y, Xie J, Chen H, Duan Y. Hepatic Polarized Differentiation Promoted the Maturity and Liver Function of Human Embryonic Stem Cell-Derived Hepatocytes via Activating Hippo and AMPK Signaling Pathways. Cells. 2022; 11(24):4117. https://doi.org/10.3390/cells11244117
Chicago/Turabian StyleWang, Jue, Ping Situ, Sen Chen, Haibin Wu, Xueyan Zhang, Shoupei Liu, Yiyu Wang, Jinghe Xie, Honglin Chen, and Yuyou Duan. 2022. "Hepatic Polarized Differentiation Promoted the Maturity and Liver Function of Human Embryonic Stem Cell-Derived Hepatocytes via Activating Hippo and AMPK Signaling Pathways" Cells 11, no. 24: 4117. https://doi.org/10.3390/cells11244117
APA StyleWang, J., Situ, P., Chen, S., Wu, H., Zhang, X., Liu, S., Wang, Y., Xie, J., Chen, H., & Duan, Y. (2022). Hepatic Polarized Differentiation Promoted the Maturity and Liver Function of Human Embryonic Stem Cell-Derived Hepatocytes via Activating Hippo and AMPK Signaling Pathways. Cells, 11(24), 4117. https://doi.org/10.3390/cells11244117