Abstract
Endothelial cells (ECs) are vital structural units of the cardiovascular system possessing two principal distinctive properties: heterogeneity and plasticity. Endothelial heterogeneity is defined by differences in tissue-specific endothelial phenotypes and their high predisposition to modification along the length of the vascular bed. This aspect of heterogeneity is closely associated with plasticity, the ability of ECs to adapt to environmental cues through the mobilization of genetic, molecular, and structural alterations. The specific endothelial cytoarchitectonics facilitate a quick structural cell reorganization and, furthermore, easy adaptation to the extrinsic and intrinsic environmental stimuli, known as the epigenetic landscape. ECs, as universally distributed and ubiquitous cells of the human body, play a role that extends far beyond their structural function in the cardiovascular system. They play a crucial role in terms of barrier function, cell-to-cell communication, and a myriad of physiological and pathologic processes. These include development, ontogenesis, disease initiation, and progression, as well as growth, regeneration, and repair. Despite substantial progress in the understanding of endothelial cell biology, the role of ECs in healthy conditions and pathologies remains a fascinating area of exploration. This review aims to summarize knowledge and concepts in endothelial biology. It focuses on the development and functional characteristics of endothelial cells in health and pathological conditions, with a particular emphasis on endothelial phenotypic and functional heterogeneity.
Keywords:
endothelial cells; microcirculation; macrocirculation; development; WNT; VGF; NOTCH; BMP; TGF; EMT; EndMT; cancer; endothelial turnover; endothelial regeneration 1. Introduction
ECs are the most ubiquitous cells of the body, with a wide variety of functions. The total surface area of the ECs in the human body is approximately 3000–6000 m2, corresponding to 1 to 6 × 1013 cells [1]. The endothelium is a multifunctional tissue, an internal organ, that performs protective (hematoencephalic, intestinal, renal, and hepatic barriers), receptor, transport (oxygen, nutrients, growth factors), secretion, absorption, and synthesis functions [2]. It is also involved in general and local tissue homeostasis, vascular tone and blood pressure regulation, hemostasis, angiogenesis, innate and adaptive immune responses, inflammation, vascular remodeling, tumorigenesis, and initiation of cardiovascular diseases [3,4,5,6,7].
ECs are classified within the simple squamous epithelium group, reflecting their arrangement in a single layer of flat cells. Although the EC population demonstrates heterogeneity across different organs, it shares unique histological characteristics within the same vascular bed region and originates from a common embryonic source. Regularly manifested unifying morphological endothelial features include:
- (1)
- A flat and elongated teardrop or cobblestone-like shape with a polygonal cell outline. The endothelial shape facilitates the blood flow dynamics (laminar blood flow due to the ECs not being studded with ciliae), and it is crucial for selective barrier formation and adaptation to environmental changes [8,9,10].
- (2)
- Possession of different junction types. The junctional complexes play a crucial role in the formation and stabilization of the endothelial barrier and in regulating permeability between the blood and surrounding tissues [11,12,13,14,15]. Barrier specialization is directly related to junction types that specifically affect the endothelial layer’s permeability, endothelial cell growth, apoptosis, and intracellular signal transmission [16,17]. Cell–cell junctional contacts impact the rearrangement of the endothelial layer through different signaling pathways (e.g., PI3Ka/MYPT1/MLCP [18], Notch, Rho GTPase, Wnt/beta-catenin, and Hippo pathways [19,20,21,22];
- (3)
- The presence of a strong extracellular matrix basal lamina, reinforcing the ECs and contributing to their trophic functions, such as survival, proliferation, and differentiation [23,24,25]. The basal lamina is associated with the vessel networks that are essential for maintaining local tissue homeostasis, post-transcriptional modifications, and possibly even the regulation of gene expression [26];
- (4)
- Polarity of the endothelial cell surface. Herein, we refer to endothelial asymmetry in the structural organization and components of the apical and basolateral surfaces of ECs. This property is one of the keys to maintaining the endothelial barrier, facilitating cell migration and vectorial transport of biomolecules and enabling proper signaling. The polarity is closely related to the positioning and distribution of the centrosomes and Golgi apparatus, centrosomal microtubule proteins (Atp6ap2, Tacc3-ch-Tog, Cep41, CG-Nap), and non-centrosomal microtubule proteins (Campsap2, Par-6, Pkc3) [27,28,29,30,31,32,33,34,35,36], as well as to the sorting of proteins and asymmetric protein surface distribution [37].
Plasticity is another vital characteristic related to the reversible modification of the endothelial phenotype. This quality is multifaceted and based on the complex molecular interplay between epigenetics and genetics. Adaptational changes entail different mechanisms, including the endothelial-to-mesenchymal transition process (EndMT) [38,39], changes in transcription factors released from chromatin versus altered import or affected “extra-local“ synthesis processing [40,41], feedback to environmental cues [42,43], and metabolic modulation [44,45].
Plasticity and heterogeneity are fundamental endothelial features that determine the role of ECs in various physiological processes and predispose endothelia to participate in numerous pathological tissue alterations. Plasticity, as a predictor of endothelial heterogeneity, enables ECs to adapt their phenotype and function to the specific local requirements of their microenvironment. A complex interplay between the local microenvironmental signals and epigenetic cues regulates the unifying EC characteristics mentioned earlier. ECs are inherently changeable, which underpins their heterogeneity. The specialization and function of ECs, regional variation in blood supply, and unique yet differing endothelial cell neighborhoods in different organs dictate the distribution of distinct endothelial cells and their associated phenotypes throughout the organism [46].
This review seeks to illuminate the role of endothelial plasticity as a predictor of heterogeneity in vascular health and disease. Its actuality lies in an attempt to expose the key mechanisms behind endothelial heterogeneity and adaptation under physiological and pathological conditions.
2. Endothelial Development
2.1. Endothelial Origin, Cell Specification, and Vessel Organization
ECs are mesodermal derivates [47,48]. Endothelial development is closely connected to hematopoiesis and includes three consecutive waves of cellular formation. The first transient pool (or first wave) of ECs appears during extraembryonic hematopoiesis in the yolk sac. Between days 12 and 15 of gestation, the human yolk sac’s hemangioblasts start forming blood islands [49]. The blood islands consist of internal and external layers [50]. The external layer gives rise to the angioblasts, the multipotent progenitors of ECs, while the internal layer contains hematopoietic precursors of the future blood cells [51]. The second wave (extraembryonic hematopoiesis) includes the migration of extraembryonic precursor cells into the embryo and the formation of intraembryonic hemogenic endothelial cells (IHECs) in the aorta–gonad–mesonephros and liver [52]. The third wave (intraembryonic wave) is characterized by the production of hematopoietic stem cells from the IHECs and further colonization of the hematopoietic organs, e.g., aorta–gonad–mesonephros region, liver, and bone marrow [53]. The embryonic ECs do not demonstrate any conventional, fundamental functions (protective, signal transmission, and interstitial regulation), but take part in the structural organization and reorganization of the vascular system, including vasculogenesis (de novo formation of primitive blood vessels from angioblasts), angiogenesis (growth and development of new vessels from the pre-existing vascular network), and remodeling of the existing vessels [54]. Vasculogenesis is characterized by the mobilization and specification of the angioblasts with further differentiation into endothelial cells and proliferation, migration, coalescence, and formation of vascular plexuses (Table 1) [55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76].
Angiogenesis is a more general physiological process occurring not only in embryogenesis, but also in postnatal life. Arterial and venous angiogenesis can include various processes (endothelial sprouting and intussusceptive microvascular growth) that include morphological changes of the initial vascular plexuses, resulting in vascular network establishment (Table 3) [77,78,79]. In postnatal life, vascular network formation mainly occurs through angiogenesis (including arterio/venogenesis) or remodeling of pre-existing vessels, including collateral vessel formation [80].
Lymphatic vessel development (lymphangiogenesis) from the vein endothelium typically happens in prenatal life. In many studied cases, postnatal lymphatic vessel formation is pathological (Table 2) [58,65,72,81,82,83,84,85,86]. In 2004, a new scheme of the differentiation of the lymphatic vasculature in prenatal life was proposed by G. Oliver [87]. According to him, prenatal development of lymphatic vessels includes four phases: (1) lymphatic ECs competence (ability to respond to stimuli and initiation of LYVE 1 expression); (2) lymphatic ECs bias (formation of lymphatic structures with the involvement of Prox1 transcription factor); (3) lymphatic ECs specification (initiation of specific lymphatic marker expression, e.g., podoplanin (T1α) and neuropilin2 (NRP2); and (4) lymphatic vessel maturation (spreading of the lymphatic vessels in the body) (Table 2) [87,88]. Lymphangiogenesis is crucial for the embryo’s healthy development, clearance of interstitial metabolites, and circulatory liver metabolite supply in the adult, as well as for wound repair and regeneration of the uterine mucosa during the menstruation cycle [67,78]. Tumor metastasis, moreover, is often critically dependent on the lymphatic conduits as well as on blood vascular remodeling [89,90].

Table 2.
Morpho-functional classification of lymphatic ECs.
Table 2.
Morpho-functional classification of lymphatic ECs.
| Lymphangiogenesis Growth and Development of Lymphatic Vessels in Prenatal and Postnatal Life | ||
|---|---|---|
| Mechanism and phases | Signaling and transcriptional regulators | Markers of endothelial differentiation |
| I. Prenatal lymphangiogenesis | 1. ETS domain protein [84] 2. SOXF factors: SOX7, SOX17, and SOX18 [81,83,85,86] 3. Vascular endothelial growth factor-C (VEGF-C)/vascular endothelial growth factor-F (VEGF-F)/vascular endothelial growth factor-D (VEGF-D) [59,81,82,83,84,85,86,87,88] 4. Prox1 [58,81,83,84,85,86] 5. Forkhead box C2 (FOXC2) [81,82,84,85,86] 6. Rho family GTPase (RAC-1) [85] 7. Tyrosine kinase Syk [81,85] 8. SLP 76 [81] 9. Phosphatase-Cγ2(PLCγ2) [84] 10. Semaphorin 3F (SEMA3F) [81,84] 11. Chicken ovalbumin upstream promoter transcription factor (COUP-TFII) [58,83,85] | Platelet endothelial cell adhesion (PECAM-1) [84,85] CD34 [80,81,84,87] CXCR4 (through CXCL12 stimulation) [58,85] LYVE-1 [80,82,83,84,85,86,87] Podoplanin [58,80,81,84,87] Vascular endothelial growth factor receptor 3 (VEGFR-3) [80,81,82,83,84,85,86,87] PROX-1 (Prospero homeobox protein) [58,81,83,84,85,86] CD44 [80,87] |
| (1) Classical Lymphatic vascular development: centrifugal sprouting from primary lymph sacs arises from embryonic cardinal veins (starts at E9.5-10.5 in mice and 6–7 weeks in humans) | ||
| A. Budding and sprouting lymphatic ECs from cardinal veins [82,83,84,85,86] | ||
| B. Centrifugal migration lymphatic ECs [82,83,84,85,86] | ||
| C. Proliferation lymphatic ECs and generation of a one-way network of capillaries [82,83,84,85,86] | ||
| (2) Lymphatic vascular differentiation, according to G. Oliver (2004) [87] | ||
| A. Lymphatic competence (lymphatic ECs from a vein at the E9.0-9.5 receive the ability to answer specific lymphatic-inducing signals) [87] | ||
| B. Lymphatic bias (determination of the lymphatic ECs fate, approx. E9.0-10.5) [87] | ||
| C. Lymphatic specification (lymphatic ECs differentiate into the desired phenotype independently from microenvironmental cues, approx. E10.5.-14.5) [87] | ||
| D. LEC differentiation (maturation and separation of lymphatic vessels, approx. E14.4. postnatal life) [87] | ||
| II. Postnatal lymphangiogenesis | ||
| closely related to such pathological processes as implantation and tumorogenesis (phases similar to angiogenesis) | ||
| A. Sprouting lymphangiogenesis: occurs with or without lymph flow in pre-existing vessels [58] | ||
| B. Intussusceptive lymphangiogenesis is dependent on lymph flow [58] | ||
2.2. Pathways and Factors Involved in the Regulation of Endothelial Development
Diversified signaling pathways and transcription factors regulate both vasculogenesis and angiogenesis (Table 1), including WNT (wingless), NOTCH (“notched wings”), bone morphogenetic protein (BMP), fibroblast growth factor (FGF), and vascular endothelial growth factor (VEGF) [91]. A wide range of transcription factors participates in the regulation of endothelial development, including the E twenty-six family of transcription factors or ETS family (ETS1, ETS2, FLI 1, ERG, ETV1, ETV2, ELK3); GATA transcription factors; VEGF-A, vascular endothelial growth factor receptor 2 (VEGFR2, Flk1/KDR); stem cell leukemia/or T-cell acute lymphocytic leukemia-1 (SCL/TAL1); SMADS proteins; BMP; Lim domain only 2 (LMO2); retinoic acid; and transforming growth factor β (TGFβ) [46,92,93,94].
WNT signaling belongs to the ancient pathway responsible for endothelial development in embryogenesis and consists of canonical (β-catenin associated signaling) and non-canonical (β-catenin independent, including planar cell polarity (PCP) and WNT/Ca2+ signaling) pathways [95]. The canonical and non-canonical pathway initiation starts with interactions of the extracellular WNT ligand with the N-terminal domain of one of the seven transmembrane-span frizzled receptors (FZD receptors) [96,97]. The WNT signaling pathway plays an essential role in embryonic vascular development and the pathogenesis of a wide range of diseases, including cancer, osteoarthritis, muscle pathologies, and kidney diseases [98,99,100,101,102]. The WNT/β-catenin-associated pathway is a primary regulator of brain development, neural differentiation of pluripotent stem cells, and neurodegenerative disorders [103]. WNT/β-catenin signaling, as reported by transcription factor 7-like 2 (TCF7L2)-dependent transcription, is involved in the onset of mental disorders and behavioral deficits [104]. WNT signaling participates in angiogenesis through interaction with secreted frizzled-related protein (SFRP), which is vital for vascular growth. SFRP1 is expressed by all cultured endothelial cells and favors vascular development in all angiogenic models, whereas SFRP2 is mainly involved in angiogenic responses through the nuclear translocation of nuclear factors of activated T cells (NFATs) [105]. SFRP1 and SFRP2 are agonists of the WNT signaling pathway, while SFRP4 appears to antagonize the canonical WNT signaling [106].
NOTCH signaling is an intracellular pathway that controls cell-type specification, differentiation, proliferation, regeneration, and tissue development [107]. NOTCH regulates cell fate, switching in a growing sprout from tip to stalk (meaning initiation “endothelial nascent sprout” from tip and stalk endothelial cells) [108] in the cardiovascular system and the vascular tree. It maintains the proper arterial, venous, and capillary organization; endothelial phenotypic identity is proposed to be regulated by shear stress influence on the NOTCH1 mechanosignaling cascade in ECs [109]. Moreover, the postnatal lymphangiogenesis and differentiation from veins occur under NOTCH signaling control [110]. Its principal components are ligands, receptors, DNA binding protein, and downstream transcribed genes (regulated expression) [111]. NOTCH is composed of four transmembrane receptors (NOTCH 1,2,3,4) and NOTCH ligands (JAGGED1, JAGGED2, DELTA-LIKE 1,2,3, SERRATE). The receptor–ligand interaction drives the NOTCH receptor’s proteolytic cleavage with subsequent liberation of the NOTCH intracellular domain (NICD) into the cytoplasm, activating canonical and non-canonical signaling mechanisms [112]. The canonical pathway is characterized by the migration of the NICD into the cellular nucleus, where it regulates hairy and enhancer of split-1 (HES1) and hairy/enhancer-of-split-1 related to YRPW motif protein 1 (HEY1) target genes by forming transcriptional complexes with the recombination signal-binding proteins for immunoglobulin kappa J region (RBPJ) and mastermind-like protein 1,2,3 (MAML1,2,3). In the non-canonical pathway (RBPJ-independent NOTCH signaling), NOTCH is activated through interaction with IKKA (IkappaB kinase α) of the NF-κβ (nuclear factor kappa-light-chain-enhancer of activated B cells) pathway, with lymphoid enhancer-binding factor 1 (LEF1) of the WNT signaling, or with R-Ras, which is responsible for cell adhesion [113,114,115].
BMP is part of the TGF-β superfamily, participating in the regulation of cardiovascular and lymphatic development [116]. TGF-β regulates the interaction between ECs and the microenvironment (vascular smooth muscle cells, pericytes). TGF-β takes part in embryonic vascular development, vascular integrity, endothelial activation, and proliferation [117]. BMP activates SMAD-dependent (canonical) and multiple SMAD-independent (non-canonical) pathways. Interaction of BMP with cell surface receptors leads to the formation of heterotetrameric complexes. Type I and type II serine/threonine kinase receptors change the gene transcription and signal transduction cascade [118]. The type I receptors include activin receptor-like kinase-1,2,3,5,6. The type II receptors are transmembrane serine/threonine kinase receptors, which are constitutively active and include BMP2, type 2 activin receptor (ACTR-2A), and type 2B activin receptor (ACTR-2B) [119]. The canonical BMP signaling pathway implicates phosphorylation of SMAD1 and SMAD5 intracellular mediators responsible for initiating angiogenesis in venous and arterial ECs. Notably, the conditional knockout of SMAD1/5 in the ECs results in the downregulation of NOTCH signaling. Disruption of SMAD1/5 is the cause of abnormal vascular development and defective lineage specification of ECs [120,121]. Non-canonical BMP signaling occurs by activating TAK-1 (transforming growth factor-β activated kinase 1), PI3K/Akt (phosphatidylinositol-3 kinase/protein kinase B), and Rho-GTPases [122].
The FGF signaling interacts with transmembrane tyrosine kinase receptors, activating their pathways (PI3K/Akt, RAS/MAPK (RAS superfamily/mitogen-activated protein kinase), and phospholipase Cγ/PLCγ). The FGF protein family contains 22 signaling ligands. However, only FGF1 (acidic FGF) and FGF2 (basic FGF) are crucial for angiogenesis [123]. FGF signaling is necessary for developing ECs, injury response, repair, and the initiation of pathologies (cancer, abnormal angiogenesis) [124]. Interestingly, FGF-2 can modulate the endothelial expression of VEGF through both autocrine and paracrine mechanisms of action [125].
The ETS transcription factors are expressed in embryonic endothelial cells and stimulate initial capillary network formation through proliferation, cell spreading, and adhesion. A lack of ETS factors provokes severe defects in embryonic angiogenesis [94,126,127].
GATA is a zinc-finger family of transcription factors essential for developing the endocardium and vessels [128]. GATA-4,5,6 are mostly expressed in the cardiac endothelium, while GATA2 plays a critical role in blood cell development [129].
VEGF is a potent angiogenetic factor for the proliferation and migration of ECs during angiogenesis and vasculogenesis [130]. VEGF binds to VEGFR: VEGFR1 (or FLT1) and VEFR2 (or FLK1). FLK1 mediates almost all known angiogenesis-dependent functions (migration, survival, vascular permeability) [131,132,133]; the additional function of Flt1 relates to organ differences in physiological and pathological conditions such as angiogenesis [134], homeostasis, preeclampsia, and metastasis [135,136]. In this sense, VEGF-A/Flk1 are the earliest markers of mesodermal differentiation and potent angiogenesis promoters, responsible for physiological and pathophysiological permeability, vascular organization, cell cycling, and differentiation [137,138,139].
The SCL/TAL1 factor is a transcription factor with a helix–loop–helix structure, regulating hematopoiesis and endothelial cell reprogramming [140]. SCL/TAL1 stimulates transcription by involving a core complex on DNA containing E2a/HEB, GATA1-3, LIM domain only 2-rhombotin-like 1 (LMO1/2), LIM-only, LIM domain-binding 1 (LDB1), ETO2, runt-related transcription factor 1 (RUNX1), and FLI1 [141]. SCL/Tal-1 is essential for initiating mesodermal endothelial differentiation, vascular network formation, and postnatal angiogenesis [142].
The LMO2 transcription factor is involved in vascular remodeling through the LIM-domain zinc-finger-like structures and DNA interaction [143]. The deficiency of the LMO2 results in a decrease in endothelial migration [144].
Retinoid acid contributes to endothelial differentiation. The lack of retinoid acid leads to embryonic death, with multiple defects in vessel reorganization of the yolk sac [145]. The retinoid acid receptor is essential for angiogenesis regulation [146], EC proliferation [147], and blood barrier regulation [148].
The control of endothelial development is complex and context-dependent (Figure 1A) [149,150,151,152,153,154,155,156,157,158,159,160,161].

Figure 1.
(A). Interaction and cross-talk between different pathways in endothelial development and function. Molecular signaling cross-talk: WNT/VEGF-A: in ECs, WNT-pathway is constitutively suppressed in ECS. WNT-ligand activates FRZ receptors through the inactivation of the destruction complex (APC-c-Cbl β-TrCP- β-catenin) and translocation of β-catenin to the nucleus with further activation of VEGF-A; VEGF/NOTCH demonstrates a synergistic effect. VGF, through the VEGFR2, increases expression of DLL4, leading to NOTCH activation. NOTCH receptors demonstrate synergism (VEGF 1, VECGF3) and antagonism (VEGF2) in VEGF expression; NOTCH/BMP activates expression of the inhibitory I-SMAD protein, leading to inhibition of BMP 2/6; NOTCH/TGF-β cross-talk is characterized by downstream of ALK receptors and depends on R-SMAD and Co-SMAD activation with the formation of SMAD/NICD complex of transcription (similar to β-catenin/NCID complex). As a result, the FGF-β-induced expression of NOTCH target genes (HEY/HES) leads to migrations of ECs; BMP/WNT/TGF-β demonstrate synergetic regulation and determine ligand production of each other; WNT/TGF-β cross-talk leads to synergetic regulation of the set of shared target genes in the nucleus via the Smad/Lef/β-catenin. NOTCH/WNT cross-talk is complex and involves several regulation levels: formation of β-catenin/NICD transcriptional complex (orange lines), the interplay between NOTCH receptors and β-catenin at the membrane (pink lines), NICD phosphorylation by GSK3b (blue lines), and inhibitory interaction between CSL and Disheveled (DVL) and inhibition of effector gene expression (green). (B) Schematic representation of the pathway signaling cross-talk in ECs: APC—adenomatous polyposis coli; BMP—bone morphogenetic proteins pathway; BMP ligands: BMP 2,4,5,6,7; BMPR—bone morphogenetic proteins receptor; b-TrCP (Fbxw1 or hsSlimb)—β-transducin repeat-containing protein; b-TrCP-p—phosphorylated b-TrCP; c-Cbl—casitas B lineage lymphoma protein with Ee ligase activity; c-Cbl-p—phosphorylated c-Cbl; Co-SMADS: SMAD 4; CSL-(CBF1, suppressor of hairless, Lag1)—transcription factor activating the genes downstream of the NOTCH pathway; HES 1,2—hairy and enhancer of split 1,2; HEY 1—hairy/enhancer of split related with YRPW motif protein 1; I-SMADS: SMADS 6, 7; NICD—NOTCH intracellular domain; R-SMADS: SMADS 1,2,3,5,8; TGF-β—transforming growth factor β; TGF-b ligands: TGF-b 1. 2,3; VEGF-A—vascular endothelial growth factor A; VEGFR—vascular endothelial growth factor receptors; WNT—wingless-related integration site.
The antagonistic, agonistic (reciprocity), and synergistic interplays between the pathways result from combinations of intrinsic and extrinsic cues: type and subtype of pathways (e.g., NOTCH differs from 1 to 4), tissue specificity, timing of pathway activation (simultaneous or sequential), duration of signaling, impact of other pathways, etc. (Figure 1B) [153]. BMP and NOTCH demonstrate antagonistic effects [162,163]. BMP/WNT/TGF-β demonstrate synergetic regulation and determine the ligand production of each other; NOTCH acts downstream of VEGFR; and WNT-NOTCH cross-talk has been shown to regulate multiple effects on cell fate. WNT plays the role of agonist and antagonist for NOTCH in cardiogenesis, and WNT and TGFβ, in most cases, demonstrate an agonistic relationship in extracellular signaling [153,164,165,166].
3. Endothelial Ontogenesis
3.1. ECs and Stem Cells Interchange
During postnatal ontogenesis, a dynamic reciprocal exchange between the stem cells and ECs is vital for vascular regeneration and the maintenance of physiological and morphological integrity. Continuous endothelial regeneration relies on the ability of stem cells to be converted into ECs. There are different types of stem cells that demonstrate strong endothelial potential: human embryonic stem cells [167], adult stem cells [168], and hematopoietic stem cells [169]. The postpartum physiological (wound healing) and pathological (tumor angiogenesis) endothelial renewal occurs through the migration of pre-existing ECs from the vascular wall or via recruitment of endothelial precursors, so-called endothelial progenitor cells, from the circulatory system [170,171]. Endothelial progenitor cells are the provasculogenic subpopulation of hematopoietic stem cells, demonstrating a profound capacity for proliferation, specialization, differentiation, and maturation [172,173,174]. Endothelial progenitor cells are subdivided into erythro-myeloid progenitors, non-hematopoietic endothelial progenitor cells [175], endothelial colony-forming cells, or hematopoietic endothelial progenitor cells [176]. The primary source of non-hematopoietic endothelial progenitor cells is blood [177,178]. Bone marrow-derived hematopoietic endothelial progenitor cells can be considered as a provasculogenic population of hematopoietic stem cells. The hematopoietic stem cells and endothelial progenitor cells express specific surface markers such as CD34, CD133, CD105, FLK-1/KDR, and CXCR4 [179]. In humans and animals, the bone marrow niche is formed by specialized tissue with a specific microenvironment where cellular and molecular components regulate the cell fate and behavior of endothelial progenitor cells [180]. Bone marrow includes vascular (hematopoietic stem cells, ECs, mesenchymal stem cells, and endosteal cells (osteoprogenitor cells and osteoclasts under the control of parathormone)) niches [181]. The functional regulation of the bone marrow niches, determining the cell fate, mostly occurs via epigenetic cues: epigenetic mechanisms (histone modifications, DNA methylation, non-coding RNAs) [182], sympathetic and parasympathetic innervation [183], and metabolic and chromatin modifications [184].
3.2. Role of ECs in Hematopoiesis and Vasculogenesis
Hematopoiesis is the formation of blood cells, which includes primitive (in the yolk sac) and definitive (aorta–gonad–mesonephros region, liver, bone marrow) stages [185]. Vasculogenesis is the de novo formation of blood vessels [186]. During the early stages of embryogenesis, vasculogenesis and hematopoiesis occur in parallel and are closely linked to each other [187]. In many mammalian species, hematopoietic stem cells, hematopoietic precursors, develop directly from hemogenic endothelium [188]. Hemogenic ECs are genetically programmed to transdifferentiate into hematopoietic stem cells through endothelial-to-hematopoietic transition and EndMT [189,190]. The cooperation of EC and hematopoietic progenitor stem cells is indispensable for the formation of blood islands (hematopoietic precursors, hematopoietic progenitors) in the yolk sac, fusion, and development of the primitive capillary plexus [191]. Endothelial specification occurs under the control of the well-defined and aforementioned signaling pathways: BMP4, WNT [61], NOTCH, and Hedgehog (Hh) [192]. The Hh signaling pathway is involved in the control of embryonic and adult hematopoiesis and is required for endothelial patterning; Hh mutants demonstrate a functional lack of vascular remodeling [193]. A number of cytokines and interleukins (granulocyte-colony stimulating factor (GM-CSF), IL-3, IL-4, IL-6, IL-8) regulate hematopoiesis and contribute to endothelial growth and differentiation as well [194]. VEGF, angiopoietin-1 [195], FGF-2, TGF-α, TGF-β, HGF, tumor necrosis factor-α (TNF-α), angiogenin [194], and recombinant human erythropoietins (EPO-α and EPO-Z) [196] stimulate vasculogenesis. Moreover, the endothelial and hematopoietic fate is regulated by TAL-1 (also known as SCL), GATA-1 and 2, LMO2, RUNX-1 [188], and stromal cell-derived factor-1 alpha (SDF-1 alpha) [197]. The endothelial cell-selective adhesion molecule (ESAM) is a member of the immunoglobulin superfamily. It facilitates communication between ECs and determines interactions between ECs and hematopoietic stem cells in hematopoiesis and vasculogenesis [198,199,200].
4. Endothelial Plasticity as the Base of Endothelial Diversity
All endothelial cells are multivalent players and messengers contributing to local and organism-wide functions. ECs exhibit structural and functional plasticity, leading to diversity in the endothelial population.
Heterogeneity is determined by a combination of (1) different genes [201], e.g., homeobox genes (HOX family of transcriptional factors B3, D4) [202], E-selectin, thrombomodulin, von Willebrand Factor (vWF), platelet endothelial cell adhesion (PECAM-1), and Ephrin B4 genes [203]; and (2) epigenetic cues, e.g., physical influences (low blood flow versus high blood flow), intrinsic biochemical characteristics (regulatory proteins, hormones, transcription factors), non-coding RNAs, and functional and morphological biochemical mutual interactions [204].
The genes and receptors that are sensitive to environmental changes include neutrophin-4, transforming growth factor-β, VEGFR-3, interleukin-1 receptor type 1 precursor, E-selectin, integrin α4 precursor, C cathepsin, cytochrome P450 IVB1, Bad Protein, a myb proto-oncogene, Ku 70-kDa subunit, S100 calcium-binding protein A7, MAL, and a regulator of G-protein signaling 16/a28 [205].
Epigenetic adaptivity and reversibility are due to the modulation of the transcription profile and dedifferentiation of the initial endothelial phenotype [206]. The loss of inductive signals leads to the modification of tissue-specific receptors that affect gene expression, where non-coding RNA regulating methylation and histone modifications play an essential role [39,207]. Physical epigenetic extrinsic factors (inductive signals) such as blood flow, shear forces, hypoxia, and foreign RNA fragments are more powerful regulators. For example, blood circulation with low shear stress alone stabilizes hypoxia-inducible factor 1α (HIF-1α), influences cell reprogramming, and ultimately decreases hypoxia levels [208]. Hypoxic conditions increase VEGF production and also stimulate human fibroblast conversion to ECs [209]. Certain viral Kaposi sarcoma herpesvirus miRNAs (miR-K12-11, miR-K-12-6) delivered via extracellular vesicles, together with transcription factors, can be considered as signaling molecules that can modulate or correspond to the epigenetic environment, or act as effectors of altered environment, participate in cell reprogramming, and stimulate angiogenesis [210,211].
4.1. Target Regulation of Endothelial Plasticity and Heterogeneity
Not much is known about the factors that may determine endothelial plasticity.
The pathways thoroughly described previously play a crucial role in this process: NOTCH (generally described previously), WNT (mentioned above), BMP (described previously), TGFβ, (VEGF) [212], Lim domain only 2 (LMO2) [213], ETS variant transcription factor 2 (ETV2) [214], twist family BHLH transcription factor 1 (TWIST-1) [215], and hypoxia-inducible factor 1 (HIF1) [216]. This section specifically focuses on the characteristics associated with endothelial plasticity and heterogeneity.
4.1.1. NOTCH
NOTCH is one of the principal regulators of endothelial heterogeneity, determination, and endothelial apoptosis. NOTCH, as mentioned before, inhibits the proliferation of the sprouting ECs through direct interaction between the receptors and DLL4 [217]. Endothelial plasticity and heterogeneity are closely related to NOTCH 1 and NOTCH 4 receptors. They differently influence endothelial apoptosis and contribute to the complex regulation of vascular remodeling. Thus, NOTCH1 and NOTCH4 determine apoptosis and normal morphology and are related to cellular context and external stimuli. Activation of NOTCH1 signaling leads to an augmentation of cell survival under stress conditions via upregulation of anti-apoptotic proteins Bcl-2 and Bcl-xl [218]. Additionally, NOTCH 1 signaling in cell survival may enhance the expression of survival pathways such as PI3K/Akt [219]. Notch4 signaling is an antagonist of NOTCH1 under certain conditions. Overexpression of NOTCH4 leads to enhanced endothelial apoptosis through the induction of pro-apoptotic factors like p53 and Bax [220]. NOTCH4 leads to the dynamic restriction of vascular networks during development, contributing to the pruning of aberrant vessels [221]. NOTCH signaling participates in the transformation of the venous ECs into artery ECS via the NR2F2 (Nuclear Receptor Subfamily 2 Group F, Member 2) receptor [222].
4.1.2. WNT
WNT is one of the crucial regulators of endothelial plasticity and heterogeneity. As mentioned before, WNT signaling acts 1) all over the body through the canonical WNT/catenin β signaling [223,224,225] and non-canonical pathway WNT/Ca2+ [226,227] and 2) locally in context-related responses [228,229]. WNT determines endothelial morphology in a case of disturbed blood flow, controls endothelial integrity, regulates the distribution of tight junctions [223], and controls endothelial maturation [230]. All the aforementioned mechanisms may be considered as the basis of endothelial heterogeneity.
4.1.3. BMP
BMP signaling selectively influences ECs in a context-dependent manner without affecting endothelial survival through SMADS effectors and a non-canonical pathway via sp38, MAPK, and ERK [231]. The transcriptional activation of BMP signaling occurs under distinct epigenetic influences. For example, the turbulent shear stress induces phosphorylation of SMAD1/5/8, which provokes the activation and proliferation of ECs. Laminar shear stress leads to endothelial hibernation through the BMP9/Alki1/ENG-SMAD cascade [232,233]. Interestingly, SMADs may also interrelate with p300/CREB binding protein (CBP) signal transduction and readily create a complex with the NOTCH intracellular domain, activating NOTCH target genes. Post-transcriptionally, BMP targets are related to the reversible phosphorylation process [233,234], ubiquitination, and sumoylation [233,235], affecting cell survival [236,237].
4.1.4. TGFβ
The transforming growth factor-β (TGFβ) signaling pathway plays a vital role in the regulation of EC heterogeneity of the vascular systems. Different aspects of endothelial biology are influenced by TGFβ. This pathway signals through type I and II serine/threonine kinase receptors ALK1 (leading to SAMAD1/5/activation and promotion of the EC proliferation and migration) and ALK5 (phosphorylation of SMAD2/3 leading to inhibition of EC proliferation and migration) [238,239]. The role of TGFβ is that of a potential inducer of EndMT through the SMAD-dependent and SMAD-independent pathways [238,240].
4.1.5. VEGF
VEGF can be considered a driver of endothelial plasticity due to the heterogeneity of ECs. Deletion or mutation of the VEGFR1 has different effects in different organs: in the retina, it increases vascular density, and in the liver, there is no change [231]. Deletion of the VEGFR2 causes regression of the vascularization [231], and VEGFR3, similarly to VEGFR1, initiates hypersprouting of vessels in the retina [241]. It is interesting that different VEGF isoforms determine the phenotypic specialization of endothelial cells in different organs [242]. This explains the phenomenon of ECs of various organs responding differently to VEGF changes in the vascular bed [243].
4.1.6. LMO2
LMO2 is a crucial element in the axis of LMO2-Prdm16 (PR domain containing 16) involved in the development of hematopoietic stem cells through the endothelial-to-hematopoietic transition as a form of endothelial plasticity [244]. Moreover, in the zebrafish, LMO 2 is a main fate plasticity messenger that determines the decision between endothelial and pronephros tubule cells [245].
4.1.7. ETS (ETV2)
ETS2 plays a pivotal role in endothelial plasticity in the context of specification and early differentiation from mesodermal progenitors [246]. This transcription factor has several lines of action, participates in the migration of progenitor ECs [247], and regulates endothelial reprogramming [229,248].
ETV2 is able to directly reprogram cells (shown on fibroblasts) into endothelial fate lines [249] and to upregulate cell migration through increasing gene chromatin accessibility, regulating cell migration [247].
4.1.8. TWIST-1
The role of this transcriptor factor is strongly associated with epithelial-to-mesenchymal transition (EMT) and is well studied, mainly in tumorigenesis [250,251]. Endothelial cells actively participate in tumor neovascularization and exhibit high levels of plasticity and heterogeneity during this process. TWIST1 stimulates EMT by representing E-cadherin expression and upregulating mesenchymal markers such as vimentin [250,251].
4.1.9. HIF1
As a universal protective molecule against low oxygen concentrations, HIF1 is involved in the induction of VEGF [252] and the modification of gene expression that promotes angiogenesis and vascular remodeling [253]. Additionally, HIF1 is potentially linked with EndMT and may influence cancer and other pathological conditions [253].
To summarize, the concept of endothelial plasticity is not only an ability of ECs to adapt their own initial genetically determined programs to the local epigenetic landscape, but an ability to induce plasticity of surrounding cells belonging to different cell types. ECs are able to build an endothelial-friendly local environment [254,255,256,257], e.g., co-culturing ECs with tanycytes induces cytoskeleton remodeling in the latter through the NO-mediated signaling pathway with cGMP and involvement of prostaglandins and thromboxanes [258]. Herein, ECs indirectly control the glial–neuronal interactions in the brain. To summarize, the concept of endothelial plasticity is not only the ability of ECs to adapt their own initial genetically determined programs to the local epigenetic landscape, but the ability to induce plasticity of surrounding cells belonging to different cell types. ECs are able to build an endothelial-friendly local environment [242,243,244,245], e.g., co-culturing ECs with tanycytes induces cytoskeleton remodeling in the latter through the NO-mediated signaling pathway with cGMP and involvement of prostaglandins and thromboxanes [246]. Herein, ECs indirectly control the glial–neuronal interactions in the brain.
5. Endothelial Diversity along the Vascular Bed
5.1. Diversity of Ecs in the Microcirculation
ECs at the microcirculation level (arterioles, capillaries, post-capillary venules with a lumen diameter of less than 100 μm) demonstrate more substantial heterogeneity than the macrovessels [259,260,261]. Arterial, venous, and lymphatic micro/macrovessel ECs across the same tissue have similar morphology and demonstrate analogous transcriptomes. Despite standard endothelial features, the transcriptomes in different tissues are also entirely different [262]. Local epigenetic factors that influence microvascular diversity are shear stress, intravascular pressure, and metabolic clues [263]. Genetic influence is essential for the biochemical organization of the endothelial basal lamina, a thin sheet of extracellular matrix (ECM) located at the epithelial cells’ basal surface. It can be visualized by light microscopy and consists of a lamina lucida or lamina rara (composed of laminin, integrins, entactins, dystroglycan), a lamina densa (collagen IV type, perlecan), and a lamina fibroreticularis. The lamina lucida and lamina densa are named in the literature as the basal laminae and can be visualized only through electron microscopy [264,265,266]. The specific microvascular gene expression patterns are primarily implicated in producing particular basal lamina proteins, including laminin, collagen (4a1 and 4a2), and ECM-interacting proteins (cD36, α1 integrin, α1 integrin, β4 integrin). The microcirculation-specific genes regulate the individual biochemical characteristics of the endothelial cell membrane. Microcirculatory ECs have a surface covered with a negatively charged glycocalyx (composed of glycoproteins and glycolipids). Glycocalyx thickness varies between different parts of the microcirculatory tree [267,268]. The ECs of the fenestrated glomerulus have a thick and specialized glycocalyx (e.g., playing a role in the filtration barrier), whereas the ECs of the fenestrated sinusoidal capillaries of the liver have a thin glycocalyx layer, allowing for quick exchange between the blood and space of Disse [269].
At the microcirculatory level, endothelial morphology is strongly correlated with its functions. ECs are divided into several phenotypes: (1) continuous (have continuous basal lamina and have no fenestrations) (Figure 2) and (2) fenestrated (have fenestrations), with several subtypes (Figure 3 and Figure 4): (a) pseudo-fenestrated (fenestrations covered by fenestral diaphragm), (b) glomerular fenestrated (without fenestral diaphragm); and (c) discontinuous fenestrated cells (lack of basal lamina) (Table 3, Figure 3) [26,270,271,272,273,274,275,276,277,278,279,280,281,282,283,284,285,286,287,288,289,290,291,292,293,294,295,296,297,298,299,300,301,302,303].
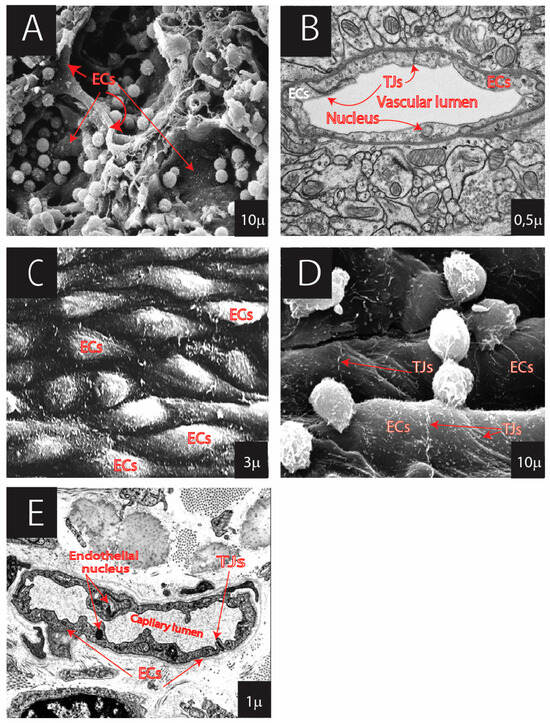
Figure 2.
Continuous ECs showing diversity in various organs: (A) medullary sinus of lymph node (SEM: surface scanning electron microscopy); (B) brain with tight junctions (TJ) of the blood–brain barrier (TEM: transmission electron microscopy); (C) ECs of arteries (SEM); (D) ECs of veins (SEM); (E) EC of capillary of dermis/skin (TEM). Kindly provided by emeritus professor P. Groscurth through https://e-learn.anatomy.uzh.ch/Anatomie/Anatomie.html.

Figure 3.
Fenestrated ECs showing diversity in the organs: (A) ECs of kidney glomerulus (SEM); (B) ECs of kidney glomerulus (TEM); (C) adrenal gland capillary with fenestrated ECs and fenestral diaphragms (FD) (TEM); (D) ECs of the sinusoid of liver (SEM). (Kindly provided by emeritus professor P. Groscurth through https://elearn.anatomy.uzh.ch/Anatomie/Anatomie.html).
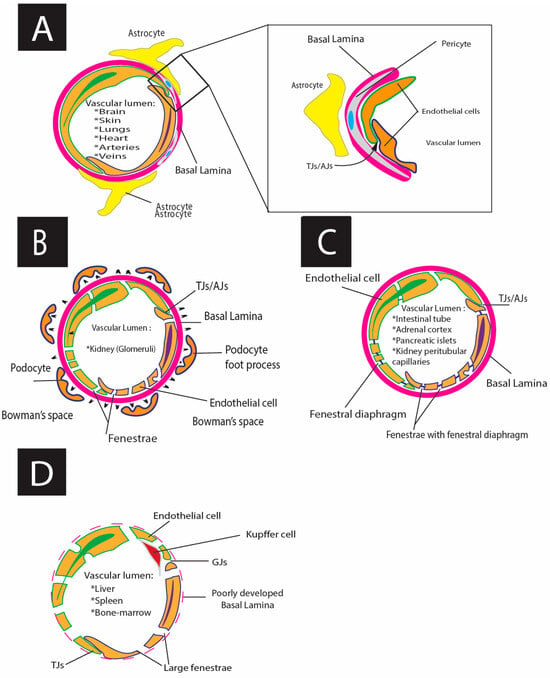
Figure 4.
Schematic representations of different types of endothelium: (A) Schematic drawing of continuous endothelium; (B) schematic drawing of fenestrated endothelium (glomerular fenestrated endothelium (true fenestrated), with fenestrae ~60–100 nm in diameter), TJ (tight junctions), and AJ (adherens junctions); (C) schematic drawing of pseudo fenestrated endothelium with fenestral diaphragm (FD (fenestrae ~60–70 nm in diameter); and (D) schematic drawing of discontinuous sinusoid endothelium with GJs (gap junctions), TJs, and large pores ~100–200nm.
Table 1.
Regulating factors of vascular development.
Table 1.
Regulating factors of vascular development.
| Vasculogenesis Differentiation of Endothelial Precursor Cells (EPCs) into ECs and De Novo Formation of the Primitive Vascular Network (ECs from Mesoderm) | ||
|---|---|---|
| Phases: | Signaling and transcriptional regulators: | Markers of endothelial differentiation: |
| I. Extraembryonic vasculogenesis: starts in ~ mice E6.5 embryos, ~3 weeks in humans (yolk sac, allantois, placenta) [59,61,65] | 1. Fibroblast growth factor (FGF) family includes 18 paracrine and endocrine factors [56,62,69,73,74] 2. The hedgehog family: Shh, Ihh, Dhh [70] 3. Vascular endothelial growth factor (VEGF) and vascular endothelial growth factor receptors (VEGFRs): VEGFR-2 and VEGFR-1 [56,57,64,67,68,69,74,75,76,84] 4. Neuropilin1(NRP-1) and Neuropilin2 (NRP2) [63,65,67,68,72,76] 5. Transforming growth factor β(TGFβ) and transforming growth factor receptors (TGFRs) [56,57,63,69,70,74,75] 6. Angiopoietins 1 and 2 (ANG1 and ANG2) binds tyrosine kinase with immunoglobulin-like and EGF-like domains 1 and 2 (Tie1 and Tie2) and affect the remodeling of capillary plexuses [70] 7. Platelet-derived growth factor (PDGF) recruitment of pericytes and smooth muscle cells [57,63,65,70,72,74,76] 8. GATA proteins [61,65,67] 9 Krüppel-like factors [61,63] 10. ETS proteins regulate ECs differentiation [61] 11. Homeodomain proteins (HOXB3 and HOXD3) participate in morphogenesis of vascular tube formation [62,70,90] 12. Epidermal growth factor like domain 7(EGFL7) participation in separation and arrangement of angioblast) [59,61] 13. Fibronectin and its receptor α5β1 [65] 14. SOXF factors: SOX7, SOX17, and SOX18 [66] 15. Granulocyte colony-stimulating factor (G-CSF) attenuated delayed tPA [63,71] 16. Overexpressed hypoxia- inducible factor (HIF-1α) stimulates endothelial progenitor cells [64,76] 17. Increase in intracellular Ca2+ concentration [68] 18. Bone morphogenetic protein (BMP) signaling pathway [63] 19. retinoid acid [67] 20. Wnt β-catenin signaling [56,61,67] 21. T- box transcription factor gene 18 (TBX18) [56] 22. Wilms tumor transcription factor 1) WT1 [56] | Vascular endothelium cadherin VE-cadherin [59,61,63,65,67,71,76] von Willebrand Factor (vWF) [62] CD34 (early angioblasts and endothelial progenitors) [59,62,63,71,72,73] T-cell acute lymphocytic leukemia (TAL1) [61,65,84] platelet endothelial cell adhesion (PECAM-1)/CD31 [56,59,61,63,73] Tyrosine kinase with immunoglobulin-like and EGF-like domains (Tie-1) and tyrosine kinase with immunoglobulin-like and EGF-like domains (Tie-2) [61,63,65] Flk1 and Flt1 [61,65] Thrombospondin type1 domain-containing protein 1 (THSD1) [62] |
| A—Assemblation of blood island within the mesodermal layer (yolk sac) [61] | ||
| B—Hemangioblast formation from blood islands (hemangioblast inner part gives rise to the hematopoietic precursors, the outer part give rise to angioblast, differentiation in situ) [61] | ||
| C—Primitive extraembryonic vascular (may contain primitive erythrocytes) plexus organization and ECs differentiation happens in association with hematopoietic precursors in blood islands [61,65] | ||
| II. Intraembryonic vasculogenesis: It starts in ~ mice E7.3 embryos and gives rise to the endocardium great vessels and is not associated with blood formation. There are two types of intra-embryonic hemangioblast forming vascular plexus (usually without erythrocytes) [61,65,73] | ||
| A—Hemangioblast from splanchnopleuric mesoderm (visceral) associated with the endoderm (production of hematopoietic cells and paraaortic splanchnopleura) [63,69] | ||
| AA—Hemangioblast from somatopleuric mesoderm (parietal) associated with ectoderm (give rise to all types of cells, except hematopoietic stem cells) [63,69,73] | ||
| B—Primitive intraembryonic vascular plexuses formation: ECs differentiate from mesoderm as solitary angioblast without the concomitant differentiation of hematopoietic stem cells, except for small regions in the aorta (paraaortic clusters) [61] | ||
| III. Functional vascular network formation in vasculogenesis: primary capillary extraembryonic plexuses anastomose with intraembryonic vasculature through the vitelline arteries and veins and then connect with developing heart tube [61] | ||
| Angiogenesis Growth of primary and secondary vascular plexus from pre-existing blood vessels (vessels which lack a fully developed tunica media) in prenatal or postnatal life | ||
| Mechanism and phases: | Signaling and transcriptional regulators: | Markers of endothelial differentiation: |
| I. Sprouting mechanism—based on endothelial cell migration, proliferation, and tube formation with or without blood flow (the result is new vascular tube formation) [56,64,67,68,69,73,84] | 1. Vascular endothelial growth factor VEGF [56,57,61,62,63,65,67,69,70,71,72,74,75] 2. Transforming growth factor β,α (TGFβ,α) [64,66,70,71] 3. Angiopoietins and tyrosine kinase with immunoglobulin-like and EGF-like domain receptors (Tie receptors) [65,70,74] 4. Platelet-derived growth factor (PDGF) and Platelet-derived growth factor receptor-β (PDGFR-β) [65,73] 5. SOXF factors: SOX7 and SOX18 [66] 6. Atypical chemokine receptor CXCR7(ACKR3) [76] 7. Angiopoietins 1 and 2 (ANG1 and ANG 2) [69,70,74] 8. Granulocyte colony-stimulating factor (G-CSF) attenuated delayed tPA [63,71] 9. hypoxia-inducible factor HIF-1α [65,71] 10. Increase in intracellular Ca2+ concentration [68] 11. High mobility group box 1 (HMGB-1) [60] 12. Connective tissue growth factor (CCN2) [60] 13. Delta/jagged-NOTCH signaling [61,63,65] 14. Metalloproteinases MMP [63,69] 15. Ephrin- B (EPH-B) [63,69] 16. Semaphorins (SEMA 3 proteins) [63,65,70] 17. Rho-associated protein kinase (ROCK) [63] 18. Chicken ovalbumin upstream promoter transcription factor (Coup-TFII) [84] | Vascular endothelium cadherin (VE-cadherin) [59,61,63,65,67,71,76] von Willebrand Factor (vWF) [62] CD34 [59,62,63,71,72,73] Platelet endothelial cell adhesion (PECAM-1)/CD31 [56,59,61,63,73] Endoglin/CD105/ [65,72] THSD1 Thrombospondin type1 domain-containing protein (THSD1) 1 [62] Krüppel-like factor 4(KLF4) [63] A disintegrin-like and metalloprotease with thrombospondin type 1 repeats 13 (ADAMTS-18) [62] Aminopeptidase N APN(CD13) in tumorigenesis [72] Intercellular adhesion molecule-1(ICAM-1 or CD54) [72] |
| A. Neovessel growth—the disintegration of the basal lamina of existing vessel, migration and proliferation of ECs, lumen formation, and loops organization by sprouts and anastomoses [226,239] | ||
| B. Neovessel stabilization—delay of the endothelial proliferation, basal lamina reconstruction, coverage of the immature vessel with pericytes [63,67] | ||
| II. Intussusceptive microvascular growth (IMG) mechanism: based on the division of existing vessel lumens by formation and insertion of tissue folds and interstitial cellular columns into the lumen of pre-existing vessels (lumen expansion occurs through the organization of new units of extracellular matrix). Blood flow-dependent process [63,69] | ||
| A. Interendothelial “transluminal bridge” formation: ECs located at the opposite side of the capillary wall move near to each [63] | ||
| B. “Cylinder tissue bridge” establishment: tissue form as a cylinder bridge, which perforates endothelial bilayer and extends across the lumen. Then ECs cover the cylinder tissue bridge involving the cytoplasmic extensions of myofibroblast and their microfilaments inside the cylinder’s core [63] | ||
| C.” Pillar” formation: the framing of the pillar by pericytes close to the lateral part wall [63] | ||
| D.” Pillar” growth: pillar growth into an intercapillary mesh [63] | ||
Continuous ECs are present in tissues with low but well-controlled exchange. This endothelial cell type’s primary characterization is the continuous basal lamina and lack of fenestra (Figure 2, Table 3) [304]. In contrast, the fenestrated and sinusoidal endothelium is mainly located in the organs with high uni- or bidirectional substrate diffusion.
Pseudo-fenestrated ECs are located in endocrine tissues, gastrointestinal mucosa, and renal peritubular capillaries. They have a continuous basal lamina and pores (60–70 nm in diameter), with a thin diaphragm (Figure 3, Table 3).
Discontinuous fenestrated ECs (or sinusoidal ECs) are characterized by a lack of a continuous basal lamina, multiple fenestrations with 50–100 nm in diameter aggregated into a group of 10–100 fenestrae (liver “sieve plates”), and big pores (100–200 nm in diameter) without a diaphragm. These types of ECs are typical for the liver, spleen, and BM (Figure 4, Table 3).
Glomerular fenestrated ECs take the intermediate position between common pseudo-fenestrated and discontinuous fenestrated ECs: they have a basal lamina, pores with diameters of 60–80, and no diaphragms (Table 3) [293,297,305]. The local microcirculatory functions and morphology contribute to permeability regulation, i.e., an exchange between the vascular lumen and the extravascular tissue [263,306,307]. The above-mentioned phenotypic differences of ECs directly affect microcirculatory endothelial permeability [308].
High endothelial venules (HEVs) are a special part of the microcirculatory bed [309]. These vessels belong to post-capillary venules, which mostly accompany lymphoid tissues such as lymph nodes, Peyer’s patches, and tonsils. The specificity of these cells is strongly related to immune system function and serves to facilitate the entry of lymphocytes from the bloodstream into the lymphoid tissues (place of immune response initiation and regulation). The ECs of HEVs have a flat and cuboidal shape, unlike the flat and elongated morphology of typical ECs. Notably, the surfaces of these types of ECs display a specific glycosylation pattern composed of Pereferial node addressins (PNAd) and express high levels of sialomucins, which contribute to lymphocyte homing through binding L-selectin to lymphocytes [310]. The listed structural and functional characteristics facilitate the mediation of lymphocyte trafficking into lymphoid tissues, supporting effective immune surveillance and response [311].
Critical structural elements of the endothelia, regulating the cellular diffusion process (especially across the continuous endothelium) and layer stability, are cell-to-cell junctions [312]. Junctional endothelial diversity is another marker of EC heterogeneity, characterized by variations in the expression of junctional proteins [313]. The junctions connect to cytoskeletal proteins and several signaling proteins that contribute to the maintenance of shape and polarity [314,315]. There are three types of cellular junctions: tight junctions (TJs), adherence junctions (AJs), and gap junctions (GJs) [272].
TJs are associated with barrier tissues, where the fast interchange between the blood and tissues does not exist or is strictly regulated. These types of junctions are typical of the endothelium in the brain, lungs, endocardium, nerve capillaries, fat, and muscle tissue. They display the role of a selective wall, allowing for the select paracellular passage of ions between cells [316,317]. The claudin family (26 proteins in humans), occludin, and tricellulin are responsible for the TJs’ barrier specificity [318,319]. In these tissues, TJs are not only responsible for the barrier function (blood–brain barrier, air–blood barrier, and blood–nerve barrier), but also for the physical stabilization of macrostructures with regular physical stress, e.g., muscle and endocardial ECs adapt quickly to shape changing during muscle contraction [320,321,322,323,324].
AJs are cell–cell adhesion complexes interacting with the F-actin cytoskeleton and contribute to tissue homeostasis, embryogenesis, stabilization, initiation of cell–cell adhesion, and control of intracellular signaling. These multiprotein complexes are composed of cadherins and nectins [325]. TJs and AJs are zipper-like elements localized at the lateral membrane of the ECs [326]. The function and stability of EC AJs are regulated by various types of interactions with actin cytoskeleton participating in modulation of endothelial permeability as a reaction to different types of stimuli [327].
GJs are members of the connexin protein family which form intercellular channels and provide direct communication patterns between endothelial and surrounding cells: electrical coupling and flow of metabolites in exchange [328]. GJs proteins are predominantly tightly associated with the plasma membrane and, in addition to being passively transported, may also play a role in the rough endoplasmic reticulum, mitochondria, and Golgi apparatus [329]. GJs participate in transmitting vasodilatory signals from the capillary network to arterioles, and they conduct signals from the endothelial to the muscle cell layer in some vascular beds [330,331]. They contribute to the chronic remodeling of vessels through the induction of cellular stiffness, actin rearrangement, and activation of pro-inflammatory genes that result in disease development [332,333]. GJs are mainly located in organs with a rapid molecular exchange between the blood and the surrounding compartment (endocrine glands, lymphatic capillaries, liver sinusoids, spleen, and BM). GJs facilitate fast information exchange that helps the whole endothelium to respond to a focal signal in a coupled fashion, like a syncitium [62,334,335,336].
5.2. Diversity of Ecs in the Large Blood Vessels
The expression of genes specific for the macrocirculation bed contributes to modulation and biosynthesis of the corresponding ECM: fibronectin, collagen (5α1 and 5α), and osteonectin [337]. The main epigenetic microcirculatory factors are arterial pressure, hypercholesterolemia, and inflammation [338,339,340].
Morpho-functionally, arterial ECs (AECs) and venous ECs (VenECs) specialize in the non-stop conduction of oxygen-rich/poor blood towards or away from the heart. Macrocirculatory ECs participate in the systemic control of blood pressure and the regulation of blood flow (shear stress) and vasomotor tone (circumferential wall stress). Therefore, they are also involved in the maintenance of the total peripheral vascular resistance (elastance and compliance) and vascular capacitance, activation and migration of blood cells/immune cells, as well as in vascular disease development [263,306,307]. AECs have elongated and narrow shapes. They contribute to the tunica intima, which is reinforced by smooth muscle cells. In comparison with VenECs, AECs do not participate in the formation of valves. Additionally, AECs provide fewer conduits for transmigration of immune cells [341,342] and play a lesser role in terms of inflammation when compared with the venous endothelium [343]. Arterial and venous ECs belong to the continuous endothelial cell type. AECs and VenECs contain TJs and AJs, which provide cell–cell interactions and secure vessel stability (providing laminar blood flow and reducing vascular shear stress).
5.3. Lymphatic Macro/Microcirculation
The lymphatic system belongs to the drainage system of the body, and is closely related to blood circulation [344]. It furnishes the peripheral immune system [345] and nervous system with drainage of the extracellular liquid and corresponding clearance of antigens contributing to the maintenance of brain homeostasis [346,347]. The lymphatic system’s essential functions include the removal of interstitial fluid and the maintenance of local homeostasis; local tissue immunological supervision; and host defense, which entails cell trafficking and transcytotic delivery in the guidance and support of immunocompetent cells. It is also involved in the absorption of dietary lipids and participates in vessel and organo-vascular morphogenesis [348]. The morpho-physiological organization of the lymphatic bed determines its functionality.
The lymphatic capillaries consist of a single layer of lymphatic ECs (LECs) characterized by different cellular integrity levels and the absence of a basal lamina, in contrast to most blood vessels [349]. The “initial blind-ended” capillaries eliminate intercellular overflow and regulate the macromolecular balance in the interstitial space. LECs in the initial capillaries interconnect directly with the interstitial matrix, forming the specific discontinuous oak-leaf shape and button-like junctions (buttons) [350]. The larger collecting capillaries drain into the thoracic or right lymphatic duct and, finally, to the brachiocephalic veins. The level of cell–cell integrity between “collecting” LECs is higher as compared to the “initial” LECs, which closely cooperate with smooth muscle cells or pericytes, build continuous zipper-like junctions (zippers), and additionally acquire intraluminal valves. This architecture provides permanent unilateral lymphatic flow [351]. Ontogenetically, the zippers are older than the buttons [352]. Both junction types demonstrate a high capacity for plasticity under various physiological and/or pathophysiological demands; mature junctions are able to reorganize junctional proteins and the cytoskeleton [283]. Inflammation and infection stimulate the transformation of the existing button-like junctions into zipper junctions [302]. Organ-specific functions may be determined by the source of lymphatic origin and cellular microenvironment [348]. In early development, the lymphatic vessels arise mainly from large veins. In the postnatal period, lymphatic development occurs by reorganizing lymphatic capillaries or by transdifferentiation of venous, mesenteric, and hemogenic endothelium [353]. Notably, human and mouse myeloid lineage cells can successfully transdifferentiate to LECs through the activation of toll-like receptor-4 (TLR4) [354]. PROX1, SOX18, NOTCH, Wingless-related integration site (WNT), COUP transcription factor 2, AM-CRL-RAMP2, angiopoietin-TIE, VEGF, VEGFR3, and sphingosine 1-phosphate (S1P)- sphingosine 1-phosphate receptor (S1PR1) participate in the control of the lymphatic fate [355,356,357,358,359]. VEGFR3, VEGFC, PROX1, NOTCH, and LYVE l are responsible for lymphatic migration and sprouting; the specification of LECs is derived by WNT, BMP, and JAGGED1/NOTCH1 signaling pathways [360]. The angiopoietin-TIE is necessary for cardiovascular and lymphatic development, including remodeling [361,362]. S1P and S1PR1 also participate in LEC remodeling, development, lymphatic valve formation, endothelial barrier function, dilatation of vessels, and inflammation [363]. The dysfunction of the lymphatic system may result in the initiation and progression of a wide range of diseases. Decreased lymphatic drainage and cerebrospinal fluid flow aggravate glioblastoma progression and may influence the onset of pre-senility in the elderly [347,364]. Metabolic disorders such as diabetes mellitus, obesity, and metabolic syndrome are closely related to the dysfunction of the lymphatic system. They are characterized by chronic inflammation, low inflammatory answer, development of secondary lymphedema, and lipedema [365,366].
6. Selected Tissue-Specific Endothelial Phenotypes
The brain, heart, lungs, liver, kidneys, gut, and endocrine organs have distinct endothelial subpopulations displaying specific characteristics.
Brain endothelial cells (BECs), along with pericytes and astrocytes, form the blood–brain barrier and enforce the protective barrier function [367]. The relationships between pericytes and ECs (including BECs) are dynamic and vital for the integrity and maintenance of vessel walls [368]. Functionally, pericytes play a vital role in modulating blood flow, supporting angiogenesis, and contributing to the stability and maturation of blood vessels [369]. These functions, first of all, are related to specific morphological organization and, secondly, to metabolic interactions between these types of cells. Morphologically, pericytes are embedded in the basal lamina of the ECs (including BECs) within capillaries and postcapillary venules [370,371,372]. This anatomical proximity facilitates direct cell–cell communication, mainly through the Gap junctions, positioning pericytes as principal homeostasis regulators of the local microenvironment and vessel stability [370]. Metabolic interaction between ECs (including BECs) and pericytes, based on pericytes’ preferential use of oxidative phosphorylation (OXPHOS) for ATP production, makes pericytes the primary stabilizers of vessel wall integrity in the vascular system [373]. Through these diverse functions, pericytes ensure the proper functioning of the vascular system and respond to various physiological and pathological conditions over the vascular bed.
BEC morphology itself is specific and characterized by numerous caveolae, endothelial TJs, AJs, and an increased number of mitochondria [374]. The BEC surface displays dozens of specific proteins, regulating cell–cell communication and molecular transport: claudin-5, occludin 3,12 (OCCLN 3,12), endothelial cell-selective adhesion molecule (ESAM), junction molecules 1, 3 (JAM1,3), tricellulin- and lipolysis-stimulated lipoprotein receptor (LSR), vascular endothelium cadherin (VE-cadherin), N-cadherin, and transporters (Glut-1, Slc2a1) [375,376]. Notably, the BECs do not express thrombomodulin [377]. The morphology, proteome composition, and expression of CD31 and von Willebrand factor differ in various brain parts and types of vessels (capillaries, arterioles, venules) [378,379]. The local cellular environment mediates BEC functionality: microglia change BBB (blood–brain barrier) permeability, also causing systemic inflammation [380], and BEC–macrophage communication instigates barrier dysfunction in patients with hypertension [381]. Plasmodium parasites initiate BBB disruption, resulting in edema [382]. Gamma interferon contributes to the BBB leakage process [383].
Endocardial ECs (EECs) display characteristics typical of the continuous vascular endothelium. The EECs form a thin cell layer with varying thicknesses ranging from 50 to 300 μm. Primarily, they contribute to normal blood flow and physiological cardiac function. The total amount of EECs in the heart corresponds to 2–3% of the heart mass [384]. These cells are characterized by the unique presence of numerous microvilli on the surface, and their basal lamina is composed of delicate collagen and elastic fibers [385]. Compared to vascular ECs of arteries, veins, and capillaries, the EECs have a broader shape; particular cellular connections (e.g., GJs) and intercellular spaces define the specificity of these EECs. The GJs control the transendothelial permeability of EECs through quick passage of charged ions (mostly Ca2+), messenger molecules, and small metabolites [386]. Metabolically active EECs use fatty acid and lactate as energy sources [387] and control long-chain fatty acid (LCFA) delivery and metabolite transport to myocytes [388]. GLUT-1,3,4 mediates the primary glucose uptake in EECs, while GTPases participate in cellular protein transport [1,389]. The condition of EECs is regulated by systemic and local NO synthases, endothelin-1 (ET-1), angiotensin II, prostacyclin, natriuretic peptides A and B, VEGF, hepatocyte growth factor (HGF), FGF, interleukin-6 (IL-6), thrombospondins, and insulin-like growth factor-1 (IGF-1) [390,391]. EECs express various markers that are typical for all endothelial cells, including CD31, CD34, vWf, caveolin, neuregulin-1 (NRG-1), and VE-cadherin [392]. The heart endothelium can be considered as an active paracrine, endocrine, and autocrine organ that produces cardioprotective substances like secretory leukocyte protease inhibitor (rhSLP1) [393]. Simultaneously, EECs firmly contact the cardiomyocyte surface (between the two types of cells, there is a thick fibrillary basal lamina). This cell–cell cross-talk is essential for heart regeneration and remodeling [390,394]. Release and diffusion of signaling molecules within this space participates in heart inotropy [395]. However, any extrinsic cues (factors located out of the cardiovascular system) provoke disbalance in endothelial physiology, e.g., hyperthyroidism is regarded as a factor of cardio-cerebrovascular dysfunction [396], and autoimmune diseases as well as systemic and local inflammation can damage the morphology or physiology of vascular ECs and EECs [397]. Hypoxia may cause upregulation of vWf in EECs through high-mobility group box-1 (HMGB1) and activation of toll-like receptor-2 (TLR2), resulting in an augmentation of the plasma sodium concentration, an increase in E selectin and P selectin, downregulation of anti-thrombotic factors [398], and altered innate immunity reactions [399,400].
The pulmonary ECs (PECs) contribute to the gas exchange with the external microenvironment, formation of the air–blood barrier in the alveoli, maintenance of pulmonary and systemic vascular homeostasis, and immune response (through mitochondrial activation of innate immune mechanisms that stimulate lymphatic delivery to the lymphatic nodes and promotion of adaptive immunity). They also provide a failsafe mechanism to balance blood pressure in the lungs and, possibly, to regulate coagulation with PGI2 prostaglandin [401,402]. PECs are classified as vascular (macrovascular and microvascular) or alveolar ECs. Recently, Car4+ high PECs (with high levels of Car4 and CD34 expression) were identified by scRNA-seq analysis. This cell subpopulation is located throughout the lung periphery and is primed to respond to VEGFA signaling. The number of Car4+ high PECs increases in the regenerating areas of pulmonary tissues after influenza infection [403]. The alveolar endothelium is divided into two intermingled cell types: aerocytes (specialized in the gas exchange and leukocyte trafficking) and general capillary cells (functioning as stem/progenitor cells and responsible for regulating the vasomotor tone of capillaries) [404]. The pulmonary endothelial barrier plays a vital role in vascular homeostasis maintenance. TJs of PECs are formed by occludins, claudins, and junctional adhesion molecules. Notably, primarily vascular endothelial cadherin comprises the AJs [405]. PECS is a continuous type of endothelial cell with an epithelioid shape. When metabolically active, these cells produce prostacyclin, bradykinin, angiotensin, endothelin-1, prothrombotic and anti-thrombotic factors, and other anti-inflammatory cytokines [406,407]. PECS express vWf, endothelial NO synthase, cadherins, CD31, and angiotensin I-converting enzyme (ACEI) [408,409,410].
The endothelial population of the kidney ECs (KECs) is diverse. KECs include glomerular ECs (GECs) and peritubular capillary ECs (PCECs). Both types of cells have a continuous basal lamina; however, PCECs belong to the pseudo-fenestrated and GECs to the true-fenestrated ECs (Table 1) [411]. Pseudo-fenestrations of PCECs represent incomplete fenestration with an overlying fenestral diaphragm, controlling molecular passage and exerting a “sieving” function or regulating the counter-current-based gradient in the medulla and cortex [412]. Fenestral diaphragms are typical for PCECs and intestinal ECs (but not for GECs and form open holes); they are composed of radial heparan sulfate proteoglycan fibrils (glycocalyx tufts) acting as a permselective barrier and regulating the passage of water and small molecules [296]. Electron microscopy of GECs has revealed a thick (around 200 nm) glycocalyx layer [411,413,414]. GECs express vWf, vascular cell adhesion protein 1 (VCAM1), and intercellular adhesion molecule-1 (ICAM-1). GECs, together with mesenchymal cells, produce HGF, Kruppel-like factor (KLF), and insulin-like growth factor-binding proteins (IGFBPs) through the activation of the c-Met receptor [415]. PCECs sit on the continuous basal lamina, which separates them from the pericytes. PCECs display some immunohistochemical characteristics of macrophages: OKM5 (medullary expression only, there is no expression of OKM5 in GECs) and interleukin-1 (IL-1) expression [416,417]. In adults, PCECs display CD31 and VE-cadherin, a low level of vWf, and overexpression of the plasmalemma vesicle-associated protein 1 (PV1). GECs do not have the PV1 [286]. According to recent data, CD34 has been revealed in the peritubular microvasculature of the adult human kidney, and the level of expression usually correlates with the severity of glomerular and tubulointerstitial damage [286,418,419].
Intestinal ECs (IECs) belong histologically to the pseudo-fenestrated endothelium with TJs and fenestral diaphragms. These cells are essential for local and general immune responses and the development of intestinal inflammation [420]. The IECs possess fenestrated diaphragms. This fenestrated diaphragm, along with the gastrointestinal mucosa, forms a selective barrier between the extracellular and intravascular space (so-called gut-blood barrier) [421]. The healthy intestinal barrier is impermeable to 70 kDa molecules [422]. TJs and claudin proteins regulate the functionality of the intestinal microvascular endothelial barrier. The overexpression of claudin-1 increases the antiviral and antibacterial resistance of IECs by intensifying the mucosal and endothelial integrity [423]. Downregulation of claudin-5 and claudin-8 increases barrier permeability [424]. The tight junctions belong to a dynamic structure that can adapt its protein composition according to external (pathological or physiological) stimuli [425]. The primary markers of IECs are CD31, vWf, VE-cadherin [426], and E-selectin. P-selectin, VCAM-1, and ICAM-1 cannot be detected in the basal, unstimulated state. However, the pro-inflammatory cytokines Il-1β and tumor necrosis factor-α (TNF-α) can induce the biosynthesis of the latter proteins [427,428].
The liver sinusoid ECs (LSECs) are metabolically active, organ-specific ECs with great transdifferentiation potential. Phenotypically, LSECs form a discontinuous fenestrated endothelium with a lacking basal lamina [429]. LSECs are organized in sieve plates and display plenty of fenestrations: ~2–20 fenestrations per μm2, corresponding to 2–10% of the LSECs surface. The diameter and distribution of fenestrations allow for rapid exchange between the space of Disse and the blood [430]. LECs are essential for: (1) the primary selective barrier (protecting liver parenchyma); (2) the formation of scavenger and endocytosis systems of the liver; (3) immune response; (4) paracrine signaling; and (5) liver regeneration [431,432]. Discontinuous fenestrated LECs express such specific markers as stabilin-1, stabilin-2, liver endothelial differentiation association protein (LEDA-1), and CD32b [433]. Additionally, mannose receptors (CD206) and toll-like receptors are present on the surface of the LSEC, while the CD31 and vWf are not expressed in the LSCs under normal conditions or in young individuals, but may be present in ECs of larger blood vessels and lymphatic vessels in the liver [434,435]. VEGFR3 is often used as a specific marker for LECs (FLT-4) [436].
Splenic sinusoidal ECs (SSECs) are the most abundant non-immune cells of the spleen which participate in forming the splenic sinusoidal wall and create a discontinuous endothelial layer adjacent to the fenestrated basal membrane. The spleen is a secondary lymphatic organ involved in immunological supervision, clearance of the blood, and maturation of immune cells [437]. The endothelium forms filamentous structures, contributing to radial construction and free, yet limited and retarded, blood passage [438,439]. Open blood flow is found in splenic cords [440]. The fibers around the SSECs are associated with cellular VE-cadherin, β-catenin, p120 catenin, and actin filaments [441]. Thus, SSECs, together with spleen littoral cells, specialize in the filtration of senescent red blood cells [442]. SSECs are characterized by expression of CD31, CD8 α/α, CD271, stabilin-1, and CD206 [442,443]. The vWf is another marker of SSECs which is upregulated by hypothermic conditions [444].
7. Alteration of Endothelial Cells and Pathologies
7.1. Cancer
ECs are structural units of the circulatory system which are essential for cancer development [445]. Cancer onset and progression are associated with the metabolic reprogramming of ECs and the formation of specific cancer-hospitable “niches” or “sub-niches” that enable tumor-associated AG and facilitate tumor growth in the body [446]. Tumor progression is an active metabolic process requiring persisting neovascularization through VG, AG, LyAG, intussusceptive AG, vessel co-option, and vasculogenic mimicry [447]. VG in tumor development is characterized by the recruitment of endothelial progenitor cells, contributing substantially to tumor formation. Similarly to AG in other contexts, cancer-related AG involves the formation of vessels from mature ECs through a sprouting process [448]. Intussusceptive AG constitutes a splitting process of an initial vessel lumen through invagination, thereby creating a new vessel [449]. The usage and alteration of existing vessels for cancer cell spreading is called “co-option” [450]. Vasculogenic mimicry in tumorigenesis is the formation of vasculogenic-like channels without the EC layer [LIT]. This is necessary for the direct transport of the tumor cells in the vascular bed [451]. Notably, the amount of ECs and precursor blood cells in patients with different types of cancer (cell lung cancer, ovarian cancer, leukemia, myeloid leukemia, hepatocellular carcinoma, and breast carcinoma) is substantially increased [452], since the cells are more often released from the BM or the systemic vascular wall [453,454]. Different populations and subpopulations of ECs, cancer-associated fibroblasts, and tumor-associated immune cells (especially myeloid-derived innate cells) can be reprogrammed by the tumor environment and incorporated into the tumor stroma [455]. Tumor progression and neovascularization require the transformation of ECs and stromal cells into morphologically abnormal, aneuploid tumor-derived ECs (TECs). TEC lines in tumor vessels display loosened junctions and increased vascular permeability, leading to tumor dissemination and the formation of hemorrhages [456]. Membrane storage in transformed endothelial cells may also contribute to the formation of larger vessels in the form of VVOs (vesiculo-vacuolar organelles) that add to the rapid extension of the bilayer surface area [457]. Cancer-related EC reprogramming results from metabolic re-education and morphological transformation, e.g., through EMT/EndMT [458]. Tumor EMT/EndMT is a dysregulated process compared to EMT/EndMT in embryonic development and tissue regeneration [459]. This process is induced and controlled by the following conditions and factors: hypoxia [460], TGF-β [461], survivin [462], endosulfan [463], and gremlin-1 [45,464]. It has recently been demonstrated that ECs promote metastatic cancer dissemination through differential expression of adhesion molecules (E-selectin, P-selectin, integrin, transpanin, trombospodins), secretion of CCL5, and enhancing autophagy [465,466]. ECs in healthy body organs are also programmed by the transfer of tumor molecules from the environment and can aid in the metastasis of the original tumor tissue [467].
Table 3.
Morpho-functional classification of vascular ECs.
Table 3.
Morpho-functional classification of vascular ECs.
| Morpho-Functional Characteristics | ECs Types | ||||
|---|---|---|---|---|---|
| Continuous | Fenestrated Endothelium | Lymphatic | |||
| Pseudo-Fenestrated Fenestrated | Glomerular Fenestrated Endothelium (True Fenestrated) | Disconntimous Fenestrated (Sinusoid) Endothelium | |||
| Localization | Brain [1,277,278] Skin [1,270] Lungs [1,295,296,302] Heart [1,270,293,295,299,302] Arteries [1,270,272,289,292,295,299] Veins [1,270,295,296,299] | Intestinal tube [270,275,278,287,293,296] Adrenal cortex [273,278,293,295] Pancreatic islets [278,293] Kidney peritubular capillaries [278,289,293,296,301] | Kidney (Glomeruli) [270,275,290,293,295,296] | Liver [270,278,293,296,299] Spleen [270,278,293] Bone-marrow [275,278,293,295,299] | Lymphatic vessels and lymph nodes [232,278,283,288,300,302] |
| Function | Highly selective barrier: transfer of water and small solutes (diameter ~6 nm), transport of big molecules occurs through channels or transcytosis [270,278] | Size and selective charge barrier: permeable for small molecules and water, but impermeable for macromolecules (e.g., albumin, peptide hormones) and blood cells [278,286,293] | Low-selective barrier: permeable for small molecules and water and macromolecules (e.g., albumin) but impermeable for cells from ultrafiltrate [290,293] | Non-selective barrier: permeable for water, macromolecules, and blood cells [291,293,297] | Non-selective barrier of lymphatic capillaries (sinusoid lymphatic ECs): permeable for macromolecules and immune cells (high permeability) Selective barrier, collecting lymphatic vessels demonstrate low permeability [288] |
| Basal lamina | Yes [270,293] | Yes [293] | Yes [293] | Absent or poorly developed [270,291,293] | Lymphatic capillaries (initial capillaries): highly incomplete perforated basal lamina and discontinuous junctions (buttons) [288,294] Collecting lymphatic vessels: continuous basal lamina, continuous junctions (zippers) [294] |
| Fenestra, nm | No [1,270,276,293] | 60–70 [273,293] | 60–100 [293,301] | “Sieve plates”: 50–100 “Gaps”: 100–200 [293,296,297] | No [288] |
| Fenestral diaphragm | No [270,276] | Yes [1,273,293] | No [296] | No [296] | No [288] |
| Glycocalyx | Yes [275] | Yes [293,296] | Yes [275,290,301] | Yes [269,377,378] | Yes [303] |
| Non-specific Markers | CD31: Heart (low expression), skin [277,285] CD34: Heart, skin [292] von Willebrand factor (vWF): Heart (low expression), skin, lungs [277,292] CD62E or E -selectin (inducible) [270,277,292] CD62P or P selectin (inducible) [270,277,292] CD106 or VCAM-1 (inducible) [277] CD54 or ICAM (inducible) [277] Flt-1 or vascular endothelial growth factor receptor 1 (VEGFR1, inducible) [277] KDR/Flk or vascular endothelial growth factor receptor 2 (VEGFR2, inducible) [277] CD144 human [26,280] | CD31 [277,285,292] CD34 [277,285,292] von Willebrand factor (vWF) peritubular ECs (low expression) Fli-1 (nuclear) [277,285,292] CD62E or E -selectin (inducible) [277,285,292] CD62P or P selectin (inducible) [277,285,292] CD106 or VCAM-1 (inducible) [277,285,292] CD54 or ICAM (inducible) [277,285,292] Flt-1 or vascular endothelial growth factor receptor 1 (VEGFR1, inducible) [277,285,292] KDR/Flk or vascular endothelial growth factor receptor 2 (VEGFR2, inducible) [277,285,292] vWf [277,285,292] | CD31 [277,285,292] CD34 [277,285,292] Fli-1(nuclear) [277,285,292] von Willebrand factor (vWF) expression) [277,285,292] CD62E or E -selectin (inducible) [277,285,292] CD62P or P selectin (inducible) [277,285,292] CD106 or VCAM-1 (inducible) [277,285,292] CD54 or ICAM (inducible) [277,285,292] Flt-1 or vascular endothelial growth factor receptor 1 (VEGFR1, inducible) [277,285,292] KDR/Flk or vascular endothelial growth factor receptor 2 (VEGFR2, inducible) [277,285,292] | CD31: Liver, Spleen; Bone marrow [277,292] CD34: Bone marrow [277,292] von Willebrand factor (vWF): liver, spleen [61,78] Fli-1: Liver, spleen, bone marrow [277,292] CD62E or E -selectin (inducible) [277,291] CD62P or P selectin (inducible) [277] CD106 or VCAM-1 (inducible) [277] CD54 or ICAM (inducible) [277] Flt-1 or vascular endothelial growth factor receptor 1 (VEGFR1, inducible) [277] KDR/Flk or vascular endothelial growth factor receptor2 (VEGFR2, inducible) [277] | CD31 [277,285,292] CD34 [278,284,292] Fli-1 [284,292] von Willebrand factor (vWF) [284,292] CD62E or E -selectin (inducible) [277,284] CD62P or P selectin (inducible) [277] CD106 or VCAM-1 (inducible) [277] CD54 or ICAM (inducible) [277] Flt-1 or vascular endothelial growth factor receptor 1 (VEGFR1, inducible) [277] KDR/Flk or vascular endothelial growth factor receptor 2 (VEGFR2, inducible) [277] |
| Specific markers | Angiotensin-converting enzyme ACE or CD143 (human heart and lungs) [277,278] Thrombomodulin (TM): absent in brain endothelial cells [277,278] Tissue non-specific alkaline phosphatase or TNAP: brain (mouse and human and rat) [276,278] Thrombospondin type 1 domain or THSD1: vessels (mouse and human) [278,281] P-glycoprotein or MDR 1a: brain and lungs (mouse and human) [20,277] CD73/ transferrin receptor: brain (mouse and human) [277] Platelet-derived growth factor receptor: brain (human and mouse) [277,282] Sca-1 (mouse pulmonary ECs) [277] HLA-DR (human, pulmonary ECs) [280] Glut-1: brain (human and mouse) [277] | PV1 (human and mouse peritubular capillary) [293,295,296,299] MAdCAM-1 (venules intestinal) [277] Nephrin (human Pancreatic islet) [280] CD117 (mouse pancreatic islet) [278] | ADAMTS-13 (mouse and human) [298] | CD32b (human liver sinusoidal) [280,336,434] LYVE-1 (mouse liver and spleen sinusoidal) [278,304] PV-1 (mouse spleen sinusoidals) [280,296] Angiotensin-converting enzyme (ACE or CD143) Stabilin 1,2 [434] Liver-endothelial differentiationassociation protein (LEDA-1) [434] | CD90 (human and mouse) [280] Flt-4 or vascular endothelial growth factor receptor 3 (VEGFR3, human and mouse) [277] Desmoplakin [277] Podoplanin or PDPN (human) [278,280] LYVE-1 (human and mouse) [278,280] Prox-1 (human and mouse) [280,294] Clever-1 or Stabilin-1 or FEEL-1 (human and mouse) [231,280,303] |
| Junctions | Tight junctions/adherence junctions [272] | Tight junctions/gap junctions [272,279] | Tight junctions/gap junctions [272,279] | Gap junctions/tight junctions [272,279,302] | Buttons (discontinuous button-like junctions, with openings at the borders of adjacent lymphatic ECs) enriched with adherents and tight junction proteins [350,353] Zippers (continuous zipper-like junctions without openings at the borders of adjacent lymphatic ECs) enriched with adherent and tight junction proteins [350,353] |
7.2. Endothelial Turnover, Regeneration, and Repair
The turnover of the ECs in the vascular system includes replacing the old endothelial cells with new endothelial cells. This process reflects the balance between endothelial proliferation and apoptosis rates. Endothelial-to-endothelial turnover takes from 47 days to 23,000 days and increases under the influence of intrinsic or extrinsic factors such as hemodynamic forces, shear stress, pressure, ischemia, certain vascular and non-vascular diseases, and continuous local or systematic endothelial injury [468,469,470,471]. Regulation of the endothelial-to-endothelial turnover is controlled by Rasa 3 (decreasing level of Rasa3 increases cell adhesion), occludin, VE-cadherin, ZO-1 (ensuring maintenance of vascular integrity in vertebrates), junctional adhesion molecule C (JAM-C-1 responsible for the endothelial cell migration), VEGF (induces endothelial leakage through the Src-mediated degradation of VE-cadherin), angiopoietin-1, mitogen-activated protein-4 kinase-4 (MAP4K4), and Il-2 [472,473,474,475,476,477]. The turnover is a neighbor-depending process, requiring extra- and intracellular matrix reorganization and loss of cell–cell contacts. The leading suppliers of cells for EC turnover are blood-circulating endothelial cells, local endothelial progenitor cells, and bone marrow-derived SCs [468,478]. The endothelial turnover in the brain is low. However, the situation changes in the case of brain injury, where an increased amount of the neurovascular unit cells (ECs, pericytes, astrocytes) or endothelial progenitors may participate in endothelial recovery [479]. Compared to other microvascular ECs, the human brain endothelial cells react differently to shear stress factors, i.e., the turnover rate decreases with the increasing shear stress level [480]. According to the “nuclear bomb-derived C14 analysis,” the heart/cardiac endothelial cell turnover is high (more than 15% of all ECs per year) compared with mesenchymal cells (less than 4% per year) or cardiomyocytes (less than 1% per year) [481]. In contrast to the brain ECs, in injury of the heart, the pre-existing ECs are the primary source for endothelial recovery [482]. The daily pulmonary endothelial turnover involves approximately 1% of all ECs [483]. In the case of lung injury, the local EPCs are recruited in the turnover process [484]. The sinusoidal endothelial cell progenitors can replace the mature LSECs and the intrahepatic or resident sinusoidal ECs. Interestingly, those mature LSECs can proliferate and expand with the involvement of VEGF and FGF. BM progenitor cells, on the other hand, do not participate in liver turnover under physiological conditions, but become relevant in liver pathologies [291,485,486]. The splenic endothelial turnover recruits the local EPCs, which closely interact with the sinusoidal endothelial cells [487]. Endothelial derivates make up around 30% of the spleen [488]. Local EPCs (colony-forming and pro-angiogenic cells) are the principal elements of kidney EC turnover [489]. The intestinal EC homeostasis also builds on a unique, local intestinal stem cell niche for renewal and proliferation [490].
Endothelial repair is a vital process for endothelial physiology. Persistent endothelial damage and impaired endothelial regeneration lead to local or systemic endothelial dysfunction and loss of the primary barrier function. Endothelial repair comprises shock, proliferation, acclimation, and endothelial maturation stages. Augmentation of encoding transcripts (FOAS, FOSB, FOSL1 FOSL2, JUNB), stressor protein (activator protein-1, AP-1), and VE-cadherin two hours after endothelial damage are signs of the shock stage. Increased Myc activity, cyclins D1/E2 (cell-cycle genes), and VEGF growth factor initiate proliferative mechanisms. The acclimation process is associated with a robust inflammatory response and remodeling of the basal extracellular matrix. The last phase of endothelial regeneration involves reorganization of the extracellular matrix through the activation and expression of the COL5A1, COL5A2, COL8A1, NID1, and LAMC 3 genes [491]. Additionally, BMP [492,493], hypoxia-inducible factor 1 α (HIF-1α), apolipoprotein (A-I), high-density lipoproteins (HDL), zinc finger transcription factor (ZFP580), syndecan-1, and ixmyelocel-T stimulate endothelial regeneration [494]. Endothelial regeneration can be facilitated by intercellular interplay with surrounding cells (regeneration satellites). Smooth muscle cells, for example, mediate the EC recruitment in a PKCδ-dependent manner by releasing the chemokine (C-X-C motif) ligand 7 (CXCL7) [495]. Angiogenic T-cells and monocytes also contribute to endothelial repair [496,497]. M2 macrophages stimulate endothelial proliferation [498] in lung injury and may attach to endothelial cells in this reaction [498,499,500].
Endothelial turnover, regeneration, and repair are crucial processes for maintaining vascular health throughout life and in the post-injury period. These processes are tightly regulated to ensure proper endothelial function. Quality control mechanisms maintain the functionality of the endothelial layer, degradation processes remove damaged cells, and regeneration and repair mechanisms restore vascular integrity.
These processes must be precisely balanced; dysregulation of any of them leads to the initiation of various pathological conditions, including inflammation, cancer, cardiovascular issues, nervous dysfunctions, etc.
Key endothelial control mechanisms:
(1) Autophagy: removing damaged organelles and proteins [501]; (2) unfold protein response (UPR) is realized when the misfolded proteins are accumulated in the endoplasmic reticulum [502,503]; (3) DNA damage responses (DDR) are crucial for the detection and repair of DNA damage [504,505].
The degradation processes consist of (1) the ubiquitin–proteasome system (UPS) (tags damaged or unnecessary proteins with ubiquitin for degradation) [506,507]; (2) lysosomal degradation (breaks down cellular waste) [508]; and (3) caspase-mediated degradation (activated during apoptosis) [509].
Regeneration mechanisms are composed of: (1) proliferation of existing ECs [510,511]; (2) transdifferentiation (cells change from one type to another and replenish the endothelial layer) [512]; (3) recruiting of circulating cells (circulating cells differentiate into mature ECs to aid in repair) [513].
Repair mechanisms include (1) wound healing (natural response to injury) [514]; (2) shear stress active adaptation (ECs adapt to changes in blood flow) [515]; and (3) EndMT (ECs transform into mesenchymal cells to aid in tissue repair and remodeling) [516].
Author Contributions
A.L. wrote the review; L.F., C.M.H. and K.F. revised the article. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| AECs | arterial ECs |
| ANG1 and ANG 2 | angiopoietins 1 and 2 |
| ACE1 | Angiotensin 1-converting enzyme |
| BMP | bone morphogenetic proteins pathway |
| DNA | deoxyribonucleic acid |
| ECM | extracellular matrix |
| ECs | endothelial cells |
| EMT | epithelial–mesenchymal transition |
| EndMT | endothelial-to-mesenchymal transition |
| ER71 (ETV2) | ETS variant transcription factor 2 |
| ESAM | Endothelial cell-selective adhesion molecule |
| ETS family | E twenty-six family transcription factors |
| FGF | fibroblast growth factor |
| FLI1 | friend leukemia integration-1 transcription factor |
| HES1, 2 | hairy and enhancer of split 1, 2 |
| HEY1 | hairy/enhancer of split related with YRPW motif protein 1 |
| HGF | hepatocyte growth factor |
| HIF-1α | hypoxia-inducible factor 1α |
| Hh | Hedgehog |
| ICAM-1 | intercellular adhesion molecule-1 (or CD54) |
| IHECs | intraembryonic hemogenic endothelial cells |
| LMO2 | Lim domain only 2 |
| LSECs | liver sinusoid ECs |
| LYVE 1 | lymphatic vessel endothelial hyaluronan receptor |
| MAML1,2,3 | mastermind-like protein 1,2,3 |
| MAP4K4 | mitogen-activated protein-4-kinase 4 |
| NRP 1,2 | neuropilin1, 2 |
| PI3K/Akt | phosphoinositide 3-kinase/protein kinase B |
| PDGF | Platelet-derived growth factor |
| PDGFR-β | Platelet-derived growth factor receptor-β |
| RNA | ribonucleic acid |
| RUNX1 | runt-related transcription factor 1 |
| S1PR1 | sphingosine 1-phosphate (S1P)–sphingosine 1-phosphate receptor |
| SCL/TAL1 or TAL1 or SCL | stem cell leukemia/or T-cell acute lymphocytic leukemia-1 |
| scRNA-seq analysis | single-cell RNA sequencing analysis |
| SFRP | secret frizzled-related protein |
| SMADs | homologies to SMA (“small” worm phenotype) and MAD family (“Mothers Against Decapentaplegic”) genes |
| SSECs | spleen sinusoid ECs |
| TAK-1 | transforming growth factor-beta activated kinase 1 |
| TCF7L2 | transcription factor 7-like 2 (TCF4) |
| TGFβ | α-transforming growth factor β |
| TIE | tyrosine kinase with immunoglobulin-like and EGF-like domains |
| TLR2 | toll-like receptor-2 |
| TNF-α | tumor necrosis factor-α |
| TWIST1 | twist family of basic helix-loop-helix protein 38 (bHLHa38) transcription factor 1 |
| VCAM1 | vascular cell adhesion protein1 |
| VE-cadherin | vascular endothelial cadherin (cadherin-5 or CD144) |
| VEGF | vascular endothelial growth factor |
| VEGFR-1 (Flt 1) | vascular endothelial growth factor receptor 1 |
| VEGFR2 (Flk1/KDR) | vascular endothelial growth factor receptor 2 |
| VenECs | venous ECs |
| vWf | von Willebrand factor |
| WNT | Wingless-related integration site |
References
- Krüger-Genge, A.; Blocki, A.; Franke, R.-P.; Jung, F. Vascular Endothelial Cell Biology: An Update. Int. J. Mol. Sci. 2019, 20, 4411. [Google Scholar] [CrossRef] [PubMed]
- Paul, K.; Raymond, T.; Frances, P. Endothelium. In Mechanisms of Vascular Disease: A Reference Book for Vascular Specialists; Fitridge, R., Thompson, M., Eds.; Barr Smith Press: Adelaide, Australia, 2011; pp. 1–12. ISBN 978-0-9871718-2-5. [Google Scholar]
- Michiels, C. Endothelial Cell Functions. J. Cell. Physiol. 2003, 196, 430–443. [Google Scholar] [CrossRef] [PubMed]
- Eelen, G.; de Zeeuw, P.; Treps, L.; Harjes, U.; Wong, B.W.; Carmeliet, P. Endothelial Cell Metabolism. Physiol. Rev. 2018, 98, 3–58. [Google Scholar] [CrossRef] [PubMed]
- Wong, W.T.; Huang, N.F.; Botham, C.M.; Sayed, N.; Cooke, J.P. Endothelial Cells Derived from Nuclear Reprogramming. Circ. Res. 2012, 111, 1363–1375. [Google Scholar] [CrossRef] [PubMed]
- Al-Soudi, A.; Kaaij, M.H.; Tas, S.W. Endothelial Cells: From Innocent Bystanders to Active Participants in Immune Responses. Autoimmun. Rev. 2017, 16, 951–962. [Google Scholar] [CrossRef]
- Jeong, H.W.; Hernández-Rodríguez, B.; Kim, J.M.; Kim, K.P.; Enriquez-Gasca, R.; Yoon, J.; Adams, S.; Schöler, H.R.; Vaquerizas, J.M.; Adams, R.H. Transcriptional Regulation of Endothelial Cell Behavior during Sprouting Angiogenesis. Nat. Commun. 2017, 8, 726. [Google Scholar] [CrossRef] [PubMed]
- Singh, I. Textbook of Human Histology (with Colour Atlas and Parctical Guide), 6th ed.; Vij, J., Ed.; Jaypee Brothers Medical Publishers (P) Ltd.: New Dehli, India, 2011; ISBN 978-93-80704-34-0. [Google Scholar]
- Potter, C.M.F.; Schobesberger, S.; Lundberg, M.H.; Weinberg, P.D.; Mitchell, J.A.; Gorelik, J. Shape and Compliance of Endothelial Cells after Shear Stress In Vitro or from Different Aortic Regions: Scanning Ion Conductance Microscopy Study. PLoS ONE 2012, 7, e31228. [Google Scholar] [CrossRef]
- Malek, A.M.; Izumo, S. Mechanism of Endothelial Cell Shape Change and Cytoskeletal Remodeling in Response to Fluid Shear Stress. J. Cell Sci. 1996, 109, 713–726. [Google Scholar] [CrossRef]
- Eroschenko, V.P. Atlas of Histology, 10th ed.; Taylor, C., Ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2008; p. 19106. ISBN 13: 978-0-7817-7057-6. [Google Scholar]
- Milton, S.G.; Knutson, V.P. Comparison of the Function of the Tight Junctions of Endothelial Cells and Epithelial Cells in Regulating the Movement of Electrolytes and Macromolecules across the Cell Monolayer. J. Cell. Physiol. 1990, 144, 498–504. [Google Scholar] [CrossRef]
- Liepsch, D.W.; Levesque, M.; Nerem, R.M.; Moravec, S.T. Correlation of Laser-Doppler-Velocity Measurements and Endothelial Cell Shape in a Stenosed Dog Aorta. Adv. Exp. Med. Biol. 1988, 242, 43–50. [Google Scholar] [CrossRef]
- Palhol, J.S.C.; Balia, M.; Sánchez-Román Terán, F.; Labarchède, M.; Gontier, E.; Battefeld, A. Direct Association with the Vascular Basement Membrane Is a Frequent Feature of Myelinating Oligodendrocytes in the Neocortex. Fluids Barriers CNS 2023, 20, 24. [Google Scholar] [CrossRef]
- Stratman, A.N.; Davis, G.E. Endothelial Cell-Pericyte Interactions Stimulate Basement Membrane Matrix Assembly: Influence on Vascular Tube Remodeling, Maturation, and Stabilization. Microsc. Microanal. 2012, 18, 68–80. [Google Scholar] [CrossRef] [PubMed]
- Lampugnani, M.G.; Dejana, E. Interendothelial Junctions: Structure, Signalling and Functional Roles. Curr. Opin. Cell Biol. 1997, 9, 674–682. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.-Z.; Lei, S. Changes of Junctions of Endothelial Cells in Coronary Sclerosis: A Review. Chronic Dis. Transl. Med. 2016, 2, 22–26. [Google Scholar] [CrossRef]
- Angulo-Urarte, A.; Casado, P.; Castillo, S.D.; Kobialka, P.; Kotini, M.P.; Figueiredo, A.M.; Castel, P.; Rajeeve, V.; Milà-Guasch, M.; Millan, J.; et al. Endothelial Cell Rearrangements during Vascular Patterning Require PI3-Kinase-Mediated Inhibition of Actomyosin Contractility. Nat. Commun. 2018, 9, 4826. [Google Scholar] [CrossRef] [PubMed]
- Canse, C.; Yildirim, E.; Yaba, A. Overview of Junctional Complexes during Mammalian Early Embryonic Development. Front. Endocrinol. 2023, 14, 1150017. [Google Scholar] [CrossRef]
- Taddei, A.; Giampietro, C.; Conti, A.; Orsenigo, F.; Breviario, F.; Pirazzoli, V.; Potente, M.; Daly, C.; Dimmeler, S.; Dejana, E. Endothelial Adherens Junctions Control Tight Junctions by VE-Cadherin-Mediated Upregulation of Claudin-5. Nat. Cell Biol. 2008, 10, 923–934. [Google Scholar] [CrossRef]
- Ahmad, U.S.; Uttagomol, J.; Wan, H. The Regulation of the Hippo Pathway by Intercellular Junction Proteins. Life 2022, 12, 1792. [Google Scholar] [CrossRef]
- Nakahara, S.; Tsutsumi, K.; Zuinen, T.; Ohta, Y. FilGAP, a Rho-ROCK-Regulated GAP for Rac, Controls Adherens Junctions in MDCK Cells. J. Cell Sci. 2015, 128, 2047–2056. [Google Scholar] [CrossRef]
- Mills, S.E. Histology for Pathologists, 4th ed.; Pine, J.W., Ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2012; p. 19103. ISBN 978-1-4511-1303-7. [Google Scholar]
- Witjas, F.M.R.; van den Berg, B.M.; van den Berg, C.W.; Engelse, M.A.; Rabelink, T.J. Concise Review: The Endothelial Cell Extracellular Matrix Regulates Tissue Homeostasis and Repair. Stem Cells Transl. Med. 2019, 8, 375–382. [Google Scholar] [CrossRef]
- Rekad, Z.; Ruff, M.; Radwanska, A.; Grall, D.; Ciais, D.; Van Obberghen-Schilling, E. Coalescent RNA-Localizing and Transcriptional Activities of SAM68 Modulate Adhesion and Subendothelial Basement Membrane Assembly. Elife 2023, 12, e85165. [Google Scholar] [CrossRef] [PubMed]
- Marcu, R.; Choi, Y.J.; Xue, J.; Fortin, C.L.; Wang, Y.; Nagao, R.J.; Xu, J.; MacDonald, J.W.; Bammler, T.K.; Murry, C.E.; et al. Human Organ-Specific Endothelial Cell Heterogeneity. iScience 2018, 4, 20–35. [Google Scholar] [CrossRef] [PubMed]
- Aird, W.C. Endothelial Cell Heterogeneity. Cold Spring Harb. Perspect. Med. 2012, 2, a006429. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.R.; Rajan, K.C.; Blanks, A.; Li, Y.; Prieto, M.C.; Meadows, S.M. Endothelial Cell Polarity and Extracellular Matrix Composition Require Functional ATP6AP2 during Developmental and Pathological Angiogenesis. JCI Insight 2022, 7, e154379. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.; Veloso, A.; Wu, J.; Katrukha, E.A.; Akhmanova, A. Control of Endothelial Cell Polarity and Sprouting Angiogenesis by Noncentrosomal Microtubules. Elife 2018, 7, e33864. [Google Scholar] [CrossRef] [PubMed]
- Wolpe, A.G.; Ruddiman, C.A.; Hall, P.J.; Isakson, B.E. Polarized Proteins in Endothelium and Their Contribution to Function. J. Vasc. Res. 2021, 58, 65–91. [Google Scholar] [CrossRef]
- Castiglioni, V.G.; Pires, H.R.; Bertolini, R.R.; Riga, A.; Kerver, J.; Boxem, M. Epidermal PAR-6 and PKC-3 Are Essential for Postembryonic Development of Caenorhabditis Elegans and Control Non-Centrosomal Microtubule Organization. bioRxiv 2020, 2020.07.23.217679. [Google Scholar] [CrossRef]
- Sanchez, A.D.; Branon, T.C.; Cote, L.E.; Papagiannakis, A.; Liang, X.; Pickett, M.; Shen, K.; Jacobs-Wagner, C.; Ting, A.; Feldman, J. Proximity Labeling at Non-Centrosomal Microtubule-Organizing Centers Reveals VAB-10B and WDR-62 as Distinct Microtubule Regulators. bioRxiv 2020, 2020.08.29.272369. [Google Scholar] [CrossRef]
- Becker, R.; Vergarajauregui, S.; Billing, F.; Sharkova, M.; Lippolis, E.; Mamchaoui, K.; Ferrazzi, F.; Engel, F.B. Myogenin Controls via AKAP6 Non-Centrosomal Microtubule-Organizing Center Formation at the Nuclear Envelope. Elife 2021, 10, e65672. [Google Scholar] [CrossRef]
- Prassanawar, S.S.; Sarkar, T.; Panda, D. CEP41, a Ciliopathy-Linked Centrosomal Protein, Regulates Microtubule Assembly and Cell Division. bioRxiv 2023. [Google Scholar] [CrossRef]
- Ong, S.T.; Chalasani, M.L.S.; Fazil, M.H.U.T.; Prasannan, P.; Kizhakeyil, A.; Wright, G.D.; Kelleher, D.; Verma, N.K. Centrosome- and Golgi-Localized Protein Kinase N-Associated Protein Serves as a Docking Platform for Protein Kinase A Signaling and Microtubule Nucleation in Migrating T-Cells. Front. Immunol. 2018, 9, 397. [Google Scholar] [CrossRef] [PubMed]
- Rajeev, R.; Mukhopadhyay, S.; Bhagyanath, S.; Priya, M.R.S.D.; Manna, T.K. TACC3–Ch-TOG Interaction Regulates Spindle Microtubule Assembly by Controlling Centrosomal Recruitment of γ-TuRC. Biosci. Rep. 2023, 43, 20221882. [Google Scholar] [CrossRef] [PubMed]
- Worzfeld, T.; Schwaninger, M. Apicobasal Polarity of Brain Endothelial Cells. J. Cereb. Blood Flow Metab. 2016, 36, 340–362. [Google Scholar] [CrossRef] [PubMed]
- Yamashiro, Y.; Ramirez, K.; Nagayama, K.; Tomita, S.; Kubota, Y.; Yanagisawa, H. Partial Endothelial-to-Mesenchymal Transition (EndMT) Contributes to Lumen Re-Organization after Carotid Artery Ligation. bioRxiv 2021, 2021.08.13.456319. [Google Scholar] [CrossRef]
- Piera-Velazquez, S.; Jimenez, S.A. Endothelial to mesenchymal transition: Role in physiology and in the pathogenesis of human diseases. Physiol. Rev. 2019, 99, 1281–1324. [Google Scholar] [CrossRef] [PubMed]
- Portillo-Ledesma, S.; Chung, S.; Hoffman, J.; Schlick, T. Regulation of Chromatin Architecture by Transcription Factor Binding. Elife 2023, 12, RP91320. [Google Scholar] [CrossRef]
- Trojanowski, J.; Rippe, K. Transcription Factor Binding and Activity on Chromatin. Curr. Opin. Syst. Biol. 2022, 31, 100438. [Google Scholar] [CrossRef]
- Afshar, Y.; Ma, F.; Quach, A.; Jeong, A.; Sunshine, H.L.; Freitas, V.; Jami-Alahmadi, Y.; Helaers, R.; Li, X.; Pellegrini, M.; et al. Transcriptional Drifts Associated with Environmental Changes in Endothelial Cells. Elife 2023, 12, 81370. [Google Scholar] [CrossRef] [PubMed]
- Atkins, G.B.; Jain, M.K. Role of Krüppel-Like Transcription Factors in Endothelial Biology. Circ. Res. 2007, 100, 1686–1695. [Google Scholar] [CrossRef]
- Pasut, A.; Becker, L.M.; Cuypers, A.; Carmeliet, P. Endothelial Cell Plasticity at the Single-Cell Level. Angiogenesis 2021, 24, 311. [Google Scholar] [CrossRef]
- Rohlenova, K.; Goveia, J.; García-Caballero, M.; Subramanian, A.; Kalucka, J.; Treps, L.; Falkenberg, K.D.; de Rooij, L.P.M.H.; Zheng, Y.; Lin, L.; et al. Single-Cell RNA Sequencing Maps Endothelial Metabolic Plasticity in Pathological Angiogenesis. Cell Metab. 2020, 31, 862–877.e14. [Google Scholar] [CrossRef]
- Minami, T.; Muramatsu, M.; Kume, T. Organ/Tissue-Specific Vascular Endothelial Cell Heterogeneity in Health and Disease. Biol. Pharm. Bull. 2019, 42, 1609–1619. [Google Scholar] [CrossRef] [PubMed]
- Noden, D.M. Embryonic Origins and Assembly of Blood Vessels. Am. Rev. Respir. Dis. 1989, 140, 1097–1103. [Google Scholar] [CrossRef] [PubMed]
- Hatou, S.; Shimmura, S. Review: Corneal Endothelial Cell Derivation Methods from ES/IPS Cells. Inflamm. Regen. 2019, 39, 19. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.; Guerrero-Zayas, M.I.; Wallingford, M.C.; Ortiz-Pineda, P.; Mager, J.; Tremblay, K.D. Visceral Endoderm Expression of Yin-Yang1 (YY1) Is Required for VEGFA Maintenance and Yolk Sac Development. PLoS ONE 2013, 8, e58828. [Google Scholar] [CrossRef]
- Havrilak, J.A.; Melton, K.R.; Shannon, J.M. Endothelial Cells Are Not Required for Specification of Respiratory Progenitors. Dev. Biol. 2017, 427, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Papaioannou, V.E. Early Embryonic Mesoderm Development. In Handbook of Stem Cells; Elsevier: Amsterdam, The Netherlands, 2004; Volume 1, pp. 273–278. ISBN 9780080533735. [Google Scholar]
- Julien, E.; El Omar, R.; Tavian, M. Origin of the Hematopoietic System in the Human Embryo. FEBS Lett. 2016, 590, 3987–4001. [Google Scholar] [CrossRef] [PubMed]
- Serrado Marques, J.; Teixeira, V.; Jacinto, A.; Tavares, A. Identification of Novel Hemangioblast Genes in the Early Chick Embryo. Cells 2018, 7, 9. [Google Scholar] [CrossRef] [PubMed]
- Coffin, J.D.; Harrison, J.; Schwartz, S.; Heimark, R. Angioblast Differentiation and Morphogenesis of the Vascular Endothelium in the Mouse Embryo. Dev. Biol. 1991, 148, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Vokes, S.A.; Krieg, P.A. Endoderm Is Required for Vascular Endothelial Tube Formation, but Not for Angioblast Specification. Development 2002, 129, 775–785. [Google Scholar] [CrossRef]
- Borasch, K.; Richardson, K.; Plendl, J. Cardiogenesis with a Focus on Vasculogenesis and Angiogenesis. Anat. Histol. Embryol. 2020, 49, 643–655. [Google Scholar] [CrossRef]
- Darland, D.C.; D’Amore, P.A. Blood Vessel Maturation: Vascular Development Comes of Age. J. Clin. Investig. 1999, 103, 157–158. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Flores, L.; Gutiérrez, R.; Gayoso, S.; García, M.P.; González-Gómez, M.; Díaz-Flores, L.; Sánchez, R.; Carrasco, J.L.; Madrid, J.F. Intussusceptive Angiogenesis and Its Counterpart Intussusceptive Lymphangiogenesis. Histol. Histopathol. 2020, 35, 1083–1103. [Google Scholar] [CrossRef] [PubMed]
- Drake, C.J. Embryonic and Adult Vasculogenesis. Birth Defects Res. Part C Embryo Today Rev. 2003, 69, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Fahmy-Garcia, S.; van Driel, M.; Witte-Buoma, J.; Walles, H.; van Leeuwen, J.P.T.M.; van Osch, G.J.V.M.; Farrell, E. NELL-1, HMGB1, and CCN2 Enhance Migration and Vasculogenesis, But Not Osteogenic Differentiation Compared to BMP2. Tissue Eng. Part A 2018, 24, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, J.E.; Kelley, R.W.; Patterson, C. Mechanisms of Endothelial Differentiation in Embryonic Vasculogenesis. Arter. Thromb. Vasc. Biol. 2005, 25, 2246–2254. [Google Scholar] [CrossRef] [PubMed]
- Goncharov, N.V.; Nadeev, A.D.; Jenkins, R.O.; Avdonin, P.V. Markers and Biomarkers of Endothelium: When Something Is Rotten in the State. Oxid. Med. Cell. Longev. 2017, 2017, 9759735. [Google Scholar] [CrossRef] [PubMed]
- Kolte, D.; McClung, J.A.; Aronow, W.S. Vasculogenesis and Angiogenesis; Elsevier Inc.: Amsterdam, The Netherlands, 2016; ISBN 9780128023853. [Google Scholar]
- Kütscher, C.; Lampert, F.M.; Kunze, M.; Markfeld-Erol, F.; Stark, G.B.; Finkenzeller, G. Overexpression of Hypoxia-Inducible Factor-1 Alpha Improves Vasculogenesis-Related Functions of Endothelial Progenitor Cells. Microvasc. Res. 2016, 105, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Lanzer, P. PanVascular Medicine, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 1–5004. [Google Scholar] [CrossRef]
- Lilly, A.J.; Lacaud, G.; Kouskoff, V. SOXF Transcription Factors in Cardiovascular Development. Semin. Cell Dev. Biol. 2017, 63, 50–57. [Google Scholar] [CrossRef]
- Marcelo, K.L.; Goldie, L.C.; Hirschi, K.K. Regulation of Endothelial Cell Differentiation and Specification. Circ. Res. 2013, 112, 1272–1287. [Google Scholar] [CrossRef]
- Moccia, F.; Negri, S.; Shekha, M.; Faris, P.; Guerra, G. Endothelial Ca2+ Signaling, Angiogenesis and Vasculogenesis: Just What It Takes to Make a Blood Vessel. Int. J. Mol. Sci. 2019, 20, 3962. [Google Scholar] [CrossRef]
- Patan, S. Vasculogenesis and Angiogenesis. Cancer Treat Res. 2004, 117, 3–32. [Google Scholar] [CrossRef] [PubMed]
- Patel-Hett, S.; D’Amore, P.A. Signal Transduction in Vasculogenesis and Developmental Angiogenesis. Int. J. Dev. Biol. 2011, 55, 353–369. [Google Scholar] [CrossRef] [PubMed]
- Dela Peña, I.C.; Yoo, A.; Tajiri, N.; Acosta, S.A.; Ji, X.; Kaneko, Y.; Borlongan, C.V. Granulocyte Colony-Stimulating Factor Attenuates Delayed TPA-Induced Hemorrhagic Transformation in Ischemic Stroke Rats by Enhancing Angiogenesis and Vasculogenesis. J. Cereb. Blood Flow Metab. 2015, 35, 338–346. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D.; Tamma, R.; Ruggieri, S.; Annese, T.; Crivellato, E. Surface Markers: An Identity Card of Endothelial Cells. Microcirculation 2020, 27, e12587. [Google Scholar] [CrossRef] [PubMed]
- Risau, W.; Flamme, I. Vasculogenesis. Annu. Rev. Cell. Dev. Biol. 1995, 11, 73–91. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, T.; Kudo, M. Signaling Pathways Governing Tumor Angiogenesis. Oncology 2011, 81, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Vailhé, B.; Vittet, D.; Feige, J.J. In Vitro Models of Vasculogenesis and Angiogenesis. Lab. Investig. 2001, 81, 439–452. [Google Scholar] [CrossRef]
- Wei, S.T.; Huang, Y.C.; Hsieh, M.L.; Lin, Y.J.; Shyu, W.C.; Chen, H.C.; Hsieh, C.H. Atypical Chemokine Receptor ACKR3/CXCR7 Controls Postnatal Vasculogenesis and Arterial Specification by Mesenchymal Stem Cells via Notch Signaling. Cell Death Dis. 2020, 11, 307. [Google Scholar] [CrossRef]
- Garcia, M.D.; Larina, I.V. Vascular Development and Hemodynamic Force in the Mouse Yolk Sac. Front. Physiol. 2014, 5, 308. [Google Scholar] [CrossRef]
- Beedie, S.L.; Diamond, A.J.; Fraga, L.R.; Figg, W.D.; Vargesson, N. Vertebrate Embryos as Tools for Anti-Angiogenic Drug Screening and Function. Reprod. Toxicol. 2017, 70, 49–59. [Google Scholar] [CrossRef]
- Mozes, G.; Gloviczki, P. Venous Embryology and Anatomy. Vein Book 2007, 15–25. [Google Scholar] [CrossRef]
- Wolf, K.; Hu, H.; Isaji, T.; Dardik, A. Molecular Identity of Arteries, Veins, and Lymphatics. J. Vasc. Surg. 2019, 69, 253–262. [Google Scholar] [CrossRef]
- Alitalo, K.; Tammela, T.; Petrova, T.V. Lymphangiogenesis in Development and Human Disease. Nature 2005, 438, 946–953. [Google Scholar] [CrossRef] [PubMed]
- Bruyère, F.; Noël, A. Lymphangiogenesis: In Vitro and in Vivo Models. FASEB J. 2010, 24, 8–21. [Google Scholar] [CrossRef] [PubMed]
- Escobedo, N.; Oliver, G. Lymphangiogenesis: Origin, Specification, and Cell Fate Determination. Annu. Rev. Cell Dev. Biol. 2016, 32, 677–691. [Google Scholar] [CrossRef] [PubMed]
- Kaipainen, A.; Bielenberg, D.R. Hemangiogenesis versus Lymphangiogenesis. Encycl. Eye 2010, 227–232. [Google Scholar] [CrossRef]
- Norrmén, C.; Tammela, T.; Petrova, T.V.; Alitalo, K. Biological Basis of Therapeutic Lymphangiogenesis. Circulation 2011, 123, 1335–1351. [Google Scholar] [CrossRef] [PubMed]
- Tammela, T.; Alitalo, K. Lymphangiogenesis: Molecular Mechanisms and Future Promise. Cell 2010, 140, 460–476. [Google Scholar] [CrossRef] [PubMed]
- Oliver, G. Lymphatic Vasculature Development. Nat. Rev. Immunol. 2004, 4, 35–45. [Google Scholar] [CrossRef]
- Butler, M.G.; Isogai, S.; Weinstein, B.M. Lymphatic Development. Birth Defects Res. Part C Embryo Today Rev. 2009, 87, 222–231. [Google Scholar] [CrossRef]
- Matanes, E.; Gotlieb, W.H. Pathophysiological and Anatomical Basis of Lymphatic Transit of Cancer Cells and Role of the Lymphatic System: A Review of Published Literature. Chin. Clin. Oncol. 2021, 10, 14. [Google Scholar] [CrossRef] [PubMed]
- Leong, S.P.; Pissas, A.; Scarato, M.; Gallon, F.; Pissas, M.H.; Amore, M.; Wu, M.; Faries, M.B.; Lund, A.W. The Lymphatic System and Sentinel Lymph Nodes: Conduit for Cancer Metastasis. Clin. Exp. Metastasis 2022, 39, 139. [Google Scholar] [CrossRef] [PubMed]
- Sakabe, M.; Fan, J.; Odaka, Y.; Liu, N.; Hassan, A.; Duan, X.; Stump, P.; Byerly, L.; Donaldson, M.; Hao, J.; et al. YAP/TAZ-CDC42 Signaling Regulates Vascular Tip Cell Migration. Proc. Natl. Acad. Sci. USA 2017, 114, 10918–10923. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yue, R.; Wei, B.; Gao, G.; Du, J.; Pei, G. Lysophosphatidic Acid Acts as a Nutrient-derived Developmental Cue to Regulate Early Hematopoiesis. EMBO J. 2014, 33, 1383–1396. [Google Scholar] [CrossRef] [PubMed]
- Mylona, A.; Andrieu-Soler, C.; Thongjuea, S.; Martella, A.; Soler, E.; Jorna, R.; Hou, J.; Kockx, C.; Van Ijcken, W.; Lenhard, B.; et al. Genome-Wide Analysis Shows That Ldb1 Controls Essential Hematopoietic Genes/Pathways in Mouse Early Development and Reveals Novel Players in Hematopoiesis. Blood 2013, 121, 2902–2913. [Google Scholar] [CrossRef] [PubMed]
- Meadows, S.M.; Myers, C.T.; Krieg, P.A. Regulation of Endothelial Cell Development by ETS Transcription Factors. Semin. Cell Dev. Biol. 2011, 22, 976–984. [Google Scholar] [CrossRef] [PubMed]
- Komiya, Y.; Habas, R. Wnt Signal Transduction Pathways. Organogenesis 2008, 4, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Blagodatski, A.; Klimenko, A.; Jia, L.; Katanaev, V.L. Small Molecule Wnt Pathway Modulators from Natural Sources: History, State of the Art and Perspectives. Cells 2020, 9, 589. [Google Scholar] [CrossRef] [PubMed]
- Vladar, E.K.; Königshoff, M. Noncanonical Wnt Planar Cell Polarity Signaling in Lung Development and Disease. Biochem. Soc. Trans. 2020, 48, 231–243. [Google Scholar] [CrossRef]
- Lories, R.J.; Monteagudo, S. Review Article: Is Wnt Signaling an Attractive Target for the Treatment of Osteoarthritis? Rheumatol. Ther. 2020, 7, 259–270. [Google Scholar] [CrossRef]
- Tocci, J.M.; Felcher, C.M.; Solá, M.E.G.; Kordon, E.C. R-Spondin-Mediated WNT Signaling Potentiation in Mammary and Breast Cancer Development. IUBMB Life 2020, 72, 1546–1559. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Tao, F.; Zhang, X.; Zhang, Y.; Sun, X.; Wu, D. Role of Wnt/β-Catenin Signaling in the Chemoresistance Modulation of Colorectal Cancer. Biomed. Res. Int. 2020, 2020, 9390878. [Google Scholar] [CrossRef] [PubMed]
- Huybrechts, Y.; Mortier, G.; Boudin, E.; Van Hul, W. WNT Signaling and Bone: Lessons From Skeletal Dysplasias and Disorders. Front. Endocrinol. 2020, 11, 165. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Castañeda, J.R.; Rodelo-Haad, C.; Pendon-Ruiz de Mier, M.V.; Martin-Malo, A.; Santamaria, R.; Rodriguez, M. Klotho/FGF23 and Wnt Signaling as Important Players in the Comorbidities Associated with Chronic Kidney Disease. Toxins 2020, 12, 185. [Google Scholar] [CrossRef] [PubMed]
- Noelanders, R.; Vleminckx, K. How Wnt Signaling Builds the Brain: Bridging Development and Disease. Neuroscientist 2017, 23, 314–329. [Google Scholar] [CrossRef] [PubMed]
- Bem, J.; Brożko, N.; Chakraborty, C.; Lipiec, M.A.; Koziński, K.; Nagalski, A.; Szewczyk, Ł.M.; Wiśniewska, M.B. Wnt/β-Catenin Signaling in Brain Development and Mental Disorders: Keeping TCF7L2 in Mind. FEBS Lett. 2019, 593, 1654–1674. [Google Scholar] [CrossRef] [PubMed]
- Courtwright, A.; Siamakpour-Reihani, S.; Arbiser, J.L.; Banet, N.; Hilliard, E.; Fried, L.; Livasy, C.; Ketelsen, D.; Nepal, D.B.; Perou, C.M.; et al. SFRP2 Stimulates Angiogenesis via a Calcineurin/NFAT Signaling Pathway. Cancer Res. 2009, 69, 4621. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Liu, X.; Zheng, H.; Zhu, H.; Mai, W.; Huang, X.; Huang, Y. Multiple Roles of SFRP2 in Cardiac Development and Cardiovascular Disease. Int. J. Biol. Sci. 2020, 16, 730–738. [Google Scholar] [CrossRef] [PubMed]
- Hasan, S.S.; Tsaryk, R.; Lange, M.; Wisniewski, L.; Moore, J.C.; Lawson, N.D.; Wojciechowska, K.; Schnittler, H.; Siekmann, A.F. Endothelial Notch Signalling Limits Angiogenesis via Control of Artery Formation. Nat. Cell Biol. 2017, 19, 928–940. [Google Scholar] [CrossRef]
- Blanco, R.; Gerhardt, H. VEGF and Notch in Tip and Stalk Cell Selection. Cold Spring Harb. Perspect. Med. 2012, 3, a006569. [Google Scholar] [CrossRef]
- Vega, R.; Carretero, M.; Travasso, R.D.M.; Bonilla, L.L. Notch Signaling and Taxis Mechanims Regulate Early Stage Angiogenesis: A Mathematical and Computational Model. PLoS Comput. Biol. 2020, 16, e1006919. [Google Scholar] [CrossRef] [PubMed]
- MacK, J.J.; Luisa Iruela-Arispe, M. NOTCH Regulation of the Endothelial Cell Phenotype. Curr. Opin. Hematol. 2018, 25, 212–218. [Google Scholar] [CrossRef]
- Roca, C.; Adams, R.H. Regulation of Vascular Morphogenesis by Notch Signaling. Genes. Dev. 2007, 21, 2511–2524. [Google Scholar] [CrossRef]
- Luo, Z.; Shang, X.; Zhang, H.; Wang, G.; Massey, P.A.; Barton, S.R.; Kevil, C.G.; Dong, Y. Notch Signaling in Osteogenesis, Osteoclastogenesis, and Angiogenesis. Am. J. Pathol. 2019, 189, 1495–1500. [Google Scholar] [CrossRef]
- Andersen, P.; Uosaki, H.; Shenje, L.T.; Kwon, C. Non-Canonical Notch Signaling: Emerging Role and Mechanism. Trends Cell Biol. 2012, 22, 257–265. [Google Scholar] [CrossRef]
- Herrick, D.B.; Guo, Z.; Jang, W.; Schnittke, N.; Schwob, J.E. Canonical Notch Signaling Directs the Fate of Differentiating Neurocompetent Progenitors in the Mammalian Olfactory Epithelium. J. Neurosci. 2018, 38, 5022–5037. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, B.; Meloty-Kapella, L.; Weinmaster, G. Canonical and Non-Canonical Notch Ligands. Curr. Top. Dev. Biol. 2010, 92, 73–129. [Google Scholar] [CrossRef]
- Beets, K.; Staring, M.W.; Criem, N.; Maas, E.; Schellinx, N.; De Sousa Lopes, S.M.C.; Umans, L.; Zwijsen, A. BMP-SMAD Signalling Output Is Highly Regionalized in Cardiovascular and Lymphatic Endothelial Networks. BMC Dev. Biol. 2016, 16, 34. [Google Scholar] [CrossRef] [PubMed]
- Jakobsson, L.; van Meeteren, L.A. Transforming Growth Factor β Family Members in Regulation of Vascular Function: In the Light of Vascular Conditional Knockouts. Exp. Cell Res. 2013, 319, 1264–1270. [Google Scholar] [CrossRef]
- Herrera, B.; Addante, A.; Sánchez, A. BMP Signalling at the Crossroad of Liver Fibrosis and Regeneration. Int. J. Mol. Sci. 2018, 19, 39. [Google Scholar] [CrossRef]
- Dyer, L.; Patterson, C. Development of the Endothelium: An Emphasis on Heterogeneity. Semin. Thromb. Hemost. 2010, 36, 227–235. [Google Scholar] [CrossRef][Green Version]
- Marks-Bluth, J.; Khanna, A.; Chandrakanthan, V.; Thoms, J.; Bee, T.; Eich, C.; Kang, Y.C.; Knezevic, K.; Qiao, Q.; Fitch, S.; et al. SMAD1 and SMAD5 Expression Is Coordinately Regulated by FLI1 and GATA2 during Endothelial Development. Mol. Cell. Biol. 2015, 35, 2165–2172. [Google Scholar] [CrossRef]
- Dyer, L.A.; Pi, X.; Patterson, C. The Role of BMPs in Endothelial Cell Function and Dysfunction. Trends Endocrinol. Metab. 2014, 25, 472–480. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.N.; Green, J.; Wang, Z.; Deng, Y.; Qiao, M.; Peabody, M.; Zhang, Q.; Ye, J.; Yan, Z.; Denduluri, S.; et al. Bone Morphogenetic Protein (BMP) Signaling in Development and Human Diseases. Genes Dis. 2014, 1, 87–105. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Bao, L.; Zhao, M.; Cao, J.; Zheng, H. Progress in Research on the Role of FGF in the Formation and Treatment of Corneal Neovascularization. Front. Pharmacol. 2020, 11, 111. [Google Scholar] [CrossRef]
- Oladipupo, S.S.; Smith, C.; Santeford, A.; Park, C.; Sene, A.; Wiley, L.A.; Osei-Owusu, P.; Hsu, J.; Zapata, N.; Liu, F.; et al. Endothelial Cell FGF Signaling Is Required for Injury Response but Not for Vascular Homeostasis. Proc. Natl. Acad. Sci. USA 2014, 111, 13379–13384. [Google Scholar] [CrossRef] [PubMed]
- Seghezzi, G.; Patel, S.; Ren, C.J.; Gualandris, A.; Pintucci, G.; Robbins, E.S.; Shapiro, R.L.; Galloway, A.C.; Rifkin, D.B.; Mignatti, P. Fibroblast Growth Factor-2 (FGF-2) Induces Vascular Endothelial Growth Factor (VEGF) Expression in the Endothelial Cells of Forming Capillaries: An Autocrine Mechanism Contributing to Angiogenesis. J. Cell Biol. 1998, 141, 1659–1673. [Google Scholar] [CrossRef]
- Garry, D.J. Etv2 Is a Master Regulator of Hematoendothelial Lineages. Trans. Am. Clin. Climatol. Assoc. 2016, 127, 212–223. [Google Scholar]
- Koyano-Nakagawa, N.; Kweon, J.; Iacovino, M.; Shi, X.; Rasmussen, T.L.; Borges, L.; Zirbes, K.M.; Li, T.; Perlingeiro, R.C.R.; Kyba, M.; et al. Etv2 Is Expressed in the Yolk Sac Hematopoietic and Endothelial Progenitors and Regulates Lmo2 Gene Expression. Stem Cells 2012, 30, 1611–1623. [Google Scholar] [CrossRef]
- Dobrzycki, T.; Lalwani, M.; Telfer, C.; Monteiro, R.; Patient, R. The Roles and Controls of GATA Factors in Blood and Cardiac Development. IUBMB Life 2020, 72, 39–44. [Google Scholar] [CrossRef]
- Whitcomb, J.; Gharibeh, L.; Nemer, M. From Embryogenesis to Adulthood: Critical Role for GATA Factors in Heart Development and Function. IUBMB Life 2020, 72, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Yan, X.; Sun, X.; Shen, X.; Yin, H.; Wang, C.; Liu, Y.; Lu, C.; Fu, H.; Yang, S.; et al. Synergistic Effects of Dual-Presenting VEGF- A Nd BDNF-Mimetic Peptide Epitopes from Self-Assembling Peptide Hydrogels on Peripheral Nerve Regeneration. Nanoscale 2019, 11, 19943–19958. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.K.; Chauhan, S.K.; Kay, E.D.; Dana, R. Flt-1 Regulates Vascular Endothelial Cell Migration via a Protein Tyrosine Kinase-7–Dependent Pathway. Blood 2011, 117, 5762. [Google Scholar] [CrossRef] [PubMed]
- Volpe, M.C.; Ciucci, G.; Zandomenego, G.; Vuerich, R.; Ring, N.A.R.; Vodret, S.; Salton, F.; Marchesan, P.; Braga, L.; Marcuzzo, T.; et al. Flt1 Produced by Lung Endothelial Cells Impairs ATII Cell Transdifferentiation and Repair in Pulmonary Fibrosis. Cell Death Dise. 2023, 14, 437. [Google Scholar] [CrossRef] [PubMed]
- Holmes, K.; Roberts, O.L.; Thomas, A.M.; Cross, M.J. Vascular Endothelial Growth Factor Receptor-2: Structure, Function, Intracellular Signalling and Therapeutic Inhibition. Cell. Signal. 2007, 19, 2003–2012. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Xu, J.; Wei, Q.; Zhao, Z.; Chen, X.; Cui, H.; Zhang, Y. VEGF/Flk1 Mechanism Is Involved in Roxarsone Promotion of Rat Endothelial Cell Growth and B16F10 Xenograft Tumor Angiogenesis. Sci. Rep. 2019, 9, 17417. [Google Scholar] [CrossRef] [PubMed]
- Morris, B.J.; Chen, R.; Donlon, T.A.; Kallianpur, K.J.; Masaki, K.H.; Willcox, B.J. Vascular Endothelial Growth Factor Receptor 1 Gene (FLT1) Longevity Variant Increases Lifespan by Reducing Mortality Risk Posed by Hypertension. Aging 2023, 15, 3967–3983. [Google Scholar] [CrossRef]
- Otowa, Y.; Moriwaki, K.; Sano, K.; Shirakabe, M.; Yonemura, S.; Shibuya, M.; Rossant, J.; Suda, T.; Kakeji, Y.; Hirashima, M. Flt1/VEGFR1 Heterozygosity Causes Transient Embryonic Edema. Sci. Rep. 2016, 6, 27186. [Google Scholar] [CrossRef]
- Park-Windhol, C.; D’Amore, P.A. Disorders of Vascular Permeability. Trends Endocrinol. Metab. 2016, 11, 251–281. [Google Scholar] [CrossRef]
- Harde, E.; Nicholson, L.; Furones Cuadrado, B.; Bissen, D.; Wigge, S.; Urban, S.; Segarra, M.; Ruiz de Almodóvar, C.; Acker-Palmer, A. EphrinB2 Regulates VEGFR2 during Dendritogenesis and Hippocampal Circuitry Development. Elife 2019, 8, e49819. [Google Scholar] [CrossRef]
- Large, C.L.; Vitali, H.E.; Whatley, J.D.; Red-Horse, K.; Sharma, B. In Vitro Model of Coronary Angiogenesis. J. Vis. Exp. 2020, 2020, e60558. [Google Scholar] [CrossRef]
- Real, P.J.; Ligero, G.; Ayllon, V.; Ramos-Mejia, V.; Bueno, C.; Gutierrez-Aranda, I.; Navarro-Montero, O.; Lako, M.; Menendez, P. SCL/TAL1 Regulates Hematopoietic Specification from Human Embryonic Stem Cells. Mol. Ther. 2012, 20, 1443–1453. [Google Scholar] [CrossRef] [PubMed]
- Hoang, T.; Lambert, J.A.; Martin, R. SCL/TAL1 in Hematopoiesis and Cellular Reprogramming, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2016; Volume 118. [Google Scholar]
- Porcher, C.; Chagraoui, H.; Kristiansen, M.S. SCL/TAL1: A Multifaceted Regulator from Blood Development to Disease. Blood 2017, 129, 2051–2060. [Google Scholar] [CrossRef] [PubMed]
- Matrone, G.; Meng, S.; Gu, Q.; Lv, J.; Fang, L.; Chen, K.; Cooke, J.P. Lmo2 Modulates Sphk1 and Promotes Endothelial Cell Migration. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 1860–1868. [Google Scholar] [CrossRef] [PubMed]
- Ganta, V.C.; Annex, B.H. LMO2 and Endothelial Cell Migration in Developmental and Postnatal Angiogenesis. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 1806–1808. [Google Scholar] [CrossRef] [PubMed]
- Cañete, A.; Cano, E.; Muñoz-Chápuli, R.; Carmona, R. Role of Vitamin a/Retinoic Acid in Regulation of Embryonic and Adult Hematopoiesis. Nutrients 2017, 9, 159. [Google Scholar] [CrossRef] [PubMed]
- Saito, A.; Sugawara, A.; Uruno, A.; Kudo, M.; Kagechika, H.; Sato, Y.; Owada, Y.; Kondo, H.; Sato, M.; Kurabayashi, M.; et al. All-Trans Retinoic Acid Induces in Vitro Angiogenesis via Retinoic Acid Receptor: Possible Involvement of Paracrine Effects of Endogenous Vascular Endothelial Growth Factor Signaling. Endocrinology 2007, 148, 1412–1423. [Google Scholar] [CrossRef] [PubMed]
- Lai, L.; Bohnsack, B.L.; Niederreither, K.; Hirschi, K.K. Retinoic Acid Regulates Endothelial Cell Proliferation during Vasculogenesis. Development 2003, 130, 6465–6474. [Google Scholar] [CrossRef]
- Pawlikowski, B.; Wragge, J.; Siegenthaler, J.A. Retinoic Acid Signaling in Vascular Development. Genesis 2019, 57, e23287. [Google Scholar] [CrossRef] [PubMed]
- Binshtok, U.; Sprinzak, D. Molecular Mechanisms of Notch Signaling; Springer International Publishing: Cham, Switzerland, 2018; Volume 1066, ISBN 978-3-319-89511-6. [Google Scholar]
- Cunha, S.I.; Magnusson, P.U.; Dejana, E.; Lampugnani, M.G. Deregulated TGF-β/BMP Signaling in Vascular Malformations. Circ. Res. 2017, 121, 981–999. [Google Scholar] [CrossRef]
- Garside, V.C.; Chang, A.C.; Karsan, A.; Hoodless, P.A. Co-Ordinating Notch, BMP, and TGF-β Signaling during Heart Valve Development. Cell. Mol. Life Sci. 2013, 70, 2899–2917. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Wang, X.F. Signaling Cross-Talk between TGF-β/BMP and Other Pathways. Cell Res. 2009, 19, 71–88. [Google Scholar] [CrossRef] [PubMed]
- Holderfield, M.T.; Hughes, C.C.W. Cross-talk between Vascular Endothelial Growth Factor, Notch, and Transforming Growth Factor-β in Vascular Morphogenesis. Circ. Res. 2008, 102, 637–652. [Google Scholar] [CrossRef] [PubMed]
- LaFoya, B.; Munroe, J.A.; Mia, M.M.; Detweiler, M.A.; Crow, J.J.; Wood, T.; Roth, S.; Sharma, B.; Albig, A.R. Notch: A Multi-Functional Integrating System of Microenvironmental Signals. Dev. Biol. 2016, 418, 227–241. [Google Scholar] [CrossRef]
- Lyle, C.L.; Belghasem, M.; Chitalia, V.C. C-Cbl: An Important Regulator and a Target in Angiogenesis and Tumorigenesis. Cells 2019, 8, 498. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, R.; Pinheiro, P.; Joseph, N.; Peterkin, T.; Koth, J.; Repapi, E.; Bonkhofer, F.; Kirmizitas, A.; Patient, R. Transforming Growth Factor β Drives Hemogenic Endothelium Programming and the Transition to Hematopoietic Stem Cells. Dev. Cell 2016, 38, 358–370. [Google Scholar] [CrossRef] [PubMed]
- Olsen, J.J.; Pohl, S.Ö.; Deshmukh, A.; Visweswaran, M.; Ward, N.C.; Arfuso, F.; Agostino, M.; Dharmarajan, A. The Role of Wnt Signalling in Angiogenesis. Clin. Biochem. Rev. 2017, 38, 131–142. [Google Scholar] [PubMed]
- Sam, S.A.; Teel, J.; Tegge, A.N.; Bharadwaj, A.; Murali, T.M. XTALKDB: A Database of Signaling Pathway Cross-talk. Nucleic Acids Res. 2017, 45, D432–D439. [Google Scholar] [CrossRef] [PubMed]
- Takebe, N.; Percy Ivy, S. Controversies in Cancer Stem Cells: Targeting Embryonic Signaling Pathways. Clin. Cancer Res. 2010, 16, 3106–3112. [Google Scholar] [CrossRef]
- Thurston, G.; Kitajewski, J. VEGF and Delta-Notch: Interacting Signalling Pathways in Tumour Angiogenesis. Br. J. Cancer 2008, 99, 1204–1209. [Google Scholar] [CrossRef]
- Wang, Y.; Shen, R.-W.; Han, B.; Li, Z.; Xiong, L.; Zhang, F.-Y.; Cong, B.-B.; Zhang, B. Notch Signaling Mediated by TGF-β/Smad Pathway in Concanavalin A-Induced Liver Fibrosis in Rats. World J. Gastroenterol. 2017, 23, 2330. [Google Scholar] [CrossRef] [PubMed]
- Itoh, F.; Itoh, S.; Goumans, M.J.; Valdimarsdottir, G.; Iso, T.; Dotto, G.P.; Hamamori, Y.; Kedes, L.; Kato, M.; Ten Dijke, P. Synergy and Antagonism between Notch and BMP Receptor Signaling Pathways in Endothelial Cells. EMBO J. 2004, 23, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Yao, J.; Wang, L.; Zhang, D.; Zhang, L.; Reynolds, E.X.; Yu, T.; Boström, K.I.; Yao, Y. Cross-talk between BMP and Notch Induces Sox2 in Cerebral Endothelial Cells. Cells 2019, 8, 549. [Google Scholar] [CrossRef]
- Benn, A.; Hiepen, C.; Osterland, M.; Schütte, C.; Zwijsen, A.; Knaus, P. Role of Bone Morphogenetic Proteins in Sprouting Angiogenesis: Differential BMP Receptor-Dependent Signaling Pathways Balance Stalk vs. Tip Cell Competence. FASEB J. 2017, 31, 4720–4733. [Google Scholar] [CrossRef]
- Caliceti, C.; Nigro, P.; Rizzo, P.; Ferrari, R. ROS, Notch, and Wnt Signaling Pathways: Cross-talk between Three Major Regulators of Cardiovascular Biology. Biomed Res. Int. 2014, 2014, 318714. [Google Scholar] [CrossRef] [PubMed]
- Foulquier, S.; Daskalopoulos, E.P.; Lluri, G.; Hermans, K.C.M.; Deb, A.; Blankesteijn, W.M. WNT Signaling in Cardiac and Vascular Disease. Pharmacol. Rev. 2018, 70, 68–141. [Google Scholar] [CrossRef] [PubMed]
- Levenberg, S.; Zoldan, J.; Basevitch, Y.; Langer, R. Endothelial Potential of Human Embryonic Stem Cells. Blood 2007, 110, 806–814. [Google Scholar] [CrossRef] [PubMed]
- Banno, K.; Yoder, M.C. Endothelial Stem and Progenitor Cells for Regenerative Medicine. Curr. Stem Cell Rep. 2019, 5, 101–108. [Google Scholar] [CrossRef]
- Wei, Q.; Frenette, P.S. Niches for Hematopoietic Stem Cells and Their Progeny. Immunity 2018, 48, 632–648. [Google Scholar] [CrossRef]
- Gussin, H.A.E.; Bischoff, F.Z.; Hoffman, R.; Elias, S. Endothelial Precursor Cells in the Peripheral Blood of Pregnant Women. J. Soc. Gynecol. Investig. 2002, 9, 357–361. [Google Scholar] [CrossRef]
- Urbich, C.; Dimmeler, S. Endothelial Progenitor Cells: Characterization and Role in Vascular Biology. Circ. Res. 2004, 95, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Rafii, S. Circulating Endothelial Precursors: Mystery, Reality, and Promise. J. Clin. Investig. 2000, 105, 17–19. [Google Scholar] [CrossRef]
- Szpera-Goździewicz, A.; Majcherek, M.; Boruczkowski, M.; Goździewicz, T.; Dworacki, G.; Wicherek, L.; Bręborowicz, G.H. Circulating Endothelial Cells, Circulating Endothelial Progenitor Cells, and von Willebrand Factor in Pregnancies Complicated by Hypertensive Disorders. Am. J. Reprod. Immunol. 2017, 77, e12625. [Google Scholar] [CrossRef] [PubMed]
- Zahran, A.M.; Mohamed, I.L.; El Asheer, O.M.; Tamer, D.M.; Abo-ELela, M.G.M.; Abdel-Rahim, M.H.; El-Badawy, O.H.B.; Elsayh, K.I. Circulating Endothelial Cells, Circulating Endothelial Progenitor Cells, and Circulating Microparticles in Type 1 Diabetes Mellitus. Clin. Appl. Thromb. 2019, 25, 1076029618825311. [Google Scholar] [CrossRef] [PubMed]
- Plein, A.; Fantin, A.; Denti, L.; Pollard, J.W.; Ruhrberg, C. Erythro-Myeloid Progenitors Contribute Endothelial Cells to Blood Vessels. Nature 2018, 562, 223–228. [Google Scholar] [CrossRef]
- Díaz del Moral, S.; Barrena, S.; Muñoz-Chápuli, R.; Carmona, R. Embryonic Circulating Endothelial Progenitor Cells. Angiogenesis 2020, 23, 531–541. [Google Scholar] [CrossRef]
- Nieda, M.; Nicol, A.; Denning-Kendall, P.; Sweetenham, J.; Bradley, B.; Hows, J. Endothelial Cell Precursors Are Normal Components of Human Umbilical Cord Blood. Br. J. Haematol. 1997, 98, 775–777. [Google Scholar] [CrossRef]
- Kirton, J.P.; Xu, Q. Endothelial Precursors in Vascular Repair. Microvasc. Res. 2010, 79, 193–199. [Google Scholar] [CrossRef]
- Asahara, T.; Kawamoto, A.; Masuda, H. Concise Review: Circulating Endothelial Progenitor Cells for Vascular Medicine. Stem Cells 2011, 29, 1650–1655. [Google Scholar] [CrossRef]
- Heissig, B.; Hattori, K.; Dias, S.; Friedrich, M.; Ferris, B.; Hackett, N.R.; Crystal, R.G.; Besmer, P.; Lyden, D.; Moore, M.A.S.; et al. Recruitment of Stem and Progenitor Cells from the Bone Marrow Niche Requires MMP-9 Mediated Release of Kit-Ligand. Cell 2002, 109, 625–637. [Google Scholar] [CrossRef]
- Shiozawa, Y.; Havens, A.M.; Pienta, K.J.; Taichman, R.S. The Bone Marrow Niche: Habitat to Hematopoietic and Mesenchymal Stem Cells, and Unwitting Host to Molecular Parasites. Leukemia 2008, 22, 941–950. [Google Scholar] [CrossRef] [PubMed]
- Sui, B.D.; Zheng, C.X.; Li, M.; Jin, Y.; Hu, C.H. Epigenetic Regulation of Mesenchymal Stem Cell Homeostasis. Trends Cell Biol. 2020, 30, 97–116. [Google Scholar] [CrossRef]
- Fielding, C.; Méndez-Ferrer, S. Neuronal Regulation of Bone Marrow Stem Cell Niches. F1000Res 2020, 9, 614. [Google Scholar] [CrossRef] [PubMed]
- Shapira, S.N.; Christofk, H.R. Metabolic Regulation of Tissue Stem Cells. Trends Cell Biol. 2020, 30, 566–576. [Google Scholar] [CrossRef] [PubMed]
- Jagannathan-Bogdan, M.; Zon, L.I. Hematopoiesis. Development 2013, 140, 2463–2467. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Li, G.; Deng, L.; Kuang, B.; Li, X. The Roles of FGF10 in Vasculogenesis and Angiogenesis. Biomed. Res. 2017, 28, 1329–1332. [Google Scholar]
- Ribatti, D.; Vacca, A.; De Falco, G.; Ria, R.; Roncali, L.; Dammacco, F. Role of Hematopoietic Growth Factors in Angiogenesis. Acta Haematol. 2001, 106, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Goldie, L.C.; Nix, M.K.; Hirschi, K.K. Embryonic Vasculogenesis and Hematopoietic Specification. Organogenesis 2008, 4, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Kauts, M.L.; Vink, C.S.; Dzierzak, E. Hematopoietic (Stem) Cell Development—How Divergent Are the Roads Taken? FEBS Lett. 2016, 590, 3975–3986. [Google Scholar] [CrossRef]
- Krenning, G.; Barauna, V.G.; Krieger, J.E.; Harmsen, M.C.; Moonen, J.R.A.J. Endothelial Plasticity: Shifting Phenotypes through Force Feedback. Stem Cells Int. 2016, 2016, 9762959. [Google Scholar] [CrossRef]
- Caolo, V.; Molin, D.G.M.; Post, M.J. Notch Regulation of Hematopoiesis, Endothelial Precursor Cells, and Blood Vessel Formation: Orchestrating the Vasculature. Stem Cells Int. 2012, 2012, 805602. [Google Scholar] [CrossRef] [PubMed]
- Testa, U.; Castelli, G.; Pelosi, E. Role of Endothelial Progenitor Cells in Vascular Development, Homestatic Maintenance of Blood Vessels and in Injury-Mediated Reparative Response. Stem Cell Investig. 2020, 7, 7. [Google Scholar] [CrossRef]
- Kim, P.G.; Albacker, C.E.; Lu, Y.F.; Jang, I.H.; Lim, Y.; Heffner, G.C.; Arora, N.; Bowman, T.V.; Lin, M.I.; Lensch, M.W.; et al. Signaling Axis Involving Hedgehog, Notch, and Scl Promotes the Embryonic Endothelial-to-Hematopoietic Transition. Proc. Natl. Acad. Sci. USA 2013, 110, E141–E150. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D.; Vacca, A.; Roncali, L.; Dammacco, F. Hematopoiesis and Angiogenesis: A Link between Two Apparently Independent Processes. J. Hematother. Stem Cell Res. 2000, 19, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Hattori, K.; Dias, S.; Heissig, B.; Hackett, N.R.; Lyden, D.; Tateno, M.; Hicklin, D.J.; Zhu, Z.; Witte, L.; Crystal, R.G.; et al. Vascular Endothelial Growth Factor and Angiopoietin-1 Stimulate Postnatal Hematopoiesis by Recruitment of Vasculogenic and Hematopoietic Stem Cells. J. Exp. Med. 2001, 193, 1005–1014. [Google Scholar] [CrossRef]
- D’Alessio, A.; Moccia, F.; Li, J.H.; Micera, A.; Kyriakides, T.R. Angiogenesis and Vasculogenesis in Health and Disease. Biomed. Res. Int. 2015, 2015, 2–4. [Google Scholar] [CrossRef]
- Heissig, B.; Werb, Z.; Rafii, S.; Hattori, K. Role of C-Kit/Kit Ligand Signaling in Regulating Vasculogenesis. Thromb. Haemost. 2003, 90, 570–576. [Google Scholar] [CrossRef] [PubMed]
- Ueda, T.; Yokota, T.; Okuzaki, D.; Uno, Y.; Mashimo, T.; Kubota, Y.; Sudo, T.; Ishibashi, T.; Shingai, Y.; Doi, Y.; et al. Endothelial Cell-Selective Adhesion Molecule Contributes to the Development of Definitive Hematopoiesis in the Fetal Liver. Stem Cell Rep. 2019, 13, 992–1005. [Google Scholar] [CrossRef]
- Cangara, H.M.; Ishida, T.; Hara, T.; Sun, L.; Toh, R.; Rikitake, Y.; Kundu, R.K.; Quertermous, T.; Hirata, K.I.; Hayashi, Y. Role of Endothelial Cell-Selective Adhesion Molecule in Hematogeneous Metastasis. Microvasc. Res. 2010, 80, 133–141. [Google Scholar] [CrossRef]
- Inoue, M.; Ishida, T.; Yasuda, T.; Toh, R.; Hara, T.; Cangara, H.M.; Rikitake, Y.; Taira, K.; Sun, L.; Kundu, R.K.; et al. Endothelial Cell-Selective Adhesion Molecule Modulates Atherosclerosis through Plaque Angiogenesis and Monocyte-Endothelial Interaction. Microvasc. Res. 2010, 80, 179–187. [Google Scholar] [CrossRef]
- Cahill, P.A.; Redmond, E.M. Vascular Endothelium-Gatekeeper of Vessel Health. Atherosclerosis 2016, 248, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Nova-Lampeti, E.; Aguilera, V.; Oporto, K.; Guzmán, P.; Ormazábal, V.; Zúñiga, F.; Escudero, C.; Aguayo, C. Hox Genes in Adult Tissues and Their Role in Endothelial Cell Differentiation and Angiogenesis. Endothel. Dysfunct. Old Concept. New Chall. 2018, 10. [Google Scholar] [CrossRef]
- Minami, T.; Aird, W.C. Endothelial Cell Gene Regulation. Trends Cardiovasc. Med. 2005, 15, 174.e1–174.e24. [Google Scholar] [CrossRef] [PubMed]
- Gaskell, T.; Englund, M.C.O.; Hyllner, J. Human Embryonic Stem Cells. Regenerative Medicine-from Protocol to Patient: 2. Stem Cell Science and Technology; Springer: Berlin/Heidelberg, Germany, 2016; pp. 27–49. [Google Scholar] [CrossRef]
- Lacorre, D.-A.; Baekkevold, E.S.; Garrido, I.; Brandtzaeg, P.; Haraldsen, G.; Amalric, F.; Girard, J.-P. Plasticity of Endothelial Cells: Rapid Dedifferentiation of Freshly Isolated High Endothelial Venule Endothelial Cells Outside the Lymphoid Tissue Microenvironment. Blood 2004, 103, 4164–4172. [Google Scholar] [CrossRef] [PubMed]
- Cleuren, A.C.A.A.; van der Ent, M.A.; Jiang, H.; Hunker, K.L.; Yee, A.; Siemieniak, D.R.; Molema, G.; Aird, W.C.; Ganesh, S.K.; Ginsburg, D. The in Vivo Endothelial Cell Translatome Is Highly Heterogeneous across Vascular Beds. Proc. Natl. Acad. Sci. USA 2019, 116, 23618–23624. [Google Scholar] [CrossRef]
- Russell-Hallinan, A.; Watson, C.J.; O’Dwyer, D.; Grieve, D.J.; O’Neill, K.M. Epigenetic Regulation of Endothelial Cell Function by Nucleic Acid Methylation in Cardiac Homeostasis and Disease. Cardiovasc. Drugs Ther. 2021, 35, 1025. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Huang, R.T.; Hamanaka, R.B.; Krause, M.; Oh, M.J.; Kuo, C.H.; Nigdelioglu, R.; Meliton, A.Y.; Witt, L.; Dai, G.; et al. HIF-1α Is Required for Disturbed Flow-Induced Metabolic Reprogramming in Human and Porcine Vascular Endothelium. Elife 2017, 6, e25217. [Google Scholar] [CrossRef] [PubMed]
- Van Pham, P.; Vu, N.B.; Nguyen, H.T.; Huynh, O.T.; Truong, M.T.H. Significant Improvement of Direct Reprogramming Efficacy of Fibroblasts into Progenitor Endothelial Cells by ETV2 and Hypoxia. Stem Cell Res. Ther. 2016, 7, 104. [Google Scholar] [CrossRef] [PubMed]
- Lucero, R.; Zappulli, V.; Sammarco, A.; Murillo, O.D.; Cheah, P.S.; Srinivasan, S.; Tai, E.; Ting, D.T.; Wei, Z.; Roth, M.E.; et al. Glioma-Derived MiRNA-Containing Extracellular Vesicles Induce Angiogenesis by Reprogramming Brain Endothelial Cells. Cell Rep. 2020, 30, 2065–2074.e4. [Google Scholar] [CrossRef]
- Hansen, A.; Henderson, S.; Lagos, D.; Nikitenko, L.; Coulter, E.; Roberts, S.; Gratrix, F.; Plaisance, K.; Renne, R.; Bower, M.; et al. KSHV-Encoded MiRNAs Target MAF to Induce Endothelial Cell Reprogramming. Genes. Dev. 2010, 24, 195–205. [Google Scholar] [CrossRef]
- Wang, L.; Xiang, M.; Liu, Y.; Sun, N.; Lu, M.; Shi, Y.; Wang, X.; Meng, D.; Chen, S.; Qin, J. Human Induced Pluripotent Stem Cells Derived Endothelial Cells Mimicking Vascular Inflammatory Response under Flow. Biomicrofluidics 2016, 10, 014106. [Google Scholar] [CrossRef] [PubMed]
- Han, J.-K.; Chang, S.-H.; Cho, H.-J.; Choi, S.-B.; Ahn, H.-S.; Lee, J.; Jeong, H.; Youn, S.-W.; Lee, H.-J.; Kwon, Y.-W.; et al. Direct Conversion of Adult Skin Fibroblasts to Endothelial Cells by Defined Factors. Circulation 2014, 130, 1168–1178. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Park, C.; Han, J.W.; Kim, J.Y.; Cho, K.; Kim, E.J.; Kim, S.; Lee, S.-J.; Oh, S.Y.; Tanaka, Y.; et al. Direct Reprogramming of Human Dermal Fibroblasts Into Endothelial Cells Using ER71/ETV2. Circ. Res. 2017, 120, 848–861. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, K.; Das, A. TWIST1 -Reprogrammed Endothelial Cell Transplantation Potentiates Neovascularization-Mediated Diabetic Wound Tissue Regeneration. Diabetes 2020, 69, 1232–1247. [Google Scholar] [CrossRef] [PubMed]
- Souilhol, C.; Serbanovic-Canic, J.; Fragiadaki, M.; Chico, T.J.; Ridger, V.; Roddie, H.; Evans, P.C. Endothelial Responses to Shear Stress in Atherosclerosis: A Novel Role for Developmental Genes. Nat. Rev. Cardiol. 2019, 17, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Lobov, I.; Mikhailova, N. The Role of Dll4/Notch Signaling in Normal and Pathological Ocular Angiogenesis: Dll4 Controls Blood Vessel Sprouting and Vessel Remodeling in Normal and Pathological Conditions. J. Ophthalmol. 2018, 2018, 3565292. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Chen, F.; Clifton, N.; Sullivan, D.M.; Dalton, W.S.; Gabrilovich, D.I.; Nefedova, Y. Combined Inhibition of Notch Signaling and Bcl-2/Bcl-XL Results in Synergistic Anti-Myeloma Effect. Mol. Cancer Ther. 2010, 9, 3200. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, Y.; Banerjee, S.; Sarkar, F.H. Exploitation of the Notch Signaling Pathway as a Novel Target for Cancer Therapy. Anticancer Res. 2008, 28, 3621–3630. [Google Scholar] [PubMed]
- Yang, X.; Klein, R.; Tian, X.; Cheng, H.T.; Kopan, R.; Shen, J. Notch Activation Induces Apoptosis in Neural Progenitor Cells through a P53-Dependent Pathway. Dev. Biol. 2004, 269, 81–94. [Google Scholar] [CrossRef]
- Segarra, M.; Williams, C.K.; De La Luz Sierra, M.; Bernardo, M.; McCormick, P.J.; Maric, D.; Regino, C.; Choyke, P.; Tosato, G. Dll4 Activation of Notch Signaling Reduces Tumor Vascularity and Inhibits Tumor Growth. Blood 2008, 112, 1904. [Google Scholar] [CrossRef]
- Sajinovic, T.; Baier, G. New Insights into the Diverse Functions of the NR2F Nuclear Orphan Receptor Family. Front. Biosci. 2023, 28, 13. [Google Scholar] [CrossRef] [PubMed]
- Manukjan, N.; Ahmed, Z.; Fulton, D.; Blankesteijn, W.M.; Foulquier, S. A Systematic Review of WNT Signaling in Endothelial Cell Oligodendrocyte Interactions: Potential Relevance to Cerebral Small Vessel Disease. Cells 2020, 9, 1545. [Google Scholar] [CrossRef]
- Laksitorini, M.D.; Yathindranath, V.; Xiong, W.; Hombach-Klonisch, S.; Miller, D.W. Modulation of Wnt/β-Catenin Signaling Promotes Blood-Brain Barrier Phenotype in Cultured Brain Endothelial Cells. Sci. Rep. 2019, 9, 19718. [Google Scholar] [CrossRef] [PubMed]
- Corada, M.; Nyqvist, D.; Orsenigo, F.; Caprini, A.; Giampietro, C.; Taketo, M.M.; Iruela-Arispe, M.L.; Adams, R.H.; Dejana, E. The Wnt/β-Catenin Pathway Modulates Vascular Remodeling and Specification by Upregulating Dll4/Notch Signaling. Dev. Cell 2010, 18, 938. [Google Scholar] [CrossRef] [PubMed]
- Korn, C.; Scholz, B.; Hu, J.; Srivastava, K.; Wojtarowicz, J.; Arnsperger, T.; Adams, R.H.; Boutros, M.; Augustin, H.G.; Augustin, I. Endothelial Cell-Derived Non-Canonical Wnt Ligands Control Vascular Pruning in Angiogenesis. Development 2014, 141, 1757–1766. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, J.R.; Fortunato, I.C.; Fonseca, C.G.; Pezzarossa, A.; Barbacena, P.; Dominguez-Cejudo, M.A.; Vasconcelos, F.F.; Santos, N.C.; Carvalho, F.A.; Franco, C.A. Non-Canonical Wnt Signaling Regulates Junctional Mechanocoupling during Angiogenic Collective Cell Migration. Elife 2019, 8, e45853. [Google Scholar] [CrossRef]
- Meyer, I.S.; Jungmann, A.; Dieterich, C.; Zhang, M.; Lasitschka, F.; Werkmeister, S.; Haas, J.; Müller, O.J.; Boutros, M.; Nahrendorf, M.; et al. The Cardiac Microenvironment Uses Non-canonical WNT Signaling to Activate Monocytes after Myocardial Infarction. EMBO Mol. Med. 2017, 9, 1279. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Lu, Y.; Zhang, Y.; Wu, J.; McVicar, A.; Chen, Y.; Zhu, S.; Zhu, G.; Lu, Y.; Zhang, J.; et al. Cbfβ Regulates Wnt/β-Catenin, Hippo/Yap, and TGFβ Signaling Pathways in Articular Cartilage Homeostasis and Protects from ACLT Surgery-Induced Osteoarthritis. Elife 2024, 13, e95640. [Google Scholar] [CrossRef]
- Gastfriend, B.D.; Nishihara, H.; Canfield, S.G.; Foreman, K.L.; Engelhardt, B.; Palecek, S.P.; Shusta, E.V. Wnt Signaling Mediates Acquisition of Blood-Brain Barrier Properties in Naive Endothelium Derived from Human Pluripotent Stem Cells. Elife 2021, 10, e70992. [Google Scholar] [CrossRef]
- Kulikauskas, M.R.; Shaka, X.; Bautch, V.L. The Versatility and Paradox of BMP Signaling in Endothelial Cell Behaviors and Blood Vessel Function. Cell. Mol. Life Sci. 2022, 79, 77. [Google Scholar] [CrossRef]
- Baeyens, N.; Larrivée, B.; Ola, R.; Hayward-Piatkowskyi, B.; Dubrac, A.; Huang, B.; Ross, T.D.; Coon, B.G.; Min, E.; Tsarfati, M.; et al. Defective Fluid Shear Stress Mechanotransduction Mediates Hereditary Hemorrhagic Telangiectasia. J. Cell Biol. 2016, 214, 807–816. [Google Scholar] [CrossRef]
- Han, O.; Pak, B.; Jin, S.W. The Role of BMP Signaling in Endothelial Heterogeneity. Front. Cell Dev. Biol. 2021, 9, 673396. [Google Scholar] [CrossRef]
- Nickel, J.; Mueller, T.D. Specification of BMP Signaling. Cells 2019, 8, 1579. [Google Scholar] [CrossRef]
- Zhu, H.; Kavsak, P.; Abdollah, S.; Wrana, J.L.; Thomsen, G.H. A SMAD Ubiquitin Ligase Targets the BMP Pathway and Affects Embryonic Pattern Formation. Nature 1999, 400, 687–693. [Google Scholar] [CrossRef]
- Masckauchán, T.N.H.; Shawber, C.J.; Funahashi, Y.; Li, C.M.; Kitajewski, J. Wnt/Beta-Catenin Signaling Induces Proliferation, Survival and Interleukin-8 in Human Endothelial Cells. Angiogenesis 2005, 8, 43–51. [Google Scholar] [CrossRef]
- Tajadura, V.; Hansen, M.H.; Smith, J.; Charles, H.; Rickman, M.; Farrell-Dillon, K.; Claro, V.; Warboys, C.; Ferro, A. β-Catenin Promotes Endothelial Survival by Regulating ENOS Activity and Flow-Dependent Anti-Apoptotic Gene Expression. Cell Death Dis. 2020, 11, 493. [Google Scholar] [CrossRef]
- Zhang, Y.; Alexander, P.B.; Wang, X.F. TGF-β Family Signaling in the Control of Cell Proliferation and Survival. Cold Spring Harb. Perspect. Biol. 2016, 9, a022145. [Google Scholar] [CrossRef]
- Lebrin, F.; Deckers, M.; Bertolino, P.; Ten Dijke, P. TGF-β Receptor Function in the Endothelium. Cardiovasc. Res. 2005, 65, 599–608. [Google Scholar] [CrossRef]
- Ma, J.; Sanchez-Duffhues, G.; Goumans, M.J.; ten Dijke, P. TGF-β-Induced Endothelial to Mesenchymal Transition in Disease and Tissue Engineering. Front. Cell Dev. Biol. 2020, 8, 260. [Google Scholar] [CrossRef]
- Li, P.; Ferrara, N. Vascular Heterogeneity: VEGF Receptors Make Blood Vessels Special. J. Exp. Med. 2022, 219, 212984. [Google Scholar] [CrossRef]
- Hennigs, J.K.; Matuszcak, C.; Trepel, M.; Körbelin, J. Vascular Endothelial Cells: Heterogeneity and Targeting Approaches. Cells 2021, 10, 2712. [Google Scholar] [CrossRef]
- Gurevich, D.B.; David, D.T.; Sundararaman, A.; Patel, J.; Minchiotti, G.; Fico, A.; Brakebusch, C. Endothelial Heterogeneity in Development and Wound Healing. Cells 2021, 10, 2338. [Google Scholar] [CrossRef]
- Matrone, G.; Xia, B.; Chen, K.; Denvir, M.A.; Baker, A.H.; Cooke, J.P. Fli1+ Cells Transcriptional Analysis Reveals an Lmo2-Prdm16 Axis in Angiogenesis. Proc. Natl. Acad. Sci. USA 2021, 118, e2008559118. [Google Scholar] [CrossRef]
- Mattonet, K.; Riemslagh, F.W.; Guenther, S.; Prummel, K.D.; Kesavan, G.; Hans, S.; Ebersberger, I.; Brand, M.; Burger, A.; Reischauer, S.; et al. Endothelial versus Pronephron Fate Decision Is Modulated by the Transcription Factors Cloche/Npas4l, Tal1, and Lmo2. Sci. Adv. 2022, 8, 31. [Google Scholar] [CrossRef]
- Kataoka, H.; Hayashi, M.; Nakagawa, R.; Tanaka, Y.; Izumi, N.; Nishikawa, S.; Jakt, M.L.; Tarui, H.; Nishikawa, S.I. Etv2/ER71 Induces Vascular Mesoderm from Flk1+PDGFRα+ Primitive Mesoderm. Blood 2011, 118, 6975–6986. [Google Scholar] [CrossRef]
- Singh, B.N.; Sierra-Pagan, J.E.; Gong, W.; Das, S.; Theisen, J.W.M.; Skie, E.; Garry, M.G.; Garry, D.J. ETV2 (Ets Variant Transcription Factor 2)- Rhoj Cascade Regulates Endothelial Progenitor Cell Migration During Embryogenesis. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 2875–2890. [Google Scholar] [CrossRef]
- Kim, T.M.; Lee, R.H.; Kim, M.S.; Lewis, C.A.; Park, C. ETV2/ER71, the Key Factor Leading the Paths to Vascular Regeneration and Angiogenic Reprogramming. Stem Cell Res. Ther. 2023, 14, 41. [Google Scholar] [CrossRef]
- Morita, R.; Suzuki, M.; Kasahara, H.; Shimizu, N.; Shichita, T.; Sekiya, T.; Kimura, A.; Sasaki, K.I.; Yasukawa, H.; Yoshimura, A. ETS Transcription Factor ETV2 Directly Converts Human Fibroblasts into Functional Endothelial Cells. Proc. Natl. Acad. Sci. USA 2015, 112, 160–165. [Google Scholar] [CrossRef]
- Martin, A.; Cano, A. Tumorigenesis: Twist1 Links EMT to Self-Renewal. Nat. Cell Biol. 2010, 12, 924–925. [Google Scholar] [CrossRef]
- Liu, T.; Zhao, X.; Zheng, X.; Zheng, Y.; Dong, X.; Zhao, N.; Liao, S.; Sun, B. The EMT Transcription Factor, Twist1, as a Novel Therapeutic Target for Pulmonary Sarcomatoid Carcinomas. Int. J. Oncol. 2020, 56, 750–760. [Google Scholar] [CrossRef]
- Bui, B.P.; Nguyen, P.L.; Lee, K.; Cho, J. Hypoxia-Inducible Factor-1: A Novel Therapeutic Target for the Management of Cancer, Drug Resistance, and Cancer-Related Pain. Cancers 2022, 14, 6054. [Google Scholar] [CrossRef] [PubMed]
- Ziello, J.E.; Jovin, I.S.; Huang, Y. Hypoxia-Inducible Factor (HIF)-1 Regulatory Pathway and Its Potential for Therapeutic Intervention in Malignancy and Ischemia. Yale J. Biol. Med. 2007, 80, 51. [Google Scholar]
- Fernández-Sarmiento, J.; Salazar-Pelaéz, L.M.; Carcillo, J.A. The Endothelial Glycocalyx: A Fundamental Determinant of Vascular Permeability in Sepsis. Pediatr. Crit. Care Med. 2020, 21, e291–e300. [Google Scholar] [CrossRef] [PubMed]
- Jackson, W.F. Microcirculation, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2012; Volume 2, ISBN 9780123815101. [Google Scholar]
- Gilbert, S.F. Epigenetic Landscaping: Waddington’s Use of Cell Fate Bifurcation Diagrams. Biol. Philos. 1991, 6, 135–154. [Google Scholar] [CrossRef]
- Jambusaria, A.; Hong, Z.; Zhang, L.; Srivastava, S.; Jana, A.; Toth, P.T.; Dai, Y.; Malik, A.B.; Rehman, J. Endothelial Heterogeneity across Distinct Vascular Beds during Homeostasis and Inflammation. Elife 2020, 9, e51413. [Google Scholar] [CrossRef] [PubMed]
- De Seranno, S.; Estrella, C.; Loyens, A.; Cornea, A.; Ojeda, S.R.; Beauvillain, J.-C.; Prevot, V. Development/Plasticity/Repair Vascular Endothelial Cells Promote Acute Plasticity in Ependymoglial Cells of the Neuroendocrine Brain. J. Neurosci. 2004, 24, 10353–10363. [Google Scholar] [CrossRef] [PubMed]
- Oak, M.H.; Auger, C.; Belcastro, E.; Park, S.H.; Lee, H.H.; Schini-Kerth, V.B. Potential Mechanisms Underlying Cardiovascular Protection by Polyphenols: Role of the Endothelium. Free. Radic. Biol. Med. 2018, 122, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Hadland, B.K.; Varnum-Finney, B.; Poulos, M.G.; Moon, R.T.; Butler, J.M.; Rafii, S.; Bernstein, I.D. Endothelium and NOTCH Specify and Amplify Aorta-Gonad-Mesonephros-Derived Hematopoietic Stem Cells. J. Clin. Investig. 2015, 125, 2032–2045. [Google Scholar] [CrossRef] [PubMed]
- Slovinski, A.P.; Hajjar, L.A.; Ince, C. Microcirculation in Cardiovascular Diseases. J. Cardiothorac. Vasc. Anesth. 2019, 33, 3458–3468. [Google Scholar] [CrossRef]
- Kalucka, J.; de Rooij, L.P.M.H.; Goveia, J.; Rohlenova, K.; Dumas, S.J.; Meta, E.; Conchinha, N.V.; Taverna, F.; Teuwen, L.A.; Veys, K.; et al. Single-Cell Transcriptome Atlas of Murine Endothelial Cells. Cell 2020, 180, 764–779.e20. [Google Scholar] [CrossRef]
- Pries, A.R.; Secomb, T.W. Modeling Structural Adaptation of Microcirculation. Microcirculation 2008, 15, 753–764. [Google Scholar] [CrossRef] [PubMed]
- Chan, F.L.; Inoue, S. Lamina Lucida of Basement Membrane: An Artefact. Microsc. Res. Tech. 1994, 28, 59. [Google Scholar] [CrossRef]
- Sekiguchi, R.; Yamada, K.M. Basement Membranes in Development and Disease. Curr. Top. Dev. Biol. 2018, 130, 143–191. [Google Scholar]
- Jayadev, R.; Sherwood, D.R. Basement Membranes. Curr. Biol. 2017, 27, R207–R211. [Google Scholar] [CrossRef] [PubMed]
- Sieve, I.; Münster-Kühnel, A.K.; Hilfiker-Kleiner, D. Regulation and Function of Endothelial Glycocalyx Layer in Vascular Diseases. Vasc. Pharmacol. 2018, 100, 26–33. [Google Scholar] [CrossRef]
- Delgadillo, L.F.; Marsh, G.A.; Waugh, R.E. Endothelial Glycocalyx Layer Properties and Its Ability to Limit Leukocyte Adhesion. Biophys. J. 2020, 118, 1564–1575. [Google Scholar] [CrossRef]
- Reiterer, M.; Branco, C.M. Endothelial Cells and Organ Function: Applications and Implications of Understanding Unique and Reciprocal Remodelling. FEBS J. 2020, 287, 1088–1100. [Google Scholar] [CrossRef] [PubMed]
- Aird, W.C. Phenotypic Heterogeneity of the Endothelium: I. Structure, Function, and Mechanisms. Circ. Res. 2007, 100, 158–173. [Google Scholar] [CrossRef]
- Azcutia, V.; Stefanidakis, M.; Tsuboi, N.; Mayadas, T.; Croce, K.J.; Fukuda, D.; Aikawa, M.; Newton, G.; Luscinskas, F.W. Endothelial CD47 Promotes Vascular Endothelial-Cadherin Tyrosine Phosphorylation and Participates in T Cell Recruitment at Sites of Inflammation In Vivo. J. Immunol. 2012, 189, 2553–2562. [Google Scholar] [CrossRef]
- Bazzoni, G.; Dejana, E. Endothelial Cell-to-Cell Junctions: Molecular Organization and Role in Vascular Homeostasis. Physiol. Rev. 2004, 84, 869–901. [Google Scholar] [CrossRef]
- Bearer, E.L.; Orci, L.; Sors, P. Endothelial Fenestral Diaphragms: A Quick-Freeze, Deep-Etch Study. J. Cell Biol. 1985, 100, 418–428. [Google Scholar] [CrossRef] [PubMed]
- Campbell, L.; Abulrob, A.N.G.; Kandalaft, L.E.; Plummer, S.; Hollins, A.J.; Gibbs, A.; Gumbleton, M. Constitutive Expression of P-Glycoprotein in Normal Lung Alveolar Epithelium and Functionality in Primary Alveolar Epithelial Cultures. J. Pharmacol. Exp. Ther. 2003, 304, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Fogo, A.B.; Kon, V. The Glomerulus-a View from the inside-the Endothelial Cell. J. Pharmacol. Exp. Ther. 2010, 42, 1388–1397. [Google Scholar] [CrossRef] [PubMed]
- Fonta, C. Neuronal Tissue-Nonspecific Alkaline Phosphatase (TNAP); Fonta, C., Négyessy, L., Eds.; Subcellular Biochemistry; Springer Netherlands: Dordrecht, The Netherland, 2015; Volume 76, ISBN 978-94-017-7196-2. [Google Scholar]
- Garlanda, C.; Dejana, E. Brief Reviews Heterogeneity of Endothelial Cells. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 1193–1202. [Google Scholar] [CrossRef]
- Goncharov, N.V.; Popova, P.I.; Avdonin, P.P.; Kudryavtsev, I.V.; Serebryakova, M.K.; Korf, E.A.; Avdonin, P.V. Markers of Endothelial Cells in Normal and Pathological Conditions. Biochem. (Mosc.) Suppl. Ser. A Membr. Cell Biol. 2020, 14, 167–183. [Google Scholar] [CrossRef] [PubMed]
- Govindarajan, R.; Chakraborty, S.; Johnson, K.E.; Falk, M.M.; Wheelock, M.J.; Johnson, K.R.; Mehta, P.P. Assembly of Connexin43 into Gap Junctions Is Regulated Differentially by E-Cadherin and N-Cadherin in Rat Liver Epithelial Cells. Mol. Biol. Cell 2010, 21, 4089–4107. [Google Scholar] [CrossRef] [PubMed]
- Grant, D.; Wanner, N.; Frimel, M.; Erzurum, S.; Asosingh, K. Comprehensive Phenotyping of Endothelial Cells Using Flow Cytometry 1: Murine. Cytom. Part A 2020, 99, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Haasdijk, R.A.; Den Dekker, W.K.; Cheng, C.; Tempel, D.; Szulcek, R.; Bos, F.L.; Hermkens, D.M.A.; Chrifi, I.; Brandt, M.M.; Van Dijk, C.; et al. THSD1 Preserves Vascular Integrity and Protects against Intraplaque Haemorrhaging in ApoE-/- Mice. Cardiovasc. Res. 2016, 110, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Ishii, Y.; Hamashima, T.; Yamamoto, S.; Sasahara, M. Pathogenetic Significance and Possibility as a Therapeutic Target of Platelet Derived Growth Factor. Pathol. Int. 2017, 67, 235–246. [Google Scholar] [CrossRef]
- Kakei, Y.; Akashi, M.; Shigeta, T.; Hasegawa, T.; Komori, T. Alteration of Cell-Cell Junctions in Cultured Human Lymphatic Endothelial Cells with Inflammatory Cytokine Stimulation. Lymphat. Res. Biol. 2014, 12, 136–143. [Google Scholar] [CrossRef]
- Keuschnigg, J.; Karinen, S.; Auvinen, K.; Irjala, H.; Mpindi, J.P.; Kallioniemi, O.; Hautaniemi, S.; Jalkanen, S.; Salmi, M. Plasticity of Blood- and Lymphatic Endothelial Cells and Marker Identification. PLoS ONE 2013, 8, e74293. [Google Scholar] [CrossRef]
- Lalor, P.F.; Lai, W.K.; Curbishley, S.M.; Shetty, S.; Adams, D.H. Human Hepatic Sinusoidal Endothelial Cells Can Be Distinguished by Expression of Phenotypic Markers Related to Their Specialised Functions in Vivo. World J. Gastroenterol. 2006, 12, 5429–5439. [Google Scholar] [CrossRef] [PubMed]
- Ligresti, G.; Nagao, R.J.; Xue, J.; Choi, Y.J.; Xu, J.; Ren, S.; Aburatani, T.; Anderson, S.K.; MacDonald, J.W.; Bammler, T.K.; et al. A Novel Three-Dimensional Human Peritubular Microvascular System. J. Am. Soc. Nephrol. 2016, 27, 2370–2381. [Google Scholar] [CrossRef] [PubMed]
- DiBartola, S.P. Fluid, Electrolyte, and Acid-Base Disorders in Small Animal Practice; Elsevier: Amsterdam, The Netherlands, 2012; Volume 53, ISBN 9781437706543. [Google Scholar]
- McMillan, D.B.; Harris, R.J. An Atlas of Comparative Vertebrate Histology; Donald, B., McMillan, R.J.H., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; ISBN 9780124104242. [Google Scholar]
- Mink, D.; Schiller, A.; Kriz, W.; Taugner, R. Interendothelial Junctions in Kidney Vessels. Cell Tissue Res. 1984, 236, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Obeidat, M.; Obeidat, M.; Ballermann, B.J. Glomerular Endothelium: A Porous Sieve and Formidable Barrier. Exp. Cell Res. 2012, 318, 964–972. [Google Scholar] [CrossRef] [PubMed]
- Poisson, J.; Lemoinne, S.; Boulanger, C.; Durand, F.; Moreau, R.; Valla, D.; Rautou, P.E. Liver Sinusoidal Endothelial Cells: Physiology and Role in Liver Diseases. J. Hepatol. 2017, 66, 212–227. [Google Scholar] [CrossRef]
- Pusztaszeri, M.P.; Seelentag, W.; Bosman, F.T. Immunohistochemical Expression of Endothelial Markers CD31, CD34, von Willebrand Factor, and Fli-1 in Normal Human Tissues. J. Histochem. Cytochem. 2006, 54, 385–395. [Google Scholar] [CrossRef]
- Satchell, S.C.; Braet, F. Glomerular Endothelial Cell Fenestrations: An Integral Component of the Glomerular Filtration Barrier. Am. J. Physiol. Renal. Physiol. 2009, 296, F947–F956. [Google Scholar] [CrossRef]
- Sleeman, J.P.; Krishnan, J.; Kirkin, V.; Baumann, P. Markers for the Lymphatic Endothelium: In Search of the Holy Grail? Microsc. Res. Tech. 2001, 55, 61–69. [Google Scholar] [CrossRef]
- Stan, R.V. Endothelial Stomatal and Fenestral Diaphragms in Normal Vessels and Angiogenesis. J. Cell. Mol. Med. 2007, 11, 621–643. [Google Scholar] [CrossRef]
- Stan, R.V.; Tse, D.; Deharvengt, S.J.; Smits, N.C.; Xu, Y.; Luciano, M.R.; McGarry, C.L.; Buitendijk, M.; Nemani, K.V.; Elgueta, R.; et al. The Diaphragms of Fenestrated Endothelia: Gatekeepers of Vascular Permeability and Blood Composition. Dev. Cell 2012, 23, 1203–1218. [Google Scholar] [CrossRef] [PubMed]
- Svistounov, D.; Warren, A.; McNerney, G.P.; Owen, D.M.; Zencak, D.; Zykova, S.N.; Crane, H.; Huser, T.; Quinn, R.J.; Smedsrød, B.; et al. The Relationship between Fenestrations, Sieve Plates and Rafts in Liver Sinusoidal Endothelial Cells. PLoS ONE 2012, 7, e46134. [Google Scholar] [CrossRef] [PubMed]
- Tati, R.; Kristoffersson, A.C.; Ståhl, A.L.; Mörgelin, M.; Motto, D.; Satchell, S.; Mathieson, P.; Manea-Hedström, M.; Karpman, D. Phenotypic Expression of ADAMTS13 in Glomerular Endothelial Cells. PLoS ONE 2011, 6, e21587. [Google Scholar] [CrossRef]
- Tse, D.; Stan, R.V. Morphological Heterogeneity of Endothelium. Semin. Thromb. Hemost. 2010, 36, 236–245. [Google Scholar] [CrossRef]
- Wallez, Y.; Huber, P. Endothelial Adherens and Tight Junctions in Vascular Homeostasis, Inflammation and Angiogenesis. Biochim. Biophys. Acta Biomembr. 2008, 1778, 794–809. [Google Scholar] [CrossRef]
- Zhang, D.; Isaka, Y.; Imamura, R.; Ichimaru, N.; Shi, Y.; Imai, E.; Tian, Y.; Ohtsuka, A.; Takahara, S. Glycocalyx Damage Estimated Using Colloidal Iron Staining. Cell Transplant. 2008, 17, 159–163. [Google Scholar] [CrossRef]
- Zhang, F.; Zarkada, G.; Yi, S.; Eichmann, A. Lymphatic Endothelial Cell Junctions: Molecular Regulation in Physiology and Diseases. Front. Physiol. 2020, 11, 509. [Google Scholar] [CrossRef]
- Shang, T.; Liang, J.; Kapron, C.M.; Liu, J. Pathophysiology of Aged Lymphatic Vessels. Aging 2019, 11, 6602–6613. [Google Scholar] [CrossRef] [PubMed]
- Hunt, N.J.; Lockwood, G.P.; Warren, A.; Mao, H.; McCourt, P.A.G.; Le Couteur, D.G.; Cogger, V.C. Manipulating Fenestrations in Young and Old Liver Sinusoidal Endothelial Cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 316, G144–G154. [Google Scholar] [CrossRef]
- McCarron, J.G.; Lee, M.D.; Wilson, C. The Endothelium Solves Problems That Endothelial Cells Do Not Know Exist. Trends Pharmacol. Sci. 2017, 38, 322–338. [Google Scholar] [CrossRef]
- Orsenigo, F.; Conze, L.L.; Jauhiainen, S.; Corada, M.; Lazzaroni, F.; Malinverno, M.; Sundell, V.; Cunha, S.I.; Brännström, J.; Globisch, M.A.; et al. Mapping Endothelial-Cell Diversity in Cerebral Cavernous Malformations at Single-Cell Resolution. Elife 2020, 9, e61413. [Google Scholar] [CrossRef]
- Paz, N.G.; Amore, P.A.D. Arterial versus Venous Endothelial Cells Nathaniel. Cell Tissue Res. 2014, 335, 5–16. [Google Scholar] [CrossRef]
- Risau, W. Development and Differentiation of Endothelium. Kidney Int. Suppl. 1998, 54, 3–6. [Google Scholar] [CrossRef]
- Jones, E.; Gallimore, A.; Ager, A. Defining High Endothelial Venules and Tertiary Lymphoid Structures in Cancer. Methods Mol. Biol. 2018, 1845, 99–118. [Google Scholar] [CrossRef]
- Blanchard, L.; Girard, J.P. High Endothelial Venules (HEVs) in Immunity, Inflammation and Cancer. Angiogenesis 2021, 24, 719–753. [Google Scholar] [CrossRef]
- Vella, G.; Guelfi, S.; Bergers, G. High Endothelial Venules: A Vascular Perspective on Tertiary Lymphoid Structures in Cancer. Front. Immunol. 2021, 12, 736670. [Google Scholar] [CrossRef]
- Vallance, P. Blood Vessels and the Endothelium. In Oxford Textbook of Medicine; Oxford Textbooks: Oxford, UK, 2003; Volume 2, pp. 1715–1720. [Google Scholar] [CrossRef]
- Dejana, E.; Orsenigo, F.; Lampugnani, M.G. The Role of Adherens Junctions and VE-Cadherin in the Control of Vascular Permeability. J. Cell Sci. 2008, 121, 2115–2122. [Google Scholar] [CrossRef]
- Dejana, E. Endothelial Cell-Cell Junctions: Happy Together. Nat. Rev. Mol. Cell Biol. 2004, 5, 261–270. [Google Scholar] [CrossRef]
- Freund-Michel, V.; Muller, B.; Marthan, R.; Savineau, J.P.; Guibert, C. Expression and Role of Connexin-Based Gap Junctions in Pulmonary Inflammatory Diseases. Pharmacol. Ther. 2016, 164, 105–119. [Google Scholar] [CrossRef]
- Hartsock, A.; Nelson, W.J. Adherens and Tight Junctions: Structure, Function and Connections to the Actin Cytoskeleton. Biochim. Biophys. Acta Biomembr. 2008, 1778, 660–669. [Google Scholar] [CrossRef]
- Campbell, H.K.; Maiers, J.L.; DeMali, K.A. Interplay between Tight Junctions & Adherens Junctions. Exp. Cell Res. 2017, 358, 39–44. [Google Scholar] [CrossRef]
- Winkler, L.; Blasig, R.; Breitkreuz-Korff, O.; Berndt, P.; Dithmer, S.; Helms, H.C.; Puchkov, D.; Devraj, K.; Kaya, M.; Qin, Z.; et al. Tight Junctions in the Blood–Brain Barrier Promote Edema Formation and Infarct Size in Stroke–Ambivalent Effects of Sealing Proteins. J. Cereb. Blood Flow Metab. 2020, 41, 132–145. [Google Scholar] [CrossRef]
- Bhat, A.A.; Uppada, S.; Achkar, I.W.; Hashem, S.; Yadav, S.K.; Shanmugakonar, M.; Al-Naemi, H.A.; Haris, M.; Uddin, S. Tight Junction Proteins and Signaling Pathways in Cancer and Inflammation: A Functional Cross-talk. Front. Physiol. 2019, 10, 1942. [Google Scholar] [CrossRef]
- Takeshita, Y.; Sato, R.; Kanda, T. Blood–Nerve Barrier (BNB) Pathology in Diabetic Peripheral Neuropathy and In Vitro Human BNB Model. Int. J. Mol. Sci. 2020, 22, 62. [Google Scholar] [CrossRef]
- Stamatovic, S.M.; Johnson, A.M.; Keep, R.F.; Andjelkovic, A.V. Junctional Proteins of the Blood-Brain Barrier: New Insights into Function and Dysfunction. Tissue Barriers 2016, 4, e1154641. [Google Scholar] [CrossRef]
- Wittekindt, O.H. Tight Junctions in Pulmonary Epithelia during Lung Inflammation. Pflüg. Arch. Eur. J. Physiol. 2017, 469, 135–147. [Google Scholar] [CrossRef]
- Bing, S.; Chen, Z.; Gu, J.; Tse, G.; Jiang, J.; Huang, F.; Zhao, C. Tight Junction Proteins and Gap Junction Proteins Play Important Roles in High Fat Dietary Atherosclerosis Pathogenesis. Int. J. Clin. Exp. Pathol. 2016, 9, 7969–7976. [Google Scholar]
- Tym, K.; Mathieu-Costello, O. Structural and Functional Changes in the Microvasculature of Disused Skeletal Muscle. Front. Biosci. 2001, 6, d45. [Google Scholar] [CrossRef]
- Harris, T.J.C.; Tepass, U. Adherens Junctions: From Molecules to Morphogenesis. Nat. Rev. Mol. Cell Biol. 2010, 11, 502–514. [Google Scholar] [CrossRef]
- Gonschior, H.; Haucke, V.; Lehmann, M. Super-Resolution Imaging of Tight and Adherens Junctions: Challenges and Open Questions. Int. J. Mol. Sci. 2020, 21, 744. [Google Scholar] [CrossRef]
- Gavard, J.; Gutkind, S.J. VE-Cadherin and Claudin-5: It Takes Two to Tango. Nat. Cell Biol. 2008, 10, 883. [Google Scholar] [CrossRef]
- Komarova, Y.A.; Kruse, K.; Mehta, D.; Malik, A.B. Protein Interactions at Endothelial Junctions and Signaling Mechanisms Regulating Endothelial Permeability. Circ. Res. 2017, 120, 179–206. [Google Scholar] [CrossRef]
- Peracchia, C. Connexin/Innexin Channels in Cytoplasmic Organelles. Are There Intracellular Gap Junctions? A Hypothesis! Int. J. Mol. Sci. 2020, 21, 2163. [Google Scholar] [CrossRef]
- Socha, M.J.; Segal, S.S. Vascular Aging: Microvascular Mechanisms Limiting Skeletal Muscle Blood Flow with Advancing Age. J. Appl. Physiol. 2018, 125, 1851. [Google Scholar] [CrossRef]
- Kovacs-Oller, T.; Ivanova, E.; Bianchimano, P.; Sagdullaev, B.T. The Pericyte Connectome: Spatial Precision of Neurovascular Coupling Is Driven by Selective Connectivity Maps of Pericytes and Endothelial Cells and Is Disrupted in Diabetes. Cell Discov. 2020, 6, 39. [Google Scholar] [CrossRef]
- Maibier, M.; Bintig, W.; Goede, A.; Höpfner, M.; Kuebler, W.M.; Secomb, T.W.; Nitzsche, B.; Pries, A.R. Gap Junctions Regulate Vessel Diameter in Chick Chorioallantoic Membrane Vasculature by Both Tone-Dependent and Structural Mechanisms. Microcirculation 2020, 27, e12590. [Google Scholar] [CrossRef]
- Okamoto, T.; Kawamoto, E.; Takagi, Y.; Akita, N.; Hayashi, T.; Park, E.J.; Suzuki, K.; Shimaoka, M. Gap Junction-Mediated Regulation of Endothelial Cellular Stiffness. Sci. Rep. 2017, 7, 6134. [Google Scholar] [CrossRef]
- Favero, G.; Paganelli, C.; Buffoli, B.; Rodella, L.F.; Rezzani, R. Endothelium and Its Alterations in Cardiovascular Diseases: Life Style Intervention. Biomed Res. Int. 2014, 2014, 801896. [Google Scholar] [CrossRef]
- Xu, B.; Broome, U.; Uzunel, M.; Nava, S.; Ge, X.; Kumagai-Braesch, M.; Hultenby, K.; Christensson, B.; Ericzon, B.G.; Holgersson, J.; et al. Capillarization of Hepatic Sinusoid by Liver Endothelial Cell-Reactive Autoantibodies in Patients with Cirrhosis and Chronic Hepatitis. Am. J. Pathol. 2003, 163, 1275–1289. [Google Scholar] [CrossRef]
- Murray, J.W.; Spray, D.C. Gap and Tight Junctions in Liver. In The Liver; Wiley: Hoboken, NJ, USA, 2020; pp. 160–173. [Google Scholar]
- Chi, J.T.J.-T.; Chang, H.Y.; Haraldsen, G.; Jahnsen, F.L.; Troyanskaya, O.G.; Chang, D.S.; Wang, Z.; Rockson, S.G.; van de Rijn, M.; Botstein, D.; et al. Endothelial Cell Diversity Revealed by Global Expression Profiling. Proc. Natl. Acad. Sci. USA 2003, 100, 10623–10628. [Google Scholar] [CrossRef]
- Turunen, M.P.; Aavik, E.; Ylä-Herttuala, S. Epigenetics and Atherosclerosis. Biochim. Biophys. Acta Gen. Subj. 2009, 1790, 886–891. [Google Scholar] [CrossRef]
- Friso, S.; Carvajal, C.A.; Fardella, C.E.; Olivieri, O. Epigenetics and Arterial Hypertension: The Challenge of Emerging Evidence. Transl. Res. 2015, 165, 154–165. [Google Scholar] [CrossRef]
- Gorabi, A.M.; Penson, P.E.; Banach, M.; Motallebnezhad, M.; Jamialahmadi, T.; Sahebkar, A. Epigenetic Control of Atherosclerosis via DNA Methylation: A New Therapeutic Target? Life Sci. 2020, 253, 117682. [Google Scholar] [CrossRef]
- Certo, M.; Elkafrawy, H.; Pucino, V.; Cucchi, D.; Cheung, K.C.P.; Mauro, C. Endothelial Cell and T-cell Cross-talk: Targeting Metabolism as a Therapeutic Approach in Chronic Inflammation. Br. J. Pharmacol. 2021, 178, 2041. [Google Scholar] [CrossRef]
- Howe, K.L.; Cybulsky, M.; Fish, J.E. The Endothelium as a Hub for Cellular Communication in Atherogenesis: Is There Directionality to the Message? Front. Cardiovasc. Med. 2022, 9, 888390. [Google Scholar] [CrossRef]
- Przysinda, A.; Feng, W.; Li, G. Diversity of Organism-Wide and Organ-Specific Endothelial Cells. Curr. Cardiol. Rep. 2020, 22, 4–9. [Google Scholar] [CrossRef]
- Glinskii, O.V.; Huxley, V.H.; Xie, L.; Bunyak, F.; Palaniappan, K.; Glinsky, V.V. Complex Non-Sinus-Associated Pachymeningeal Lymphatic Structures: Interrelationship with Blood Microvasculature. Front. Physiol. 2019, 10, 1364. [Google Scholar] [CrossRef]
- Xiang, M.; Grosso, R.A.; Takeda, A.; Pan, J.; Bekkhus, T.; Brulois, K.; Dermadi, D.; Nordling, S.; Vanlandewijck, M.; Jalkanen, S.; et al. A Single-Cell Transcriptional Roadmap of the Mouse and Human Lymph Node Lymphatic Vasculature. Front. Cardiovasc. Med. 2020, 7, 52. [Google Scholar] [CrossRef]
- He, W.; Jing, Y.; Wan, Q.; Xiao, K.; Chen, K.; Lu, Y.; Li, L.; Tang, Y.; Deng, Y.; Yao, Z.; et al. The Anatomy and Metabolome of the Lymphatic System in the Brain in Health and Disease. Brain Pathol. 2020, 30, 392–404. [Google Scholar] [CrossRef]
- Ma, Q.; Schlegel, F.; Bachmann, S.B.; Schneider, H.; Decker, Y.; Rudin, M.; Weller, M.; Proulx, S.T.; Detmar, M. Lymphatic Outflow of Cerebrospinal Fluid Is Reduced in Glioma. Sci. Rep. 2019, 9, 14815. [Google Scholar] [CrossRef]
- Petrova, T.V.; Koh, G.Y. Biological Functions of Lymphatic Vessels. Science 2020, 369, eaax4063. [Google Scholar] [CrossRef]
- Morfoisse, F.; Noel, A. Lymphatic and Blood Systems: Identical or Fraternal Twins? International Journal of Biochemistry and Cell Biol. 2019, 114, 105562. [Google Scholar] [CrossRef]
- Liu, X.; Gu, X.; Ma, W.; Oxendine, M.; Gil, H.J.; Davis, G.E.; Cleaver, O.; Oliver, G. Rasip1 Controls Lymphatic Vessel Lumen Maintenance by Regulating Endothelial Cell Junctions. Development 2018, 145, dev.165092. [Google Scholar] [CrossRef]
- Oliver, G.; Kipnis, J.; Randolph, G.J.; Harvey, N.L. The Lymphatic Vasculature in the 21st Century: Novel Functional Roles in Homeostasis and Disease. Cell 2020, 182, 270–296. [Google Scholar] [CrossRef]
- Yao, L.C.; Baluk, P.; Srinivasan, R.S.; Oliver, G.; McDonald, D.M. Plasticity of Button-like Junctions in the Endothelium of Airway Lymphatics in Development and Inflammation. Am. J. Pathol. 2012, 180, 2561–2575. [Google Scholar] [CrossRef]
- Kazenwadel, J.; Harvey, N.L. ScienceDirect Lymphatic Endothelial Progenitor Cells: Origins and Roles in Lymphangiogenesis. Curr. Opin. Immunol. 2018, 53, 81–87. [Google Scholar] [CrossRef]
- Volk-Draper, L.D.; Hall, K.L.; Wilber, A.C.; Ran, S. Lymphatic Endothelial Progenitors Originate from Plastic Myeloid Cells Activated by Toll-like Receptor-4. PLoS ONE 2017, 12, e0179257. [Google Scholar] [CrossRef]
- Shoemaker, L.D.; Fuentes, L.F.; Santiago, S.M.; Allen, B.M.; Cook, D.J.; Steinberg, G.K.; Chang, S.D. Human Brain Arteriovenous Malformations Express Lymphatic-Associated Genes. Ann. Clin. Transl. Neurol. 2014, 1, 982–995. [Google Scholar] [CrossRef]
- Antila, S.; Karaman, S.; Nurmi, H.; Airavaara, M.; Voutilainen, M.H.; Mathivet, T.; Chilov, D.; Li, Z.; Koppinen, T.; Park, J.H.; et al. Development and Plasticity of Meningeal Lymphatic Vessels. J. Exp. Med. 2017, 214, 3645–3667. [Google Scholar] [CrossRef]
- Pawlak, J.B.; Caron, K.M. Lymphatic Programing and Specialization in Hybrid Vessels. Front. Physiol. 2020, 11, 114. [Google Scholar] [CrossRef]
- Surya, V.N.; Michalaki, E.; Huang, E.Y.; Fuller, G.G.; Dunn, A.R. Sphingosine 1-Phosphate Receptor 1 Regulates the Directional Migration of Lymphatic Endothelial Cells in Response to Fluid Shear Stress. J. R. Soc. Interface 2016, 13, 20160823. [Google Scholar] [CrossRef]
- Dellinger, M.T.; Meadows, S.M.; Wynne, K.; Cleaver, O.; Brekken, R.A. Vascular Endothelial Growth Factor Receptor-2 Promotes the Development of the Lymphatic Vasculature. PLoS ONE 2013, 8, e74686. [Google Scholar] [CrossRef]
- Semo, J.; Nicenboim, J.; Yaniv, K. Development of the Lymphatic System: New Questions and Paradigms. Development 2016, 143, 924–935. [Google Scholar] [CrossRef]
- Gancz, D.; Perlmoter, G.; Yaniv, K. Formation and Growth of Cardiac Lymphatics during Embryonic Development, Heart Regeneration, and Disease. Cold Spring Harb. Perspect. Biol. 2020, 12, a037176. [Google Scholar] [CrossRef]
- Shen, B.; Shang, Z.; Wang, B.; Zhang, L.; Zhou, F.; Li, T.; Chu, M.; Jiang, H.; Wang, Y.; Qiao, T.; et al. Genetic Dissection of Tie Pathway in Mouse Lymphatic Maturation and Valve Development. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 1221–1230. [Google Scholar] [CrossRef]
- Engelbrecht, E.; Levesque, M.V.; He, L.; Vanlandewijck, M.; Nitzsche, A.; Niazi, H.; Kuo, A.; Singh, S.A.; Aikawa, M.; Holton, K.; et al. Sphingosine 1-Phosphate-Regulated Transcriptomes in Heterogenous Arterial and Lymphatic Endothelium of the Aorta. Elife 2020, 9, e52690. [Google Scholar] [CrossRef]
- Da Mesquita, S.; Papadopoulos, Z.; Dykstra, T.; Brase, L.; Farias, F.G.; Wall, M.; Jiang, H.; Kodira, C.D.; de Lima, K.A.; Herz, J.; et al. Meningeal Lymphatics Affect Microglia Responses and Anti-Aβ Immunotherapy. Nature 2021, 593, 255–260. [Google Scholar] [CrossRef]
- Norden, P.R.; Kume, T. The Role of Lymphatic Vascular Function in Metabolic Disorders. Front. Physiol. 2020, 11, 404. [Google Scholar] [CrossRef]
- Ho, Y.C.; Srinivasan, R.S. Lymphatic Vasculature in Energy Homeostasis and Obesity. Front. Physiol. 2020, 11, 3. [Google Scholar] [CrossRef]
- Goodall, E.F.; Leach, V.; Wang, C.; Cooper-Knock, J.; Heath, P.R.; Baker, D.; Drew, D.R.; Jill Saffrey, M.; Simpson, J.E.; Romero, I.A.; et al. Age-Associated MRNA and MiRNA Expression Changes in the Blood-Brain Barrier. Int. J. Mol. Sci. 2019, 20, 3097. [Google Scholar] [CrossRef]
- Geevarghese, A.; Herman, I.M. Pericyte-Endothelial Cross-Talk: Implications and Opportunities for Advanced Cellular Therapies. Transl. Res. 2014, 163, 296. [Google Scholar] [CrossRef] [PubMed]
- Longden, T.A.; Zhao, G.; Hariharan, A.; Lederer, W.J. Pericytes and the Control of Blood Flow in Brain and Heart. Annu. Rev. Physiol. 2023, 85, 137. [Google Scholar] [CrossRef] [PubMed]
- Bergers, G.; Song, S. The Role of Pericytes in Blood-Vessel Formation and Maintenance. Neuro Oncol. 2005, 7, 452. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Fu, J.; Huang, Y.; Duan, R.; Zhang, W.; Wang, C.; Wang, S.; Hu, X.; Zhao, H.; Wang, L.; et al. Biology and Function of Pericytes in the Vascular Microcirculation. Anim. Model Exp. Med. 2023, 6, 337–345. [Google Scholar] [CrossRef]
- Van Splunder, H.; Villacampa, P.; Martínez-Romero, A.; Graupera, M. Pericytes in the Disease Spotlight Cell Biology. Trends Cell Biol. 2024, 34, 58–71. [Google Scholar] [CrossRef] [PubMed]
- Van Noorden, C.J.F.; Yetkin-Arik, B.; Serrano Martinez, P.; Bakker, N.; van Breest Smallenburg, M.E.; Schlingemann, R.O.; Klaassen, I.; Majc, B.; Habic, A.; Bogataj, U.; et al. New Insights in ATP Synthesis as Therapeutic Target in Cancer and Angiogenic Ocular Diseases. J. Histochem. Cytochem. 2024, 72, 329–352. [Google Scholar] [CrossRef] [PubMed]
- Vinet, L.; Zhedanov, A. The Blood-Brain and Other Neural Barriers; Nag, S., Ed.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2011; Volume 686, ISBN 978-1-60761-937-6. [Google Scholar]
- Liebner, S.; Dijkhuizen, R.M.; Reiss, Y.; Plate, K.H.; Agalliu, D.; Constantin, G. Functional Morphology of the Blood–Brain Barrier in Health and Disease. Acta Neuropathol. 2018, 135, 311–336. [Google Scholar] [CrossRef] [PubMed]
- Helms, H.C.; Abbott, N.J.; Burek, M.; Cecchelli, R.; Couraud, P.O.; Deli, M.A.; Förster, C.; Galla, H.J.; Romero, I.A.; Shusta, E.V.; et al. In Vitro Models of the Blood-Brain Barrier: An Overview of Commonly Used Brain Endothelial Cell Culture Models and Guidelines for Their Use. J. Cereb. Blood Flow Metab. 2015, 36, 862–890. [Google Scholar] [CrossRef] [PubMed]
- Van Hinsbergh, V.W.M. Endothelium-Role in Regulation of Coagulation and Inflammation. Semin. Immunopathol. 2012, 34, 93–106. [Google Scholar] [CrossRef]
- Wilhelm, I.; Nyúl-Tóth, Á.; Suciu, M.; Hermenean, A.; Krizbai, I.A. Heterogeneity of the Blood-Brain Barrier. Tissue Barriers 2016, 4, e1143544. [Google Scholar] [CrossRef]
- Mbagwu, S.I.; Filgueira, L. Differential Expression of CD31 and Von Willebrand Factor on Endothelial Cells in Different Regions of the Human Brain: Potential Implications for Cerebral Malaria Pathogenesis. Brain Sci. 2020, 10, 31. [Google Scholar] [CrossRef]
- Haruwaka, K.; Ikegami, A.; Tachibana, Y.; Ohno, N.; Konishi, H.; Hashimoto, A.; Matsumoto, M.; Kato, D.; Ono, R.; Kiyama, H.; et al. Dual Microglia Effects on Blood Brain Barrier Permeability Induced by Systemic Inflammation. Nat. Commun. 2019, 10, 5816. [Google Scholar] [CrossRef]
- Santisteban, M.; Ahn, S.J.; Lane, D.; Faraco, G.; Garcia-Bonilla, L.; Racchumi, G.; Poon, C.; Schaeffer, S.; Segarra, S.; Körbelin, J.; et al. Endothelium-Macrophage Cross-talk Mediates Blood-Brain Barrier Dysfunction in Hypertension. Hypertension 2020, 76, 795–807. [Google Scholar] [CrossRef]
- Tunon-Ortiz, A.; Lamb, T.J. Blood Brain Barrier Disruption in Cerebral Malaria: Beyond Endothelial Cell Activation. PLoS Pathog. 2019, 15, e1007786. [Google Scholar] [CrossRef] [PubMed]
- Bonney, S.; Seitz, S.; Ryan, C.A.; Jones, K.L.; Clarke, P.; Tyler, K.L.; Siegenthaler, J.A. Gamma Interferon Alters Junctional Integrity via Rho Kinase, Resulting in Blood-Brain Barrier Leakage in Experimental Viral Encephalitis. mBio 2019, 10, 10–1128. [Google Scholar] [CrossRef]
- Ross, O.N.; Moore, C.M.; Suggett, D.J.; Macintyre, H.L.; Geider, R.J. Endothelial-Cardiomyocyte Interactions. Annu. Rev Physiol. 2008, 53, 1835–1852. [Google Scholar] [CrossRef]
- Brutsaert, D.L.; De Keulenaer, G.W.; Fransen, P.; Mohan, P.; Kaluza, G.L.; Andries, L.J.; Rouleau, J.-L.; Sys, S.U. The Cardiac Endothelium: Functional Morphology, Development, and Physiology. Prog. Cardiovasc. Dis. 1996, 39, 239–262. [Google Scholar] [CrossRef]
- Smiljic, S. The Clinical Significance of Endocardial Endothelial Dysfunction. Medicina 2017, 53, 295–302. [Google Scholar] [CrossRef]
- Piper, H.M.; Spahr, R.; Mertens, S.; Krützfeldt, A.; Watanabe, H. Microvascular Endothelial Cells from Heart. In Cell Culture Techniques in Heart and Vessel Research; Springer: Berlin\Heidelberg, Germany, 1990; pp. 158–177. [Google Scholar]
- Taylor, J.; Fischer, A. Endothelial Cells Dictate Cardiac Fuel Source. Aging 2019, 11, 1083–1084. [Google Scholar] [CrossRef]
- Yazdani, S.; Jaldin-Fincati, J.R.; Pereira, R.V.S.; Klip, A. Endothelial Cell Barriers: Transport of Molecules between Blood and Tissues. Traffic 2019, 20, 390–403. [Google Scholar] [CrossRef]
- Colliva, A.; Braga, L.; Giacca, M.; Zacchigna, S. Endothelial Cell–Cardiomyocyte Cross-talk in Heart Development and Disease. J. Physiol. 2020, 598, 2923–2939. [Google Scholar] [CrossRef]
- Segers, V.F.M.; Brutsaert, D.L.; De Keulenaer, G.W. Cardiac Remodeling: Endothelial Cells Have More to Say than Just NO. Front. Physiol. 2018, 9, 382. [Google Scholar] [CrossRef] [PubMed]
- Pratumvinit, B.; Reesukumal, K.; Janebodin, K.; Ieronimakis, N.; Reyes, M. Isolation, Characterization, and Transplantation of Cardiac Endothelial Cells. Biomed Res. Int. 2013, 2013, 359412. [Google Scholar] [CrossRef]
- Kongpol, K.; Nernpermpisooth, N.; Prompunt, E.; Kumphune, S. Endothelial-Cell-Derived Human Secretory Leukocyte Protease Inhibitor (SLPI) Protects Cardiomyocytes against Ischemia/Reperfusion Injury. Biomolecules 2019, 9, 678. [Google Scholar] [CrossRef]
- Talman, V.; Kivelä, R. Cardiomyocyte—Endothelial Cell Interactions in Cardiac Remodeling and Regeneration. Front. Cardiovasc. Med. 2018, 5, 101. [Google Scholar] [CrossRef]
- Mebazaa, A.; Wetzel, R.; Cherian, M.; Abraham, M. Comparison between Endocardial and Great Vessel Endothelial Cells: Morphology, Growth, and Prostaglandin Release. Am. J. Physiol. Heart Circ. Physiol. 1995, 268, H250–H259. [Google Scholar] [CrossRef]
- Yu, T.; Jing, M.; Gao, Y.; Liu, C.; Liu, L.; Jia, H.; Liu, P.; Chang, M. Study on the Relationship between Hyperthyroidism and Vascular Endothelial Cell Damage. Sci. Rep. 2020, 10, 6992. [Google Scholar] [CrossRef]
- Dal Lin, C.; Tona, F.; Osto, E. The Cross-talk between the Cardiovascular and the Immune System. Vasc. Biol. 2019, 1, H83–H88. [Google Scholar] [CrossRef]
- Mojiri, A.; Alavi, P.; Lorenzana Carrillo, M.A.; Nakhaei-Nejad, M.; Sergi, C.M.; Thebaud, B.; Aird, W.C.; Jahroudi, N. Endothelial Cells of Different Organs Exhibit Heterogeneity in von Willebrand Factor Expression in Response to Hypoxia. Atherosclerosis 2019, 282, 1–10. [Google Scholar] [CrossRef]
- Borsig, L. Selectins in Cancer Immunity. Glycobiology 2018, 28, 648. [Google Scholar] [CrossRef]
- González-Tajuelo, R.; Silván, J.; Pérez-Frías, A.; De La Fuente-Fernández, M.; Tejedor, R.; Espartero-Santos, M.; Vicente-Rabaneda, E.; Juarranz, Á.; Muñoz-Calleja, C.; Castañeda, S.; et al. P-Selectin Preserves Immune Tolerance in Mice and Is Reduced in Human Cutaneous Lupus. Sci. Rep. 2017, 7, srep41841. [Google Scholar] [CrossRef]
- Huertas, A.; Guignabert, C.; Barberà, J.A.; Bärtsch, P.; Bhattacharya, J.; Bhattacharya, S.; Bonsignore, M.R.; Dewachter, L.; Dinh-Xuan, A.T.; Dorfmüller, P.; et al. Pulmonary Vascular Endothelium: The Orchestra Conductor in Respiratory Diseases. Eur. Respir. J. 2018, 51, 1700745. [Google Scholar] [CrossRef] [PubMed]
- Fundamentals of Vascular Biology; Springer Nature: Dordrecht, The Netherlands, 2019. [CrossRef]
- Niethamer, T.K.; Stabler, C.T.; Leach, J.P.; Zepp, J.A.; Morley, M.P.; Babu, A.; Zhou, S.; Morrisey, E.E. Defining the Role of Pulmonary Endothelial Cell Heterogeneity in the Response to Acute Lung Injury. Elife 2020, 9, e53072. [Google Scholar] [CrossRef] [PubMed]
- Gillich, A.; Zhang, F.; Farmer, C.G.; Travaglini, K.J.; Tan, S.Y.; Gu, M.; Zhou, B.; Feinstein, J.A.; Krasnow, M.A.; Metzger, R.J. Capillary Cell-Type Specialization in the Alveolus. Nature 2020, 7, 785–789. [Google Scholar] [CrossRef] [PubMed]
- Millar, F.R.; Summers, C.; Griffiths, M.J.; Toshner, M.R.; Proudfoot, A.G. The Pulmonary Endothelium in Acute Respiratory Distress Syndrome: Insights and Therapeutic Opportunities. Thorax 2016, 71, 462–473. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.R. Human Pulmonary Endothelial Cells in Culture: Activities of Cells from Arteries and Cells from Veins. J. Clin. Investig. 1980, 65, 841–850. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, J.N.; Verin, A.D. Pulmonary Vascular Endothelial Cells. In Endothelial Dysfunction—Old Concepts and New Challenges; IntechOpen: London, UK, 2018; Chapter 12; pp. 287–305. [Google Scholar] [CrossRef]
- Stevens, T. Functional and Molecular Heterogeneity of Pulmonary Endothelial Cells. Proc. Am. Thorac. Soc. 2011, 8, 453–457. [Google Scholar] [CrossRef] [PubMed]
- Comhair, S.A.A.; Xu, W.; Mavrakis, L.; Aldred, M.A.; Asosingh, K.; Erzurum, S.C. Human Primary Lung Endothelial Cells in Culture. Am. J. Respir. Cell Mol. Biol. 2012, 46, 723–730. [Google Scholar] [CrossRef] [PubMed]
- Murray, G.P.; Post, S.R.; Post, G.R. ABO Blood Group Is a Determinant of von Willebrand Factor Protein Levels in Human Pulmonary Endothelial Cells. J. Clin. Pathol. 2020, 73, 347–349. [Google Scholar] [CrossRef]
- Johansson, B.B. The Role of the Endothelial Cell Surface Charge for Blood-Brain Barrier Function. In New Concepts of a Blood—Brain Barrier; Springer: Boston, MA, USA, 1995; Volume 30, pp. 63–70. [Google Scholar]
- Segal, M.S.; Baylis, C.; Johnson, R.J. Endothelial Health and Diversity in the Kidney. J. Am. Soc. Nephrol. 2006, 17, 323–324. [Google Scholar] [CrossRef]
- Maestroni, S.; Zerbini, G. Glomerular Endothelial Cells versus Podocytes as the Cellular Target in Diabetic Nephropathy. Acta Diabetol. 2018, 55, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Satchell, S.C.; Neal, C.R.; McKenzie, E.A.; Tooke, J.E.; Mathieson, P.W. Glomerular Endothelial Glycocalyx Constitutes a Barrier to Protein Permeability. J. Am. Soc. Nephrol. 2007, 18, 2885–2893. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Lee, K.; Chuang, P.Y.; Liu, Z.H.; He, J.C. Glomerular Endothelial Cell Injury and Cross Talk in Diabetic Kidney Disease. Am. J. Physiol. Renal. Physiol. 2015, 308, F287. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.K.; Molitoris, B.A. Renal Endothelial Injury and Microvascular Dysfunction in Acute Kidney Injury. Semin. Nephrol. 2015, 35, 96–107. [Google Scholar] [CrossRef]
- Jourde-Chiche, N.; Fakhouri, F.; Dou, L.; Bellien, J.; Burtey, S.; Frimat, M.; Jarrot, P.A.; Kaplanski, G.; Le Quintrec, M.; Pernin, V.; et al. Endothelium Structure and Function in Kidney Health and Disease. Nat. Rev. Nephrol. 2019, 15, 87–108. [Google Scholar] [CrossRef] [PubMed]
- Noroozinia, F.; Mahmoudzadeh, L.; Gharalari, F.H.; Makhdoomi, K.; Abbasi, A. Relationship between Interstitial CD34 Positive Cells and Active Phase of Lupus Nephritis. Eur. J. Rheumatol. 2018, 5, 254–257. [Google Scholar] [CrossRef]
- Masum, M.A.; Ichii, O.; Elewa, Y.H.A.; Nakamura, T.; Kon, Y. Local CD34-Positive Capillaries Decrease in Mouse Models of Kidney Disease Associating with the Severity of Glomerular and Tubulointerstitial Lesions. BMC Nephrol. 2017, 18, 280. [Google Scholar] [CrossRef]
- Ogawa, H.; Rafiee, P.; Fisher, P.J.; Johnson, N.A.; Otterson, M.F.; Binion, D.G. Butyrate Modulates Gene and Protein Expression in Human Intestinal Endothelial Cells. Biochem. Biophys. Res. Commun. 2003, 309, 512–519. [Google Scholar] [CrossRef]
- Vancamelbeke, M.; Vermeire, S. The Intestinal Barrier: A Fundamental Role in Health and Disease. Expert Rev. Gastroenterol. Hepatol. 2017, 11, 821–834. [Google Scholar] [CrossRef]
- Thomas, H. Intestinal Tract: Gut Endothelial Cells-Another Line of Defence. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 4. [Google Scholar] [CrossRef]
- Lin, X.; Jiang, S.; Jiang, Z.; Zheng, C.; Gou, Z. Effects of Equol on H2O2-Induced Oxidative Stress in Primary Chicken Intestinal Epithelial Cells. Poult. Sci. 2016, 95, 1380–1386. [Google Scholar] [CrossRef] [PubMed]
- Chelakkot, C.; Ghim, J.; Ryu, S.H. Mechanisms Regulating Intestinal Barrier Integrity and Its Pathological Implications. Exp. Mol. Med. 2018, 50, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Hernandez, V.; Quiros, M.; Nusrat, A. Intestinal Epithelial Claudins: Expression and Regulation in Homeostasis and Inflammation. Ann. N. Y. Acad. Sci. 2017, 1397, 66–79. [Google Scholar] [CrossRef] [PubMed]
- Haraldsen, G.; Rugtveit, J.; Kvale, D.; Scholz, T.; Muller, W.A.; Hovig, T.; Brandtzaeg, P. Isolation and Longterm Culture of Human Intestinal Microvascular Endothelial Cels. Gut 1995, 37, 225–234. [Google Scholar] [CrossRef]
- Haraldsen, G.; Kvale, D.; Lien, B.; Farstad, I.N.; Brandtzaeg, P. Cytokine-Regulated Expression of E-Selectin, Intercellular Adhesion Molecule-1 (ICAM-1), and Vascular Cell Adhesion Molecule-1 (VCAM-1) in Human Microvascular Endothelial Cells. J. Immunol. 1996, 156, 2558–2565. [Google Scholar] [CrossRef] [PubMed]
- Cromer, W.E.; Mathis, J.M.; Granger, D.N.; Chaitanya, G.V.; Alexander, J.S. Role of the Endothelium in Inflammatory Bowel Diseases. World J. Gastroenterol. 2011, 17, 578–593. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, V.; Harris, E.N.; Kidambi, S. SECs (Sinusoidal Endothelial Cells), Liver Microenvironment, and Fibrosis. Biomed. Res. Int. 2017, 2017, 4097205. [Google Scholar] [CrossRef] [PubMed]
- Cogger, V.C.; Le Couteur, D.G. Fenestrations in the Liver Sinusoidal Endothelial Cell. In The Liver; John Wiley & Sons, Ltd.: Chichester, UK, 2009; pp. 389–406. ISBN 9780470723135. [Google Scholar]
- Shubham, S.; Kumar, D.; Rooge, S.; Maras, J.S.; Maheshwari, D.; Nautiyal, N.; Kumari, R.; Bhat, A.; Kumar, G.; Rastogi, A.; et al. Cellular and Functional Loss of Liver Endothelial Cells Correlates with Poor Hepatocyte Regeneration in Acute-on-Chronic Liver Failure. Hepatol. Int. 2019, 13, 777–787. [Google Scholar] [CrossRef] [PubMed]
- Baiocchini, A.; Del Nonno, F.; Taibi, C.; Visco-Comandini, U.; D’Offizi, G.; Piacentini, M.; Falasca, L. Liver Sinusoidal Endothelial Cells (LSECs) Modifications in Patients with Chronic Hepatitis C. Sci. Rep. 2019, 9, 8760. [Google Scholar] [CrossRef]
- Géraud, C.; Schledzewski, K.; Demory, A.; Klein, D.; Kaus, M.; Peyre, F.; Sticht, C.; Evdokimov, K.; Lu, S.; Schmieder, A.; et al. Liver Sinusoidal Endothelium: A Microenvironment-Dependent Differentiation Program in Rat Including the Novel Junctional Protein Liver Endothelial Differentiation-Associated Protein-1. Hepatology 2010, 52, 313–326. [Google Scholar] [CrossRef]
- Sørensen, K.K.; Simon-Santamaria, J.; McCuskey, R.S.; Smedsrød, B. Liver Sinusoidal Endothelial Cells. Compr. Physiol. 2015, 5, 1751–1774. [Google Scholar] [CrossRef] [PubMed]
- DeLeve, L.D.; Maretti-Mira, A.C. Liver Sinusoidal Endothelial Cell: An Update. Semin. Liver Dis. 2017, 37, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, K.K.; Smedsrød, B. The Liver Sinusoidal Endothelial Cell. In The Liver; Wiley: Hoboken, NJ, USA, 2020; pp. 422–434. ISBN 9780470723135. [Google Scholar]
- Steiniger, B.S. Human Spleen Microanatomy: Why Mice Do Not Suffice. Immunology 2015, 145, 334–346. [Google Scholar] [CrossRef] [PubMed]
- De Bruyn, P.P.H.; Cho, Y. Contractile Structures in Endothelial Cells of Splenic Sinusoids. J. Ultrastruct. Res. 1974, 49, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Weiss, L. Electron Microscopy of the Red Pulp of Human Spleen. Am. J. Anat. 1972, 134, 425–457. [Google Scholar] [CrossRef] [PubMed]
- Structure and Functions of the Human Spleen: Relationship between Microcirculation and Splenic Functions. Available online: https://pubmed.ncbi.nlm.nih.gov/2689682/ (accessed on 4 June 2024).
- Uehara, K.; Uehara, A. Differentiated Localizations of Phosphorylated Focal Adhesion Kinase in Endothelial Cells of Rat Splenic Sinus. Cell Tissue Res. 2016, 364, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Salama, M.E.; Hu, C.S.; Li, Y.; Wang, X.; Hoffman, R. The Characteristics of Vessel Lining Cells in Normal Spleens and Their Role in the Pathobiology of Myelofibrosis. Blood Adv. 2018, 2, 1130–1145. [Google Scholar] [CrossRef]
- Almenar, S.; Rios-Navarro, C.; Ortega, M.; Molina, P.; Ferrandez-Izquierdo, A.; Ruiz-Sauri, A. Anatomy, Immunohistochemistry, and Numerical Distribution of Human Splenic Microvessels. Ann. Anat. 2019, 224, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Horioka, K.; Tanaka, H.; Isozaki, S.; Konishi, H.; Addo, L.; Takauji, S.; Druid, H. Low Temperature Induces Von-Willebrand Factor Expression via Increased Early Growth Response 1 Transcriptional Activity in Splenic Sinusoidal Endothelial Cells: Activation of the Splenic Vascular Endothelium in Low Temperatures. Biochem. Biophys. Res. Commun. 2020, 526, 239–245. [Google Scholar] [CrossRef]
- Williams, I.M.; Wu, J.C. Generation of Endothelial Cells from Human Pluripotent Stem Cells Methods, Considerations, and Applications. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 1317–1329. [Google Scholar] [CrossRef]
- Jayaraman, A.; Kumar, P.; Marin, S.; De Atauri, P.; Mateo, F.; Thomson, T.M.; Centelles, J.J.; Graham, S.F.; Cascante, M. Untargeted Metabolomics Reveals Distinct Metabolic Reprogramming in Endothelial Cells Co-Cultured with CSC and Non-CSC Prostate Cancer Cell Subpopulations. PLoS ONE 2018, 13, e0192175. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Wang, J.; Zhao, W.; Peng, Z.; Liu, X.; Li, B.; Zhang, H.; Shan, B.; Zhang, C.; Duan, C. Vasculogenic Mimicry in Carcinogenesis and Clinical Applications. J. Hematol. Oncol. 2020, 13, 19. [Google Scholar] [CrossRef] [PubMed]
- Lugano, R.; Ramachandran, M.; Dimberg, A. Tumor Angiogenesis: Causes, Consequences, Challenges and Opportunities. Cell. Mol. Life Sci. 2020, 77, 1745–1770. [Google Scholar] [CrossRef]
- Mentzer, S.J.; Konerding, M.A. Intussusceptive Angiogenesis: Expansion and Remodeling of Microvascular Networks. Angiogenesis 2014, 17, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Kuczynski, E.A.; Reynolds, A.R. Vessel Co-Option and Resistance to Anti-Angiogenic Therapy. Angiogenesis 2020, 23, 55–74. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Won, Y.J.; Shim, J.H.; Kim, H.J.; Kim, J.; Hong, H.N.; Kim, B.S. Morphological Characteristics of Vasculogenic Mimicry and Its Correlation with EphA2 Expression in Gastric Adenocarcinoma. Sci. Rep. 2019, 9, 3414. [Google Scholar] [CrossRef]
- Chopra, H.; Hung, M.K.; Kwong, D.L.; Zhang, C.F.; Pow, E.H.N. Insights into Endothelial Progenitor Cells: Origin, Classification, Potentials, and Prospects. Stem Cells Int. 2018, 2018, 9847015. [Google Scholar] [CrossRef] [PubMed]
- Carmeliet, P.; Jain, R.K. Angiogenesis in Cancer and Other Diseases. Nature 2000, 407, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Bertolini, F.; Shaked, Y.; Mancuso, P.; Kerbel, R.S. The Multifaceted Circulating Endothelial Cell in Cancer: Towards Marker and Target Identification. Nat. Rev. Cancer 2006, 6, 835–845. [Google Scholar] [CrossRef]
- Werb, Z.; Lu, P. The Role of Stroma in Tumor Development. Cancer J. 2015, 21, 250–253. [Google Scholar] [CrossRef]
- Lin, P.P. Aneuploid Circulating Tumor-Derived Endothelial Cell (CTEC): A Novel Versatile Player in Tumor Neovascularization and Cancer Metastasis. Cells 2020, 9, 1539. [Google Scholar] [CrossRef]
- Dvorak, A.M.; Feng, D. The Vesiculo-Vacuolar Organelle (VVO). A New Endothelial Cell Permeability Organelle. J. Histochem. Cytochem. 2001, 49, 419–431. [Google Scholar] [CrossRef] [PubMed]
- Bacci, M.; Ippolito, L.; Magnelli, L.; Giannoni, E.; Chiarugi, P. Stromal-Induced Mitochondrial Re-Education: Impact on Epithelial-to-Mesenchymal Transition and Cancer Aggressiveness. Semin. Cell Dev. Biol. 2020, 98, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; Weinberg, R.A. The Basics of Epithelial-Mesenchymal Transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef] [PubMed]
- Hapke, R.Y.; Haake, S.M. Hypoxia-Induced Epithelial to Mesenchymal Transition in Cancer. Cancer Lett. 2020, 487, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Hua, W.; ten Dijke, P.; Kostidis, S.; Giera, M.; Hornsveld, M. TGFβ-Induced Metabolic Reprogramming during Epithelial-to-Mesenchymal Transition in Cancer. Cell. Mol. Life Sci. 2020, 77, 2103–2123. [Google Scholar] [CrossRef]
- Yang, F.; Ma, J.; Wan, J.; Ha, W.; Fang, C.; Lu, H.; Zhang, W. Epithelial-Mesenchymal Transition of Circulating Tumor Cells in Prostate Cancer Is Promoted by Survivin. J. Int. Med. Res. 2020, 48, 0300060519892395. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Guo, Y.; Hu, Y.; Sun, Y.; Xu, D. Endosulfan Triggers Epithelial-Mesenchymal Transition via PTP4A3-Mediated TGF-β Signaling Pathway in Prostate Cancer Cells. Sci. Total Environ. 2020, 731, 139234. [Google Scholar] [CrossRef] [PubMed]
- Sung, N.J.; Kim, N.H.; Bae, N.Y.; Jo, H.S.; Park, S.A. DHA Inhibits Gremlin-1-Induced Epithelial-to-Mesenchymal Transition via ERK Suppression in Human Breast Cancer Cells. Biosci. Rep. 2020, 40, BSR20200164. [Google Scholar] [CrossRef]
- Choi, H.; Moon, A. Cross-talk between Cancer Cells and Endothelial Cells: Implications for Tumor Progression and Intervention. Arch. Pharm. Res. 2018, 41, 711–724. [Google Scholar] [CrossRef]
- Zhao, R.; Bei, X.; Yang, B.; Wang, X.; Jiang, C.; Shi, F.; Wang, X.; Zhu, Y.; Jing, Y.; Han, B.; et al. Endothelial Cells Promote Metastasis of Prostate Cancer by Enhancing Autophagy. J. Exp. Clin. Cancer Res. 2018, 37, 221. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, A.; Costa-Silva, B.; Shen, T.L.; Rodrigues, G.; Hashimoto, A.; Tesic Mark, M.; Molina, H.; Kohsaka, S.; Di Giannatale, A.; Ceder, S.; et al. Tumour Exosome Integrins Determine Organotropic Metastasis. Nature 2015, 527, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Woywodt, A.; Bahlmann, F.H.; De Groot, K.; Haller, H.; Haubitz, M. Circulating Endothelial Cells: Life, Death, Detachment and Repair of the Endothelial Cell Layer. Nephrol. Dial. Transplant. 2002, 17, 1728–1730. [Google Scholar] [CrossRef] [PubMed]
- Davies, P.F.; Remuzzitt, A.; Gordon, E.J.; Dewey, C.F.; Gimbrone, M.A. Turbulent Fluid Shear Stress Induce Vascular Endothelial Cell Turnover in Vitrio. Proc. Natl. Acad. Sci. USA 1986, 83, 2114–2117. [Google Scholar] [CrossRef] [PubMed]
- Caplan, B.A.; Schwartz, C.J. Increased Endothelial Cell Turnover in Areas of in Vivo Evans Blue Uptake in the Pig Aorta. Atherosclerosis 1973, 17, 401–417. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, T.; Mesa-Tejada, R.; Quick, C.M.; Bollen, A.W.; Joshi, S.; Pile-Spellman, J.; Lawton, M.T.; Young, W.L. Evidence of Increased Endothelial Cell Turnover in Brain Arteriovenous Malformations. Neurosurgery 2001, 49, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Molina-Ortiz, P.; Orban, T.; Martin, M.; Habets, A.; Dequiedt, F.; Schurmans, S. Rasa3 Controls Turnover of Endothelial Cell Adhesion and Vascular Lumen Integrity by a Rap1-Dependent Mechanism. PLoS Genet 2018, 14, e1007195. [Google Scholar] [CrossRef]
- Berardi, D.E.; Tarbell, J.M. Stretch and Shear Interactions Affect Intercellular Junction Protein Expression and Turnover in Endothelial Cells. Cell. Mol. Bioeng. 2009, 2, 320–331. [Google Scholar] [CrossRef]
- Kostelnik, K.B.; Barker, A.; Schultz, C.; Mitchell, T.P.; Rajeeve, V.; White, I.J.; Aurrand-Lions, M.; Nourshargh, S.; Cutillas, P.; Nightingale, T.D. Dynamic Trafficking and Turnover of JAM-C Is Essential for Endothelial Cell Migration. PLoS Biol. 2019, 17, 320–331. [Google Scholar] [CrossRef]
- Gao, F.; Artham, S.; Sabbineni, H.; Al-Azayzih, A.; Peng, X.D.; Hay, N.; Adams, R.H.; Byzova, T.V.; Somanath, P.R. Akt1 Promotes Stimuli-Induced Endothelial-Barrier Protection through FoxO-Mediated Tight-Junction Protein Turnover. Cell. Mol. Life Sci. 2016, 73, 3917–3933. [Google Scholar] [CrossRef]
- Botros, L.; Pronk, M.C.A.; Juschten, J.; Liddle, J.; Morsing, S.K.H.; Van Buul, J.D.; Bates, R.H.; Tuinman, P.R.; Van Bezu, J.S.M.; Huveneers, S.; et al. Bosutinib Prevents Vascular Leakage by Reducing Focal Adhesion Turnover and Reinforcing Junctional Integrity. J. Cell Sci. 2020, 133, jcs240077. [Google Scholar] [CrossRef] [PubMed]
- Wylezinski, L.S.; Hawiger, J. Interleukin 2 Activates Brain Microvascular Endothelial Cells Resulting in Destabilization of Adherens Junctions. J. Biol. Chem. 2016, 291, 22913–22923. [Google Scholar] [CrossRef] [PubMed]
- Foteinos, G.; Hu, Y.; Xiao, Q.; Metzler, B.; Xu, Q. Rapid Endothelial Turnover in Atherosclerosis-Prone Areas Coincides with Stem Cell Repair in Apolipoprotein E-Deficient Mice. Circulation 2008, 117, 1856–1863. [Google Scholar] [CrossRef] [PubMed]
- Bogorad, M.I.; DeStefano, J.G.; Linville, R.M.; Wong, A.D.; Searson, P.C. Cerebrovascular Plasticity: Processes That Lead to Changes in the Architecture of Brain Microvessels. J. Cereb. Blood Flow Metab. 2019, 39, 1413–1432. [Google Scholar] [CrossRef] [PubMed]
- DeStefano, J.G.; Xu, Z.S.; Williams, A.J.; Yimam, N.; Searson, P.C. Effect of Shear Stress on IPSC-Derived Human Brain Microvascular Endothelial Cells (DhBMECs). Fluids Barriers CNS 2017, 14, 20. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, O.; Zdunek, S.; Felker, A.; Salehpour, M.; Alkass, K.; Bernard, S.; Sjostrom, S.L.; Szewczykowska, M.; Jackowska, T.; Dos Remedios, C.; et al. Dynamics of Cell Generation and Turnover in the Human Heart. Cell 2015, 161, 1566–1575. [Google Scholar] [CrossRef] [PubMed]
- Greif, D.M.; Eichmann, A. Basic Research: Where Do New Endothelial Cells Come from in the Injured Heart? Nat. Rev. Cardiol. 2017, 14, 507–508. [Google Scholar] [CrossRef] [PubMed]
- Bowden, D.H. Cell Turnover in the Lung. Am. Rev. Respir. Dis. 1983, 128, 46–48. [Google Scholar] [CrossRef]
- Chen, F.; Fine, A. Stem Cells in Lung Injury and Repair. Am. J. Pathol. 2016, 186, 2544–2550. [Google Scholar] [CrossRef]
- Alban, F.T.; Gyamfi, D.; van Golen, R.F.; Heger, M. Reactive Oxygen and Nitrogen Species and Liver Ischemia-Reperfusion Injury: Role of Lipoic Acid. In The Liver; Elsevier: Amsterdam, The Netherlands, 2018; pp. 109–119. ISBN 9780128039519. [Google Scholar]
- Van Golen, R.F.; Reiniers, M.J.; Vrisekoop, N.; Zuurbier, C.J.; Olthof, P.B.; van Rheenen, J.; van Gulik, T.M.; Parsons, B.J.; Heger, M. The Mechanisms and Physiological Relevance of Glycocalyx Degradation in Hepatic Ischemia/Reperfusion Injury. Antioxid Redox Signal. 2014, 21, 1098–1118. [Google Scholar] [CrossRef]
- Zhao, X.; Wu, N.; Huang, L. Endothelial Progenitor Cells and Spleen: New Insights in Regeneration Medicine. Cytotherapy 2010, 12, 7–16. [Google Scholar] [CrossRef]
- Crane, G.M.; Liu, Y.-C.; Chadburn, A. Spleen: Development, Anatomy and Reactive Lymphoid Proliferations. Semin. Diagn. Pathol. 2020, 38, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Basile, D.P.; Collett, J.A.; Yoder, M.C. Endothelial Colony-Forming Cells and pro-Angiogenic Cells: Clarifying Definitions and Their Potential Role in Mitigating Acute Kidney Injury. Acta Physiol. 2018, 222, e12914. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.J.M.; Lo, Y.H.; Mah, A.T.; Kuo, C.J. The Intestinal Stem Cell Niche: Homeostasis and Adaptations. Trends Cell Biol. 2018, 28, 1062–1078. [Google Scholar] [CrossRef] [PubMed]
- Shirali, A.S.; Romay, M.C.; McDonald, A.I.; Su, T.; Steel, M.E.; Iruela-Arispe, M.L. A Multi-Step Transcriptional Cascade Underlies Vascular Regeneration in Vivo. Sci. Rep. 2018, 8, 5430. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Wang, S.; He, Y.; Hu, H.; Yao, B.; Zhang, Y. Regulation of Bone Morphogenetic Protein 4 on Epithelial Tissue. J. Cell Commun. Signal. 2020, 14, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Subileau, M.; Merdzhanova, G.; Ciais, D.; Collin-Faure, V.; Feige, J.J.; Bailly, S.; Vittet, D. Bone Morphogenetic Protein 9 Regulates Early Lymphatic-Specified Endothelial Cell Expansion during Mouse Embryonic Stem Cell Differentiation. Stem Cell Rep. 2019, 12, 98–111. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.B.; Chen, K.; Zhang, Y.M.; Gong, T. Common Injuries and Repair Mechanisms in the Endothelial Lining. Chin. Med. J. 2018, 131, 2338–2345. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Zhou, T.; Sekhar Pilli, V.S.; Phan, N.; Wang, Q.; Gupta, K.; Liu, Z.; Sheibani, N.; Liu, B. Novel Paracrine Functions of Smooth Muscle Cells in Supporting Endothelial Regeneration Following Arterial Injury. Circ. Res. 2019, 124, 1253–1265. [Google Scholar] [CrossRef]
- O’Carroll, L.; Wardrop, B.; Murphy, R.P.; Ross, M.D.; Harrison, M. Circulating Angiogenic Cell Response to Sprint Interval and Continuous Exercise. Eur. J. Appl. Physiol. 2019, 119, 743–752. [Google Scholar] [CrossRef]
- D’Alessio, F.R.; Kurzhagen, J.T.; Rabb, H. Reparative T Lymphocytes in Organ Injury. J. Clin. Investig. 2019, 129, 2608–2618. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Song, J.; Wang, Y.; Chen, Z.; Zhang, L.; Yu, J.; Zhu, D.; Zhong, M. M2 Macrophages Promote Pulmonary Endothelial Cells Regeneration in Sepsis-Induced Acute Lung Injury. Ann. Transl. Med. 2019, 7, 142. [Google Scholar] [CrossRef]
- Osorio-Valencia, S.; Zhou, B. Roles of Macrophages and Endothelial Cells and Their Cross-talk in Acute Lung Injury. Biomedicines 2024, 12, 632. [Google Scholar] [CrossRef] [PubMed]
- Guan, T.; Zhou, X.; Zhou, W.; Lin, H. Regulatory T Cell and Macrophage Cross-talk in Acute Lung Injury: Future Perspectives. Cell Death Discov. 2023, 9, 9. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F. Autophagy in Vascular Endothelial Cells. Clin. Exp. Pharmacol. Physiol. 2016, 43, 1021–1028. [Google Scholar] [CrossRef] [PubMed]
- Lenna, S.; Han, R.; Trojanowska, M. ER Stress and Endothelial Dysfunction. IUBMB Life 2014, 66, 530. [Google Scholar] [CrossRef] [PubMed]
- Kaplon, R.E.; Chung, E.; Reese, L.; Cox-York, K.; Seals, D.R.; Gentile, C.L. Activation of the Unfolded Protein Response in Vascular Endothelial Cells of Nondiabetic Obese Adults. J. Clin. Endocrinol. Metab. 2013, 98, E1505. [Google Scholar] [CrossRef]
- Nagane, M.; Yasui, H.; Kuppusamy, P.; Yamashita, T.; Inanami, O. DNA Damage Response in Vascular Endothelial Senescence: Implication for Radiation-Induced Cardiovascular Diseases. J. Radiat. Res. 2021, 62, 564–573. [Google Scholar] [CrossRef] [PubMed]
- Nikfarjam, S.; Singh, K.K. DNA Damage Response Signaling: A Common Link between Cancer and Cardiovascular Diseases. Cancer Med. 2023, 12, 4380–4404. [Google Scholar] [CrossRef]
- Kim, S.J.; Lee, W.J.; Cho, K.J. P62 Links the Autophagy Pathway and the Ubiquitin–Proteasome System in Endothelial Cells during Atherosclerosis. Int. J. Mol. Sci. 2021, 22, 7791. [Google Scholar] [CrossRef]
- Stangl, K.; Stangl, V. The Ubiquitin-Proteasome Pathway and Endothelial (Dys)Function. Cardiovasc. Res. 2010, 85, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Bhat, O.M.; Li, P.L. Lysosome Function in Cardiovascular Diseases. Cell Physiol. Biochem. 2021, 55, 277. [Google Scholar] [CrossRef]
- Lu, Y.; Nanayakkara, G.; Sun, Y.; Liu, L.; Xu, K.; Drummer, C.; Shao, Y.; Saaoud, F.; Choi, E.T.; Jiang, X.; et al. Procaspase-1 Patrolled to the Nucleus of Proatherogenic Lipid LPC-Activated Human Aortic Endothelial Cells Induces ROS Promoter CYP1B1 and Strong Inflammation. Redox Biol. 2021, 47, 102142. [Google Scholar] [CrossRef] [PubMed]
- Naito, H.; Iba, T.; Takakura, N. Mechanisms of New Blood-Vessel Formation and Proliferative Heterogeneity of Endothelial Cells. Int. Immunol. 2020, 32, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Jerka, D.; Bonowicz, K.; Piekarska, K.; Gokyer, S.; Derici, U.S.; Hindy, O.A.; Altunay, B.B.; Yazgan, I.; Steinbrink, K.; Kleszczyński, K.; et al. Unraveling Endothelial Cell Migration: Insights into Fundamental Forces, Inflammation, Biomaterial Applications, and Tissue Regeneration Strategies. ACS Appl. Bio Mater. 2024, 7, 2054–2069. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Tu, J.; Gao, Q.; Li, X.; Zhu, Z.; Jin, Y.; Zhang, Y.; Xie, J.; Zhu, P.; Zhao, B.; et al. WITHDRAWN: BBB Recovery after Stroke by Self-Replenishing E-Pericytes Transdifferentiated from Local Endothelial Cells. bioRxiv 2024, 2024.01.07.573171. [Google Scholar] [CrossRef]
- Kalies, K.; Knöpp, K.; Wurmbrand, L.; Korte, L.; Dutzmann, J.; Pilowski, C.; Koch, S.; Sedding, D. Isolation of Circulating Endothelial Cells Provides Tool to Determine Endothelial Cell Senescence in Blood Samples. Sci. Rep. 2024, 14, 4271. [Google Scholar] [CrossRef] [PubMed]
- Lallo, V.; Bracaglia, L.G. Influencing Endothelial Cells’ Roles in Inflammation and Wound Healing Through Nucleic Acid Delivery. Tissue Eng. Part A 2024, 30, 272–286. [Google Scholar] [CrossRef]
- Pinto, T.S.; Feltran, G.D.S.; Fernandes, C.J.D.C.; de Camargo Andrade, A.F.; Coque, A.D.C.; Silva, S.L.; Abuderman, A.A.; Zambuzzi, W.F.; Foganholi da Silva, R.A. Epigenetic Changes in Shear-Stressed Endothelial Cells. Cell Biol. Int. 2024, 48, 665–681. [Google Scholar] [CrossRef]
- Alvandi, Z.; Bischoff, J. Endothelial-Mesenchymal Transition in Cardiovascular Disease. Arterioscler. Thromb. Vasc. Biol. 2021, 41, 2357–2369. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).