
Establishment of an In Vitro Model to Study Viral Infections of the Fish Intestinal Epithelium


Abstract
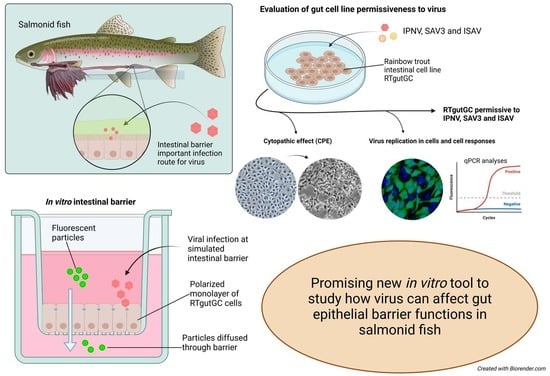
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Virus Propagation
2.3. Virus Infection Assays and Assessment of CPE in RTgutGC Cells
2.4. Immunofluorescence Antibody Technique (IFAT)
2.5. Quantitative Real-Time PCR (qPCR) Analyses of Virus Replication and Cellular Responses
2.6. Permeability Test with Fluorescent Bovine Serum Albumin (BSA)
2.7. Statistical Analyses
3. Results
3.1. Assessment of CPE
3.2. Detection of Virus Proteins in Infected RTgutGC Using IFAT
3.3. Correlation between Viral Replication and Induction of Antiviral Responses
3.4. Effects of Virus Infection on Epithelial Barrier Function
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Houston, R.D.; Gheyas, A.; Hamilton, A.; Guy, D.R.; Tinch, A.E.; Taggart, J.B.; McAndrew, B.J.; Haley, C.S.; Bishop, S.C. Detection and confirmation of a major QTL affecting resistance to infectious pancreatic necrosis (IPN) in Atlantic salmon (Salmo salar). Dev. Biol. 2008, 132, 199–204. [Google Scholar] [CrossRef]
- Aslam, M.L.; Robledo, D.; Krasnov, A.; Moghadam, H.K.; Hillestad, B.; Houston, R.D.; Baranski, M.; Boison, S.; Robinson, N.A. Quantitative trait loci and genes associated with salmonid alphavirus load in Atlantic salmon: Implications for pancreas disease resistance and tolerance. Sci. Rep. 2020, 10, 10393. [Google Scholar] [CrossRef]
- Sommerset, I.; Wiik-Nilsen, J.; Oliveira, V.H.S.; Moldal, T.; Bornø, G.; Haukaas, A.; Brun, E. Norwegian Fish Health Report 2022; Norwegian Veterinary Institute: Oslo, Norway, 2023. [Google Scholar]
- Turner, J.R. Intestinal mucosal barrier function in health and disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Neumann, N.F.; Stafford, J.L.; Barreda, D.; Ainsworth, A.J.; Belosevic, M. Antimicrobial mechanisms of fish phagocytes and their role in host defense. Dev. Comp. Immunol. 2001, 25, 807–825. [Google Scholar] [CrossRef]
- Smith, P.D.; MacDonald, T.T.; Blumberg, R.R.S. Principles of Mucosal Immunology; Garland Science: New York, NY, USA, 2013. [Google Scholar]
- Løkka, G.; Koppang, E.O. Antigen sampling in the fish intestine. Dev. Comp. Immunol. 2016, 64, 138–149. [Google Scholar] [CrossRef]
- Jung, S.-M.; Kim, S. In vitro models of the small intestine for studying intestinal diseases. Front. Microbiol. 2022, 12, 767038. [Google Scholar] [CrossRef] [PubMed]
- Kendall, L.V.; Owiny, J.R.; Dohm, E.D.; Knapek, K.J.; Lee, E.S.; Kopanke, J.H.; Fink, M.; Hansen, S.A.; Ayers, J.D. Replacement, refinement, and reduction in animal studies with biohazardous agents. ILAR J. 2018, 59, 177–194. [Google Scholar] [CrossRef]
- Kawano, A.; Haiduk, C.; Schimer, K.; Hanner, R.; Lee, L.E.J.; Dixon, B.; Bols, N.C. Development of a rainbow trout intestinal epithelial cell line and its response to lipopolysaccharide. Aquac. Nutr. 2011, 17, e241–e252. [Google Scholar] [CrossRef]
- Wang, J.; Lei, P.; Gamil, A.A.A.; Lagos, L.; Yue, Y.; Schirmer, K.; Mydland, L.T.; Øverland, M.; Krogdahl, Å.; Kortner, T.M. Rainbow trout (Oncorhynchus mykiss) intestinal epithelial cells as a model for studying gut immune function and effects of functional feed ingredients. Front. Immunol. 2019, 10, 152. [Google Scholar] [CrossRef]
- Minghetti, M.; Drieschner, C.; Bramaz, N.; Schug, H.; Schirmer, K. A fish intestinal epithelial barrier model established from the rainbow trout (Oncorhynchus mykiss) cell line, RTgutGC. Cell Biol. Toxicol. 2017, 33, 539–555. [Google Scholar] [CrossRef]
- Geppert, M.; Sigg, L.; Schirmer, K. A novel two-compartment barrier model for investigating nanoparticle transport in fish intestinal epithelial cells. Environ. Sci. Nano 2016, 3, 388–395. [Google Scholar] [CrossRef]
- Pasquariello, R.; Verdile, N.; Pavlovic, R.; Panseri, S.; Schirmer, K.; Brevini, T.A.L.; Gandolfi, F. New stable cell lines derived from the proximal and distal intestine of rainbow trout (Oncorhynchus mykiss) retain several properties observed in vivo. Cells 2021, 10, 1555. [Google Scholar] [CrossRef]
- Schug, H.; Yue, Y.; Krese, R.; Fischer, S.; Kortner, T.M.; Schirmer, K. Time- and concentration-dependent expression of immune and barrier genes in the RTgutGC fish intestinal model following immune stimulation. Fish Shellfish Immunol. 2019, 88, 308–317. [Google Scholar] [CrossRef]
- Prabhu, P.A.J.; Stewart, T.; Silva, M.; Amlund, H.; Ørnsrud, R.; Lock, E.J.; Waagbo, R.; Hogstrand, C. Zinc uptake in fish intestinal epithelial model RTgutGC: Impact of media ion composition and methionine chelation. J. Trace Elem. Med. Biol. 2018, 50, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Langan, L.M.; Harper, G.M.; Owen, S.F.; Purcell, W.M.; Jackson, S.K.; Jha, A.N. Application of the rainbow trout derived intestinal cell line (RTgutGC) for ecotoxicological studies: Molecular and cellular responses following exposure to copper. Ecotoxicology 2017, 26, 1117–1133. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, P.; Wacyk, J.; Morales-Lange, B.; Rojas, V.; Guzmán, F.; Dixon, B.; Mercado, L. Immunomodulatory effect of cathelicidins in response to a β-glucan in intestinal epithelial cells from rainbow trout. Dev. Comp. Immunol. 2015, 51, 160–169. [Google Scholar] [CrossRef]
- Santi, N.; Vakharia, V.N.; Evensen, Ø. Identification of putative motifs involved in the virulence of infectious pancreatic necrosis virus. Virology 2004, 322, 31–40. [Google Scholar] [CrossRef]
- Guo, T.C.; Johansson, D.X.; Haugland, Ø.; Liljeström, P.; Evensen, Ø. A 6K-deletion variant of salmonid alphavirus is non-viable but can be rescued through RNA recombination. PLoS ONE 2014, 9, e100184. [Google Scholar] [CrossRef]
- Godoy, M.G.; Kibenge, M.J.T.; Suarez, R.; Lazo, E.; Heisinger, A.; Aguinaga, J.; Bravo, D.; Mendoza, J.; Llegues, K.O.; Avendaño-Herrera, R.; et al. Infectious salmon anaemia virus (ISAV) in Chilean Atlantic salmon (Salmo salar) aquaculture: Emergence of low pathogenic ISAV-HPR0 and re-emergence of virulent ISAV-HPR∆: HPR3 and HPR14. Virol. J. 2013, 10, 344. [Google Scholar] [CrossRef]
- Kärber, G. Beitrag zur kollektiven Behandlung pharmakologischer Reihenversuche. Naunyn-Schmiedebergs Arch. Für Exp. Pathol. Pharmakol. 1931, 162, 480–483. [Google Scholar] [CrossRef]
- Evensen, Ø.; Rimstad, E. Immunohistochemical identification of infectious pancreatic necrosis virus in paraffin-embedded tissues of Atlantic salmon (Salmo salar). J. Vet. Diagn. Investig. 1990, 2, 288–293. [Google Scholar] [CrossRef]
- Xu, C.; Guo, T.-C.; Mutoloki, S.; Haugland, Ø.; Marjara, I.S.; Evensen, Ø. Alpha interferon and not gamma interferon inhibits salmonid alphavirus subtype 3 replication in vitro. J. Virol. 2010, 84, 8903–8912. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Pearce, S.C.; Coia, H.G.; Karl, J.P.; Pantoja-Feliciano, I.G.; Zachos, N.C.; Racicot, K. Intestinal in vitro and ex vivo models to study host-microbiome interactions and acute stressors. Front. Physiol. 2018, 9, 1584. [Google Scholar] [CrossRef] [PubMed]
- Verdile, N.; Pasquariello, R.; Brevini, T.A.L.; Gandolfi, F. The 3D pattern of the rainbow trout (Oncorhynchus mykiss) enterocytes and intestinal stem cells. Int. J. Mol. Sci. 2020, 21, 9192. [Google Scholar] [CrossRef]
- Bussolaro, D.; Wright, S.L.; Schnell, S.; Schirmer, K.; Bury, N.R.; Arlt, V.M. Co-exposure to polystyrene plastic beads and polycyclic aromatic hydrocarbon contaminants in fish gill (RTgill-W1) and intestinal (RTgutGC) epithelial cells derived from rainbow trout (Oncorhynchus mykiss). Environ. Pollut. 2019, 248, 706–714. [Google Scholar] [CrossRef]
- Dayeh, V.R.; Bols, N.C.; Tanneberger, K.; Schirmer, K.; Lee, L.E. The use of fish-derived cell lines for investigation of environmental contaminants: An update following OECD’s fish toxicity testing framework No. 171. Curr. Protoc. Toxicol. 2013, 56, 1.5.1–1.5.20. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.; Minghetti, M. Effect of chloride concentration on the cytotoxicity, bioavailability, and bioreactivity of copper and silver in the rainbow trout gut cell line, RTgutGC. Ecotoxicology 2022, 31, 626–636. [Google Scholar] [CrossRef] [PubMed]
- Langan, L.M.; Arossa, S.; Owen, S.F.; Jha, A.N. Assessing the impact of benzo[a]pyrene with the in vitro fish gut model: An integrated approach for eco-genotoxicological studies. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2018, 826, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Oldham, D.; Dudefoi, W.; Minghetti, M. Development of an in vitro digestion system to study the bioavailability and bioreactivity of zinc sulfate and Zn-bioplex in fish using the RTgutGC cell line. ACS Food Sci. Technol. 2023, 3, 141–149. [Google Scholar] [CrossRef]
- Schug, H.; Maner, J.; Hülskamp, M.; Begnaud, F.; Debonneville, C.; Berthaud, F.; Gimeno, S.; Schirmer, K. Extending the concept of predicting fish acute toxicity in vitro to the intestinal cell line RTgutGC. Altex 2020, 37, 37–46. [Google Scholar] [CrossRef]
- Doherty, L.; Poynter, S.J.; Aloufi, A.; DeWitte-Orr, S.J. Fish viruses make dsRNA in fish cells: Characterization of dsRNA production in rainbow trout (Oncorhynchus mykiss) cells infected with viral haemorrhagic septicaemia virus, chum salmon reovirus and frog virus 3. J. Fish Dis. 2016, 39, 1133–1137. [Google Scholar] [CrossRef] [PubMed]
- Nerbøvik, I.G.; Solheim, M.A.; Eggestøl, H.; Rønneseth, A.; Jakobsen, R.A.; Wergeland, H.I.; Haugland, G.T. Molecular cloning of MDA5, phylogenetic analysis of RIG-I-like receptors (RLRs) and differential gene expression of RLRs, interferons and proinflammatory cytokines after in vitro challenge with IPNV, ISAV and SAV in the salmonid cell line TO. J. Fish Dis. 2017, 40, 1529–1544. [Google Scholar] [CrossRef] [PubMed]
- Sadasiv, E.C. Immunological and pathological responses of salmonids to infectious pancreatic necrosis virus (IPNV). Annu. Rev. Fish Dis. 1995, 5, 209–223. [Google Scholar] [CrossRef]
- Sundh, H.; Calabrese, S.; Jutfelt, F.; Niklasson, L.; Olsen, R.-E.; Olsen, R.-E.; Sundell, K. Translocation of infectious pancreatic necrosis virus across the intestinal epithelium of Atlantic salmon (Salmo salar L.). Aquaculture 2011, 321, 85–92. [Google Scholar] [CrossRef]
- Chen, L.; Evensen, Ø.; Mutoloki, S. IPNV antigen uptake and distribution in Atlantic salmon following oral administration. Viruses 2015, 7, 2507–2517. [Google Scholar] [CrossRef] [PubMed]
- Gamil, A.A.A.; Evensen, Ø.; Mutoloki, S. Infection profiles of selected Aquabirnavirus isolates in CHSE cells. PLoS ONE 2015, 10, e0134173. [Google Scholar] [CrossRef] [PubMed]
- Gamil, A.A.A.; Mutoloki, S.; Evensen, Ø. A piscine birnavirus induces inhibition of protein synthesis in CHSE-214 cells primarily through the induction of eIF2α phosphorylation. Viruses 2015, 7, 1987–2005. [Google Scholar] [CrossRef]
- Gamil, A.A.A.; Xu, C.; Mutoloki, S.; Evensen, Ø. PKR activation favors infectious pancreatic necrosis virus replication in infected cells. Viruses 2016, 8, 173. [Google Scholar] [CrossRef]
- Collet, B.; Munro, E.S.; Gahlawat, S.; Acosta, F.; Garcia, J.; Roemelt, C.; Zou, J.; Secombes, C.J.; Ellis, A.E. Infectious pancreatic necrosis virus suppresses type I interferon signalling in rainbow trout gonad cell line but not in Atlantic salmon macrophages. Fish Shellfish Immunol. 2007, 22, 44–56. [Google Scholar] [CrossRef]
- Gahlawat, S.K.; Ellis, A.E.; Collet, B. Expression of interferon and interferon-induced genes in Atlantic salmon Salmo salar cell lines SHK-1 and TO following infection with Salmon AlphaVirus SAV. Fish Shellfish Immunol. 2009, 26, 672–675. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Gamil, A.A.A.; Gadan, K.; Mikalsen, A.B.; Evensen, Ø. Antiviral innate immune responses: Infectious pancreatic necrosis virus and salmonid alphavirus. In Principles of Fish Immunology: From Cells and Molecules to Host Protection; Buchmann, K., Secombes, C.J., Eds.; Springer International Publishing: Cham, Switzerland, 2022; pp. 463–494. [Google Scholar] [CrossRef]
- Mandal, S.C.; Weidmann, M.; Albalat, A.; Carrick, E.; Morro, B.; MacKenzie, S. Polarized trout epithelial cells regulate transepithelial electrical resistance, gene expression, and the phosphoproteome in response to viral infection. Front. Immunol. 2020, 11, 1809. [Google Scholar] [CrossRef] [PubMed]
- Jami, R.; Mérour, E.; Bernard, J.; Lamoureux, A.; Millet, J.K.; Biacchesi, S. The C-terminal domain of salmonid alphavirus nonstructural protein 2 (nsP2) is essential and sufficient to block RIG-I pathway induction and interferon-mediated antiviral response. J. Virol. 2021, 95, e0115521. [Google Scholar] [CrossRef]
- Mikalsen, A.B.; Teig, A.; Helleman, A.L.; Mjaaland, S.; Rimstad, E. Detection of infectious salmon anaemia virus (ISAV) by RT-PCR after cohabitant exposure in Atlantic salmon Salmo salar. Dis. Aquat. Org. 2001, 47, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Aamelfot, M.; McBeath, A.; Christiansen, D.H.; Matejusova, I.; Falk, K. Infectious salmon anaemia virus (ISAV) mucosal infection in Atlantic salmon. Vet. Res. 2015, 46, 120. [Google Scholar] [CrossRef]
- Weli, S.C.; Aamelfot, M.; Dale, O.B.; Koppang, E.O.; Falk, K. Infectious salmon anaemia virus infection of Atlantic salmon gill epithelial cells. J. Virol. 2013, 10, 5. [Google Scholar] [CrossRef]
- Aamelfot, M.; Dale, O.B.; Weli, S.C.; Koppang, E.O.; Falk, K. Expression of the infectious salmon anemia virus receptor on atlantic salmon endothelial cells correlates with the cell tropism of the virus. J. Virol. 2012, 86, 10571–10578. [Google Scholar] [CrossRef]
- Aamelfot, M.; Weli, S.C.; Dale, O.B.; Koppang, E.O.; Falk, K. Characterisation of a monoclonal antibody detecting Atlantic salmon endothelial and red blood cells, and its association with the infectious salmon anaemia virus cell receptor. J. Anat. 2013, 222, 547–557. [Google Scholar] [CrossRef]
- Poynter, S.J.; Lisser, G.R.J.; Monjo, A.L.-A.; DeWitte-Orr, S.J. Sensors of infection: Viral nucleic acid PRRs in fish. Biology 2015, 4, 460–493. [Google Scholar] [CrossRef]
- Poynter, S.J.; DeWitte-Orr, S.J. Fish interferon-stimulated genes: The antiviral effectors. Dev. Comp. Immunol. 2016, 65, 218–225. [Google Scholar] [CrossRef]
- Poynter, S.J.; DeWitte-Orr, S.J. Understanding viral dsRNA-mediated innate immune responses at the cellular level using a rainbow trout model. Front. Immunol. 2018, 9, 829. [Google Scholar] [CrossRef] [PubMed]
- Poynter, S.J.; DeWitte-Orr, S.J. Length-dependent innate antiviral effects of double-stranded RNA in the rainbow trout (Oncorhynchus mykiss) cell line, RTG-2. Fish Shellfish Immunol. 2015, 46, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Rombout, J.H.; Abelli, L.; Picchietti, S.; Scapigliati, G.; Kiron, V. Teleost intestinal immunology. Fish Shellfish Immunol. 2011, 31, 616–626. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Løkka, G.; Gamil, A.A.A.; Evensen, Ø.; Kortner, T.M. Establishment of an In Vitro Model to Study Viral Infections of the Fish Intestinal Epithelium. Cells 2023, 12, 1531. https://doi.org/10.3390/cells12111531
Løkka G, Gamil AAA, Evensen Ø, Kortner TM. Establishment of an In Vitro Model to Study Viral Infections of the Fish Intestinal Epithelium. Cells. 2023; 12(11):1531. https://doi.org/10.3390/cells12111531
Chicago/Turabian StyleLøkka, Guro, Amr A. A. Gamil, Øystein Evensen, and Trond M. Kortner. 2023. "Establishment of an In Vitro Model to Study Viral Infections of the Fish Intestinal Epithelium" Cells 12, no. 11: 1531. https://doi.org/10.3390/cells12111531
APA StyleLøkka, G., Gamil, A. A. A., Evensen, Ø., & Kortner, T. M. (2023). Establishment of an In Vitro Model to Study Viral Infections of the Fish Intestinal Epithelium. Cells, 12(11), 1531. https://doi.org/10.3390/cells12111531