

In Vitro Human Haematopoietic Stem Cell Expansion and Differentiation


Abstract
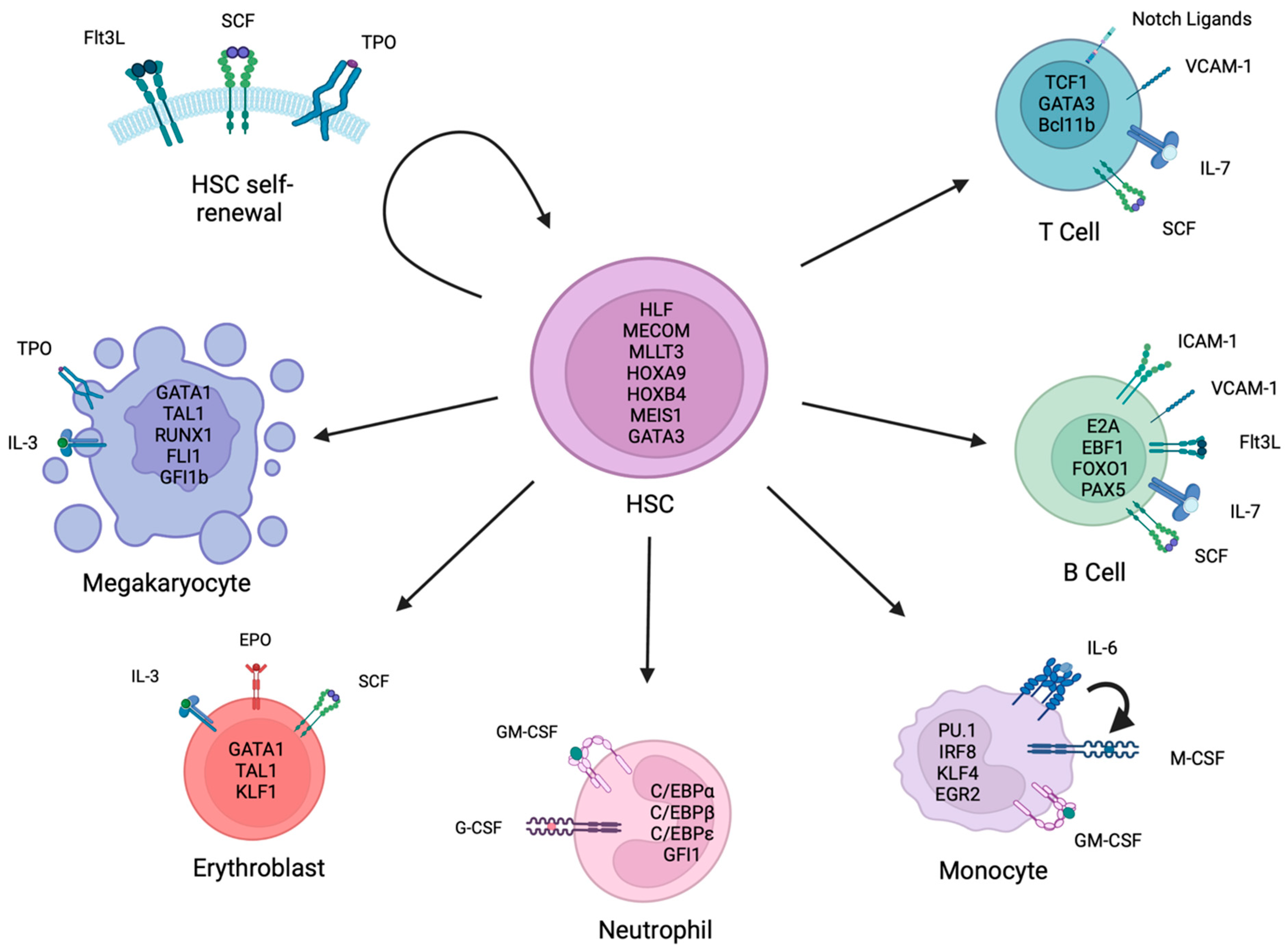
:1. Introduction
2. In Vitro Maintenance and Expansion of HSPCs
2.1. Signalling Pathways
2.2. Small-Molecules

{kind=link}
{kind=link}
| Compound | Activity | Expansion Effect on HSPC Culture | Refs. |
|---|---|---|---|
| TEPA | Copper chelator | 30.5-fold CD34+CD38− HSPCs 172-fold CFU activity Improved engraftment in xenograft transplants | [78] |
| 219-fold total nucleated cells 6-fold CD34+ HSPCs 37.8-fold CFU activity | [81] | ||
| 400-fold total nucleated cells 77-fold CD34+ HSPCs | [82] | ||
| SR1 | AHR | 50-fold CD34+ HSPCs 17-fold SRCs | [6] |
| 854-fold total nucleated cells 330-fold CD34+ HSPCs | [83] | ||
| NAM | SIRT1 | 80-fold CD34+ HSPCs 9-fold SRCs | [84] |
| 486-fold total nucleated cells 72-fold CD34+ HSPCs | [85] | ||
| 33-fold CD34+ HSPCs | [86] | ||
| dmPGE2 | 15-PGDH | 1.4-fold CFU activity Improved engraftment in xenograft transplants 2.2-fold increase in BM homing | [90,91] |
| UM171 | LSD1 | 13.4-fold SRCs | [94] |
| 35.4-fold CD34+ HSPCs Improved engraftment in xenograft transplants | [96] | ||
| NR101 | THPO | 4.9-fold CD34+CD38− HSPCs 2.9-fold SRCs | [98] |
| Eltrombopag | THPO/Iron Chelator | 1.42-fold CD34+CD38− HSPCs | [99] |
| C7 | p38 | 1554-fold CD34+CD38−CD45+CD45RA− HSPCs 2.5-fold SRCs | [103] |
| JNK-IN-8 | JNK | 8-fold CD34+CD38−CD45RA−CD90+ HSPCs 3.88-fold SRCs | [104] |
| 005A | p18 | 2.72-fold CFU activity Improved engraftment in xenograft transplants | [110] |
| VPA | HDAC | 194.7-fold CD34+CD45+ HSPCs 6-fold SRCs | [116] |
| VPA | HDAC | 213-fold CD34+ HSPCs 36-fold SRCs | [115] |
| Garcinol/ isogarcinol | HATs | 4.5-/7.4-fold CD34+CD38− HSPCs 2.5-fold SRCs (garcinol) | [117] |
| 5azaD | DNMT | 12.5-fold CD34+CD90+ HSPCs 9.6-fold SRCs | [118] |
| TSA | HDAC | ||
| CPI-203 | BET | 5–10-fold Lin−CD34+CD38−CD45RA−CD90+CD49f+ HSPCs 1.5–3-fold SRCs | [121] |
| BIO | GSK-3 | 2-fold CFU activity | [124] |
| CHIR99021 | GSK-3 | 7-fold total nucleated cells 5-fold SRCs | [125] |
| Rapamycin | mTOR |
3. In Vitro Differentiation to Megakaryocytes
4. In Vitro Differentiation to Erythrocytes
5. In Vitro Differentiation into Myeloid Cells
5.1. Neutrophils
| Cell Source | Method | Cells Generated | Reference |
|---|---|---|---|
| UCB CD34+ | THPO, FLT3L, G-CSF | 6–23% CD16+ (CD32hiCD64hi) | [63] |
| UCB or BM CD34+ | IL-3, GM-CSF, G-CSF, M-CSF | 100× expansion, 10–70% CD33+, <20% CD14/CD15+ | [236] |
| PB CD34+ PBMCs | SCF, IL-3, GM-CSF, G-CSF | 130–220× expansion, by morphology: 65% granulocytic, 11% band/segmented neutrophils, 5% monocyte/macrophages | [231] |
| UCB CD34+ HSPCs | SCF, IL-3, FLT3L, G-CSF | 60–70% CD15+, 75% MPO+ | [230] |
| G-CSF mobilised PB CD34+ | SCF, FLT3L, G-CSF | 30× expansion, 80% mature neutrophils (by morphology), CD16blo, | [238] |
| BM CD34+ | SCF, THPO, IL-3, G-CSF | 76% band/segmented neutrophils, CD15+CD66+ | [259] |
| UCB CD34+ | SCF, G-CSF, THPO mimetic | 5800× expansion, 61% metamyelocytes/band/segmented neutrophils, 73% CD15+ | [237] |
| UCB CD34+ | Phase I: SCF, IL-3, FLT3L, GM-CSF; phase II: G-CSF | 8900–49,000× expansion, 59% CD66b+ | [232] |
| UCB CD34+ | SCF, FLT3L, IL3, THPO, EPO, with MS-5 stromal cells | Pan-myeloid differentiation, 23% CD34+, 12% CD14+ monocytes, 5% CD66b+ granulocytes, 8% CD41+ megakaryocytes, 11% CD235a+ erythrocytes | [240] |
| UCB or PB CD34+ | Staged combinations of SCF, IL-3, FLT3L, GM-CSF, G-CSF | 50–70× expansion, 70–92% CD15+, 43–57% CD66b+ | [233] |
5.2. Monocytes
| Cell Source | Method | Cells Generated | Reference |
|---|---|---|---|
| G-CSF mobilised PB CD34+ | M-CSF, mast cell growth factor (MGF), IL-6 | No expansion, 55% CD33+CD14+, 62% CD33+HLA-DR+ | [270] |
| G-CSF mobilised PB CD34+ | SCF and IL-2 | Majority CD33+, 2.5% CD33+CD56dim, NK-like monocytes | [273] |
| UCB CD34+ | SCF, IL-3, FLT3L and M-CSF | 300× expansion, 45% CD14+, 22% CD16+ | [272] |
| G-CSF mobilised PB CD34+ | SCF, FLT3L, IL-3, IL-6, GM-CSF, M-CSF with 0.5% buminate | 360× expansion, 90% CD11b+, 65% CD64+, 45% CD14+ | [278] |
| G-CSF mobilised PB | SCF, FLT3L, IL-3, IL-6 M-CSF with 1% Buminate | 90% lysozyme positive, CD14+CD64+CD16+HLA-DR+ | [271] |
| UCB CD34+ | SCF, IL-6, FLT3L or SCF, THPO and FLT3L and viral transduction of M-CSF | 350× expansion, 70–80% CD33+CD14+ | [283] |
6. In Vitro Differentiation into Lymphoid Cells
6.1. B Cells
| Cell Source | Method | Cytokines | Cell Generated | References |
|---|---|---|---|---|
| Nucleated BM cells | BM fibroblast feeder layer | None | CD10+/CD20+ B-lineage cells | [315] |
| UCB CD34+ | Murine fetal stromal feeder cell layer (MS-5) with cytokines | SCF, G-CSF | CD19+/IgM+ | [316] |
| BM CD34+Lineage−CD38− | Murine fetal stromal feeder cell layer (AFT024) with cytokines | IL-2, SCF, FLT3L, IL-7, IL-3 | CD10+/CD19+ B-lineage cells | [317] |
| Fetal BM CD34+Lineage− | Human fetal BM stromal cells | None | IgM+ immature B cells | [318] |
| UCB CD34+ | Human BM stromal cell layer with cytokines (and additional antibodies) | SCF, FLT3L; (anti-Activin A and anti-TGFβ antibodies) | IgM+ immature B cells | [314] |
| UCB CD34+ | Murine stromal cell layer (S17) | Il-10, IL-4, FLT3L, IL-2 | IgM/IgG-secreting B cells | [319,320] |
| Fetal liver CD34+ | Murine stromal cell (OP9) with cytokines | TSLP | Mature CD34−CD38+CD19+IgM+IgD+ B cells | [321] |
| UCB or BM CD34+ | ICAM-1 coated plate with cytokines | IL-6, SCF, FLT3L, IL-7 | IgM+ CD19+ immature B cells | [301] |
| UCB CD34+ | Cytokines only | IL-6, FLT3L, SCF, IL-7 | CD10+CD79α+CD19+ pro-B cells | [322] |
6.2. T Cells
| Cell Source | Method | Cytokines | Cell Generated | Reference |
|---|---|---|---|---|
| UCB CD34+CD38− | OP9-DLL1 stromal feeder cell with cytokines | FLT3L, IL-7 | CD4+CD8+ DP T cells | [333] |
| UCB CD34+CD38−CD3−CD19− | Culture on MS5 then transfer to DLL4-coated plate | IL-2, IL-15, SCF | CD7+CD3+ | [356] |
| UCB CD34+CD38− | Murine stromal feeder layer (Tst-4/hDLL1) | None | CD5+CD7+ immature T cells | [336] |
| UCB CD34+ | DLL4-coated plate with cytokines | SCF, THPO, FLT3L, IL-7 | CD5+CD7+CD1a+ immature T cells | [345] |
| UCB CD34+ | DLL4- and VCAM-1 coated plate with cytokines | SCF, THPO, FLT3L, IL-7 | CD7+ pro-T cells | [349] |
| UCB CD34+ | DLL4- and VCAM-1 coated plate with cytokines | SCF, THPO, FLT3L, IL-7, IL-3, TNFa | CD4+CD8+ DP T cells | [351] |
| UCB and BM CD34+ | ATO system with cytokines | FLT3L, IL-7 | CD4+ and CD8+ mature SP T cells | [342] |
| UCB CD34+ | DLL4-coated microbeads with cytokines | FLT3L, SCF, IL-7 | CD7+CD5+ immature T cells | [350] |
7. Summary
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Boulais, P.E.; Frenette, P.S. Making Sense of Hematopoietic Stem Cell Niches. Blood 2015, 125, 2621–2629. [Google Scholar] [CrossRef] [Green Version]
- Pinho, S.; Frenette, P.S. Haematopoietic Stem Cell Activity and Interactions with the Niche. Nat. Rev. Mol. Cell Biol. 2019, 20, 303–320. [Google Scholar] [CrossRef] [PubMed]
- Morrison, S.J.; Scadden, D.T. The Bone Marrow Niche for Haematopoietic Stem Cells. Nature 2014, 505, 327–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Notta, F.; Doulatov, S.; Laurenti, E.; Poeppl, A.; Jurisica, I.; Dick, J.E. Isolation of Single Human Hematopoietic Stem Cells Capable of Long-Term Multilineage Engraftment. Science 2011, 333, 218–221. [Google Scholar] [CrossRef]
- Laurenti, E.; Göttgens, B. From Haematopoietic Stem Cells to Complex Differentiation Landscapes. Nature 2018, 553, 418–426. [Google Scholar] [CrossRef]
- Boitano, A.E.; Wang, J.; Romeo, R.; Bouchez, L.C.; Parker, A.E.; Sutton, S.E.; Walker, J.R.; Flaveny, C.A.; Perdew, G.H.; Denison, M.S.; et al. Aryl Hydrocarbon Receptor Antagonists Promote the Expansion of Human Hematopoietic Stem Cells. Science 2010, 329, 1345–1348. [Google Scholar] [CrossRef] [Green Version]
- Eaves, C.J. Hematopoietic Stem Cells: Concepts, Definitions, and the New Reality. Blood 2015, 125, 2605–2613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehnertz, B.; Chagraoui, J.; MacRae, T.; Tomellini, E.; Corneau, S.; Mayotte, N.; Boivin, I.; Durand, A.; Gracias, D.; Sauvageau, G. HLF Expression Defines the Human Hematopoietic Stem Cell State. Blood 2021, 138, 2642–2654. [Google Scholar] [CrossRef]
- Christodoulou, C.; Spencer, J.A.; Yeh, S.-C.A.; Turcotte, R.; Kokkaliaris, K.D.; Panero, R.; Ramos, A.; Guo, G.; Seyedhassantehrani, N.; Esipova, T.V.; et al. Live-Animal Imaging of Native Haematopoietic Stem and Progenitor Cells. Nature 2020, 578, 278–283. [Google Scholar] [CrossRef]
- Calvanese, V.; Nguyen, A.T.; Bolan, T.J.; Vavilina, A.; Su, T.; Lee, L.K.; Wang, Y.; Lay, F.D.; Magnusson, M.; Crooks, G.M.; et al. MLLT3 Governs Human Haematopoietic Stem-Cell Self-Renewal and Engraftment. Nature 2019, 576, 281–286. [Google Scholar] [CrossRef]
- Xiang, P.; Wei, W.; Hofs, N.; Clemans-Gibbon, J.; Maetzig, T.; Lai, C.K.; Dhillon, I.; May, C.; Ruschmann, J.; Schneider, E.; et al. A Knock-in Mouse Strain Facilitates Dynamic Tracking and Enrichment of MEIS1. Blood Adv. 2017, 1, 2225–2235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frelin, C.; Herrington, R.; Janmohamed, S.; Barbara, M.; Tran, G.; Paige, C.J.; Benveniste, P.; Zuñiga-Pflücker, J.-C.; Souabni, A.; Busslinger, M.; et al. GATA-3 Regulates the Self-Renewal of Long-Term Hematopoietic Stem Cells. Nat. Immunol. 2013, 14, 1037–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawrence, H.J.; Christensen, J.; Fong, S.; Hu, Y.-L.; Weissman, I.; Sauvageau, G.; Humphries, R.K.; Largman, C. Loss of Expression of the Hoxa-9 Homeobox Gene Impairs the Proliferation and Repopulating Ability of Hematopoietic Stem Cells. Blood 2005, 106, 3988–3994. [Google Scholar] [CrossRef] [Green Version]
- Buske, C. Deregulated Expression of HOXB4 Enhances the Primitive Growth Activity of Human Hematopoietic Cells. Blood 2002, 100, 862–868. [Google Scholar] [CrossRef] [PubMed]
- Sauvageau, G.; Thorsteinsdottir, U.; Eaves, C.J.; Lawrence, H.J.; Largman, C.; Lansdorp, P.M.; Humphries, R.K. Overexpression of HOXB4 in Hematopoietic Cells Causes the Selective Expansion of More Primitive Populations in Vitro and in Vivo. Genes Dev. 1995, 9, 1753–1765. [Google Scholar] [CrossRef] [Green Version]
- Staal, F.J.T.; Chhatta, A.; Mikkers, H. Caught in a Wnt Storm: Complexities of Wnt Signaling in Hematopoiesis. Exp. Hematol. 2016, 44, 451–457. [Google Scholar] [CrossRef]
- Nusse, R.; Clevers, H. Wnt/β-Catenin Signaling, Disease, and Emerging Therapeutic Modalities. Cell 2017, 169, 985–999. [Google Scholar] [CrossRef] [Green Version]
- Pajcini, K.V.; Speck, N.A.; Pear, W.S. Notch Signaling in Mammalian Hematopoietic Stem Cells. Leukemia 2011, 25, 1525–1532. [Google Scholar] [CrossRef] [Green Version]
- Siebel, C.; Lendahl, U. Notch Signaling in Development, Tissue Homeostasis, and Disease. Physiol. Rev. 2017, 97, 1235–1294. [Google Scholar] [CrossRef] [Green Version]
- Blank, U.; Karlsson, G.; Karlsson, S. Signaling Pathways Governing Stem-Cell Fate. Blood 2008, 111, 492–503. [Google Scholar] [CrossRef]
- Reya, T.; Duncan, A.W.; Ailles, L.; Domen, J.; Scherer, D.C.; Willert, K.; Hintz, L.; Nusse, R.; Weissman, I.L. A Role for Wnt Signalling in Self-Renewal of Haematopoietic Stem Cells. Nature 2003, 423, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Helgason, C.D.; Antonchuk, J.; Bodner, C.; Humphries, R.K. Homeostasis and Regeneration of the Hematopoietic Stem Cell Pool Are Altered in SHIP-Deficient Mice. Blood 2003, 102, 3541–3547. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, M.; Bonnet, D.; Kapp, U.; Wang, J.C.Y.; Murdoch, B.; Dick, J.E. Quantitative Analysis Reveals Expansion of Human Hematopoietic Repopulating Cells after Short-Term Ex Vivo Culture. J. Exp. Med. 1997, 186, 619–624. [Google Scholar] [CrossRef] [PubMed]
- Conneally, E.; Cashman, J.; Petzer, A.; Eaves, C. Expansion in Vitro of Transplantable Human Cord Blood Stem Cells Demonstrated Using a Quantitative Assay of Their Lympho-Myeloid Repopulating Activity in Nonobese Diabetic– Scid/Scid Mice. Proc. Natl. Acad. Sci. USA 1997, 94, 9836–9841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walasek, M.A.; van Os, R.; de Haan, G. Hematopoietic Stem Cell Expansion: Challenges and Opportunities: HSC Expansion: Challenges and Opportunities. Ann. N. Y. Acad. Sci. 2012, 1266, 138–150. [Google Scholar] [CrossRef] [Green Version]
- Himburg, H.A.; Termini, C.M.; Schlussel, L.; Kan, J.; Li, M.; Zhao, L.; Fang, T.; Sasine, J.P.; Chang, V.Y.; Chute, J.P. Distinct Bone Marrow Sources of Pleiotrophin Control Hematopoietic Stem Cell Maintenance and Regeneration. Cell Stem Cell 2018, 23, 370–381.e5. [Google Scholar] [CrossRef] [Green Version]
- Himburg, H.A.; Yan, X.; Doan, P.L.; Quarmyne, M.; Micewicz, E.; McBride, W.; Chao, N.J.; Slamon, D.J.; Chute, J.P. Pleiotrophin Mediates Hematopoietic Regeneration via Activation of RAS. J. Clin. Investig. 2014, 124, 4753–4758. [Google Scholar] [CrossRef] [Green Version]
- Himburg, H.A.; Harris, J.R.; Ito, T.; Daher, P.; Russell, J.L.; Quarmyne, M.; Doan, P.L.; Helms, K.; Nakamura, M.; Fixsen, E.; et al. Pleiotrophin Regulates the Retention and Self-Renewal of Hematopoietic Stem Cells in the Bone Marrow Vascular Niche. Cell Rep. 2012, 2, 964–975. [Google Scholar] [CrossRef] [Green Version]
- Himburg, H.A.; Muramoto, G.G.; Daher, P.; Meadows, S.K.; Russell, J.L.; Doan, P.; Chi, J.-T.; Salter, A.B.; Lento, W.E.; Reya, T.; et al. Pleiotrophin Regulates the Expansion and Regeneration of Hematopoietic Stem Cells. Nat. Med. 2010, 16, 475–482. [Google Scholar] [CrossRef]
- Santulli, G. Angiopoietin-Like Proteins: A Comprehensive Look. Front. Endocrinol. 2014, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Bhardwaj, G.; Murdoch, B.; Wu, D.; Baker, D.P.; Williams, K.P.; Chadwick, K.; Ling, L.E.; Karanu, F.N.; Bhatia, M. Sonic Hedgehog Induces the Proliferation of Primitive Human Hematopoietic Cells via BMP Regulation. Nat. Immunol. 2001, 2, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Bai, T.; Li, J.; Sinclair, A.; Imren, S.; Merriam, F.; Sun, F.; O’Kelly, M.B.; Nourigat, C.; Jain, P.; Delrow, J.J.; et al. Expansion of Primitive Human Hematopoietic Stem Cells by Culture in a Zwitterionic Hydrogel. Nat. Med. 2019, 25, 1566–1575. [Google Scholar] [CrossRef]
- Williams, D.E.; Eisenman, J.; Baird, A.; Rauch, C.; Van Ness, K.; March, C.J.; Park, L.S.; Martin, U.; Mochizukl, D.Y.; Boswell, H.S.; et al. Identification of a Ligand for the C-Kit Proto-Oncogene. Cell 1990, 63, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.H.; Suggs, S.V.; Langley, K.E.; Lu, H.S.; Ting, J.; Okino, K.H.; Morris, C.F.; McNiece, I.K.; Jacobsen, F.W.; Mendlaz, E.A.; et al. Primary Structure and Functional Expression of Rat and Human Stem Cell Factor DNAs. Cell 1990, 63, 203–211. [Google Scholar] [CrossRef]
- Zsebo, K.M.; Williams, D.A.; Geissler, E.N.; Broudy, V.C.; Martin, F.H.; Atkins, H.L.; Hsu, R.-Y.; Birkett, N.C.; Okino, K.H.; Murdock, D.C.; et al. Stem Cell Factor Is Encoded at the SI Locus of the Mouse and Is the Ligand for the C-Kit Tyrosine Kinase Receptor. Cell 1990, 63, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, I.D.; Andrews, R.G.; Zsebo, K.M. Recombinant Human Stem Cell Factor Enhances the Formation of Colonies by CD34+ and CD34+lin- Cells, and the Generation of Colony-Forming Cell Progeny From CD34+lin- Cells Cultured With Interleukin-3, Granulocyte Colony-Stimulating Factor, or Granulocyte-Macrophage Colony-Stimulating Factor. Blood 1991, 77, 2316–2321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duarte, R.F.; Frank, D.A. The Synergy Between Stem Cell Factor (SCF) and Granulocyte Colony-Stimulating Factor (G-CSF): Molecular Basis and Clinical Relevance. Leuk. Lymphoma 2002, 43, 1179–1187. [Google Scholar] [CrossRef]
- Ding, L.; Saunders, T.L.; Enikolopov, G.; Morrison, S.J. Endothelial and Perivascular Cells Maintain Haematopoietic Stem Cells. Nature 2012, 481, 457–462. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, S.; Iwama, A.; Takayanagi, S.; Morita, Y.; Eto, K.; Ema, H.; Nakauchi, H. Cytokine Signals Modulated via Lipid Rafts Mimic Niche Signals and Induce Hibernation in Hematopoietic Stem Cells. EMBO J. 2006, 25, 3515–3523. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, K.; Araki, K.Y.; Naka, K.; Arai, F.; Takubo, K.; Yamazaki, S.; Matsuoka, S.; Miyamoto, T.; Ito, K.; Ohmura, M.; et al. Foxo3a Is Essential for Maintenance of the Hematopoietic Stem Cell Pool. Cell Stem Cell 2007, 1, 101–112. [Google Scholar] [CrossRef] [Green Version]
- Rönnstrand, L. Signal Transduction via the Stem Cell Factor Receptor/c-Kit. CMLS Cell. Mol. Life Sci. 2004, 61, 2535–2548. [Google Scholar] [CrossRef] [PubMed]
- Bartley, T. Identification and Cloning of a Megakaryocyte Growth and Development Factor That Is a Ligand for the Cytokine Receptor MpI. Cell 1994, 77, 1117–1124. [Google Scholar] [CrossRef] [PubMed]
- de Sauvage, F.J.; Hass, P.E.; Spencer, S.D.; Malloy, B.E.; Gurney, A.L.; Spencer, S.A.; Darbonne, W.C.; Henzel, W.J.; Wong, S.C.; Kuang, W.-J.; et al. Stimulation of Megakaryocytopoiesis and Thrombopoiesis by the C-Mpl Ligand. Nature 1994, 369, 533–538. [Google Scholar] [CrossRef]
- Kaushansky, K.; Lok, S.; Holly, R.D.; Broudy, V.C.; Lin, N.; Bailey, M.C.; Forstrom, J.W.; Buddle, M.M.; Oort, P.J.; Hagen, F.S.; et al. Promotion of Megakaryocyte Progenitor Expansion and Differentiation by the C-Mpl Ligand Thrombopoietin. Nature 1994, 369, 568–571. [Google Scholar] [CrossRef] [PubMed]
- Lok, S.; Kaushansky, K.; Holly, R.D.; Kuijper, J.L.; Lofton-Day, C.E.; Oort, P.J.; Grant, F.J.; Heipel, M.D.; Burkhead, S.K.; Kramer, J.M.; et al. Cloning and Expression of Murine Thrombopoietin CDNA and Stimulation of Platelet Production in Vivo. Nature 1994, 369, 565–568. [Google Scholar] [CrossRef]
- Sohma, Y.; Akahori, H.; Seki, N.; Hori, T.; Ogami, K.; Kato, T.; Shimada, Y.; Kawamura, K.; Miyazaki, H. Molecular Cloning and Chromosomal Localization of the Human Thrombopoietin Gene. FEBS Lett. 1994, 353, 57–61. [Google Scholar] [CrossRef] [Green Version]
- Wendling, F.; Maraskovsky, E.; Debili, N.; Florindo, C.; Teepe, M.; Titeux, M.; Methia, N.; Breton-Gorius, J.; Cosman, D.; Vainchenker, W. C-Mpl Ligand Is a Humoral Regulator of Megakaryocytopoiesis. Nature 1994, 369, 571–574. [Google Scholar] [CrossRef]
- Qian, H.; Buza-Vidas, N.; Hyland, C.D.; Jensen, C.T.; Antonchuk, J.; Månsson, R.; Thoren, L.A.; Ekblom, M.; Alexander, W.S.; Jacobsen, S.E.W. Critical Role of Thrombopoietin in Maintaining Adult Quiescent Hematopoietic Stem Cells. Cell Stem Cell 2007, 1, 671–684. [Google Scholar] [CrossRef] [Green Version]
- Yoshihara, H.; Arai, F.; Hosokawa, K.; Hagiwara, T.; Takubo, K.; Nakamura, Y.; Gomei, Y.; Iwasaki, H.; Matsuoka, S.; Miyamoto, K.; et al. Thrombopoietin/MPL Signaling Regulates Hematopoietic Stem Cell Quiescence and Interaction with the Osteoblastic Niche. Cell Stem Cell 2007, 1, 685–697. [Google Scholar] [CrossRef] [Green Version]
- Fox, N.; Priestley, G.; Papayannopoulou, T.; Kaushansky, K. Thrombopoietin Expands Hematopoietic Stem Cells after Transplantation. J. Clin. Investig. 2002, 110, 389–394. [Google Scholar] [CrossRef]
- Kovtonyuk, L.V.; Manz, M.G.; Takizawa, H. Enhanced Thrombopoietin but Not G-CSF Receptor Stimulation Induces Self-Renewing Hematopoietic Stem Cell Divisions in Vivo. Blood 2016, 127, 3175–3179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, S.; Roberts, A.W.; Metcalf, D.; Alexander, W.S. Hematopoietic Stem Cell Deficiencies in Mice Lacking C-Mpl, the Receptor for Thrombopoietin. Proc. Natl. Acad. Sci. USA 1998, 95, 1195–1200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyakawa, Y.; Drachman, J.G.; Gallis, B.; Kaushansky, A.; Kaushansky, K. A Structure-Function Analysis of Serine/Threonine Phosphorylation of the Thrombopoietin Receptor, c-Mpl. J. Biol. Chem. 2000, 275, 32214–32219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyakawa, Y.; Oda, A.; Druker, B.; Kato, T.; Miyazaki, H.; Handa, M.; Ikeda, Y. Recombinant Thrombopoietin Induces Rapid Protein Tyrosine Phosphorylation of Janus Kinase 2 and Shc in Human Blood Platelets. Blood 1995, 86, 23–27. [Google Scholar] [CrossRef] [Green Version]
- Miyakawa, Y.; Oda, A.; Druker, B.; Miyazaki, H.; Handa, M.; Ohashi, H.; Ikeda, Y. Thrombopoietin Induces Tyrosine Phosphorylation of Stat3 and Stat5 in Human Blood Platelets. Blood 1996, 87, 439–446. [Google Scholar] [CrossRef] [Green Version]
- Yamada, M.; Komatsu, N.; Okada, K.; Kato, T.; Miyazaki, H.; Miura, Y. Thrombopoietin Induces Tyrosine Phosphorylation and Activation of Mitogen-Activated Protein Kinases in a Human Thrombopoietin-Dependent Cell Line. Biochem. Biophys. Res. Commun. 1995, 217, 230–237. [Google Scholar] [CrossRef]
- Lyman, S.D.; James, L.; Bos, T.V.; de Vries, P.; Brasel, K.; Gliniak, B.; Hollingsworth, L.T.; Picha, K.S.; McKenna, H.J.; Splett, R.R.; et al. Molecular Cloning of a Ligand for the Flt3flk-2 Tyrosine Kinase Receptor: A Proliferative Factor for Primitive Hematopoietic Cells. Cell 1993, 75, 1157–1167. [Google Scholar] [CrossRef]
- Hannum, C.; Culpepper, J.; Campbell, D.; McClanahan, T.; Zurawski, S.; Kastelein, R.; Bazan, J.F.; Hudak, S.; Wagner, J.; Mattson, J.; et al. Ligand for FLT3/FLK2 Receptor Tyrosine Kinase Regulates Growth of Haematopoietic Stem Cells and Is Encoded by Variant RNAs. Nature 1994, 368, 643–648. [Google Scholar] [CrossRef]
- Masson, K.; Rönnstrand, L. Oncogenic Signaling from the Hematopoietic Growth Factor Receptors C-Kit and Flt3. Cell. Signal. 2009, 21, 1717–1726. [Google Scholar] [CrossRef] [Green Version]
- Levis, M.; Small, D. FLT3: ITDoes Matter in Leukemia. Leukemia 2003, 17, 1738–1752. [Google Scholar] [CrossRef] [Green Version]
- Tenen, D.G. Disruption of Differentiation in Human Cancer: AML Shows the Way. Nat. Rev. Cancer 2003, 3, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Broxmeyer, H.E.; Lu, L.; Cooper, S.; Ruggieri, L.; Li, Z.H.; Lyman, S.D. Flt3 Ligand Stimulates/Costimulates the Growth of Myeloid Stem/Progenitor Cells. Exp. Hematol. 1995, 23, 1121–1129. [Google Scholar] [PubMed]
- Yoo, E.-S.; Ryu, K.-H.; Park, H.-Y.; Seong, C.-M.; Chung, W.-S.; Kim, S.-C.; Choi, Y.-M.; Hahn, M.-J.; Woo, S.-Y.; Seoh, J.-Y. Myeloid Differentiation of Human Cord Blood CD34+ Cells during Ex Vivo Expansion Using Thrombopoietin, Flt3-Ligand and/or Granulocyte-Colony Stimulating Factor. Br. J. Haematol. 1999, 105, 1034–1040. [Google Scholar] [CrossRef]
- Hunter, C.A.; Jones, S.A. IL-6 as a Keystone Cytokine in Health and Disease. Nat. Immunol. 2015, 16, 448–457. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, P.C.; Behrmann, I.; Haan, S.; Hermanns, H.M.; MÜller-Newen, G.; Schaper, F. Principles of Interleukin (IL)-6-Type Cytokine Signalling and Its Regulation. Biochem. J. 2003, 374, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.E.; O’Keefe, R.A.; Grandis, J.R. Targeting the IL-6/JAK/STAT3 Signalling Axis in Cancer. Nat. Rev. Clin. Oncol. 2018, 15, 234–248. [Google Scholar] [CrossRef]
- Skiniotis, G.; Boulanger, M.J.; Garcia, K.C.; Walz, T. Signaling Conformations of the Tall Cytokine Receptor Gp130 When in Complex with IL-6 and IL-6 Receptor. Nat. Struct. Mol. Biol. 2005, 12, 545–551. [Google Scholar] [CrossRef]
- Butler, J.M.; Nolan, D.J.; Vertes, E.L.; Varnum-Finney, B.; Kobayashi, H.; Hooper, A.T.; Seandel, M.; Shido, K.; White, I.A.; Kobayashi, M.; et al. Endothelial Cells Are Essential for the Self-Renewal and Repopulation of Notch-Dependent Hematopoietic Stem Cells. Cell Stem Cell 2010, 6, 251–264. [Google Scholar] [CrossRef] [Green Version]
- Poulos, M.G.; Guo, P.; Kofler, N.M.; Pinho, S.; Gutkin, M.C.; Tikhonova, A.; Aifantis, I.; Frenette, P.S.; Kitajewski, J.; Rafii, S.; et al. Endothelial Jagged-1 Is Necessary for Homeostatic and Regenerative Hematopoiesis. Cell Rep. 2013, 4, 1022–1034. [Google Scholar] [CrossRef] [Green Version]
- Negishi, N.; Suzuki, D.; Ito, R.; Irie, N.; Matsuo, K.; Yahata, T.; Nagano, K.; Aoki, K.; Ohya, K.; Hozumi, K.; et al. Effective Expansion of Engrafted Human Hematopoietic Stem Cells in Bone Marrow of Mice Expressing Human Jagged1. Exp. Hematol. 2014, 42, 487–494.e1. [Google Scholar] [CrossRef]
- Delaney, C.; Varnum-Finney, B.; Aoyama, K.; Brashem-Stein, C.; Bernstein, I.D. Dose-Dependent Effects of the Notch Ligand Delta1 on Ex Vivo Differentiation and in Vivo Marrow Repopulating Ability of Cord Blood Cells. Blood 2005, 106, 2693–2699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delaney, C.; Heimfeld, S.; Brashem-Stein, C.; Voorhies, H.; Manger, R.L.; Bernstein, I.D. Notch-Mediated Expansion of Human Cord Blood Progenitor Cells Capable of Rapid Myeloid Reconstitution. Nat. Med. 2010, 16, 232–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohishi, K.; Varnum-Finney, B.; Bernstein, I.D. Delta-1 Enhances Marrow and Thymus Repopulating Ability of Human CD34+CD38– Cord Blood Cells. J. Clin. Investig. 2002, 110, 1165–1174. [Google Scholar] [CrossRef] [PubMed]
- de Lima, M.; McNiece, I.; Robinson, S.N.; Munsell, M.; Eapen, M.; Horowitz, M.; Alousi, A.; Saliba, R.; McMannis, J.D.; Kaur, I.; et al. Cord-Blood Engraftment with Ex Vivo Mesenchymal-Cell Coculture. N. Engl. J. Med. 2012, 367, 2305–2315. [Google Scholar] [CrossRef] [Green Version]
- Lewis, N.S.; Lewis, E.E.; Mullin, M.; Wheadon, H.; Dalby, M.J.; Berry, C.C. Magnetically Levitated Mesenchymal Stem Cell Spheroids Cultured with a Collagen Gel Maintain Phenotype and Quiescence. J. Tissue Eng. 2017, 8, 204173141770442. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.O.; Rodriguez-Romera, A.; Reyat, J.S.; Olijnik, A.-A.; Colombo, M.; Wang, G.; Wen, W.X.; Sousos, N.; Murphy, L.C.; Grygielska, B.; et al. Human Bone Marrow Organoids for Disease Modeling, Discovery, and Validation of Therapeutic Targets in Hematologic Malignancies. Cancer Discov. 2023, 13, 364–385. [Google Scholar] [CrossRef]
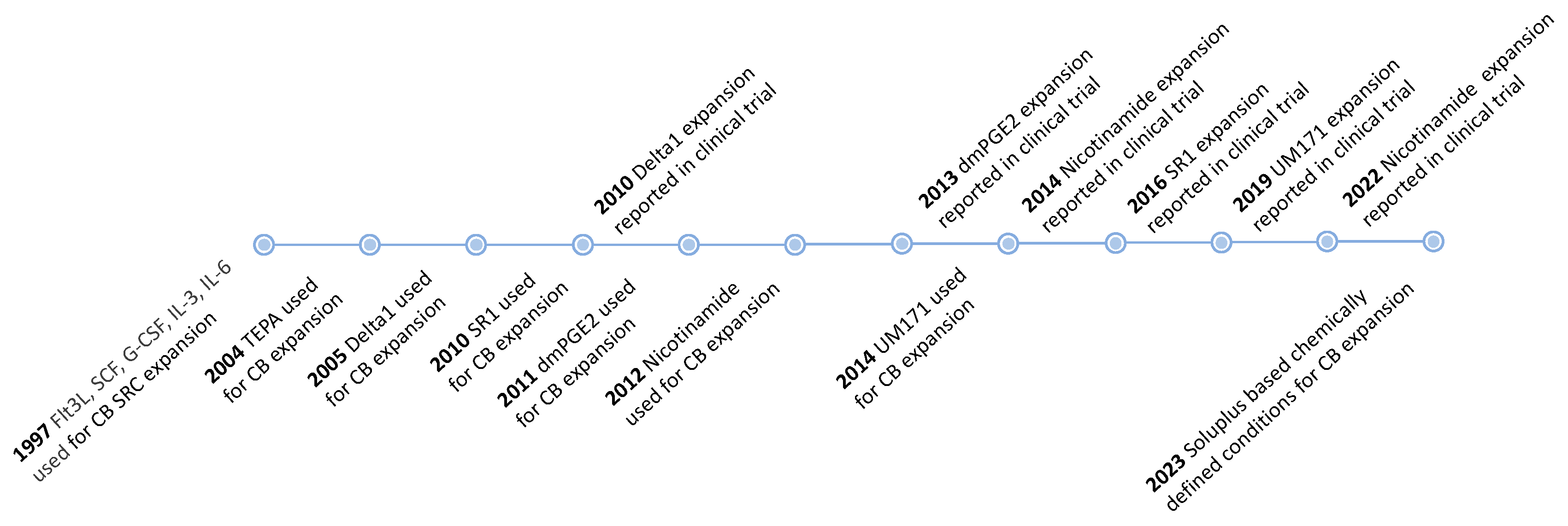
- Peled, T.; Landau, E.; Mandel, J.; Glukhman, E.; Goudsmid, N.R.; Nagler, A.; Fibach, E. Linear Polyamine Copper Chelator Tetraethylenepentamine Augments Long-Term Ex Vivo Expansion of Cord Blood-Derived CD34+ Cells and Increases Their Engraftment Potential in NOD/SCID Mice. Exp. Hematol. 2004, 32, 547–555. [Google Scholar] [CrossRef]
- Peled, T.; Mandel, J.; Goudsmid, R.N.; Landor, C.; Hasson, N.; Harati, D.; Austin, M.; Hasson, A.; Fibach, E.; Shpall, E.J.; et al. Pre-Clinical Development of Cord Blood-Derived Progenitor Cell Graft Expanded Ex Vivo with Cytokines and the Polyamine Copper Chelator Tetraethylenepentamine. Cytotherapy 2004, 6, 344–355. [Google Scholar] [CrossRef]
- Peled, T.; Landau, E.; Prus, E.; Treves, A.J.; Fibach, E. Cellular Copper Content Modulates Differentiation and Self-Renewal in Cultures of Cord Blood-Derived CD34+ Cells: Copper Modulates Differentiation/Self-Renewal of CD34 Cells. Br. J. Haematol. 2002, 116, 655–661. [Google Scholar] [CrossRef] [Green Version]
- Peled, T.; Glukhman, E.; Hasson, N.; Adi, S.; Assor, H.; Yudin, D.; Landor, C.; Mandel, J.; Landau, E.; Prus, E.; et al. Chelatable Cellular Copper Modulates Differentiation and Self-Renewal of Cord Blood–Derived Hematopoietic Progenitor Cells. Exp. Hematol. 2005, 33, 1092–1100. [Google Scholar] [CrossRef]
- de Lima, M.; McMannis, J.; Gee, A.; Komanduri, K.; Couriel, D.; Andersson, B.S.; Hosing, C.; Khouri, I.; Jones, R.; Champlin, R.; et al. Transplantation of Ex Vivo Expanded Cord Blood Cells Using the Copper Chelator Tetraethylenepentamine: A Phase I/II Clinical Trial. Bone Marrow Transpl. 2008, 41, 771–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stiff, P.J.; Montesinos, P.; Peled, T.; Landau, E.; Goudsmid, N.R.; Mandel, J.; Hasson, N.; Olesinski, E.; Glukhman, E.; Snyder, D.A.; et al. Cohort-Controlled Comparison of Umbilical Cord Blood Transplantation Using Carlecortemcel-L, a Single Progenitor–Enriched Cord Blood, to Double Cord Blood Unit Transplantation. Biol. Blood Marrow Transplant. 2018, 24, 1463–1470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, J.E.; Brunstein, C.G.; Boitano, A.E.; DeFor, T.E.; McKenna, D.; Sumstad, D.; Blazar, B.R.; Tolar, J.; Le, C.; Jones, J.; et al. Phase I/II Trial of StemRegenin-1 Expanded Umbilical Cord Blood Hematopoietic Stem Cells Supports Testing as a Stand-Alone Graft. Cell Stem Cell 2016, 18, 144–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peled, T.; Shoham, H.; Aschengrau, D.; Yackoubov, D.; Frei, G.; Rosenheimer G, N.; Lerrer, B.; Cohen, H.Y.; Nagler, A.; Fibach, E.; et al. Nicotinamide, a SIRT1 Inhibitor, Inhibits Differentiation and Facilitates Expansion of Hematopoietic Progenitor Cells with Enhanced Bone Marrow Homing and Engraftment. Exp. Hematol. 2012, 40, 342–355.e1. [Google Scholar] [CrossRef] [PubMed]
- Horwitz, M.E.; Chao, N.J.; Rizzieri, D.A.; Long, G.D.; Sullivan, K.M.; Gasparetto, C.; Chute, J.P.; Morris, A.; McDonald, C.; Waters-Pick, B.; et al. Umbilical Cord Blood Expansion with Nicotinamide Provides Long-Term Multilineage Engraftment. J. Clin. Investig. 2014, 124, 3121–3128. [Google Scholar] [CrossRef] [Green Version]
- Horwitz, M.E.; Wease, S.; Blackwell, B.; Valcarcel, D.; Frassoni, F.; Boelens, J.J.; Nierkens, S.; Jagasia, M.; Wagner, J.E.; Kuball, J.; et al. Phase I/II Study of Stem-Cell Transplantation Using a Single Cord Blood Unit Expanded Ex Vivo With Nicotinamide. JCO 2019, 37, 367–374. [Google Scholar] [CrossRef]
- Goessling, W.; North, T.E.; Loewer, S.; Lord, A.M.; Lee, S.; Stoick-Cooper, C.L.; Weidinger, G.; Puder, M.; Daley, G.Q.; Moon, R.T.; et al. Genetic Interaction of PGE2 and Wnt Signaling Regulates Developmental Specification of Stem Cells and Regeneration. Cell 2009, 136, 1136–1147. [Google Scholar] [CrossRef] [Green Version]
- Ikushima, Y.M.; Arai, F.; Hosokawa, K.; Toyama, H.; Takubo, K.; Furuyashiki, T.; Narumiya, S.; Suda, T. Prostaglandin E2 Regulates Murine Hematopoietic Stem/Progenitor Cells Directly via EP4 Receptor and Indirectly through Mesenchymal Progenitor Cells. Blood 2013, 121, 1995–2007. [Google Scholar] [CrossRef] [Green Version]
- North, T.E.; Goessling, W.; Walkley, C.R.; Lengerke, C.; Kopani, K.R.; Lord, A.M.; Weber, G.J.; Bowman, T.V.; Jang, I.-H.; Grosser, T.; et al. Prostaglandin E2 Regulates Vertebrate Haematopoietic Stem Cell Homeostasis. Nature 2007, 447, 1007–1011. [Google Scholar] [CrossRef] [Green Version]
- Goessling, W.; Allen, R.S.; Guan, X.; Jin, P.; Uchida, N.; Dovey, M.; Harris, J.M.; Metzger, M.E.; Bonifacino, A.C.; Stroncek, D.; et al. Prostaglandin E2 Enhances Human Cord Blood Stem Cell Xenotransplants and Shows Long-Term Safety in Preclinical Nonhuman Primate Transplant Models. Cell Stem Cell 2011, 8, 445–458. [Google Scholar] [CrossRef] [Green Version]
- Cutler, C.; Multani, P.; Robbins, D.; Kim, H.T.; Le, T.; Hoggatt, J.; Pelus, L.M.; Desponts, C.; Chen, Y.-B.; Rezner, B.; et al. Prostaglandin-Modulated Umbilical Cord Blood Hematopoietic Stem Cell Transplantation. Blood 2013, 122, 3074–3081. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Desai, A.; Yang, S.Y.; Bae, K.B.; Antczak, M.I.; Fink, S.P.; Tiwari, S.; Willis, J.E.; Williams, N.S.; Dawson, D.M.; et al. Inhibition of the Prostaglandin-Degrading Enzyme 15-PGDH Potentiates Tissue Regeneration. Science 2015, 348, aaa2340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desai, A.; Zhang, Y.; Park, Y.; Dawson, D.M.; Larusch, G.A.; Kasturi, L.; Wald, D.; Ready, J.M.; Gerson, S.L.; Markowitz, S.D. A Second-Generation 15-PGDH Inhibitor Promotes Bone Marrow Transplant Recovery Independently of Age, Transplant Dose and Granulocyte Colony-Stimulating Factor Support. Haematologica 2018, 103, 1054–1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fares, I.; Chagraoui, J.; Gareau, Y.; Gingras, S.; Ruel, R.; Mayotte, N.; Csaszar, E.; Knapp, D.J.H.F.; Miller, P.; Ngom, M.; et al. Pyrimidoindole Derivatives Are Agonists of Human Hematopoietic Stem Cell Self-Renewal. Science 2014, 345, 1509–1512. [Google Scholar] [CrossRef] [Green Version]
- Subramaniam, A.; Žemaitis, K.; Talkhoncheh, M.S.; Yudovich, D.; Bäckström, A.; Debnath, S.; Chen, J.; Jain, M.V.; Galeev, R.; Gaetani, M.; et al. Lysine-Specific Demethylase 1A Restricts Ex Vivo Propagation of Human HSCs and Is a Target of UM171. Blood 2020, 136, 2151–2161. [Google Scholar] [CrossRef]
- Cohen, S.; Roy, J.; Lachance, S.; Delisle, J.-S.; Marinier, A.; Busque, L.; Roy, D.-C.; Barabé, F.; Ahmad, I.; Bambace, N.; et al. Hematopoietic Stem Cell Transplantation Using Single UM171-Expanded Cord Blood: A Single-Arm, Phase 1–2 Safety and Feasibility Study. Lancet Haematol. 2020, 7, e134–e145. [Google Scholar] [CrossRef]
- Feng, Y.; Xie, X.-Y.; Yang, Y.-Q.; Sun, Y.-T.; Ma, W.-H.; Zhou, P.-J.; Li, Z.-Y.; Liu, H.-Q.; Wang, Y.-F.; Huang, Y.-S. Synthesis and Evaluation of Pyrimidoindole Analogs in Umbilical Cord Blood Ex Vivo Expansion. Eur. J. Med. Chem. 2019, 174, 181–197. [Google Scholar] [CrossRef]
- Nishino, T.; Miyaji, K.; Ishiwata, N.; Arai, K.; Yui, M.; Asai, Y.; Nakauchi, H.; Iwama, A. Ex Vivo Expansion of Human Hematopoietic Stem Cells by a Small-Molecule Agonist of c-MPL. Exp. Hematol. 2009, 37, 1364–1377.e4. [Google Scholar] [CrossRef]
- Sun, H.; Tsai, Y.; Nowak, I.; Liesveld, J.; Chen, Y. Eltrombopag, a Thrombopoietin Receptor Agonist, Enhances Human Umbilical Cord Blood Hematopoietic Stem/Primitive Progenitor Cell Expansion and Promotes Multi-Lineage Hematopoiesis. Stem Cell Res. 2012, 9, 77–86. [Google Scholar] [CrossRef] [Green Version]
- Kao, Y.-R.; Chen, J.; Narayanagari, S.-R.; Todorova, T.I.; Aivalioti, M.M.; Ferreira, M.; Ramos, P.M.; Pallaud, C.; Mantzaris, I.; Shastri, A.; et al. Thrombopoietin Receptor–Independent Stimulation of Hematopoietic Stem Cells by Eltrombopag. Sci. Transl. Med. 2018, 10, eaas9563. [Google Scholar] [CrossRef]
- Geest, C.R.; Coffer, P.J. MAPK Signaling Pathways in the Regulation of Hematopoiesis. J. Leukoc. Biol. 2009, 86, 237–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, J.; Zou, P.; Wang, J.; Li, L.; Wang, Y.; Zhou, D.; Liu, L. Inhibition of P38 MAPK Activity Promotes Ex Vivo Expansion of Human Cord Blood Hematopoietic Stem Cells. Ann. Hematol. 2012, 91, 813–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bari, S.; Zhong, Q.; Fan, X.; Poon, Z.; Lim, A.S.T.; Lim, T.H.; Dighe, N.; Li, S.; Chiu, G.N.C.; Chai, C.L.L.; et al. Ex Vivo Expansion of CD34+CD90+CD49f+ Hematopoietic Stem and Progenitor Cells from Non-Enriched Umbilical Cord Blood with Azole Compounds. Stem Cells Transl. Med. 2018, 7, 376–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, X.; Lai, W.; Xie, H.; Liu, Y.; Guo, W.; Liu, Y.; Li, Y.; Li, Y.; Zhang, J.; Chen, W.; et al. Targeting JNK Pathway Promotes Human Hematopoietic Stem Cell Expansion. Cell Discov. 2019, 5, 2. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Inesta-Vaquera, F.; Niepel, M.; Zhang, J.; Ficarro, S.B.; Machleidt, T.; Xie, T.; Marto, J.A.; Kim, N.; Sim, T.; et al. Discovery of Potent and Selective Covalent Inhibitors of JNK. Chem. Biol. 2012, 19, 140–154. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Yuan, Y.; Shen, H.; Cheng, T. Hematopoietic Stem Cell Exhaustion Impacted by P18INK4C and P21Cip1/Waf1 in Opposite Manners. Blood 2006, 107, 1200–1206. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Shen, H.; Franklin, D.S.; Scadden, D.T.; Cheng, T. In Vivo Self-Renewing Divisions of Haematopoietic Stem Cells Are Increased in the Absence of the Early G1-Phase Inhibitor, P18INK4C. Nat. Cell Biol. 2004, 6, 436–442. [Google Scholar] [CrossRef]
- Gao, Y.; Yang, P.; Shen, H.; Yu, H.; Song, X.; Zhang, L.; Zhang, P.; Cheng, H.; Xie, Z.; Hao, S.; et al. Small-Molecule Inhibitors Targeting INK4 Protein P18INK4C Enhance Ex Vivo Expansion of Haematopoietic Stem Cells. Nat. Commun. 2015, 6, 6328. [Google Scholar] [CrossRef] [Green Version]
- Xie, X.-Q.; Yang, P.; Zhang, Y.; Zhang, P.; Wang, L.; Ding, Y.; Yang, M.; Tong, Q.; Cheng, H.; Ji, Q.; et al. Discovery of Novel INK4C Small-Molecule Inhibitors to Promote Human and Murine Hematopoietic Stem Cell Ex Vivo Expansion. Sci. Rep. 2015, 5, 18115. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhang, W.; Zhang, Y.; Ding, Y.; Yang, M.; He, M.; Liu, X.; Gu, J.; Xu, S.; Feng, Z.; et al. Enhanced Self-Renewal of Human Long-Term Hematopoietic Stem Cells by a Sulfamoyl Benzoate Derivative Targeting P18INK4C. Blood Adv. 2021, 5, 3362–3372. [Google Scholar] [CrossRef]
- Bug, G.; Gül, H.; Schwarz, K.; Pfeifer, H.; Kampfmann, M.; Zheng, X.; Beissert, T.; Boehrer, S.; Hoelzer, D.; Ottmann, O.G.; et al. Valproic Acid Stimulates Proliferation and Self-Renewal of Hematopoietic Stem Cells. Cancer Res. 2005, 65, 2537–2541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Felice, L.; Tatarelli, C.; Mascolo, M.G.; Gregorj, C.; Agostini, F.; Fiorini, R.; Gelmetti, V.; Pascale, S.; Padula, F.; Petrucci, M.T.; et al. Histone Deacetylase Inhibitor Valproic Acid Enhances the Cytokine-Induced Expansion of Human Hematopoietic Stem Cells. Cancer Res. 2005, 65, 1505–1513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papa, L.; Zimran, E.; Djedaini, M.; Ge, Y.; Ozbek, U.; Sebra, R.; Sealfon, S.C.; Hoffman, R. Ex Vivo Human HSC Expansion Requires Coordination of Cellular Reprogramming with Mitochondrial Remodeling and P53 Activation. Blood Adv. 2018, 2, 2766–2779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimran, E.; Papa, L.; Djedaini, M.; Patel, A.; Iancu-Rubin, C.; Hoffman, R. Expansion and Preservation of the Functional Activity of Adult Hematopoietic Stem Cells Cultured Ex Vivo with a Histone Deacetylase Inhibitor. Stem Cells Transl. Med. 2020, 9, 531–542. [Google Scholar] [CrossRef] [Green Version]
- Chaurasia, P.; Gajzer, D.C.; Schaniel, C.; D’Souza, S.; Hoffman, R. Epigenetic Reprogramming Induces the Expansion of Cord Blood Stem Cells. J. Clin. Investig. 2014, 124, 2378–2395. [Google Scholar] [CrossRef]
- Seet, L.-F.; Teng, E.; Lai, Y.-S.; Laning, J.; Kraus, M.; Wnendt, S.; Merchav, S.; Chan, S.L. Valproic Acid Enhances the Engraftability of Human Umbilical Cord Blood Hematopoietic Stem Cells Expanded under Serum-Free Conditions*. Eur. J. Haematol. 2009, 82, 124–132. [Google Scholar] [CrossRef]
- Nishino, T.; Wang, C.; Mochizuki-Kashio, M.; Osawa, M.; Nakauchi, H.; Iwama, A. Ex Vivo Expansion of Human Hematopoietic Stem Cells by Garcinol, a Potent Inhibitor of Histone Acetyltransferase. PLoS ONE 2011, 6, e24298. [Google Scholar] [CrossRef]
- Araki, H.; Mahmud, N.; Milhem, M.; Nunez, R.; Xu, M.; Beam, C.A.; Hoffman, R. Expansion of Human Umbilical Cord Blood SCID-Repopulating Cells Using Chromatin-Modifying Agents. Exp. Hematol. 2006, 34, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Saraf, S.; Araki, H.; Petro, B.; Park, Y.; Taioli, S.; Yoshinaga, K.G.; Koca, E.; Rondelli, D.; Mahmud, N. Ex Vivo Expansion of Human Mobilized Peripheral Blood Stem Cells Using Epigenetic Modifiers: Epigenetic Modifiers and PBSC Expansion. Transfusion 2015, 55, 864–874. [Google Scholar] [CrossRef] [Green Version]
- Araki, H.; Yoshinaga, K.; Boccuni, P.; Zhao, Y.; Hoffman, R.; Mahmud, N. Chromatin-Modifying Agents Permit Human Hematopoietic Stem Cells to Undergo Multiple Cell Divisions While Retaining Their Repopulating Potential. Blood 2007, 109, 3570–3578. [Google Scholar] [CrossRef]
- Hua, P.; Hester, J.; Adigbli, G.; Li, R.; Psaila, B.; Roy, A.; Bataille, C.J.R.; Wynne, G.M.; Jackson, T.; Milne, T.A.; et al. The BET Inhibitor CPI203 Promotes Ex Vivo Expansion of Cord Blood Long-Term Repopulating HSCs and Megakaryocytes. Blood 2020, 136, 2410–2415. [Google Scholar] [CrossRef] [PubMed]
- Trowbridge, J.J.; Xenocostas, A.; Moon, R.T.; Bhatia, M. Glycogen Synthase Kinase-3 Is an in Vivo Regulator of Hematopoietic Stem Cell Repopulation. Nat. Med. 2006, 12, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Perry, J.M.; He, X.C.; Sugimura, R.; Grindley, J.C.; Haug, J.S.; Ding, S.; Li, L. Cooperation between Both Wnt/β-Catenin and PTEN/PI3K/Akt Signaling Promotes Primitive Hematopoietic Stem Cell Self-Renewal and Expansion. Genes Dev. 2011, 25, 1928–1942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, K.-H.; Holmes, T.; Palladinetti, P.; Song, E.; Nordon, R.; O’Brien, T.A.; Dolnikov, A. GSK-3β Inhibition Promotes Engraftment of Ex Vivo-Expanded Hematopoietic Stem Cells and Modulates Gene Expression. Stem Cells 2011, 29, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Nguyen-McCarty, M.; Hexner, E.O.; Danet-Desnoyers, G.; Klein, P.S. Maintenance of Hematopoietic Stem Cells through Regulation of Wnt and MTOR Pathways. Nat. Med. 2012, 18, 1778–1785. [Google Scholar] [CrossRef] [Green Version]
- Sakurai, M.; Ishitsuka, K.; Ito, R.; Wilkinson, A.C.; Kimura, T.; Mizutani, E.; Nishikii, H.; Sudo, K.; Becker, H.J.; Takemoto, H.; et al. Chemically Defined Cytokine-Free Expansion of Human Haematopoietic Stem Cells. Nature 2023, 615, 127–133. [Google Scholar] [CrossRef]
- Nakeff, A.; Maat, B. Separation of Megakaryocytes From Mouse Bone Marrow by Velocity Sedimentation. Blood 1974, 43, 591–595. [Google Scholar] [CrossRef]
- George, J.N. Platelets. Lancet 2000, 355, 1531–1539. [Google Scholar] [CrossRef]
- McArthur, K.; Chappaz, S.; Kile, B.T. Apoptosis in Megakaryocytes and Platelets: The Life and Death of a Lineage. Blood 2018, 131, 605–610. [Google Scholar] [CrossRef] [Green Version]
- Stroncek, D.F.; Rebulla, P. Platelet Transfusions. Lancet 2007, 370, 427–438. [Google Scholar] [CrossRef] [Green Version]
- Squires, J.E. Indications for Platelet Transfusion in Patients with Thrombocytopenia. Blood Transfus. 2015, 13, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Sanjuan-Pla, A.; Macaulay, I.C.; Jensen, C.T.; Woll, P.S.; Luis, T.C.; Mead, A.; Moore, S.; Carella, C.; Matsuoka, S.; Jones, T.B.; et al. Platelet-Biased Stem Cells Reside at the Apex of the Haematopoietic Stem-Cell Hierarchy. Nature 2013, 502, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Seita, J.; Weissman, I.L. Hematopoietic Stem Cell: Self-renewal versus Differentiation. WIREs Mech. Dis. 2010, 2, 640–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, E.; Nichol, J.; Hokom, M.; Hornkohl, A.; Hunt, P. Platelets Generated in Vitro from Proplatelet-Displaying Human Megakaryocytes Are Functional. Blood 1995, 85, 402–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norol, F.; Vitrat, N.; Cramer, E.; Guichard, J.; Burstein, S.A.; Vainchenker, W.; Debili, N. Effects of Cytokines on Platelet Production from Blood and Marrow CD34+ Cells. Blood 1998, 91, 830–843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proulx, C.; Dupuis, N.; St-Amour, I.; Boyer, L.; Lemieux, R. Increased Megakaryopoiesis in Cultures of CD34-Enriched Cord Blood Cells Maintained at 39 °C: Increased Megakaryopoiesis in CB Cultures. Biotechnol. Bioeng. 2004, 88, 675–680. [Google Scholar] [CrossRef]
- Matsunaga, T.; Tanaka, I.; Kobune, M.; Kawano, Y.; Tanaka, M.; Kuribayashi, K.; Iyama, S.; Sato, T.; Sato, Y.; Takimoto, R.; et al. Ex Vivo Large-Scale Generation of Human Platelets from Cord Blood CD34+ Cells. Stem Cells 2006, 24, 2877–2887. [Google Scholar] [CrossRef]
- Guan, X.; Qin, M.; Zhang, Y.; Wang, Y.; Shen, B.; Ren, Z.; Ding, X.; Dai, W.; Jiang, Y. Safety and Efficacy of Megakaryocytes Induced from Hematopoietic Stem Cells in Murine and Nonhuman Primate Models. Stem Cells Transl. Med. 2017, 6, 897–909. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, C.; Lei, X.; Wang, H.; Su, P.; Ru, Y.; Ruan, X.; Duan, E.; Feng, S.; Han, M.; et al. Integrated Biophysical and Biochemical Signals Augment Megakaryopoiesis and Thrombopoiesis in a Three-Dimensional Rotary Culture System. Stem Cells Transl. Med. 2016, 5, 175–185. [Google Scholar] [CrossRef] [Green Version]
- Guan, X.; Wang, L.; Wang, H.; Wang, H.; Dai, W.; Jiang, Y. Good Manufacturing Practice-Grade of Megakaryocytes Produced by a Novel Ex Vivo Culturing Platform. Clin. Transl. Sci. 2020, 13, 1115–1126. [Google Scholar] [CrossRef]
- Orkin, S.H.; Zon, L.I. Hematopoiesis: An Evolving Paradigm for Stem Cell Biology. Cell 2008, 132, 631–644. [Google Scholar] [CrossRef] [Green Version]
- Pimanda, J.E.; Gttgens, B. Gene Regulatory Networks Governing Haematopoietic Stem Cell Development and Identity. Int. J. Dev. Biol. 2010, 54, 1201–1211. [Google Scholar] [CrossRef] [PubMed]
- Starck, J.; Cohet, N.; Gonnet, C.; Sarrazin, S.; Doubeikovskaia, Z.; Doubeikovski, A.; Verger, A.; Duterque-Coquillaud, M.; Morle, F. Functional Cross-Antagonism between Transcription Factors FLI-1 and EKLF. Mol. Cell Biol. 2003, 23, 1390–1402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doré, L.C.; Crispino, J.D. Transcription Factor Networks in Erythroid Cell and Megakaryocyte Development. Blood 2011, 118, 231–239. [Google Scholar] [CrossRef] [Green Version]
- Tijssen, M.R.; Cvejic, A.; Joshi, A.; Hannah, R.L.; Ferreira, R.; Forrai, A.; Bellissimo, D.C.; Oram, S.H.; Smethurst, P.A.; Wilson, N.K.; et al. Genome-Wide Analysis of Simultaneous GATA1/2, RUNX1, FLI1, and SCL Binding in Megakaryocytes Identifies Hematopoietic Regulators. Dev. Cell 2011, 20, 597–609. [Google Scholar] [CrossRef] [Green Version]
- Klimchenko, O.; Mori, M.; DiStefano, A.; Langlois, T.; Larbret, F.; Lecluse, Y.; Feraud, O.; Vainchenker, W.; Norol, F.; Debili, N. A Common Bipotent Progenitor Generates the Erythroid and Megakaryocyte Lineages in Embryonic Stem Cell–Derived Primitive Hematopoiesis. Blood 2009, 114, 1506–1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, N.K.; Foster, S.D.; Wang, X.; Knezevic, K.; Schütte, J.; Kaimakis, P.; Chilarska, P.M.; Kinston, S.; Ouwehand, W.H.; Dzierzak, E.; et al. Combinatorial Transcriptional Control In Blood Stem/Progenitor Cells: Genome-Wide Analysis of Ten Major Transcriptional Regulators. Cell Stem Cell 2010, 7, 532–544. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Cantor, A.B. Common Features of Megakaryocytes and Hematopoietic Stem Cells: What’s the Connection? J. Cell. Biochem. 2009, 107, 857–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fielder, P.J.; Hass, P.; Nagel, M.; Stefanich, E.; Widmer, R.; Bennett, G.L.; Keller, G.-A.; de Sauvage, F.J.; Eaton, D. Human Platelets as a Model for the Binding and Degradation of Thrombopoietin. Blood 1997, 89, 2782–2788. [Google Scholar] [CrossRef]
- Kuter, D.; Rosenberg, R. The Reciprocal Relationship of Thrombopoietin (c-Mpl Ligand) to Changes in the Platelet Mass during Busulfan-Induced Thrombocytopenia in the Rabbit. Blood 1995, 85, 2720–2730. [Google Scholar] [CrossRef] [Green Version]
- McCarty, J.; Sprugel, K.; Fox, N.; Sabath, D.; Kaushansky, K. Murine Thrombopoietin MRNA Levels Are Modulated by Platelet Count. Blood 1995, 86, 3668–3675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Den Oudenrijn, S.; Bruin, M.; Folman, C.C.; Peters, M.; Faulkner, L.B.; De Haas, M.; Von Dem Borne, A.E.G.K.R. Mutations in the Thrombopoietin Receptor, Mpl, in Children with Congenital Amegakaryocytic Thrombocytopenia: C-Mpl Mutations in Amegakaryocytic Thrombocytopenia. Br. J. Haematol. 2000, 110, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Commins, S.P.; Borish, L.; Steinke, J.W. Immunologic Messenger Molecules: Cytokines, Interferons, and Chemokines. J. Allergy Clin. Immunol. 2010, 125, S53–S72. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H.; Ishibashi, T.; Shikama, Y.; Okano, A.; Akiyama, Y.; Uchida, T.; Maruyama, Y. Interleukin-1 Beta (IL-1 Beta) Induces Thrombocytosis in Mice: Possible Implication of IL-6. Blood 1990, 76, 2493–2500. [Google Scholar] [CrossRef] [PubMed]
- Kaser, A.; Brandacher, G.; Steurer, W.; Kaser, S.; Offner, F.A.; Zoller, H.; Theurl, I.; Widder, W.; Molnar, C.; Ludwiczek, O.; et al. Interleukin-6 Stimulates Thrombopoiesis through Thrombopoietin: Role in Inflammatory Thrombocytosis. Blood 2001, 98, 2720–2725. [Google Scholar] [CrossRef] [Green Version]
- Segal, G.M.; Stueve, T.; Adamson, J.W. Analysis of Murine Megakaryocyte Colony Size and Ploidy: Effects of Interleukin-3. J. Cell. Physiol. 1988, 137, 537–544. [Google Scholar] [CrossRef]
- Yang, Y.-C.; Ciarletta, A.B.; Temple, P.A.; Chung, M.P.; Kovacic, S.; Witek-Giannotti, J.S.; Leary, A.C.; Kriz, R.; Donahue, R.E.; Wong, G.G.; et al. Human IL-3 (Multi-CSF): Identification by Expression Cloning of a Novel Hematopoietic Growth Factor Related to Murine IL-3. Cell 1986, 47, 3–10. [Google Scholar] [CrossRef]
- Nishimura, S.; Nagasaki, M.; Kunishima, S.; Sawaguchi, A.; Sakata, A.; Sakaguchi, H.; Ohmori, T.; Manabe, I.; Italiano, J.E.; Ryu, T.; et al. IL-1α Induces Thrombopoiesis through Megakaryocyte Rupture in Response to Acute Platelet Needs. J. Cell Biol. 2015, 209, 453–466. [Google Scholar] [CrossRef] [Green Version]
- Steinberg, M.H.; Forget, B.G.; Higgs, D.R.; Weatherall, D.J. Disorders of Hemoglobin: Genetics, Pathophysiology, and Clinical Management, 2nd ed.; Cambridge University Press: Cambridge, UK, 2009; ISBN 978-0-521-87519-6. [Google Scholar]
- Hirani, R.; Mondy, P. Review of Full Blood Count Reference Interval Using a Large Cohort of First-Time Plasmapheresis Blood Donors. Pathology 2021, 53, 498–502. [Google Scholar] [CrossRef]
- Cable, C.A.; Razavi, S.A.; Roback, J.D.; Murphy, D.J. RBC Transfusion Strategies in the ICU: A Concise Review. Crit. Care Med. 2019, 47, 1637–1644. [Google Scholar] [CrossRef]
- Carson, J.L.; Stanworth, S.J.; Dennis, J.A.; Trivella, M.; Roubinian, N.; Fergusson, D.A.; Triulzi, D.; Dorée, C.; Hébert, P.C. Transfusion Thresholds for Guiding Red Blood Cell Transfusion. Cochrane Database Syst. Rev. 2021, 2022. [Google Scholar] [CrossRef]
- Neildez-Nguyen, T.M.A.; Wajcman, H.; Marden, M.C.; Bensidhoum, M.; Moncollin, V.; Giarratana, M.-C.; Kobari, L.; Thierry, D.; Douay, L. Human Erythroid Cells Produced Ex Vivo at Large Scale Differentiate into Red Blood Cells in Vivo. Nat. Biotechnol. 2002, 20, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Giarratana, M.-C.; Kobari, L.; Lapillonne, H.; Chalmers, D.; Kiger, L.; Cynober, T.; Marden, M.C.; Wajcman, H.; Douay, L. Ex Vivo Generation of Fully Mature Human Red Blood Cells from Hematopoietic Stem Cells. Nat. Biotechnol. 2005, 23, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Miharada, K.; Hiroyama, T.; Sudo, K.; Nagasawa, T.; Nakamura, Y. Efficient Enucleation of Erythroblasts Differentiated in Vitro from Hematopoietic Stem and Progenitor Cells. Nat. Biotechnol. 2006, 24, 1255–1256. [Google Scholar] [CrossRef] [PubMed]
- Giarratana, M.-C.; Rouard, H.; Dumont, A.; Kiger, L.; Safeukui, I.; Le Pennec, P.-Y.; François, S.; Trugnan, G.; Peyrard, T.; Marie, T.; et al. Proof of Principle for Transfusion of in Vitro–Generated Red Blood Cells. Blood 2011, 118, 5071–5079. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wang, C.; Wang, L.; Shen, B.; Guan, X.; Tian, J.; Ren, Z.; Ding, X.; Ma, Y.; Dai, W.; et al. Large-Scale Ex Vivo Generation of Human Red Blood Cells from Cord Blood CD34+ Cells. Stem Cells Transl. Med. 2017, 6, 1698–1709. [Google Scholar] [CrossRef]
- Hattangadi, S.M.; Wong, P.; Zhang, L.; Flygare, J.; Lodish, H.F. From Stem Cell to Red Cell: Regulation of Erythropoiesis at Multiple Levels by Multiple Proteins, RNAs, and Chromatin Modifications. Blood 2011, 118, 6258–6268. [Google Scholar] [CrossRef] [Green Version]
- Caulier, A.L.; Sankaran, V.G. Molecular and Cellular Mechanisms That Regulate Human Erythropoiesis. Blood 2022, 139, 2450–2459. [Google Scholar] [CrossRef]
- Capellera-Garcia, S.; Pulecio, J.; Dhulipala, K.; Siva, K.; Rayon-Estrada, V.; Singbrant, S.; Sommarin, M.N.E.; Walkley, C.R.; Soneji, S.; Karlsson, G.; et al. Defining the Minimal Factors Required for Erythropoiesis through Direct Lineage Conversion. Cell Rep. 2016, 15, 2550–2562. [Google Scholar] [CrossRef] [Green Version]
- Sankaran, V.G.; Ghazvinian, R.; Do, R.; Thiru, P.; Vergilio, J.-A.; Beggs, A.H.; Sieff, C.A.; Orkin, S.H.; Nathan, D.G.; Lander, E.S.; et al. Exome Sequencing Identifies GATA1 Mutations Resulting in Diamond-Blackfan Anemia. J. Clin. Investig. 2012, 122, 2439–2443. [Google Scholar] [CrossRef] [Green Version]
- Katsumura, K.R.; Bresnick, E.H.; the GATA Factor Mechanisms Group. The GATA Factor Revolution in Hematology. Blood 2017, 129, 2092–2102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakabayashi, A.; Ulirsch, J.C.; Ludwig, L.S.; Fiorini, C.; Yasuda, M.; Choudhuri, A.; McDonel, P.; Zon, L.I.; Sankaran, V.G. Insight into GATA1 Transcriptional Activity through Interrogation of Cis Elements Disrupted in Human Erythroid Disorders. Proc. Natl. Acad. Sci. USA 2016, 113, 4434–4439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez, L.; Tsukamoto, S.; Suzuki, M.; Yamamoto-Mukai, H.; Yamamoto, M.; Philipsen, S.; Ohneda, K. Ablation of Gata1 in Adult Mice Results in Aplastic Crisis, Revealing Its Essential Role in Steady-State and Stress Erythropoiesis. Blood 2008, 111, 4375–4385. [Google Scholar] [CrossRef] [Green Version]
- Fujiwara, Y.; Browne, C.P.; Cunniff, K.; Goff, S.C.; Orkin, S.H. Arrested Development of Embryonic Red Cell Precursors in Mouse Embryos Lacking Transcription Factor GATA-1. Proc. Natl. Acad. Sci. USA 1996, 93, 12355–12358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crispino, J.D.; Horwitz, M.S. GATA Factor Mutations in Hematologic Disease. Blood 2017, 129, 2103–2110. [Google Scholar] [CrossRef]
- Abdulhay, N.J.; Fiorini, C.; Verboon, J.M.; Ludwig, L.S.; Ulirsch, J.C.; Zieger, B.; Lareau, C.A.; Mi, X.; Roy, A.; Obeng, E.A.; et al. Impaired Human Hematopoiesis Due to a Cryptic Intronic GATA1 Splicing Mutation. J. Exp. Med. 2019, 216, 1050–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frontelo, P.; Manwani, D.; Galdass, M.; Karsunky, H.; Lohmann, F.; Gallagher, P.G.; Bieker, J.J. Novel Role for EKLF in Megakaryocyte Lineage Commitment. Blood 2007, 110, 3871–3880. [Google Scholar] [CrossRef] [Green Version]
- Bouilloux, F.; Juban, G.; Cohet, N.; Buet, D.; Guyot, B.; Vainchenker, W.; Louache, F.; Morlé, F. EKLF Restricts Megakaryocytic Differentiation at the Benefit of Erythrocytic Differentiation. Blood 2008, 112, 576–584. [Google Scholar] [CrossRef]
- Gnanapragasam, M.N.; McGrath, K.E.; Catherman, S.; Xue, L.; Palis, J.; Bieker, J.J. EKLF/KLF1-Regulated Cell Cycle Exit Is Essential for Erythroblast Enucleation. Blood 2016, 128, 1631–1641. [Google Scholar] [CrossRef]
- Zhou, D.; Liu, K.; Sun, C.-W.; Pawlik, K.M.; Townes, T.M. KLF1 Regulates BCL11A Expression and γ- to β-Globin Gene Switching. Nat. Genet. 2010, 42, 742–744. [Google Scholar] [CrossRef]
- Arnaud, L.; Saison, C.; Helias, V.; Lucien, N.; Steschenko, D.; Giarratana, M.-C.; Prehu, C.; Foliguet, B.; Montout, L.; de Brevern, A.G.; et al. A Dominant Mutation in the Gene Encoding the Erythroid Transcription Factor KLF1 Causes a Congenital Dyserythropoietic Anemia. Am. J. Hum. Genet. 2010, 87, 721–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borg, J.; Papadopoulos, P.; Georgitsi, M.; Gutiérrez, L.; Grech, G.; Fanis, P.; Phylactides, M.; Verkerk, A.J.M.H.; van der Spek, P.J.; Scerri, C.A.; et al. Haploinsufficiency for the Erythroid Transcription Factor KLF1 Causes Hereditary Persistence of Fetal Hemoglobin. Nat. Genet. 2010, 42, 801–805. [Google Scholar] [CrossRef]
- Magor, G.W.; Tallack, M.R.; Gillinder, K.R.; Bell, C.C.; McCallum, N.; Williams, B.; Perkins, A.C. KLF1-Null Neonates Display Hydrops Fetalis and a Deranged Erythroid Transcriptome. Blood 2015, 125, 2405–2417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porcher, C.; Chagraoui, H.; Kristiansen, M.S. SCL/TAL1: A Multifaceted Regulator from Blood Development to Disease. Blood 2017, 129, 2051–2060. [Google Scholar] [CrossRef]
- Sui, X.; Krantz, S.B.; Zhao, Z.J. Stem Cell Factor and Erythropoietin Inhibit Apoptosis of Human Erythroid Progenitor Cells through Different Signalling Pathways: Distinct Roles of PI3K in SCF and EPO Signalling. Br. J. Haematol. 2000, 110, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Nocka, K.; Majumder, S.; Chabot, B.; Ray, P.; Cervone, M.; Bernstein, A.; Besmer, P. Expression of C-Kit Gene Products in Known Cellular Targets of W Mutations in Normal and W Mutant Mice--Evidence for an Impaired c-Kit Kinase in Mutant Mice. Genes Dev. 1989, 3, 816–826. [Google Scholar] [CrossRef] [Green Version]
- Goodman, J.W.; Hall, E.A.; Miller, K.L.; Shinpock, S.G. Interleukin 3 Promotes Erythroid Burst Formation in “Serum-Free” Cultures without Detectable Erythropoietin. Proc. Natl. Acad. Sci. USA 1985, 82, 3291–3295. [Google Scholar] [CrossRef] [Green Version]
- Migliaccio, G.; Migliaccio, A.R.; Adamson, J.W. In Vitro Differentiation of Human Granulocyte/Macrophage and Erythroid Progenitors: Comparative Analysis of the Influence of Recombinant Human Erythropoietin, G-CSF, GM-CSF, and IL-3 in Serum-Supplemented and Serum-Deprived Cultures. Blood 1988, 72, 248–256. [Google Scholar] [CrossRef]
- Lee-Huang, S. Cloning and Expression of Human Erythropoietin CDNA in Escherichia Coli. Proc. Natl. Acad. Sci. USA 1984, 81, 2708–2712. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, K.; Shoemaker, C.; Rudersdorf, R.; Neill, S.D.; Kaufman, R.J.; Mufson, A.; Seehra, J.; Jones, S.S.; Hewick, R.; Fritsch, E.F.; et al. Isolation and Characterization of Genomic and CDNA Clones of Human Erythropoietin. Nature 1985, 313, 806–810. [Google Scholar] [CrossRef]
- Koury, M.J.; Bondurant, M.C. Erythropoietin Retards DNA Breakdown and Prevents Programmed Death in Erythroid Progenitor Cells. Science 1990, 248, 378–381. [Google Scholar] [CrossRef] [PubMed]
- Broudy, V.C.; Lin, N.; Brice, M.; Nakamoto, B.; Papayannopoulou, T. Erythropoietin Receptor Characteristics on Primary Human Erythroid Cells. Blood 1991, 77, 2583–2590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reissmann, K.R. Studies on the Mechanism of Erythropoietic Stimulation in Parabiotic Rats during Hypoxia. Blood 1950, 5, 372–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Liu, X.; Jaenisch, R.; Lodish, H.F. Generation of Committed Erythroid BFU-E and CFU-E Progenitors Does Not Require Erythropoietin or the Erythropoietin Receptor. Cell 1995, 83, 59–67. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wang, L.; Dey, S.; Alnaeeli, M.; Suresh, S.; Rogers, H.; Teng, R.; Noguchi, C. Erythropoietin Action in Stress Response, Tissue Maintenance and Metabolism. Int. J. Mol. Sci. 2014, 15, 10296–10333. [Google Scholar] [CrossRef] [Green Version]
- Ugo, V.; Marzac, C.; Teyssandier, I.; Larbret, F.; Lécluse, Y.; Debili, N.; Vainchenker, W.; Casadevall, N. Multiple Signaling Pathways Are Involved in Erythropoietin-Independent Differentiation of Erythroid Progenitors in Polycythemia Vera. Exp. Hematol. 2004, 32, 179–187. [Google Scholar] [CrossRef]
- Chida, D.; Miura, O.; Yoshimura, A.; Miyajima, A. Role of Cytokine Signaling Molecules in Erythroid Differentiation of Mouse Fetal Liver Hematopoietic Cells: Functional Analysis of Signaling Molecules by Retrovirus-Mediated Expression. Blood 1999, 93, 1567–1578. [Google Scholar] [CrossRef]
- Arcasoy, M.O.; Jiang, X. Co-Operative Signalling Mechanisms Required for Erythroid Precursor Expansion in Response to Erythropoietin and Stem Cell Factor. Br. J. Haematol. 2005, 130, 121–129. [Google Scholar] [CrossRef]
- Notta, F.; Zandi, S.; Takayama, N.; Dobson, S.; Gan, O.I.; Wilson, G.; Kaufmann, K.B.; McLeod, J.; Laurenti, E.; Dunant, C.F.; et al. Distinct Routes of Lineage Development Reshape the Human Blood Hierarchy across Ontogeny. Science 2016, 351, aab2116. [Google Scholar] [CrossRef] [Green Version]
- Grover, A.; Mancini, E.; Moore, S.; Mead, A.J.; Atkinson, D.; Rasmussen, K.D.; O’Carroll, D.; Jacobsen, S.E.W.; Nerlov, C. Erythropoietin Guides Multipotent Hematopoietic Progenitor Cells toward an Erythroid Fate. J. Exp. Med. 2014, 211, 181–188. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Wang, S.; Liu, D.; Gao, C.; Han, Y.; Guo, X.; Qu, X.; Li, W.; Zhang, S.; Geng, J.; et al. EpoR-TdTomato-Cre Mice Enable Identification of EpoR Expression in Subsets of Tissue Macrophages and Hematopoietic Cells. Blood 2021, 138, 1986–1997. [Google Scholar] [CrossRef] [PubMed]
- Sherwood, J.B.; Goldwasser, E. Extraction of Erythropoietin from Normal Kidneys*. Endocrinology 1978, 103, 866–870. [Google Scholar] [CrossRef] [PubMed]
- Hammond, D.; Winnick, S. Paraneoplastic Erythrocytosis and Ectopic Erythropoietins. Ann. N. Y. Acad. Sci. 1974, 230, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Fried, W. The Liver as a Source of Extrarenal Erythropoietin Production. Blood 1972, 40, 671–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naughton, B.A.; Kaplan, S.M.; Roy, M.; Burdowski, A.J.; Gordon, A.S.; Piliero, S.J. Hepatic Regeneration and Erythropoietin Production in the Rat. Science 1977, 196, 301–302. [Google Scholar] [CrossRef] [PubMed]
- Lucarelli, G.; Howard, D.; Stohlman, F. Regulation of Erythropoiesis. XV. Neonatal Erythropoiesis and the Effect of Nephrectomy *. J. Clin. Investig. 1964, 43, 2195–2203. [Google Scholar] [CrossRef]
- Zanjani, E.D.; Poster, J.; Burlington, H.; Mann, L.I.; Wasserman, L.R. Liver as the Primary Site of Erythropoietin Formation in the Fetus. J. Lab. Clin. Med. 1977, 89, 640–644. [Google Scholar]
- Janeway, C.A.J.; Travers, P.; Walport, M.; Shlomchik, M.J. The Components of the Immune System. In Immunobiology: The Immune System in Health and Disease, 5th ed.; Garland Science: New York, NY, USA, 2001. [Google Scholar]
- Friedman, A.D. Transcriptional Control of Granulocyte and Monocyte Development. Oncogene 2007, 26, 6816–6828. [Google Scholar] [CrossRef] [Green Version]
- Huber, R.; Pietsch, D.; Günther, J.; Welz, B.; Vogt, N.; Brand, K. Regulation of Monocyte Differentiation by Specific Signaling Modules and Associated Transcription Factor Networks. Cell. Mol. Life Sci. 2014, 71, 63–92. [Google Scholar] [CrossRef]
- Lawrence, S.M.; Corriden, R.; Nizet, V. The Ontogeny of a Neutrophil: Mechanisms of Granulopoiesis and Homeostasis. Microbiol. Mol. Biol. Rev. 2018, 82, e00057-17. [Google Scholar] [CrossRef] [Green Version]
- DeKoter, R.P.; Singh, H. Regulation of B Lymphocyte and Macrophage Development by Graded Expression of PU.1. Science 2000, 288, 1439–1441. [Google Scholar] [CrossRef] [PubMed]
- Pang, S.H.M.; de Graaf, C.A.; Hilton, D.J.; Huntington, N.D.; Carotta, S.; Wu, L.; Nutt, S.L. PU.1 Is Required for the Developmental Progression of Multipotent Progenitors to Common Lymphoid Progenitors. Front. Immunol. 2018, 9, 1264. [Google Scholar] [CrossRef] [PubMed]
- Bjerregaard, M.D.; Jurlander, J.; Klausen, P.; Borregaard, N.; Cowland, J.B. The in Vivo Profile of Transcription Factors during Neutrophil Differentiation in Human Bone Marrow. Blood 2003, 101, 4322–4332. [Google Scholar] [CrossRef] [Green Version]
- Johansen, L.M.; Iwama, A.; Lodie, T.A.; Sasaki, K.; Felsher, D.W.; Golub, T.R.; Tenen, D.G. C-Myc Is a Critical Target for C/EBPα in Granulopoiesis. Mol. Cell. Biol. 2001, 21, 3789–3806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahl, R.; Walsh, J.C.; Lancki, D.; Laslo, P.; Iyer, S.R.; Singh, H.; Simon, M.C. Regulation of Macrophage and Neutrophil Cell Fates by the PU.1:C/EBPα Ratio and Granulocyte Colony-Stimulating Factor. Nat. Immunol. 2003, 4, 1029–1036. [Google Scholar] [CrossRef]
- Radomska, H.S.; Bassères, D.S.; Zheng, R.; Zhang, P.; Dayaram, T.; Yamamoto, Y.; Sternberg, D.W.; Lokker, N.; Giese, N.A.; Bohlander, S.K.; et al. Block of C/EBPα Function by Phosphorylation in Acute Myeloid Leukemia with FLT3 Activating Mutations. J. Exp. Med. 2006, 203, 371–381. [Google Scholar] [CrossRef]
- Feinberg, M.W.; Wara, A.K.; Cao, Z.; Lebedeva, M.A.; Rosenbauer, F.; Iwasaki, H.; Hirai, H.; Katz, J.P.; Haspel, R.L.; Gray, S.; et al. The Kruppel-like Factor KLF4 Is a Critical Regulator of Monocyte Differentiation. EMBO J. 2007, 26, 4138–4148. [Google Scholar] [CrossRef] [Green Version]
- Hock, H.; Hamblen, M.J.; Rooke, H.M.; Traver, D.; Bronson, R.T.; Cameron, S.; Orkin, S.H. Intrinsic Requirement for Zinc Finger Transcription Factor Gfi-1 in Neutrophil Differentiation. Immunity 2003, 18, 109–120. [Google Scholar] [CrossRef] [Green Version]
- Laslo, P.; Spooner, C.J.; Warmflash, A.; Lancki, D.W.; Lee, H.-J.; Sciammas, R.; Gantner, B.N.; Dinner, A.R.; Singh, H. Multilineage Transcriptional Priming and Determination of Alternate Hematopoietic Cell Fates. Cell 2006, 126, 755–766. [Google Scholar] [CrossRef] [Green Version]
- Schönheit, J.; Kuhl, C.; Gebhardt, M.L.; Klett, F.F.; Riemke, P.; Scheller, M.; Huang, G.; Naumann, R.; Leutz, A.; Stocking, C.; et al. PU.1 Level-Directed Chromatin Structure Remodeling at the Irf8 Gene Drives Dendritic Cell Commitment. Cell Rep. 2013, 3, 1617–1628. [Google Scholar] [CrossRef] [Green Version]
- Burn, G.L.; Foti, A.; Marsman, G.; Patel, D.F.; Zychlinsky, A. The Neutrophil. Immunity 2021, 54, 1377–1391. [Google Scholar] [CrossRef]
- Rosales, C. Neutrophil: A Cell with Many Roles in Inflammation or Several Cell Types? Front. Physiol. 2018, 9, 113. [Google Scholar] [CrossRef] [PubMed]
- Freifeld, A.G.; Bow, E.J.; Sepkowitz, K.A.; Boeckh, M.J.; Ito, J.I.; Mullen, C.A.; Raad, I.I.; Rolston, K.V.; Young, J.-A.H.; Wingard, J.R.; et al. Clinical Practice Guideline for the Use of Antimicrobial Agents in Neutropenic Patients with Cancer: 2010 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2011, 52, e56–e93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pizzo, P.A. Management of Fever in Patients with Cancer and Treatment-Induced Neutropenia. N. Engl. J. Med. 1993, 328, 1323–1332. [Google Scholar] [CrossRef] [PubMed]
- Delaney, C.; Milano, F.; Cicconi, L.; Othus, M.; Becker, P.S.; Sandhu, V.; Nicoud, I.; Dahlberg, A.; Bernstein, I.D.; Appelbaum, F.R.; et al. Infusion of a Non-HLA-Matched Ex-Vivo Expanded Cord Blood Progenitor Cell Product after Intensive Acute Myeloid Leukaemia Chemotherapy: A Phase 1 Trial. Lancet Haematol. 2016, 3, e330–e339. [Google Scholar] [CrossRef] [PubMed]
- Price, T.H.; Boeckh, M.; Harrison, R.W.; McCullough, J.; Ness, P.M.; Strauss, R.G.; Nichols, W.G.; Hamza, T.H.; Cushing, M.M.; King, K.E.; et al. Efficacy of Transfusion with Granulocytes from G-CSF/Dexamethasone-Treated Donors in Neutropenic Patients with Infection. Blood 2015, 126, 2153–2161. [Google Scholar] [CrossRef]
- Desai, P.M.; Brown, J.; Gill, S.; Solh, M.M.; Akard, L.P.; Hsu, J.W.; Ustun, C.; Andreadis, C.; Frankfurt, O.; Foran, J.M.; et al. Open-Label Phase II Prospective, Randomized, Controlled Study of Romyelocel-L Myeloid Progenitor Cells to Reduce Infection during Induction Chemotherapy for Acute Myeloid Leukemia. JCO 2021, 39, 3261–3272. [Google Scholar] [CrossRef]
- De Bruyn, C.; Delforge, A.; Bernier, M.; Bron, D. Ex Vivo Expansion of Neutrophil Precursor Cells from Fresh and Cryopreserved Cord Blood Cells. Cytotherapy 2003, 5, 87–98. [Google Scholar] [CrossRef]
- Hino, M.; Suzuki, K.; Yamane, T.; Sakai, N.; Kubota, H.; Koh, K.R.; Ohta, K.; Hato, F.; Kitagawa, S.; Tatsumi, N. Ex Vivo Expansion of Mature Human Neutrophils with Normal Functions from Purified Peripheral Blood CD34+ Haematopoietic Progenitor Cells. Br. J. Haematol. 2000, 109, 314–321. [Google Scholar] [CrossRef]
- Jie, Z.; Zhang, Y.; Wang, C.; Shen, B.; Guan, X.; Ren, Z.; Ding, X.; Dai, W.; Jiang, Y. Large-Scale Ex Vivo Generation of Human Neutrophils from Cord Blood CD34+ Cells. PLoS ONE 2017, 12, e0180832. [Google Scholar] [CrossRef] [Green Version]
- Kuhikar, R.; Khan, N.; Khare, S.P.; Fulzele, A.; Melinkeri, S.; Kale, V.; Limaye, L. Neutrophils Generated in Vitro from Hematopoietic Stem Cells Isolated from Apheresis Samples and Umbilical Cord Blood Form Neutrophil Extracellular Traps. Stem Cell Res. 2021, 50, 102150. [Google Scholar] [CrossRef]
- Lachmann, N.; Ackermann, M.; Frenzel, E.; Liebhaber, S.; Brennig, S.; Happle, C.; Hoffmann, D.; Klimenkova, O.; Lüttge, D.; Buchegger, T.; et al. Large-Scale Hematopoietic Differentiation of Human Induced Pluripotent Stem Cells Provides Granulocytes or Macrophages for Cell Replacement Therapies. Stem Cell Rep. 2015, 4, 282–296. [Google Scholar] [CrossRef] [Green Version]
- Rodak, B.; Fritsma, G.; Keohane, E. Hematology: Clinical Principals and Applications; Elsevier Health Sciences: St. Louis, MO, USA, 2013. [Google Scholar]
- Egeland, T.; Steen, R.; Quarsten, H.; Gaudernack, G.; Yang, Y.-C.; Thorsby, E. Myeloid Differentiation of Purified CD34+ Cells after Stimulation with Recombinant Human Granulocyte-Monocyte Colony-Stimulating Factor (CSF), Granulocyte-CSF, Monocyte-CSF, and Interleukin-3. Blood 1991, 78, 3192–3199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timmins, N.E.; Palfreyman, E.; Marturana, F.; Dietmair, S.; Luikenga, S.; Lopez, G.; Fung, Y.L.; Minchinton, R.; Nielsen, L.K. Clinical Scale Ex Vivo Manufacture of Neutrophils from Hematopoietic Progenitor Cells. Biotechnol. Bioeng. 2009, 104, 832–840. [Google Scholar] [CrossRef] [PubMed]
- Tura, O.; Barclay, G.R.; Roddie, H.; Davies, J.; Turner, M.L. Optimal Ex Vivo Expansion of Neutrophils from PBSC CD34+ Cells by a Combination of SCF, Flt3-L and G-CSF and Its Inhibition by Further Addition of TPO. J. Transl. Med. 2007, 5, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, K.-D.; Vodyanik, M.; Slukvin, I.I. The Hematopoietic Differentiation and Production of Mature Myeloid Cells from Human Pluripotent Stem Cells. Nat. Protoc. 2011, 6, 296–313. [Google Scholar] [CrossRef]
- Bapat, A.; Keita, N.; Sharma, S. Pan-Myeloid Differentiation of Human Cord Blood Derived CD34+ Hematopoietic Stem and Progenitor Cells. J. Vis. Exp. 2019. [Google Scholar] [CrossRef]
- Hamilton, J.A. Colony-Stimulating Factors in Inflammation and Autoimmunity. Nat. Rev. Immunol. 2008, 8, 533–544. [Google Scholar] [CrossRef]
- Manz, M.G.; Miyamoto, T.; Akashi, K.; Weissman, I.L. Prospective Isolation of Human Clonogenic Common Myeloid Progenitors. Proc. Natl. Acad. Sci. USA 2002, 99, 11872–11877. [Google Scholar] [CrossRef] [Green Version]
- Blalock, W.L.; Weinstein-Oppenheimer, C.; Chang, F.; Hoyle, P.E.; Wang, X.-Y.; Algate, P.A.; Franklin, R.A.; Oberhaus, S.M.; Steelman, L.S.; McCubrey, J.A. Signal Transduction, Cell Cycle Regulatory, and Anti-Apoptotic Pathways Regulated by IL-3 in Hematopoietic Cells: Possible Sites for Intervention with Anti-Neoplastic Drugs. Leukemia 1999, 13, 1109–1166. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, J.A. GM-CSF-Dependent Inflammatory Pathways. Front. Immunol. 2019, 10, 2055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hercus, T.R.; Dhagat, U.; Kan, W.L.T.; Broughton, S.E.; Nero, T.L.; Perugini, M.; Sandow, J.J.; D’Andrea, R.J.; Ekert, P.G.; Hughes, T.; et al. Signalling by the Βc Family of Cytokines. Cytokine Growth Factor Rev. 2013, 24, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Clark, S.C.; Kamen, R. The Human Hematopoietic Colony-Stimulating Factors. Science 1987, 236, 1229–1237. [Google Scholar] [CrossRef] [PubMed]
- Welte, K.; Bonilla, M.A.; Gillio, A.P.; Boone, T.C.; Potter, G.K.; Gabrilove, J.L.; Moore, M.A.; O’Reilly, R.J.; Souza, L.M. Recombinant Human Granulocyte Colony-Stimulating Factor. Effects on Hematopoiesis in Normal and Cyclophosphamide-Treated Primates. J. Exp. Med. 1987, 165, 941–948. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, C.L.; Teng, R.; Wang, H.; Merling, R.K.; Lee, J.; Choi, U.; Koontz, S.; Wright, D.G.; Malech, H.L. Molecular Analysis of Neutrophil Differentiation from Human Induced Pluripotent Stem Cells Delineates the Kinetics of Key Regulators of Hematopoiesis. Stem Cells 2016, 34, 1513–1526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barge, R.M.; de Koning, J.P.; Pouwels, K.; Dong, F.; Löwenberg, B.; Touw, I.P. Tryptophan 650 of Human Granulocyte Colony-Stimulating Factor (G-CSF) Receptor, Implicated in the Activation of JAK2, Is Also Required for G-CSF-Mediated Activation of Signaling Complexes of the P21ras Route. Blood 1996, 87, 2148–2153. [Google Scholar] [CrossRef] [Green Version]
- Corey, S.J.; Burkhardt, A.L.; Bolen, J.B.; Geahlen, R.L.; Tkatch, L.S.; Tweardy, D.J. Granulocyte Colony-Stimulating Factor Receptor Signaling Involves the Formation of a Three-Component Complex with Lyn and Syk Protein-Tyrosine Kinases. Proc. Natl. Acad. Sci. USA 1994, 91, 4683–4687. [Google Scholar] [CrossRef] [Green Version]
- Dwivedi, P.; Greis, K.D. Granulocyte Colony Stimulating Factor Receptor (G-CSFR) Signaling in Severe Congenital Neutropenia, Chronic Neutrophilic Leukemia and Related Malignancies. Exp. Hematol. 2017, 46, 9–20. [Google Scholar] [CrossRef] [Green Version]
- de Koning, J.P.; Soede-Bobok, A.A.; Ward, A.C.; Schelen, A.M.; Antonissen, C.; van Leeuwen, D.; Löwenberg, B.; Touw, I.P. STAT3-Mediated Differentiation and Survival of Myeloid Cells in Response to Granulocyte Colony-Stimulating Factor: Role for the Cyclin-Dependent Kinase Inhibitor P27Kip1. Oncogene 2000, 19, 3290–3298. [Google Scholar] [CrossRef] [Green Version]
- Grishin, A.; Sinha, S.; Roginskaya, V.; Boyer, M.J.; Gomez-Cambronero, J.; Zuo, S.; Kurosaki, T.; Romero, G.; Corey, S.J. Involvement of Shc and Cbl-PI 3-Kinase in Lyn-Dependent Proliferative Signaling Pathways for G-CSF. Oncogene 2000, 19, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, S.E.; Novak, U.; Ziegler, S.F.; Layton, J.E. Distinct Regions of the Granulocyte Colony-Stimulating Factor Receptor Are Required for Tyrosine Phosphorylation of the Signaling Molecules JAK2, Stat3, and P42, P44MAPK. Blood 1995, 86, 3698–3704. [Google Scholar] [CrossRef] [Green Version]
- Ilaria, R.L.; Hawley, R.G.; Van Etten, R.A. Dominant Negative Mutants Implicate STAT5 in Myeloid Cell Proliferation and Neutrophil Differentiation. Blood 1999, 93, 4154–4166. [Google Scholar] [CrossRef] [Green Version]
- Hu, N.; Qiu, Y.; Dong, F. Role of Erk1/2 Signaling in the Regulation of Neutrophil Versus Monocyte Development in Response to G-CSF and M-CSF*. J. Biol. Chem. 2015, 290, 24561–24573. [Google Scholar] [CrossRef] [Green Version]
- Jack, G.D.; Zhang, L.; Friedman, A.D. M-CSF Elevates c-Fos and Phospho-C/EBPalpha(S21) via ERK Whereas G-CSF Stimulates SHP2 Phosphorylation in Marrow Progenitors to Contribute to Myeloid Lineage Specification. Blood 2009, 114, 2172–2180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williamson, E.A.; Williamson, I.K.; Chumakov, A.M.; Friedman, A.D.; Koeffler, H.P. CCAAT/Enhancer Binding Protein ϵ: Changes in Function upon Phosphorylation by P38 MAP Kinase. Blood 2005, 105, 3841–3847. [Google Scholar] [CrossRef] [Green Version]
- Dick, E.P.; Prince, L.R.; Sabroe, I. Ex Vivo-Expanded Bone Marrow CD34+ Derived Neutrophils Have Limited Bactericidal Ability. Stem Cells 2008, 26, 2552–2563. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Sica, A.; Locati, M. New Vistas on Macrophage Differentiation and Activation. Eur. J. Immunol. 2007, 37, 14–16. [Google Scholar] [CrossRef]
- Clanchy, F.I.L.; Holloway, A.C.; Lari, R.; Cameron, P.U.; Hamilton, J.A. Detection and Properties of the Human Proliferative Monocyte Subpopulation. J. Leukoc. Biol. 2006, 79, 757–766. [Google Scholar] [CrossRef] [Green Version]
- Andreesen, R.; Hennemann, B.; Krause, S.W. Adoptive Immunotherapy of Cancer Using Monocyte-Derived Macrophages: Rationale, Current Status, and Perspectives. J. Leukoc. Biol. 1998, 64, 419–426. [Google Scholar] [CrossRef] [PubMed]
- de Souza, V.C.A.; Pereira, T.A.; Teixeira, V.W.; Carvalho, H.; de Castro, M.C.A.B.; D’assunção, C.G.; de Barros, A.F.; Carvalho, C.L.; de Lorena, V.M.B.; Costa, V.M.A.; et al. Bone Marrow-Derived Monocyte Infusion Improves Hepatic Fibrosis by Decreasing Osteopontin, TGF-Β1, IL-13 and Oxidative Stress. World J. Gastroenterol. 2017, 23, 5146–5157. [Google Scholar] [CrossRef]
- Green, D.S.; Ning, F.; Duemler, A.; Myers, T.G.; Trewhitt, K.; Ekwede, I.; McCoy, A.; Houston, N.; Lee, J.; Lipkowitz, S.; et al. Intraperitoneal Monocytes plus IFNs as a Novel Cellular Immunotherapy for Ovarian Cancer: Mechanistic Characterization and Results from a Phase I Clinical Trial. Clin. Cancer Res. 2023, 29, 349–363. [Google Scholar] [CrossRef]
- Brennan, P.N.; MacMillan, M.; Manship, T.; Moroni, F.; Glover, A.; Graham, C.; Semple, S.; Morris, D.M.; Fraser, A.R.; Pass, C.; et al. Study Protocol: A Multicentre, Open-Label, Parallel-Group, Phase 2, Randomised Controlled Trial of Autologous Macrophage Therapy for Liver Cirrhosis (MATCH). BMJ Open 2021, 11, e053190. [Google Scholar] [CrossRef] [PubMed]
- Fraser, A.R.; Pass, C.; Burgoyne, P.; Atkinson, A.; Bailey, L.; Laurie, A.; McGowan, N.W.A.; Hamid, A.; Moore, J.K.; Dwyer, B.J.; et al. Development, Functional Characterization and Validation of Methodology for GMP-Compliant Manufacture of Phagocytic Macrophages: A Novel Cellular Therapeutic for Liver Cirrhosis. Cytotherapy 2017, 19, 1113–1124. [Google Scholar] [CrossRef] [Green Version]
- Chernykh, E.R.; Shevela, E.Y.; Starostina, N.M.; Morozov, S.A.; Davydova, M.N.; Menyaeva, E.V.; Ostanin, A.A. Safety and Therapeutic Potential of M2 Macrophages in Stroke Treatment. Cell Transpl. 2016, 25, 1461–1471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chomarat, P.; Banchereau, J.; Davoust, J.; Karolina Palucka, A. IL-6 Switches the Differentiation of Monocytes from Dendritic Cells to Macrophages. Nat. Immunol. 2000, 1, 510–514. [Google Scholar] [CrossRef]
- Menetrier-Caux, C.; Montmain, G.; Dieu, M.C.; Bain, C.; Favrot, M.C.; Caux, C.; Blay, J.Y. Inhibition of the Differentiation of Dendritic Cells from CD34+ Progenitors by Tumor Cells: Role of Interleukin-6 and Macrophage Colony-Stimulating Factor. Blood 1998, 92, 4778–4791. [Google Scholar] [CrossRef] [PubMed]
- Kamps, A.W.; Hendriks, D.; Smit, J.W.; Vellenga, E. Role of Macrophage Colony-Stimulating Factor in the Differentiation and Expansion of Monocytes and Dendritic Cells from CD34+ Progenitor Cells. Med. Oncol. 1999, 16, 46–52. [Google Scholar] [CrossRef]
- Clanchy, F.I.L.; Hamilton, J.A. The Development of Macrophages from Human CD34+ Haematopoietic Stem Cells in Serum-Free Cultures Is Optimized by IL-3 and SCF. Cytokine 2013, 61, 33–37. [Google Scholar] [CrossRef]
- Stec, M.; Weglarczyk, K.; Baran, J.; Zuba, E.; Mytar, B.; Pryjma, J.; Zembala, M. Expansion and Differentiation of CD14+CD16− and CD14++CD16+ Human Monocyte Subsets from Cord Blood CD34+ Hematopoietic Progenitors. J. Leukoc. Biol. 2007, 82, 594–602. [Google Scholar] [CrossRef] [PubMed]
- Sconocchia, G.; Fujiwara, H.; Rezvani, K.; Keyvanfar, K.; El Ouriaghli, F.; Grube, M.; Melenhorst, J.; Hensel, N.; Barrett, A.J. G-CSF-Mobilized CD34+ Cells Cultured in Interleukin-2 and Stem Cell Factor Generate a Phenotypically Novel Monocyte. J. Leukoc. Biol. 2004, 76, 1214–1219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyauchi, M.; Ito, Y.; Nakahara, F.; Hino, T.; Nakamura, F.; Iwasaki, Y.; Kawagoshi, T.; Koya, J.; Yoshimi, A.; Arai, S.; et al. Efficient Production of Human Neutrophils from IPSCs That Prevent Murine Lethal Infection with Immune Cell Recruitment. Blood 2021, 138, 2555–2569. [Google Scholar] [CrossRef] [PubMed]
- Duweb, A.; Gaiser, A.-K.; Stiltz, I.; El Gaafary, M.; Simmet, T.; Syrovets, T. The SC Cell Line as an in Vitro Model of Human Monocytes. J. Leukoc. Biol. 2022, 112, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Rey-Giraud, F.; Hafner, M.; Ries, C.H. In Vitro Generation of Monocyte-Derived Macrophages under Serum-Free Conditions Improves Their Tumor Promoting Functions. PLoS ONE 2012, 7, e42656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delneste, Y.; Charbonnier, P.; Herbault, N.; Magistrelli, G.; Caron, G.; Bonnefoy, J.-Y.; Jeannin, P. Interferon-γ Switches Monocyte Differentiation from Dendritic Cells to Macrophages. Blood 2003, 101, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Way, K.J.; Dinh, H.; Keene, M.R.; White, K.E.; Clanchy, F.I.L.; Lusby, P.; Roiniotis, J.; Cook, A.D.; Cassady, A.I.; Curtis, D.J.; et al. The Generation and Properties of Human Macrophage Populations from Hemopoietic Stem Cells. J. Leukoc. Biol. 2009, 85, 766–778. [Google Scholar] [CrossRef]
- Becker, S.; Warren, M.K.; Haskill, S. Colony-Stimulating Factor-Induced Monocyte Survival and Differentiation into Macrophages in Serum-Free Cultures. J. Immunol. 1987, 139, 3703–3709. [Google Scholar] [CrossRef]
- Mossadegh-Keller, N.; Sarrazin, S.; Kandalla, P.K.; Espinosa, L.; Stanley, E.R.; Nutt, S.L.; Moore, J.; Sieweke, M.H. M-CSF Instructs Myeloid Lineage Fate in Single Haematopoietic Stem Cells. Nature 2013, 497, 239–243. [Google Scholar] [CrossRef] [Green Version]
- Pixley, F.J.; Stanley, E.R. CSF-1 Regulation of the Wandering Macrophage: Complexity in Action. Trends Cell Biol. 2004, 14, 628–638. [Google Scholar] [CrossRef]
- Borzillo, G.V.; Ashmun, R.A.; Sherr, C.J. Macrophage Lineage Switching of Murine Early Pre-B Lymphoid Cells Expressing Transduced Fms Genes. Mol. Cell Biol. 1990, 10, 2703–2714. [Google Scholar] [CrossRef]
- Montano Almendras, C.P.; Thudium, C.S.; Löfvall, H.; Moscatelli, I.; Schambach, A.; Henriksen, K.; Richter, J. Forced Expression of Human Macrophage Colony-Stimulating Factor in CD34+ Cells Promotes Monocyte Differentiation in Vitro and in Vivo but Blunts Osteoclastogenesis in Vitro. Eur. J. Haematol. 2017, 98, 517–526. [Google Scholar] [CrossRef]
- Husson, H.; Mograbi, B.; Schmid-Antomarchi, H.; Fischer, S.; Rossi, B. CSF-1 Stimulation Induces the Formation of a Multiprotein Complex Including CSF-1 Receptor, c-Cbl, PI 3-Kinase, Crk-II and Grb2. Oncogene 1997, 14, 2331–2338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanagasundaram, V.; Jaworowski, A.; Hamilton, J.A. Association between Phosphatidylinositol-3 Kinase, Cbl and Other Tyrosine Phosphorylated Proteins in Colony-Stimulating Factor-1-Stimulated Macrophages. Biochem. J. 1996, 320, 69–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sampaio, N.G.; Yu, W.; Cox, D.; Wyckoff, J.; Condeelis, J.; Stanley, E.R.; Pixley, F.J. Phosphorylation of CSF-1R Y721 Mediates Its Association with PI3K to Regulate Macrophage Motility and Enhancement of Tumor Cell Invasion. J. Cell Sci. 2011, 124, 2021–2031. [Google Scholar] [CrossRef] [Green Version]
- Stanley, E.R.; Chitu, V. CSF-1 Receptor Signaling in Myeloid Cells. Cold Spring Harb. Perspect. Biol. 2014, 6, a021857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, M.; Hamilton, J.A.; Scholz, G.M.; Masendycz, P.; Macaulay, S.L.; Elsegood, C.L. Phosphatidylinostitol-3 Kinase and Phospholipase C Enhance CSF-1-Dependent Macrophage Survival by Controlling Glucose Uptake. Cell. Signal. 2009, 21, 1361–1369. [Google Scholar] [CrossRef] [PubMed]
- Takeshita, S.; Faccio, R.; Chappel, J.; Zheng, L.; Feng, X.; Weber, J.D.; Teitelbaum, S.L.; Ross, F.P. C-Fms Tyrosine 559 Is a Major Mediator of M-CSF-Induced Proliferation of Primary Macrophages *. J. Biol. Chem. 2007, 282, 18980–18990. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.; Chen, J.; Xiong, Y.; Pixley, F.J.; Yeung, Y.-G.; Stanley, E.R. Macrophage Proliferation Is Regulated through CSF-1 Receptor Tyrosines 544, 559, and 807. J. Biol. Chem. 2012, 287, 13694–13704. [Google Scholar] [CrossRef] [Green Version]
- Bourgin-Hierle, C.; Gobert-Gosse, S.; Thérier, J.; Grasset, M.-F.; Mouchiroud, G. Src-Family Kinases Play an Essential Role in Differentiation Signaling Downstream of Macrophage Colony-Stimulating Factor Receptors Mediating Persistent Phosphorylation of Phospholipase C-Gamma2 and MAP Kinases ERK1 and ERK2. Leukemia 2008, 22, 161–169. [Google Scholar] [CrossRef]
- Gobert Gosse, S.; Bourgin, C.; Liu, W.Q.; Garbay, C.; Mouchiroud, G. M-CSF Stimulated Differentiation Requires Persistent MEK Activity and MAPK Phosphorylation Independent of Grb2-Sos Association and Phosphatidylinositol 3-Kinase Activity. Cell. Signal. 2005, 17, 1352–1362. [Google Scholar] [CrossRef]
- Jackson, T.R.; Ling, R.E.; Roy, A. The Origin of B-Cells: Human Fetal B Cell Development and Implications for the Pathogenesis of Childhood Acute Lymphoblastic Leukemia. Front. Immunol. 2021, 12, 637975. [Google Scholar] [CrossRef]
- Hystad, M.E.; Myklebust, J.H.; Bø, T.H.; Sivertsen, E.A.; Rian, E.; Forfang, L.; Munthe, E.; Rosenwald, A.; Chiorazzi, M.; Jonassen, I.; et al. Characterization of Early Stages of Human B Cell Development by Gene Expression Profiling1. J. Immunol. 2007, 179, 3662–3671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.C.; Jhunjhunwala, S.; Benner, C.; Heinz, S.; Welinder, E.; Mansson, R.; Sigvardsson, M.; Hagman, J.; Espinoza, C.A.; Dutkowski, J.; et al. A Global Network of Transcription Factors, Involving E2A, EBF1 and Foxo1, That Orchestrates B Cell Fate. Nat. Immunol. 2010, 11, 635–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borghesi, L.; Aites, J.; Nelson, S.; Lefterov, P.; James, P.; Gerstein, R. E47 Is Required for V(D)J Recombinase Activity in Common Lymphoid Progenitors. J. Exp. Med. 2005, 202, 1669–1677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cobaleda, C.; Schebesta, A.; Delogu, A.; Busslinger, M. Pax5: The Guardian of B Cell Identity and Function. Nat. Immunol. 2007, 8, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Lemoine, F.M.; Dedhar, S.; Lima, G.M.; Eaves, C.J. Transformation-Associated Alterations in Interactions Between Pre-B Cells and Fibronectin. Blood 1990, 76, 2311–2320. [Google Scholar] [CrossRef] [PubMed]
- Freedman, A.S.; Rhynhart, K.; Nojima, Y.; Svahn, J.; Eliseo, L.; Benjamin, C.D.; Morimoto, C.; Vivier, E. Stimulation of Protein Tyrosine Phosphorylation in Human B Cells after Ligation of the Beta 1 Integrin VLA-4. J. Immunol. 1993, 150, 1645–1652. [Google Scholar] [CrossRef] [PubMed]
- Tse, K.W.K.; Dang-Lawson, M.; Lee, R.L.; Vong, D.; Bulic, A.; Buckbinder, L.; Gold, M.R. B Cell Receptor-Induced Phosphorylation of Pyk2 and Focal Adhesion Kinase Involves Integrins and the Rap GTPases and Is Required for B Cell Spreading*. J. Biol. Chem. 2009, 284, 22865–22877. [Google Scholar] [CrossRef] [Green Version]
- Kraus, H.; Kaiser, S.; Aumann, K.; Bönelt, P.; Salzer, U.; Vestweber, D.; Erlacher, M.; Kunze, M.; Burger, M.; Pieper, K.; et al. A Feeder-Free Differentiation System Identifies Autonomously Proliferating B Cell Precursors in Human Bone Marrow. J. Immunol. 2014, 192, 1044–1054. [Google Scholar] [CrossRef] [Green Version]
- Namen, A.E.; Lupton, S.; Hjerrild, K.; Wignall, J.; Mochizuki, D.Y.; Schmierer, A.; Mosley, B.; March, C.J.; Urdal, D.; Gillis, S.; et al. Stimulation of B-Cell Progenitors by Cloned Murine Interleukin-7. Nature 1988, 333, 571–573. [Google Scholar] [CrossRef]
- Wolf, M.L.; Buckley, J.A.; Goldfarb, A.; Law, C.L.; LeBien, T.W. Development of a Bone Marrow Culture for Maintenance and Growth of Normal Human B Cell Precursors. J. Immunol. 1991, 147, 3324–3330. [Google Scholar] [CrossRef]
- Lee, G.; Namen, A.E.; Gillis, S.; Ellingsworth, L.R.; Kincade, P.W. Normal B Cell Precursors Responsive to Recombinant Murine IL-7 and Inhibition of IL-7 Activity by Transforming Growth Factor-Beta. J. Immunol. 1989, 142, 3875–3883. [Google Scholar] [CrossRef] [PubMed]
- Dittel, B.N.; LeBien, T.W. The Growth Response to IL-7 during Normal Human B Cell Ontogeny Is Restricted to B-Lineage Cells Expressing CD34. J. Immunol. 1995, 154, 58–67. [Google Scholar] [CrossRef]
- Jiang, Q.; Li, W.Q.; Aiello, F.B.; Mazzucchelli, R.; Asefa, B.; Khaled, A.R.; Durum, S.K. Cell Biology of IL-7, a Key Lymphotrophin. Cytokine Growth Factor Rev. 2005, 16, 513–533. [Google Scholar] [CrossRef] [PubMed]
- Dias, S.; Silva, H., Jr.; Cumano, A.; Vieira, P. Interleukin-7 Is Necessary to Maintain the B Cell Potential in Common Lymphoid Progenitors. J. Exp. Med. 2005, 201, 971–979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kikuchi, K.; Lai, A.Y.; Hsu, C.-L.; Kondo, M. IL-7 Receptor Signaling Is Necessary for Stage Transition in Adult B Cell Development through up-Regulation of EBF. J. Exp. Med. 2005, 201, 1197–1203. [Google Scholar] [CrossRef] [Green Version]
- Åhsberg, J.; Tsapogas, P.; Qian, H.; Zetterblad, J.; Zandi, S.; Månsson, R.; Jönsson, J.-I.; Sigvardsson, M. Interleukin-7-Induced Stat-5 Acts in Synergy with Flt-3 Signaling to Stimulate Expansion of Hematopoietic Progenitor Cells. J. Biol. Chem. 2010, 285, 36275–36284. [Google Scholar] [CrossRef] [Green Version]
- Granato, A.; Hayashi, E.A.; Baptista, B.J.A.; Bellio, M.; Nobrega, A. IL-4 Regulates Bim Expression and Promotes B Cell Maturation in Synergy with BAFF Conferring Resistance to Cell Death at Negative Selection Checkpoints. J. Immunol. 2014, 192, 5761–5775. [Google Scholar] [CrossRef] [Green Version]
- Hipp, N.; Symington, H.; Pastoret, C.; Caron, G.; Monvoisin, C.; Tarte, K.; Fest, T.; Delaloy, C. IL-2 Imprints Human Naive B Cell Fate towards Plasma Cell through ERK/ELK1-Mediated BACH2 Repression. Nat. Commun. 2017, 8, 1443. [Google Scholar] [CrossRef] [Green Version]
- Horikawa, K.; Takatsu, K. Interleukin-5 Regulates Genes Involved in B-Cell Terminal Maturation. Immunology 2006, 118, 497–508. [Google Scholar] [CrossRef]
- Heine, G.; Drozdenko, G.; Grün, J.R.; Chang, H.-D.; Radbruch, A.; Worm, M. Autocrine IL-10 Promotes Human B-Cell Differentiation into IgM- or IgG-Secreting Plasmablasts. Eur. J. Immunol. 2014, 44, 1615–1621. [Google Scholar] [CrossRef] [Green Version]
- Ichii, M.; Oritani, K.; Yokota, T.; Nishida, M.; Takahashi, I.; Shirogane, T.; Ezoe, S.; Saitoh, N.; Tanigawa, R.; Kincade, P.W.; et al. Regulation of Human B Lymphopoiesis by the Transforming Growth Factor-β Superfamily in a Newly Established Coculture System Using Human Mesenchymal Stem Cells as a Supportive Microenvironment. Exp. Hematol. 2008, 36, 587–597. [Google Scholar] [CrossRef]
- Ryan, D.H.; Nuccie, B.L.; Abboud, C.N.; Winslow, J.M. Vascular Cell Adhesion Molecule-I and the Integrin VLA-4 Mediate Adhesion of Human B Cell Precursors to Cultured Bone Marrow Adherent Cells. J. Clin. Investig. 1991, 88, 995–1004. [Google Scholar] [CrossRef] [PubMed]
- Ohkawara, J.-I.; Ikebuchi, K.; Fujihara, M.; Sato, N.; Hirayama, F.; Yamaguchi, M.; Mori, K.J.; Sekiguchi, S. Culture System for Extensive Production of CD19+IgM+ Cells by Human Cord Blood CD34+ Progenitors. Leukemia 1998, 12, 764–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, J.S.; McCullar, V.; Punzel, M.; Lemischka, I.R.; Moore, K.A. Single Adult Human CD34+/Lin−/CD38− Progenitors Give Rise to Natural Killer Cells, B-Lineage Cells, Dendritic Cells, and Myeloid Cells. Blood 1999, 93, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Prieyl, J.A.; LeBien, T.W. Interleukin 7 Independent Development of Human B Cells. Proc. Natl. Acad. Sci. USA 1996, 93, 10348–10353. [Google Scholar] [CrossRef] [Green Version]
- Long-Term Culture System for Selective Growth of Human B-Cell Progenitors. Available online: https://www.pnas.org/doi/10.1073/pnas.92.5.1570 (accessed on 3 December 2022).
- Fluckiger, A.-C.; Sanz, E.; Garcia-Lloret, M.; Su, T.; Hao, Q.-L.; Kato, R.; Quan, S.; de la Hera, A.; Crooks, G.M.; Witte, O.N.; et al. In Vitro Reconstitution of Human B-Cell Ontogeny: From CD34+ Multipotent Progenitors to Ig-Secreting Cells. Blood 1998, 92, 4509–4520. [Google Scholar] [CrossRef] [PubMed]
- Scheeren, F.A.; van Lent, A.U.; Nagasawa, M.; Weijer, K.; Spits, H.; Legrand, N.; Blom, B. Thymic Stromal Lymphopoietin Induces Early Human B-Cell Proliferation and Differentiation. Eur. J. Immunol. 2010, 40, 955–965. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Bhattacharya, S.; Zhou, J.; Phadnis-Moghe, A.S.; Crawford, R.B.; Kaminski, N.E. Aryl Hydrocarbon Receptor Activation Suppresses EBF1 and PAX5 and Impairs Human B Lymphopoiesis. J. Immunol. 2017, 199, 3504–3515. [Google Scholar] [CrossRef] [Green Version]
- Weber, B.N.; Chi, A.W.-S.; Chavez, A.; Yashiro-Ohtani, Y.; Yang, Q.; Shestova, O.; Bhandoola, A. A Critical Role for TCF-1 in T-Lineage Specification and Differentiation. Nature 2011, 476, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Yui, M.A.; Rothenberg, E.V. Developmental Gene Networks: A Triathlon on the Course to T Cell Identity. Nat. Rev. Immunol. 2014, 14, 529–545. [Google Scholar] [CrossRef] [Green Version]
- Ormandy, L.A.; Hillemann, T.; Wedemeyer, H.; Manns, M.P.; Greten, T.F.; Korangy, F. Increased Populations of Regulatory T Cells in Peripheral Blood of Patients with Hepatocellular Carcinoma. Cancer Res. 2005, 65, 2457–2464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Themeli, M.; Rivière, I.; Sadelain, M. New Cell Sources for T Cell Engineering and Adoptive Immunotherapy. Cell Stem Cell 2015, 16, 357–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palomero, T.; Lim, W.K.; Odom, D.T.; Sulis, M.L.; Real, P.J.; Margolin, A.; Barnes, K.C.; O’Neil, J.; Neuberg, D.; Weng, A.P.; et al. NOTCH1 Directly Regulates C-MYC and Activates a Feed-Forward-Loop Transcriptional Network Promoting Leukemic Cell Growth. Proc. Natl. Acad. Sci. USA 2006, 103, 18261–18266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deftos, M.L.; Huang, E.; Ojala, E.W.; Forbush, K.A.; Bevan, M.J. Notch1 Signaling Promotes the Maturation of CD4 and CD8 SP Thymocytes. Immunity 2000, 13, 73–84. [Google Scholar] [CrossRef] [Green Version]
- De Obaldia, M.E.; Bell, J.J.; Wang, X.; Harly, C.; Yashiro-Ohtani, Y.; DeLong, J.H.; Zlotoff, D.A.; Sultana, D.A.; Pear, W.S.; Bhandoola, A. T Cell Development Requires Constraint of the Myeloid Regulator C/EBP-α by the Notch Target and Transcriptional Repressor Hes1. Nat. Immunol. 2013, 14, 1277–1284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weng, A.P.; Millholland, J.M.; Yashiro-Ohtani, Y.; Arcangeli, M.L.; Lau, A.; Wai, C.; del Bianco, C.; Rodriguez, C.G.; Sai, H.; Tobias, J.; et al. C-Myc Is an Important Direct Target of Notch1 in T-Cell Acute Lymphoblastic Leukemia/Lymphoma. Genes Dev. 2006, 20, 2096–2109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poellinger, L.; Lendahl, U. Modulating Notch Signaling by Pathway-Intrinsic and Pathway-Extrinsic Mechanisms. Curr. Opin. Genet. Dev. 2008, 18, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, T.M.; Zúñiga-Pflücker, J.C. Induction of T Cell Development from Hematopoietic Progenitor Cells by Delta-like-1 In Vitro. Immunity 2002, 17, 749–756. [Google Scholar] [CrossRef] [Green Version]
- La Motte-Mohs, R.N.; Herer, E.; Zúñiga-Pflücker, J.C. Induction of T-Cell Development from Human Cord Blood Hematopoietic Stem Cells by Delta-like 1 in Vitro. Blood 2005, 105, 1431–1439. [Google Scholar] [CrossRef] [PubMed]
- Awong, G.; Herer, E.; Surh, C.D.; Dick, J.E.; La Motte-Mohs, R.N.; Zúñiga-Pflücker, J.C. Characterization in Vitro and Engraftment Potential in Vivo of Human Progenitor T Cells Generated from Hematopoietic Stem Cells. Blood 2009, 114, 972–982. [Google Scholar] [CrossRef] [Green Version]
- Awong, G.; Herer, E.; La Motte-Mohs, R.N.; Zúñiga-Pflücker, J.C. Human CD8 T Cells Generated in Vitro from Hematopoietic Stem Cells Are Functionally Mature. BMC Immunol. 2011, 12, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, M.; Masuda, K.; Kakugawa, K.; Kawamoto, H.; Mugishima, H.; Katsura, Y. Quantification of Progenitors Capable of Generating T Cells in Human Cord Blood. Eur. J. Haematol. 2008, 80, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Mohtashami, M.; Shah, D.K.; Nakase, H.; Kianizad, K.; Petrie, H.T.; Zúñiga-Pflücker, J.C. Direct Comparison of Dll1- and Dll4-Mediated Notch Activation Levels Shows Differential Lymphomyeloid Lineage Commitment Outcomes. J. Immunol. 2010, 185, 867–876. [Google Scholar] [CrossRef] [Green Version]
- Hozumi, K.; Mailhos, C.; Negishi, N.; Hirano, K.; Yahata, T.; Ando, K.; Zuklys, S.; Holländer, G.A.; Shima, D.T.; Habu, S. Delta-like 4 Is Indispensable in Thymic Environment Specific for T Cell Development. J. Exp. Med. 2008, 205, 2507–2513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plum, J.; De Smedt, M.; Defresne, M.-P.; Leclercq, G.; Vandekerckhove, B. Human CD34+ Fetal Liver Stem Cells Differentiate to T Cells in a Mouse Thymic Microenvironment. Blood 1994, 84, 1587–1593. [Google Scholar] [CrossRef] [Green Version]
- Poznansky, M.C.; Evans, R.H.; Foxall, R.B.; Olszak, I.T.; Piascik, A.H.; Hartman, K.E.; Brander, C.; Meyer, T.H.; Pykett, M.J.; Chabner, K.T.; et al. Efficient Generation of Human T Cells from a Tissue-Engineered Thymic Organoid. Nat. Biotechnol. 2000, 18, 729–734. [Google Scholar] [CrossRef]
- Chung, B.; Montel-Hagen, A.; Ge, S.; Blumberg, G.; Kim, K.; Klein, S.; Zhu, Y.; Parekh, C.; Balamurugan, A.; Yang, O.O.; et al. Engineering the Human Thymic Microenvironment to Support Thymopoiesis In Vivo. Stem Cells 2014, 32, 2386–2396. [Google Scholar] [CrossRef] [Green Version]
- Seet, C.S.; He, C.; Bethune, M.T.; Li, S.; Chick, B.; Gschweng, E.H.; Zhu, Y.; Kim, K.; Kohn, D.B.; Baltimore, D.; et al. Generation of Mature T Cells from Human Hematopoietic Stem and Progenitor Cells in Artificial Thymic Organoids. Nat. Methods 2017, 14, 521–530. [Google Scholar] [CrossRef]
- Ikawa, T.; Kawamoto, H.; Wright, L.Y.T.; Murre, C. Long-Term Cultured E2A-Deficient Hematopoietic Progenitor Cells Are Pluripotent. Immunity 2004, 20, 349–360. [Google Scholar] [CrossRef] [Green Version]
- Gehre, N.; Nusser, A.; von Muenchow, L.; Tussiwand, R.; Engdahl, C.; Capoferri, G.; Bosco, N.; Ceredig, R.; Rolink, A.G. A Stromal Cell Free Culture System Generates Mouse Pro-T Cells That Can Reconstitute T-Cell Compartments in Vivo. Eur. J. Immunol. 2015, 45, 932–942. [Google Scholar] [CrossRef]
- Reimann, C.; Six, E.; Dal-Cortivo, L.; Schiavo, A.; Appourchaux, K.; Lagresle-Peyrou, C.; de Chappedelaine, C.; Ternaux, B.; Coulombel, L.; Beldjord, K.; et al. Human T-Lymphoid Progenitors Generated in a Feeder-Cell-Free Delta-Like-4 Culture System Promote T-Cell Reconstitution in NOD/SCID/γc−/− Mice. Stem Cells 2012, 30, 1771–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simons, L.; Ma, K.; de Chappedelaine, C.; Moiranghtem, R.D.; Elkaim, E.; Olivré, J.; Susini, S.; Appourchaux, K.; Reimann, C.; Sadek, H.; et al. Generation of Adult Human T-Cell Progenitors for Immunotherapeutic Applications. J. Allergy Clin. Immunol. 2018, 141, 1491–1494.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoyama, K.; Delaney, C.; Varnum-Finney, B.; Kohn, A.D.; Moon, R.T.; Bernstein, I.D. The Interaction of the Wnt and Notch Pathways Modulates Natural Killer versus T Cell Differentiation. Stem Cells 2007, 25, 2488–2497. [Google Scholar] [CrossRef]
- Huijskens, M.J.A.J.; Walczak, M.; Koller, N.; Briedé, J.J.; Senden-Gijsbers, B.L.M.G.; Schnijderberg, M.C.; Bos, G.M.J.; Germeraad, W.T.V. Technical Advance: Ascorbic Acid Induces Development of Double-Positive T Cells from Human Hematopoietic Stem Cells in the Absence of Stromal Cells. J. Leukoc. Biol. 2014, 96, 1165–1175. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Langley, M.A.; Singh, J.; Edgar, J.M.; Mohtashami, M.; Zúñiga-Pflücker, J.C.; Zandstra, P.W. Progenitor T-Cell Differentiation from Hematopoietic Stem Cells Using Delta-like-4 and VCAM-1. Nat. Methods 2017, 14, 531–538. [Google Scholar] [CrossRef]
- Trotman-Grant, A.C.; Mohtashami, M.; De Sousa Casal, J.; Martinez, E.C.; Lee, D.; Teichman, S.; Brauer, P.M.; Han, J.; Anderson, M.K.; Zúñiga-Pflücker, J.C. DL4-Μbeads Induce T Cell Lineage Differentiation from Stem Cells in a Stromal Cell-Free System. Nat. Commun. 2021, 12, 5023. [Google Scholar] [CrossRef]
- Edgar, J.M.; Michaels, Y.S.; Zandstra, P.W. Multi-Objective Optimization Reveals Time- and Dose-Dependent Inflammatory Cytokine-Mediated Regulation of Human Stem Cell Derived T-Cell Development. NPJ Regen. Med. 2022, 7, 1–13. [Google Scholar] [CrossRef]
- Hernández-López, C.; Varas, A.; Sacedón, R.; Jiménez, E.; Muñoz, J.J.; Zapata, A.G.; Vicente, A. Stromal Cell–Derived Factor 1/CXCR4 Signaling Is Critical for Early Human T-Cell Development. Blood 2002, 99, 546–554. [Google Scholar] [CrossRef]
- Janas, M.L.; Varano, G.; Gudmundsson, K.; Noda, M.; Nagasawa, T.; Turner, M. Thymic Development beyond β-Selection Requires Phosphatidylinositol 3-Kinase Activation by CXCR4. J. Exp. Med. 2009, 207, 247–261. [Google Scholar] [CrossRef] [Green Version]
- Ross, S.H.; Cantrell, D.A. Signaling and Function of Interleukin-2 in T Lymphocytes. Annu. Rev. Immunol. 2018, 36, 411–433. [Google Scholar] [CrossRef]
- Van Acker, H.H.; Anguille, S.; Willemen, Y.; Van den Bergh, J.M.; Berneman, Z.N.; Lion, E.; Smits, E.L.; Van Tendeloo, V.F. Interleukin-15 Enhances the Proliferation, Stimulatory Phenotype, and Antitumor Effector Functions of Human Gamma Delta T Cells. J. Hematol. Oncol. 2016, 9, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefort, N.; Benne, C.; Lelièvre, J.D.; Dorival, C.; Balbo, M.; Sakano, S.; Coulombel, L.; Lévy, Y. Short Exposure to Notch Ligand Delta-4 Is Sufficient to Induce T-Cell Differentiation Program and to Increase the T Cell Potential of Primary Human CD34+ Cells. Exp. Hematol. 2006, 34, 1720–1729. [Google Scholar] [CrossRef] [PubMed]


| Cell Source | Method | Cells Generated | Reference |
|---|---|---|---|
| PB CD34+ | human/canine serum | >95% CD41+ megakaryocytes | [134] |
| PB CD34+ | THPO | >79% CD41+ megakaryocytes | [135] |
| UCB CD34+ | THPO, SCF, IL-6, FLT3L | >80% CD41+, >50% CD41+CD42b+ megakaryocytes | [136] |
| UCB CD34+ | Phase I: THPO, SCF, FLT3L, human stromal cells; Phase II: THPO, SCF, FLT3L, IL-11, human stromal cells; Phase III: THPO, SCF, FLT3L, IL-11 | >0.5% CD41+ megakaryocytes, 4.2 × 105 platelets/starting CD34+ cell | [137] |
| UCB CD34+ | Phase I: THPO, SCF, IL-3; Phase II: THPO, IL-11 | >85% CD41+CD42b+ megakaryocytes, 1.9 × 104 platelets/starting CD34+ cell | [139] |
| UCB CD34+ | Phase I: THPO, SCF, FLT3L, IL-3, SR1; Phase II: THPO, SCF, IL-3, IL-6, IL-11, GM-CSF | >70% CD41+CD42b+ megakaryocytes | [138] |
| UCB CD34+ | Phase I: THPO, SCF, FLT3L, IL-6, SR1, C433, VPA; Phase II: THPO, SCF, IL-3, IL-6, IL-11, GM-CSF | >70% CD41+CD42b+ megakaryocytes | [140] |
| Cell Source | Method | Cells Generated | Reference |
|---|---|---|---|
| UCB CD34+ | Phase I: SCF, THPO, FLT3L, hydrocortisone; Phase II: SCF, EPO, IGF-I, hydrocortisone; Phase III: EPO, IGF1, hydrocortisone | >80% CD71+CD235a+ erythrocytes | [163] |
| UCB and PB CD34+ | Phase I: SCF, IL3, EPO, hydrocortisone; Phase II: EPO in stromal cell co-culture; Phase III: stromal cell co-culture | >80% CD71+ erythrocytes | [164] |
| UCB CD34+ | Phase I: SCF, EPO, IL-3 or SCF, EPO, IL-3, VEGF, IGF-II; Phase II: SCF, EPO; Phase III: SCF, EPO(reduced); Phase IV: plasmanate and mifepristone | >90% CD235a+RhD+ erythrocytes | [165] |
| PB CD34+ | Phase I: SCF, IL3, EPO, hydrocortisone; Phase II: SCF, EPO; Phase III: EPO | >85% CD71+CD235a+ erythrocytes | [166] |
| UCB CD34+ | Phase I: SCF, THPO, FLT3L; Phase II: SCF, FLT3L, EPO, IL3, GM-CSF; Phase III: SCF, FLT3L, EPO, IL3; Phase IV: SCF, EPO | >90% CD235a+ erythrocytes | [167] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bozhilov, Y.K.; Hsu, I.; Brown, E.J.; Wilkinson, A.C. In Vitro Human Haematopoietic Stem Cell Expansion and Differentiation. Cells 2023, 12, 896. https://doi.org/10.3390/cells12060896
Bozhilov YK, Hsu I, Brown EJ, Wilkinson AC. In Vitro Human Haematopoietic Stem Cell Expansion and Differentiation. Cells. 2023; 12(6):896. https://doi.org/10.3390/cells12060896
Chicago/Turabian StyleBozhilov, Yavor K., Ian Hsu, Elizabeth J. Brown, and Adam C. Wilkinson. 2023. "In Vitro Human Haematopoietic Stem Cell Expansion and Differentiation" Cells 12, no. 6: 896. https://doi.org/10.3390/cells12060896
APA StyleBozhilov, Y. K., Hsu, I., Brown, E. J., & Wilkinson, A. C. (2023). In Vitro Human Haematopoietic Stem Cell Expansion and Differentiation. Cells, 12(6), 896. https://doi.org/10.3390/cells12060896