

Methanolic Extract of Phoenix Dactylifera Confers Protection against Experimental Diabetic Cardiomyopathy through Modulation of Glucolipid Metabolism and Cardiac Remodeling




Abstract
:1. Introduction
2. Materials and Methods
2.1. Extract Preparation of Phoenix dactylifera L.
2.1.1. Qualitative Estimation of Bioactive Phytochemicals
2.1.2. Quantitative Estimation of Bioactive Phytochemicals
Total Phenolic Constituents (mg of Gallic Acid Equivalent/g Dry Weight of Plant)
Total Flavonoid Content (mg of Quercetin Equivalent/g Dry Weight of Plant)
2.1.3. Determination of Antihyperglycemic Activity by α-Amylase Inhibition Assay
2.1.4. Identification and Quantification of Phenolic Constituents by HPLC
2.2. In Vivo Experimental Trial
2.2.1. Animal Housing and Induction of Diabetes
2.2.2. Biochemical Parameters
Determination of Glucolipid Profile
Determination of Myocardial Profile
2.2.3. Tissue Analyses: Gene Expression and Histopathology
2.2.4. Effect of P-Coumaric Acid on In Vitro Cell Culture of Human Cardiac Fibroblasts
- i.
- Vehicle control (VC): DMEM containing 0.1% DMSO;
- ii.
- Induction (TGF-β + D): stimulation with TGF-β (10 ng/mL) in the presence of hyperglycemia (25 mmol/L D-glucose) for 48 h;
- iii.
- Treatment (PCA): Stimulation with TGF-β in the presence of hyperglycemia (25 mmol/L D-glucose) followed by treatment with PCA (30 μΜ; 48 h). PCA was dissolved in DMSO prior to dilution in DMEM, with a final concentration in experiments not exceeding 0.1% (v/v).
2.2.5. Statistical Analysis
3. Results
3.1. In Vitro
3.1.1. Qualitative and Quantitative Analysis of Phoenix dactylifera
3.1.2. Quantitative Estimation of Antihyperglycemic Activity and HPLC
3.2. In Vivo
3.2.1. Phoenix dactylifera Modulates Glucolipid Metabolic Alterations in DCM
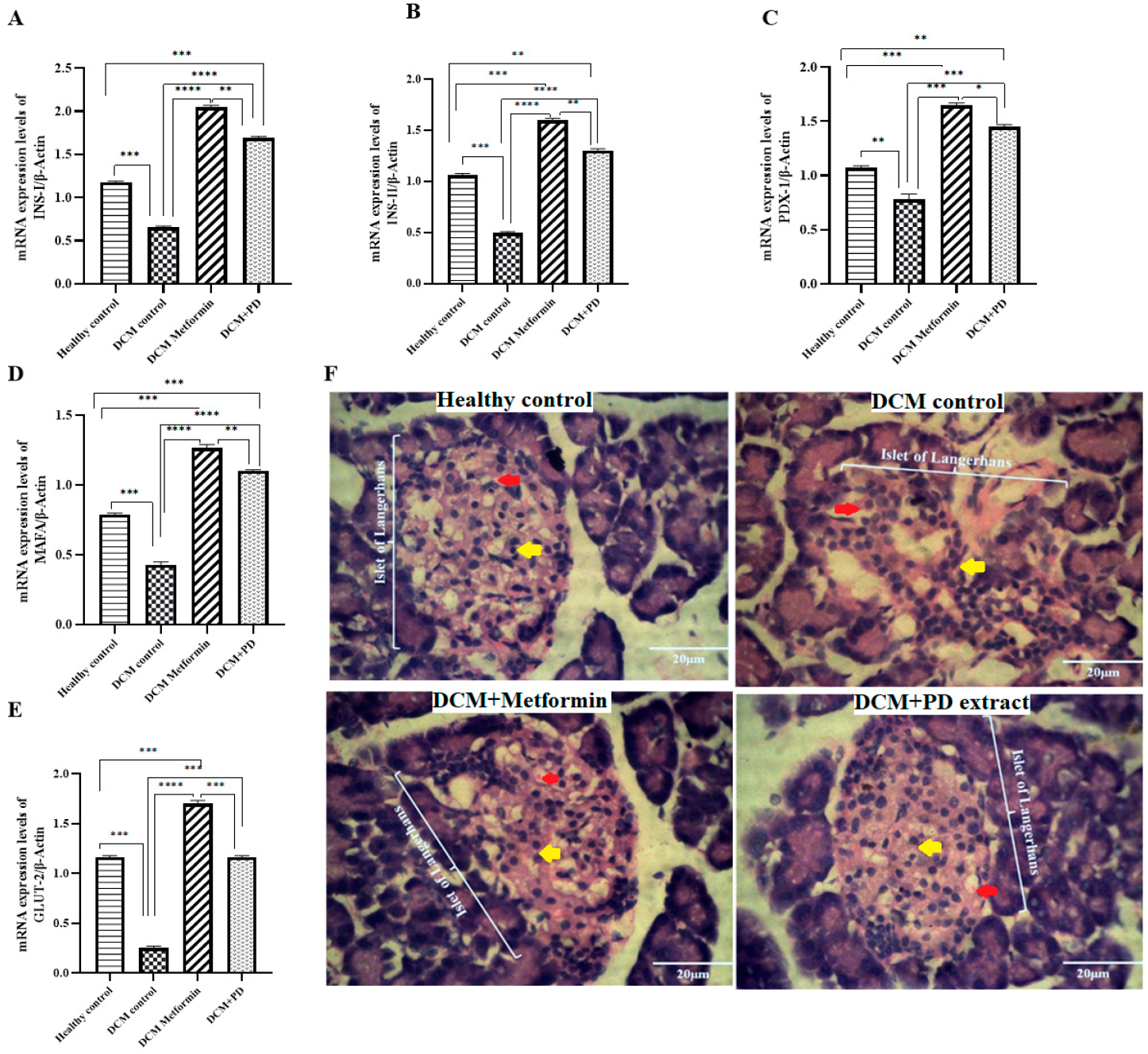
Phoenix dactylifera Upregulates Transcriptional Factors and Genes Involved in Insulin Signaling Pathway and Preserves Pancreatic Structural Integrity in DCM
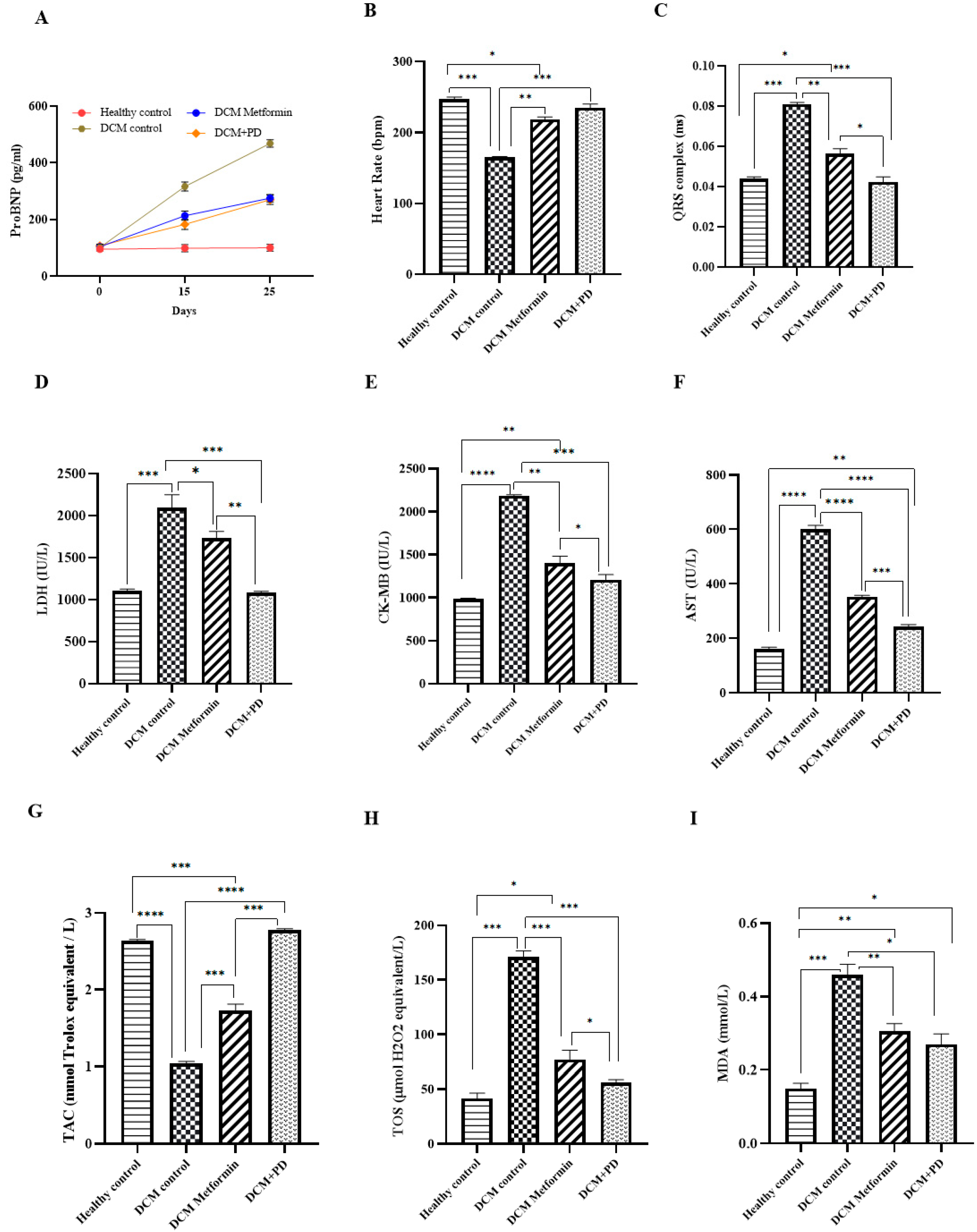
3.2.2. Phoenix dactylifera Reduces Myocardial Oxidative Stress and Protects against Injury
3.2.3. Phoenix dactylifera Confers Cardioprotection against DCM by Downregulating Pro-Inflammatory and Pro-Fibrotic Genes’ Expressions
3.2.4. p-Coumaric Acid Inhibits Hyperglycemia and TGF-β Stimulated Fibrosis in Human Cardiac Fibroblasts
4. Discussion
5. Limitations of the Study
6. Therapeutic Potential of PD Extract and Future Recommendations
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aneja, A.; Tang, W.W.; Bansilal, S.; Garcia, M.J.; Farkouh, M. Diabetic cardiomyopathy: Insights into pathogenesis, diagnostic challenges, and therapeutic options. Am. J. Med. 2008, 121, 748–757. [Google Scholar] [CrossRef]
- Cosentino, F.; Grant, P.J.; Aboyans, V.; Bailey, C.J.; Ceriello, A.; Delgado, V.; Federici, M.; Filippatos, G.; Grobbee, D.E.; Hansen, T. 2019 ESC Guidelines on diabetes, pre-diabetes, and cardiovascular diseases developed in collaboration with the EASD: The Task Force for diabetes, pre-diabetes, and cardiovascular diseases of the European Society of Cardiology (ESC) and the European Association for the Study of Diabetes (EASD). Rev. Esp. Cardiol. 2020, 41, 255–323. [Google Scholar]
- Gupta, S.K.; Dongare, S.; Mathur, R.; Mohanty, I.R.; Srivastava, S.; Mathur, S.; Nag, T. Genistein ameliorates cardiac inflammation and oxidative stress in streptozotocin-induced diabetic cardiomyopathy in rats. Mol. Cell. Biochem. 2015, 408, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Ernande, L.; Bergerot, C.; Rietzschel, E.R.; De Buyzere, M.L.; Thibault, H.; PignonBlanc, P.G.; Croisille, P.; Ovize, M.; Groisne, L.; Moulin, P. Diastolic dysfunction in patients with type 2 diabetes mellitus: Is it really the first marker of diabetic cardiomyopathy? J. Am. Soc. Echocardiogr. 2011, 24, 1268–1275.e1. [Google Scholar] [CrossRef]
- Ma, Z.-G.; Yuan, Y.-P.; Xu, S.-C.; Wei, W.-Y.; Xu, C.-R.; Zhang, X.; Wu, Q.-Q.; Liao, H.-H.; Ni, J.; Tang, Q.-Z. CTRP3 attenuates cardiac dysfunction, inflammation, oxidative stress and cell death in diabetic cardiomyopathy in rats. Diabetologia 2017, 60, 1126–1137. [Google Scholar] [CrossRef]
- Wen, H.-L.; Liang, Z.-S.; Zhang, R.; Yang, K. Anti-inflammatory effects of triptolide improve left ventricular function in a rat model of diabetic cardiomyopathy. Cardiovasc. Diabetol. 2013, 12, 50. [Google Scholar] [CrossRef]
- Salvatore, T.; Pafundi, P.C.; Galiero, R.; Albanese, G.; Di Martino, A.; Caturano, A.; Vetrano, E.; Rinaldi, L.; Sasso, F. The diabetic cardiomyopathy: The contributing pathophysiological mechanisms. Front. Med. 2021, 8, 695792. [Google Scholar] [CrossRef]
- von Bibra, H.; Sutton, M.S.J. Diastolic dysfunction in diabetes and the metabolic syndrome: Promising potential for diagnosis and prognosis. Diabetologia 2010, 53, 1033–1045. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Yan, D.; Li, S.; Liu, S.; Zeng, F.; Cheung, C.W.; Liu, H.; Irwin, M.G.; Huang, H.; Xia, Z.; et al. Allopurinol reduces oxidative stress and activates Nrf2/p62 to attenuate diabetic cardiomyopathy in rats. J. Cell. Mol. Med. 2020, 24, 1760–1773. [Google Scholar] [CrossRef]
- Huynh, K.; Bernardo, B.C.; McMullen, J.R.; Ritchie, R.H. Diabetic cardiomyopathy: Mechanisms and new treatment strategies targeting antioxidant signaling pathways. Pharmacol. Ther. 2014, 142, 375–415. [Google Scholar] [CrossRef]
- Abiola, T. Assessment of the Antidiabetic Potential of the Ethanolic Extract of Date Palm (Phoenix dactylifera) Seed in Alloxan-Induced Diabetic Rats. J. Diabetes Metab. 2018, 9, 1000784. [Google Scholar]
- Sulieman, A.M.; Elhafise, I.A.; Abdelrahim, A. Comparative study on five Sudanese date (Phoenix dactylifera L.) fruit cultivars. Food Nutr. Sci. 2012, 3, 1245–1251. [Google Scholar]
- Bakr Abdu, S. The protective role of AJWA date against the hepatotoxicity induced by ochratoxin A. Egypt. J. Nat. Toxins 2011, 8, 1. [Google Scholar]
- Azeem, S.; Khan, U.; Liaquat, A. The increasing rate of diabetes in Pakistan: A silent killer. Ann. Med. Surg. 2022, 79, 103901. [Google Scholar] [CrossRef] [PubMed]
- Zahoor, T.; Saeed, F.; Ahmad, A. Antinutritional factors in some date palm (Phoenix dactylifera L.) varieties grown in Pakistan. Internet J. Food Saf. 2011, 13, 386–390. [Google Scholar]
- Khalid, S.; Arshad, M.; Saad, B.; Imran, M.; Ul, H.B.; Ain, T.T.; Shahid, M.Z.; Atif, M.; Ullah, M.I.; Ahmed, S. Cardio protective effects of dates. Int. J. Biosci. 2020, 16, 110–123. [Google Scholar]
- Al-Farsi, M.; Alasalvar, C.; Morris, A.; Baron, M.; Shahidi, F. Comparison of antioxidant activity, anthocyanins, carotenoids, and phenolics of three native fresh and sun-dried date (Phoenix dactylifera L.) varieties grown in Oman. J. Agric. Food Chem. 2005, 53, 7592–7599. [Google Scholar] [CrossRef] [PubMed]
- Rosenblat, M.; Volkova, N.; Borochov-Neori, H.; Judeinstein, S.; Aviram, M.J.F. Anti-atherogenic properties of date vs. pomegranate polyphenols: The benefits of the combination. Food Funct. 2015, 6, 1496–1509. [Google Scholar] [CrossRef] [PubMed]
- Hasson, S.; Al-Shaqsi, M.; Albusaidi, J.; Al-Balushi, M.; Hakkim, F.; Aleemallah, G.; Al-Jabri, A. Influence of different cultivars of Phoenix dactylifera L-date fruits on blood clotting and wound healing. Asian Pac. J. Trop. Biomed. 2018, 8, 371–376. [Google Scholar] [CrossRef]
- Khan, F.; Ahmed, F.; Pushparaj, P.N.; Abuzenadah, A.; Kumosani, T.; Barbour, E.; AlQahtani, M.; Gauthaman, K. Ajwa date (Phoenix dactylifera L.) extract inhibits human breast adenocarcinoma (MCF7) cells in vitro by inducing apoptosis and cell cycle arrest. PLoS ONE 2016, 11, e0158963. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Hamid, N.A.; Abas, F.; Ismail, I.S.; Shaari, K.; Lajis, N.H. Influence of different drying treatments and extraction solvents on the metabolite profile and nitric oxide inhibitory activity of Ajwa dates. J. Food Sci. 2015, 80, H2603–H2611. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.L.; Bag, G.C. Phytochemical analysis and determination of total phenolics content in water extracts of three species of Hedychium. Int. J. PharmTech Res. 2013, 5, 1516–1521. [Google Scholar]
- Kumar, J.A.; Sharma, P.; Sikarwar, M.S. Evaluation of anti-inflammatory and anti-pyretic activity of total alcoholic extract of Croton sparsiflorus and its fractions. J. Pharm. Res. 2010, 3, 1149–1151. [Google Scholar]
- Mustafa, I.; Faisal, M.N.; Hussain, G.; Muzaffar, H.; Imran, M.; Ijaz, M.U.; Sohail, M.U.; Iftikhar, A.; Shaukat, A.; Anwar, H. Efficacy of Euphorbia helioscopia in context to a possible connection between antioxidant and antidiabetic activities: A comparative study of different extracts. BMC Complement. Med. Ther. 2021, 21, 62. [Google Scholar] [CrossRef] [PubMed]
- de Paz, G.A.; Raggio, J.; Gómez-Serranillos, M.; Palomino, O.; González-Burgos, E.; Carretero, M.; Crespo, A.J.J.o.P.; Analysis, B. HPLC isolation of antioxidant constituents from Xanthoparmelia sp. J. Pharm. Biomed. Anal. 2010, 53, 165–171. [Google Scholar] [CrossRef]
- Cheng, H.S.; Ton, S.H.; Phang, S.C.W.; Tan, J.B.L.; Kadir, K.A. Increased susceptibility of post-weaning rats on high-fat diet to metabolic syndrome. J. Adv. Res. 2017, 8, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, I.; Anwar, H.; Irfan, S.; Muzaffar, H.; Ijaz, M.U. Attenuation of carbohydrate metabolism and lipid profile by methanolic extract of Euphorbia helioscopia and improvement of beta cell function in a type 2 diabetic rat model. BMC Complement. Med. Ther. 2022, 22, 23. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Lv, X.-Y.; Li, J.; Xu, Z.-G.; Chen, L. The characterization of high-fat diet and multiple low-dose streptozotocin induced type 2 diabetes rat model. J. Diabetes Res. 2008, 2008, 704045. [Google Scholar] [CrossRef]
- Jankauskas, S.S.; Varzideh, F.; Mone, P.; Kansakar, U.; Di Lorenzo, F.; Lombardi, A.; Santulli, G. Interleukin-1 receptor associated kinase 2 is a functional downstream regulator of complement factor D that controls mitochondrial fitness in diabetic cardiomyopathy. Mil. Med. Res. 2024, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- El-Mousalamy, A.M.; Hussein, A.A.M.; Mahmoud, S.A.; Abdelaziz, A.; Shaker, G.J.B.; Access, P.O. Aqueous and methanolic extracts of palm date seeds and fruits (Phoenix dactylifera) protects against diabetic nephropathy in type II diabetic rats. Biochem. Physiol. Open Access 2016, 5, 5. [Google Scholar]
- Oza, M.J.; Kulkarni, Y.A. Formononetin alleviates diabetic cardiomyopathy by inhibiting oxidative stress and upregulating SIRT1 in rats. Asian Pac. J. Trop. Biomed. 2020, 10, 254–262. [Google Scholar]
- Albadawy, R.; Agwa, S.H.; Khairy, E.; Saad, M.; El Touchy, N.; Othman, M.; Matboli, M.J.B. Clinical Significance of HSPD1/MMP14/ITGB1/miR-6881-5P/Lnc-SPARCL1-1: 2 RNA Panel in NAFLD/NASH Diagnosis: Egyptian Pilot Study. Biomedicines 2021, 9, 1248. [Google Scholar] [CrossRef] [PubMed]
- Oršolić, N.; Landeka Jurčević, I.; Đikić, D.; Rogić, D.; Odeh, D.; Balta, V.; Perak Junaković, E.; Terzić, S.; Jutrić, D.J.A. Effect of propolis on diet-induced hyperlipidemia and atherogenic indices in mice. Antioxidants 2019, 8, 156. [Google Scholar] [CrossRef] [PubMed]
- Roy, D.C.; Wang, T.-F.; Mallick, R.; Carrier, M.; Mollanji, E.; Liu, P.; Zhang, L.; Hawken, S.; Wells, P. Growth differentiation factor-15, high-sensitivity cardiac troponin T, and N-terminal pro-B-type natriuretic peptide for predicting risk of venous thromboembolism in ambulatory cancer patients receiving chemotherapy. Thromb. Haemost. 2022, 122, 1169–1176. [Google Scholar] [CrossRef] [PubMed]
- Munir, I.; Mehmood, M.H.; Faisal, N.; Hanif, M.; Naseer, D. Pharmacological evaluation of Vitis vinifera and Zingiber zerumbet on electrocardiographic, biochemical alterations and antioxidant status in isoproterenol-induced myocardial infarction in rats. Pak. J. Pharm. Sci. 2021, 34, 1929–1937. [Google Scholar] [PubMed]
- Taye, A.; Abouzied, M.M.; Mohafez, O.M.M. Tempol ameliorates cardiac fibrosis in streptozotocin-induced diabetic rats: Role of oxidative stress in diabetic cardiomyopathy. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2013, 386, 1071–1080. [Google Scholar] [CrossRef] [PubMed]
- Maqbool, J.; Anwar, H.; Iqbal, J.; Rasul, A.; Imran, A.; Ahmad Malik, S.; Shabbir, A.; Ijaz, F.; Sajid, F.; Akram, R. Methanolic extract of Fennel (Foeniculum vulgare) escalates functional restoration following a compression injury to the sciatic nerve in a mouse model. Food Sci. Nutr. 2021, 9, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.A.; Abdelhalim, M.A.K.; Al-Ayed, M.S.; Alhomida, A.S. Effect of gold nanoparticles on glutathione and malondialdehyde levels in liver, lung and heart of rats. Saudi J. Biol. Sci. 2012, 19, 461–464. [Google Scholar] [CrossRef] [PubMed]
- Imran, A.; Xiao, L.; Ahmad, W.; Anwar, H.; Rasul, A.; Imran, M.; Aziz, N.; Razzaq, A.; Arshad, M.U.; Shabbir, A. Foeniculum vulgare (Fennel) promotes functional recovery and ameliorates oxidative stress following a lesion to the sciatic nerve in mouse model. J. Food Biochem. 2019, 43, e12983. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, D.; Bozkurt, B.; Ramasubbu, K.; Deswal, A. Relationship of hemoglobin A1C and mortality in heart failure patients with diabetes. J. Am. Coll. Cardiol. 2009, 54, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Garimella, S.; Seshayamma, V.; Rao, H.J.; Kumar, S.; Kumar, U.; Saheb, S. Effect of metformin on lipid profile of type II diabetes. Int. J. Integr. Med. Sci. 2016, 3, 449–453. [Google Scholar] [CrossRef]
- Hirano, T. Pathophysiology of diabetic dyslipidemia. J. Atheroscler. Thromb. 2018, 25, 771–782. [Google Scholar] [CrossRef]
- He, L. Metformin and systemic metabolism. Trends Pharmacol. Sci. 2020, 41, 868–881. [Google Scholar] [CrossRef] [PubMed]
- Sekhon-Loodu, S.; Rupasinghe, H.V. Evaluation of antioxidant, antidiabetic and antiobesity potential of selected traditional medicinal plants. Front. Nutr. 2019, 6, 53. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, F.E.F.; Aly, M.A.; Muhammad, A.; Raeesa, A.M.; Amr, S.M.; Ali, H.A.R.; Hasem, H.D. Hypoglycemic effects of date seed extract. Possible mechanism of action, and potential therapeutic implications. Saudi Med. J. 2013, 34, 1125–1132. [Google Scholar]
- Mia, M.A.-T.; Mosaib, M.G.; Khalil, M.I.; Islam, M.A.; Gan, S.H. Potentials and safety of date palm fruit against diabetes: A critical review. Foods 2020, 9, 1557. [Google Scholar] [CrossRef] [PubMed]
- Alqarni, M.M.; Osman, M.A.; Al-Tamimi, D.S.; Gassem, M.A.; Al-Khalifa, A.S.; Al-Juhaimi, F.; Mohamed Ahmed, I. Antioxidant and antihyperlipidemic effects of Ajwa date (Phoenix dactylifera L.) extracts in rats fed a cholesterol-rich diet. J. Food Biochem. 2019, 43, e12933. [Google Scholar] [CrossRef]
- Lee, D.; Kim, K.H.; Lee, J.; Hwang, G.S.; Lee, H.L.; Hahm, D.-H.; Huh, C.K.; Lee, S.C.; Lee, S.; Kang, K.S. Protective effect of cirsimaritin against streptozotocin-induced apoptosis in pancreatic beta cells. J. Pharm. Pharmacol. 2017, 69, 875–883. [Google Scholar] [CrossRef] [PubMed]
- Shirwaikar, A.; Rajendran, K.; Barik, R. Effect of aqueous bark extract of Garuga pinnata Roxb. in streptozotocin-nicotinamide induced type-II diabetes mellitus. J. Ethnopharmacol. 2006, 107, 285–290. [Google Scholar] [CrossRef]
- Yang, X.; Xu, Z.; Zhang, C.; Cai, Z.; Zhang, J. Metformin, beyond an insulin sensitizer, targeting heart and pancreatic β cells. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2017, 1863, 1984–1990. [Google Scholar] [CrossRef] [PubMed]
- Esfandiari, E.; Dorali, A.; Ghanadian, M.; Rashidi, B.; Mostafavi, F.S. Protective and therapeutic effects of Phoenix dactylifera leaf extract on pancreatic β-cells and diabetic parameters in streptozotocin-induced diabetic rats. Comp. Clin. Pathol. 2020, 29, 847–854. [Google Scholar] [CrossRef]
- Coşkun, R.; Çelik, A.İ.; Coşgun, M.S.; Dündar, C.; Türkoğlu, M.; Süleyman, H.J.K.H.J. Phoenix dactylifera L. Tree Fruit Exerts Cardioprotective Effect against DoxorubicinInduced Heart Damage in Rats via Inhibition of Oxidative Stress. Koşuyolu Heart J. 2022, 25, 193–199. [Google Scholar] [CrossRef]
- Sandhu, R.; Bahler, R.C. Prevalence of QRS prolongation in a community hospital cohort of patients with heart failure and its relation to left ventricular systolic dysfunction. Am. J. Cardiol. 2004, 93, 244–246. [Google Scholar] [CrossRef]
- Bai, Y.; Wang, J.; Shan, H.; Lu, Y.; Zhang, Y.; Luo, X.; Yang, B.; Wang, Z. Sphingolipid metabolite ceramide causes metabolic perturbation contributing to HERG K+ channel dysfunction. Cell. Physiol. Biochem. 2007, 20, 429–440. [Google Scholar] [CrossRef] [PubMed]
- Diniz Vilela, D.; Gomes Peixoto, L.; Teixeira, R.R.; Belele Baptista, N.; Carvalho Caixeta, D.; Vieira de Souza, A.; Machado, H.L.; Pereira, M.N.; Sabino-Silva, R.; Espindola, F.; et al. The role of metformin in controlling oxidative stress in muscle of diabetic rats. Oxid. Med. Cell. Longev. 2016, 2016, 6978625. [Google Scholar] [CrossRef] [PubMed]
- Saddi, A.A.; Mohamed, A.M.; Shaikh, A.M. Prophylactic mechanisms of Cucumis melo var. flexuosus and Phoenix dactylifera fruit extracts against diabetic cardiomyopathy in streptozotocin induced diabetic rats. Pak. J. Pharm. Sci. 2018, 31, 699. [Google Scholar] [PubMed]
- Godugu, K.; El-Far, A.H.; Al Jaouni, S.; Mousa, S.A. Nanoformulated Ajwa (Phoenix dactylifera) bioactive compounds improve the safety of doxorubicin without compromising its anticancer efficacy in breast cancer. Molecules 2020, 25, 2597. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, P.; Yeo, J.L.; Brady, E.M.; McCann, G.P. Role of inflammation in diabetic cardiomyopathy. Ther. Adv. Endocrinol. Metab. 2022, 13, 20420188221083530. [Google Scholar] [CrossRef] [PubMed]
- Chandel, N.S.; Trzyna, W.C.; McClintock, D.S.; Schumacker, P.T. Role of oxidants in NF-κB activation and TNF-α gene transcription induced by hypoxia and endotoxin. J. Immunol. 2000, 165, 1013–1021. [Google Scholar] [CrossRef] [PubMed]
- Wynn, T.A.; Ramalingam, T.R. Mechanisms of fibrosis: Therapeutic translation for fibrotic disease. Nat. Med. 2012, 18, 1028–1040. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Si, J.; Qi, K.; Li, D.; Li, T.; Tang, Y.; Ji, E.; Yang, S. Chronic intermittent hypoxia aggravated diabetic cardiomyopathy through LKB1/AMPK/Nrf2 signaling pathway. PLoS ONE 2024, 19, e0296792. [Google Scholar] [CrossRef] [PubMed]
- Hmidani, A.; Bourkhis, B.; Khouya, T.; Ramchoun, M.; Filali-Zegzouti, Y.; Alem, C. Phenolic profile and anti-inflammatory activity of four Moroccan date (Phoenix dactylifera L.) seed varieties. Heliyon 2020, 6, e03436. [Google Scholar]
- Al Alawi, R.; Alhamdani, M.S.S.; Hoheisel, J.D.; Baqi, Y. Antifibrotic and tumor microenvironment modulating effect of date palm fruit (Phoenix dactylifera L.) extracts in pancreatic cancer. Biomed. Pharmacother. 2020, 121, 109522. [Google Scholar] [CrossRef] [PubMed]
- Jankauskas, S.S.; Kansakar, U.; Varzideh, F.; Wilson, S.; Mone, P.; Lombardi, A.; Gambardella, J.; Santulli, G. Heart failure in diabetes. Metabolism 2021, 125, 154910. [Google Scholar] [CrossRef] [PubMed]
- Dyntar, D.; Eppenberger-Eberhardt, M.; Maedler, K.; Pruschy, M.; Eppenberger, H.M.; Spinas, G.A.; Donath, M.Y. Glucose and palmitic acid induce degeneration of myofibrils and modulate apoptosis in rat adult cardiomyocytes. Diabetes 2001, 50, 2105–2113. [Google Scholar] [CrossRef] [PubMed]
- Francalanci, P.; Gallo, P.; Bernucci, P.; Silver, M.D.; d’Amati, G. The pattern of desmin filaments in myocardial disarray. Hum. Pathol. 1995, 26, 262–266. [Google Scholar] [CrossRef]
- Garcia-Canadilla, P.; ook, A.C.; Mohun, T.J.; Oji, O.; Schlossarek, S.; Carrier, L.; McKenna, W.J.; Moon, J.C.; Captur, G. Myoarchitectural disarray of hypertrophic cardiomyopathy begins pre-birth. J. Anat. 2019, 235, 962–976. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Xie, H.; Liu, Y.; Gao, P.; Yang, X.; Shen, Z. Effect of metformin on all-cause and cardiovascular mortality in patients with coronary artery diseases: A systematic review and an updated meta-analysis. Cardiovasc. Diabetol. 2019, 18, 96. [Google Scholar] [CrossRef] [PubMed]
- Saafi-Ben Salah, E.B.; El Arem, A.; Louedi, M.; Saoudi, M.; Elfeki, A.; Zakhama, A.; Najjar, M.F.; Hammami, M.; Achour, L. Antioxidant-rich date palm fruit extract inhibits oxidative stress and nephrotoxicity induced by dimethoate in rat. J. Physiol. Biochem. 2012, 68, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Miki, T.; Yuda, S.; Kouzu, H.; Miura, T. Diabetic cardiomyopathy: Pathophysiology and clinical features. Heart Fail. Rev. 2013, 18, 149–166. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | NCBI Gene ID | Nucleotide Sequence | GC% | Annealing Temperature | Product Size |
|---|---|---|---|---|---|
| INS-I (Rattus norvegicus) | 16333 | FWD: CTGGGAAATGAGGTGGAAAA | 45.00 | 56.65 | 98 |
| Rev: TCCACAAGCCACGCTTCTG | 57.89 | 56.89 | |||
| INS-II (Rattus norvegicus) | 16334 | FWD: AGAAAGGTTTGGTACCTGGAATAGAGC | 44.44 | 53.23 | 121 |
| Rev: GTAGAAGAAGCTTCGCTCCCCACA | 54.16 | 53.76 | |||
| PDX-1 (Rattus norvegicus) | 29535 | FWD: CGTAGTAGCGGGACAACGAG | 60.00 | 55.25 | 71 |
| Rev: CCCGAGGTTACGGCACAAT | 57.89 | 55.08 | |||
| MAFA (Rattus norvegicus) | 366949 | FWD: GACCTGATGAAGTTCGAGGTG | 52.38 | 53.39 | 90 |
| Rev: GGGCGTCGAGGATAGCGA | 66.67 | 56.29 | |||
| GLUT-2 (Rattus norvegicus) | 6514 | FWD: GGCATGTTTTTCTGTGCCGT | 50 | 59.97 | 278 |
| Rev: AAGAACACGTAAGGCCCGAG | 55 | 60.04 | |||
| β-ACTIN (Rattus norvegicus) | 60 | FWD: CGAGTACAACCTTCTTGCAGC | 59.54 | 52.38 | 71 |
| Rev: TATCGTCATCCATGGCGAACTG | 60.55 | 50 | |||
| TGF-β (Rattus norvegicus) | 7040 | FWD: GGCCAGGAACTACCCCTGTG | 65 | 62.20 | 254 |
| Rev: GCCTAGGAGGCATAGAGCGA | 60 | 60.90 | |||
| NF-ĸB (Rattus norvegicus) | 4790 | FWD: CACCGGATCTTTCCCGCCA | 63.16 | 62.32 | 155 |
| Rev: GAGTTTCAGACGCCCGCCTA | 60 | 62.22 | |||
| TNF-α (Rattus norvegicus) | 7124 | FWD: TGAAGCCCGGCTGATGGTAG | 60 | 61.97 | 216 |
| Rev: TGTGGCCATGTCGGTTCACT | 55 | 62 | |||
| GAPDH (Rattus norvegicus) | 2597 | FWD: TCCTGTTCGACAGTCAGCCG | 60 | 62.14 | 70 |
| Rev: CCCCATGGTGTCTGAGCGAT | 60 | 61.98 | |||
| CTGF (Homo sapiens) | 1490 | FWD: TGGGAGTACGGATGCACTTT | 50 | 59.02 | 119 |
| Rev: TACCAATGACAACGCCTC | 50 | 54.63 | |||
| α-SMA (Homo sapiens) | 59 | FWD: CGTTACTACTGCTGAGCGTGA | 52.38 | 60.14 | 164 |
| Rev: AACGTTCATTTCCGATGGTG | 45 | 56.73 | |||
| β-Actin (Homo sapiens) | 60 | FWD: ACAGAGCCTCGCCTTTGCC | 63.16 | 62.91 | 71 |
| Rev: ACAGAGCCTCGCCTTTGCC | 63.16 | 62.91 |
| Phytochemical Analyses | ||
|---|---|---|
| In Vitro Qualitative Phytochemical Analysis | ||
| Phytochemicals | Test | Detection Level |
| Carbohydrates | Fehling’s test | ++ |
| Benedict’s test | ++ | |
| Proteins | Xanthopoietic | ++ |
| Alkaloids | Hager’s reagent | + |
| Flavonoids | NaOH | + |
| Phenols and tannins | Lead acetate test | +++ |
| Steroids and terpenoids | Chloroform | + |
| In Vitro Quantitative Phytochemical Analysis | ||
| Phoenix dactylfera L. | TFC (mgQE/g) | TPC (mgGAE/g) |
| 4.82 ± 0.13 | 58.33 ± 3.24 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nawaz, L.; Grieve, D.J.; Muzaffar, H.; Iftikhar, A.; Anwar, H. Methanolic Extract of Phoenix Dactylifera Confers Protection against Experimental Diabetic Cardiomyopathy through Modulation of Glucolipid Metabolism and Cardiac Remodeling. Cells 2024, 13, 1196. https://doi.org/10.3390/cells13141196
Nawaz L, Grieve DJ, Muzaffar H, Iftikhar A, Anwar H. Methanolic Extract of Phoenix Dactylifera Confers Protection against Experimental Diabetic Cardiomyopathy through Modulation of Glucolipid Metabolism and Cardiac Remodeling. Cells. 2024; 13(14):1196. https://doi.org/10.3390/cells13141196
Chicago/Turabian StyleNawaz, Laaraib, David J. Grieve, Humaira Muzaffar, Arslan Iftikhar, and Haseeb Anwar. 2024. "Methanolic Extract of Phoenix Dactylifera Confers Protection against Experimental Diabetic Cardiomyopathy through Modulation of Glucolipid Metabolism and Cardiac Remodeling" Cells 13, no. 14: 1196. https://doi.org/10.3390/cells13141196