
Role of Autonomous Neuropathy in Diabetic Bone Regeneration

 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Pharmacological Treatments
2.3. Tibial Defect Model
2.4. Histology
2.5. Image Quantification
2.6. Microcomputed Tomographic Analysis (µCT)
2.7. Image Processing
2.8. Statistical Methods
3. Results
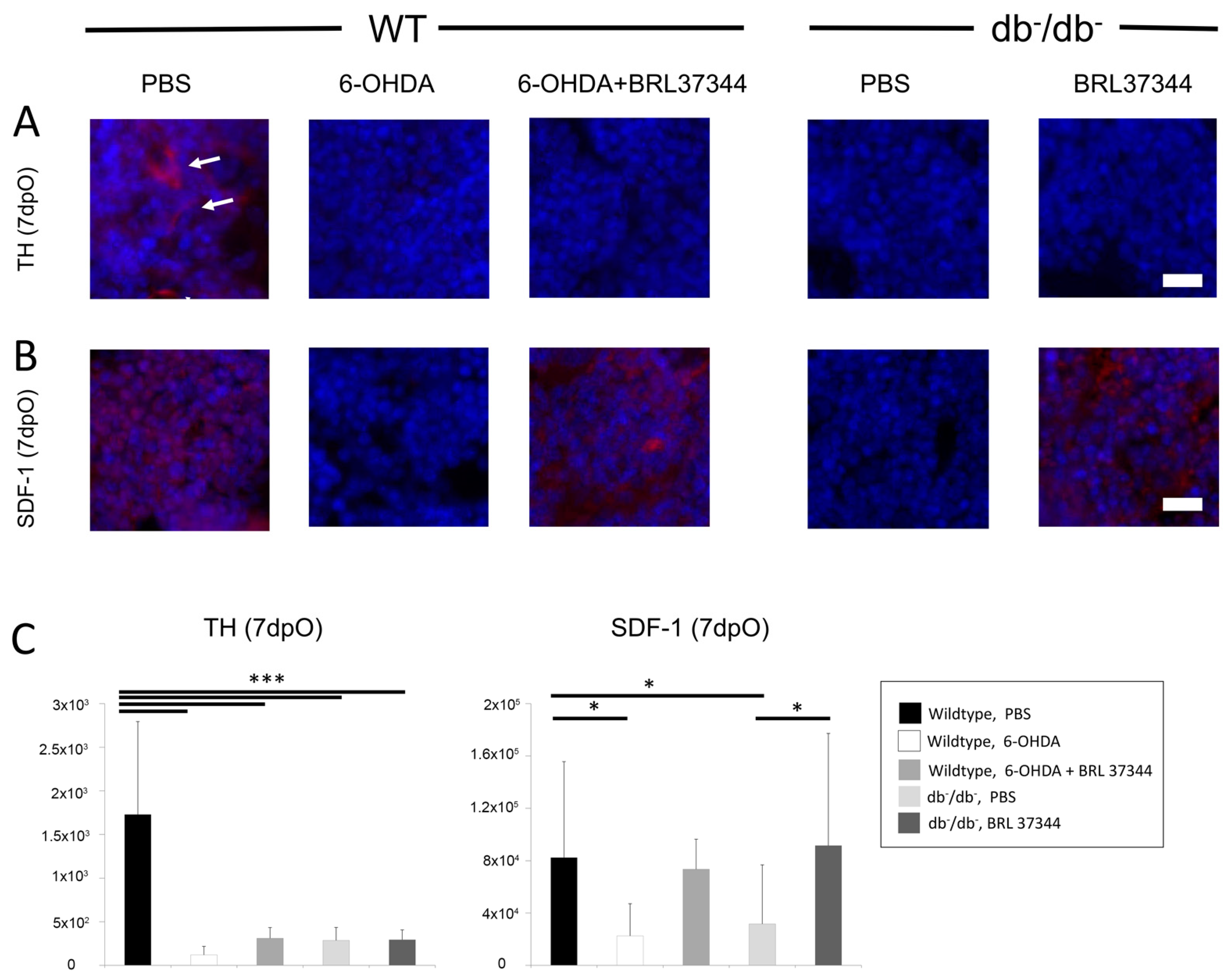
3.1. Activity of Sympathetic Nervous System in Bone of db−/db− Mice Is Decreased
3.2. Chemical Sympathectomy Reduces Bone Regeneration in Wildtype Mice Comparable to db−/db− Mice
3.3. BRL37344 Treatment Rescues Bone Regeneration of db−/db− Mice and Sympathectomized Wildtype Mice
3.4. Diminished Osteoblastogenesis and Bone Turnover Is Being Restored in Diabetic and Sympathectomized Animals by BRL37344 Treatment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kahn, S.E.; Cooper, M.E.; Del Prato, S. Pathophysiology and treatment of type 2 diabetes: Perspectives on the past, present, and future. Lancet 2014, 383, 1068–1083. [Google Scholar] [CrossRef] [Green Version]
- International Diabetes Federation. IDF Diabetes Atlas, 7th ed.; International Diabetes Federation: Brussels, Belgium, 2015. [Google Scholar]
- Baltzis, D.; Eleftheriadou, I.; Veves, A. Pathogenesis and treatment of impaired wound healing in diabetes mellitus: New insights. Adv. Ther. 2014, 31, 817–836. [Google Scholar] [CrossRef]
- Retzepi, M.; Donos, N. The effect of diabetes mellitus on osseous healing. Clin. Oral Implants Res. 2010, 21, 673–681. [Google Scholar] [CrossRef]
- Kaynak, G.; Birsel, O.; Guven, M.F.; Ogut, T. An overview of the Charcot foot pathophysiology. Diabet. Foot Ankle 2013, 4, 21117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyck, P.J.; Kratz, K.M.; Karnes, J.L.; Litchy, W.J.; Klein, R.; Pach, J.M.; Wilson, D.M.; O’Brien, P.C.; Melton, L.J., 3rd; Service, F.J. The prevalence by staged severity of various types of diabetic neuropathy, retinopathy, and nephropathy in a population-based cohort: The Rochester Diabetic Neuropathy Study. Neurology 1993, 43, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Scholte, A.J.; Schuijf, J.D.; Delgado, V.; Kok, J.A.; Bus, M.T.; Maan, A.C.; Stokkel, M.P.; Kharagitsingh, A.V.; Dibbets-Schneider, P.; van der Wall, E.E.; et al. Cardiac autonomic neuropathy in patients with diabetes and no symptoms of coronary artery disease: Comparison of 123I-metaiodobenzylguanidine myocardial scintigraphy and heart rate variability. Eur. J. Nucl. Med. Mol. Imaging 2010, 37, 1698–1705. [Google Scholar] [CrossRef] [Green Version]
- Tentolouris, N.; Marinou, K.; Kokotis, P.; Karanti, A.; Diakoumopoulou, E.; Katsilambros, N. Sudomotor dysfunction is associated with foot ulceration in diabetes. Diabet. Med. J. Br. Diabet. Assoc. 2009, 26, 302–305. [Google Scholar] [CrossRef]
- Wallner, C.; Schira, J.; Wagner, J.M.; Schulte, M.; Fischer, S.; Hirsch, T.; Richter, W.; Abraham, S.; Kneser, U.; Lehnhardt, M.; et al. Application of VEGFA and FGF-9 enhances angiogenesis, osteogenesis and bone remodeling in type 2 diabetic long bone regeneration. PLoS ONE 2015, 10, e0118823. [Google Scholar] [CrossRef] [Green Version]
- Fadini, G.P.; Ferraro, F.; Quaini, F.; Asahara, T.; Madeddu, P. Concise review: Diabetes, the bone marrow niche, and impaired vascular regeneration. Stem Cells Transl. Med. 2014, 3, 949–957. [Google Scholar] [CrossRef]
- Mendez-Ferrer, S.; Michurina, T.V.; Ferraro, F.; Mazloom, A.R.; Macarthur, B.D.; Lira, S.A.; Scadden, D.T.; Ma’ayan, A.; Enikolopov, G.N.; Frenette, P.S. Mesenchymal and haematopoietic stem cells form a unique bone marrow niche. Nature 2010, 466, 829–834. [Google Scholar] [CrossRef]
- Mendez-Ferrer, S.; Lucas, D.; Battista, M.; Frenette, P.S. Haematopoietic stem cell release is regulated by circadian oscillations. Nature 2008, 452, 442–447. [Google Scholar] [CrossRef] [PubMed]
- Takeda, S.; Elefteriou, F.; Levasseur, R.; Liu, X.; Zhao, L.; Parker, K.L.; Armstrong, D.; Ducy, P.; Karsenty, G. Leptin regulates bone formation via the sympathetic nervous system. Cell 2002, 111, 305–317. [Google Scholar] [CrossRef] [Green Version]
- Hanoun, M.; Zhang, D.; Mizoguchi, T.; Pinho, S.; Pierce, H.; Kunisaki, Y.; Lacombe, J.; Armstrong, S.A.; Duhrsen, U.; Frenette, P.S. Acute myelogenous leukemia-induced sympathetic neuropathy promotes malignancy in an altered hematopoietic stem cell niche. Cell Stem Cell 2014, 15, 365–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucas, D.; Scheiermann, C.; Chow, A.; Kunisaki, Y.; Bruns, I.; Barrick, C.; Tessarollo, L.; Frenette, P.S. Chemotherapy-induced bone marrow nerve injury impairs hematopoietic regeneration. Nat. Med. 2013, 19, 695–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albiero, M.; Poncina, N.; Tjwa, M.; Ciciliot, S.; Menegazzo, L.; Ceolotto, G.; Vigili de Kreutzenberg, S.; Moura, R.; Giorgio, M.; Pelicci, P.; et al. Diabetes causes bone marrow autonomic neuropathy and impairs stem cell mobilization via dysregulated p66Shc and Sirt1. Diabetes 2014, 63, 1353–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Z.; Wang, L.; Zhao, Y.; Cao, J.; Wang, T.; Liu, P.; Zhang, Y.; Yang, X.; Cheng, X.; Liu, B.; et al. Sympathetic denervation-induced MSC mobilization in distraction osteogenesis associates with inhibition of MSC migration and osteogenesis by norepinephrine/adrb3. PLoS ONE 2014, 9, e105976. [Google Scholar] [CrossRef] [Green Version]
- Busik, J.V.; Tikhonenko, M.; Bhatwadekar, A.; Opreanu, M.; Yakubova, N.; Caballero, S.; Player, D.; Nakagawa, T.; Afzal, A.; Kielczewski, J.; et al. Diabetic retinopathy is associated with bone marrow neuropathy and a depressed peripheral clock. J. Exp. Med. 2009, 206, 2897–2906. [Google Scholar] [CrossRef]
- Ferraro, F.; Lymperi, S.; Mendez-Ferrer, S.; Saez, B.; Spencer, J.A.; Yeap, B.Y.; Masselli, E.; Graiani, G.; Prezioso, L.; Rizzini, E.L.; et al. Diabetes impairs hematopoietic stem cell mobilization by altering niche function. Sci. Transl. Med. 2011, 3, 104ra101. [Google Scholar] [CrossRef] [Green Version]
- Song, D.; Jiang, X.; Zhu, S.; Li, W.; Khadka, A.; Hu, J. Denervation impairs bone regeneration during distraction osteogenesis in rabbit tibia lengthening. Acta Orthop. 2012, 83, 406–410. [Google Scholar] [CrossRef] [Green Version]
- Pagani, F.; Sibilia, V.; Cavani, F.; Ferretti, M.; Bertoni, L.; Palumbo, C.; Lattuada, N.; De Luca, E.; Rubinacci, A.; Guidobono, F. Sympathectomy alters bone architecture in adult growing rats. J. Cell. Biochem. 2008, 104, 2155–2164. [Google Scholar] [CrossRef]
- Niedermair, T.; Kuhn, V.; Doranehgard, F.; Stange, R.; Wieskotter, B.; Beckmann, J.; Salmen, P.; Springorum, H.R.; Straub, R.H.; Zimmer, A.; et al. Absence of substance P and the sympathetic nervous system impact on bone structure and chondrocyte differentiation in an adult model of endochondral ossification. Matrix Biol. J. Int. Soc. Matrix Biol. 2014, 38, 22–35. [Google Scholar] [CrossRef]
- Tepper, O.M.; Carr, J.; Allen, R.J., Jr.; Chang, C.C.; Lin, C.D.; Tanaka, R.; Gupta, S.M.; Levine, J.P.; Saadeh, P.B.; Warren, S.M. Decreased circulating progenitor cell number and failed mechanisms of stromal cell-derived factor-1alpha mediated bone marrow mobilization impair diabetic tissue repair. Diabetes 2010, 59, 1974–1983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horvat, S.; Bunger, L. Polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) assay for the mouse leptin receptor (Lepr(db)) mutation. Lab. Anim. 1999, 33, 380–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebbinghaus, M.; Gajda, M.; Boettger, M.K.; Schaible, H.G.; Brauer, R. The anti-inflammatory effects of sympathectomy in murine antigen-induced arthritis are associated with a reduction of Th1 and Th17 responses. Ann. Rheum. Dis. 2012, 71, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Arranz, L.; Sanchez-Aguilera, A.; Martin-Perez, D.; Isern, J.; Langa, X.; Tzankov, A.; Lundberg, P.; Muntion, S.; Tzeng, Y.S.; Lai, D.M.; et al. Neuropathy of haematopoietic stem cell niche is essential for myeloproliferative neoplasms. Nature 2014, 512, 78–81. [Google Scholar] [CrossRef]
- Behr, B.; Sorkin, M.; Manu, A.; Lehnhardt, M.; Longaker, M.T.; Quarto, N. Fgf-18 is required for osteogenesis but not angiogenesis during long bone repair. Tissue Eng. Part A 2011, 17, 2061–2069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouxsein, M.L.; Boyd, S.K.; Christiansen, B.A.; Guldberg, R.E.; Jepsen, K.J.; Muller, R. Guidelines for assessment of bone microstructure in rodents using micro-computed tomography. J. Bone Miner. Res. 2010, 25, 1468–1486. [Google Scholar] [CrossRef]
- Kim, L.R.; Whelpdale, K.; Zurowski, M.; Pomeranz, B. Sympathetic denervation impairs epidermal healing in cutaneous wounds. Wound Repair Regen. 1998, 6, 194–201. [Google Scholar] [CrossRef]
- Saburo, K.; Hiroshi, K.; Toshihiro, M. The role of sympathetic catecholaminergic nerves in wound healing. Burns 1982, 9, 135–141. [Google Scholar] [CrossRef]
- Bataille, C.; Mauprivez, C.; Hay, E.; Baroukh, B.; Brun, A.; Chaussain, C.; Marie, P.J.; Saffar, J.L.; Cherruau, M. Different sympathetic pathways control the metabolism of distinct bone envelopes. Bone 2012, 50, 1162–1172. [Google Scholar] [CrossRef]
- Vafaei, R.; Nassiri, S.M.; Siavashi, V. beta3-Adrenergic Regulation of EPC Features Through Manipulation of the Bone Marrow MSC Niche. J. Cell Biochem. 2017, 118, 4753–4761. [Google Scholar] [CrossRef]
- Wallner, C.; Abraham, S.; Wagner, J.M.; Harati, K.; Ismer, B.; Kessler, L.; Zollner, H.; Lehnhardt, M.; Behr, B. Local Application of Isogenic Adipose-Derived Stem Cells Restores Bone Healing Capacity in a Type 2 Diabetes Model. Stem Cells Transl. Med. 2016, 5, 836–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enríquez-Pérez, I.A.; Galindo-Ordoñez, K.E.; Pantoja-Ortíz, C.E.; Martínez-Martínez, A.; Acosta-González, R.I.; Muñoz-Islas, E.; Jiménez-Andrade, J.M. Streptozocin-induced type-1 diabetes mellitus results in decreased density of CGRP sensory and TH sympathetic nerve fibers that are positively correlated with bone loss at the mouse femoral neck. Neurosci. Lett. 2017, 655, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Bubb, K.J.; Ravindran, D.; Cartland, S.P.; Finemore, M.; Clayton, Z.E.; Tsang, M.; Tang, O.; Kavurma, M.M.; Patel, S.; Figtree, G.A. β3 Adrenergic Receptor Stimulation Promotes Reperfusion in Ischemic Limbs in a Murine Diabetic Model. Front. Pharmacol. 2021, 12, 666334. [Google Scholar] [CrossRef]
- Nuntapornsak, A.; Wongdee, K.; Thongbunchoo, J.; Krishnamra, N.; Charoenphandhu, N. Changes in the mRNA expression of osteoblast-related genes in response to β3-adrenergic agonist in UMR106 cells. Cell Biochem. Funct. 2010, 28, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Lambert, E.; Phillips, S.; Tursunalieva, A.; Eikelis, N.; Sari, C.; Dixon, J.; Straznicky, N.; Grima, M.; Schlaich, M.; Lambert, G. Inverse association between sympathetic nervous system activity and bone mass in middle aged overweight individuals. Bone 2018, 111, 123–128. [Google Scholar] [CrossRef]
- Koeck, F.X.; Bobrik, V.; Fassold, A.; Grifka, J.; Kessler, S.; Straub, R.H. Marked loss of sympathetic nerve fibers in chronic Charcot foot of diabetic origin compared to ankle joint osteoarthritis. J. Orthop. Res. 2009, 27, 736–741. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wagner, J.M.; Wallner, C.; Becerikli, M.; Reinkemeier, F.; von Glinski, M.; Sogorski, A.; Huber, J.; Dittfeld, S.; Becker, K.; Lehnhardt, M.; et al. Role of Autonomous Neuropathy in Diabetic Bone Regeneration. Cells 2022, 11, 612. https://doi.org/10.3390/cells11040612
Wagner JM, Wallner C, Becerikli M, Reinkemeier F, von Glinski M, Sogorski A, Huber J, Dittfeld S, Becker K, Lehnhardt M, et al. Role of Autonomous Neuropathy in Diabetic Bone Regeneration. Cells. 2022; 11(4):612. https://doi.org/10.3390/cells11040612
Chicago/Turabian StyleWagner, Johannes Maximilian, Christoph Wallner, Mustafa Becerikli, Felix Reinkemeier, Maxi von Glinski, Alexander Sogorski, Julika Huber, Stephanie Dittfeld, Kathrin Becker, Marcus Lehnhardt, and et al. 2022. "Role of Autonomous Neuropathy in Diabetic Bone Regeneration" Cells 11, no. 4: 612. https://doi.org/10.3390/cells11040612
APA StyleWagner, J. M., Wallner, C., Becerikli, M., Reinkemeier, F., von Glinski, M., Sogorski, A., Huber, J., Dittfeld, S., Becker, K., Lehnhardt, M., Dadras, M., & Behr, B. (2022). Role of Autonomous Neuropathy in Diabetic Bone Regeneration. Cells, 11(4), 612. https://doi.org/10.3390/cells11040612