

Transcriptomic Differences by RNA Sequencing for Evaluation of New Method for Long-Time In Vitro Culture of Cryopreserved Testicular Tissue for Oncologic Patients


 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
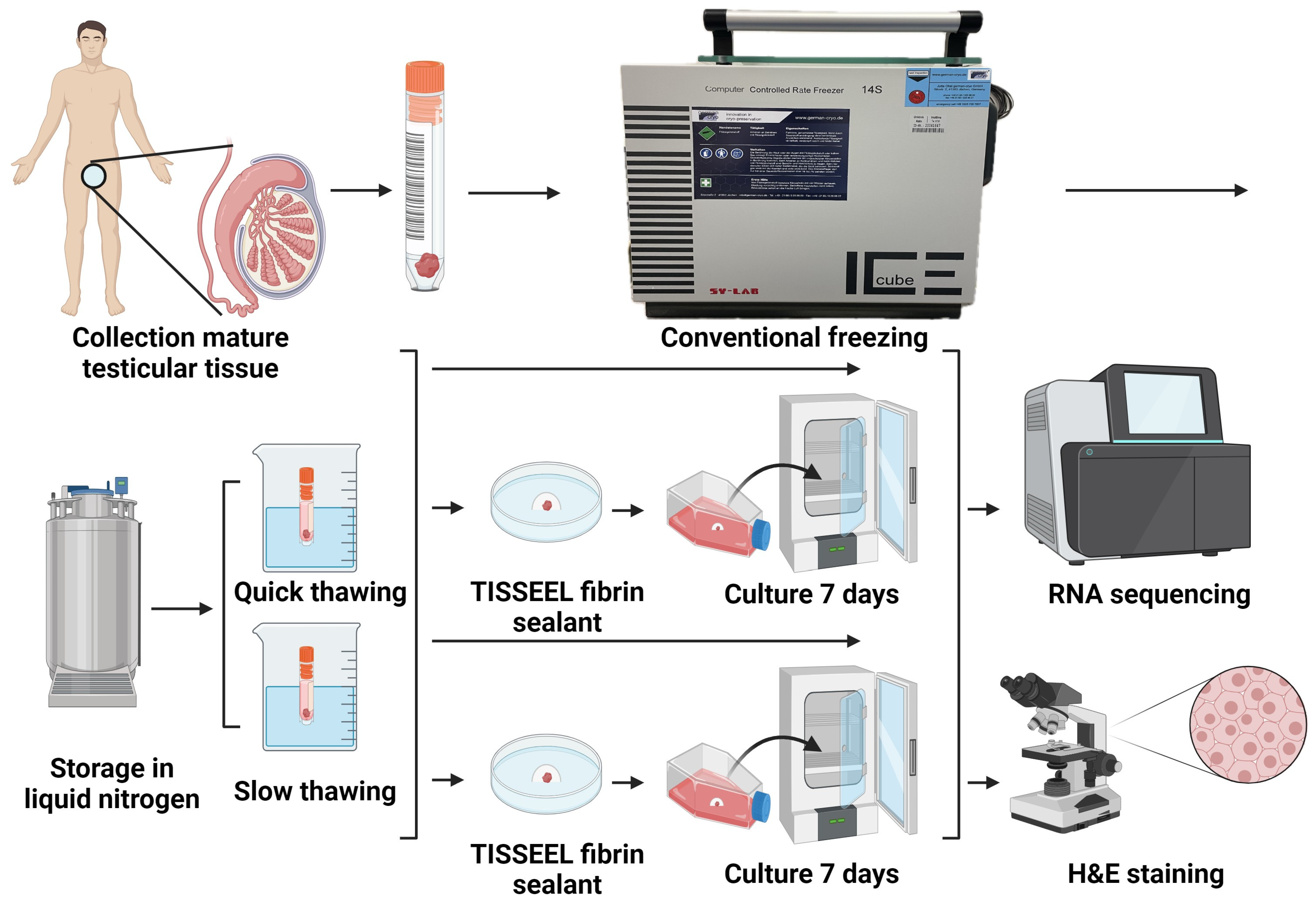
2.1. Design of Experiments
2.2. Collection of Samples
2.3. Cryopreservation (Freezing and Thawing)
2.4. 3D In Vitro Culture
2.5. Histology
2.6. Data Extraction and Sequencing
3. Results
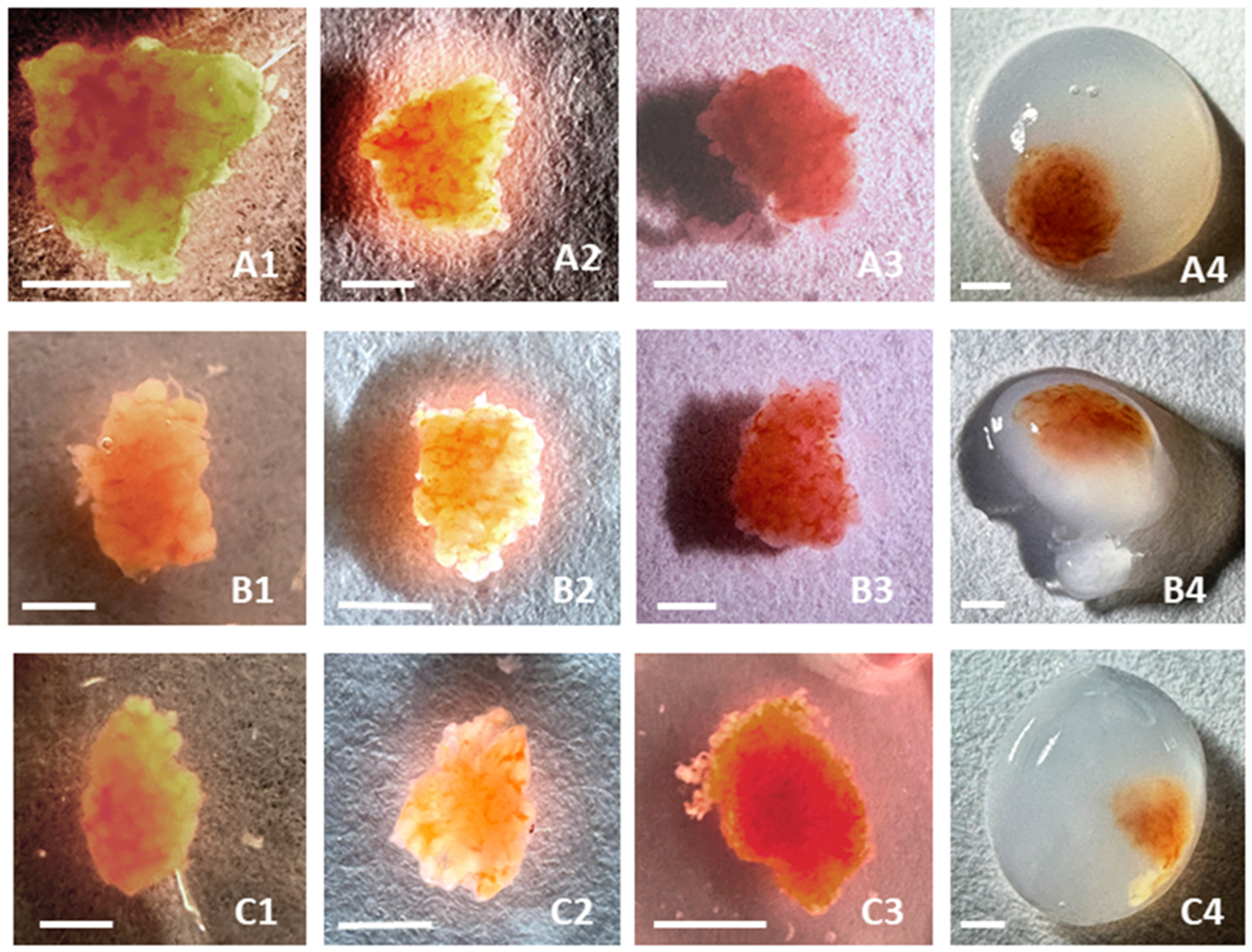
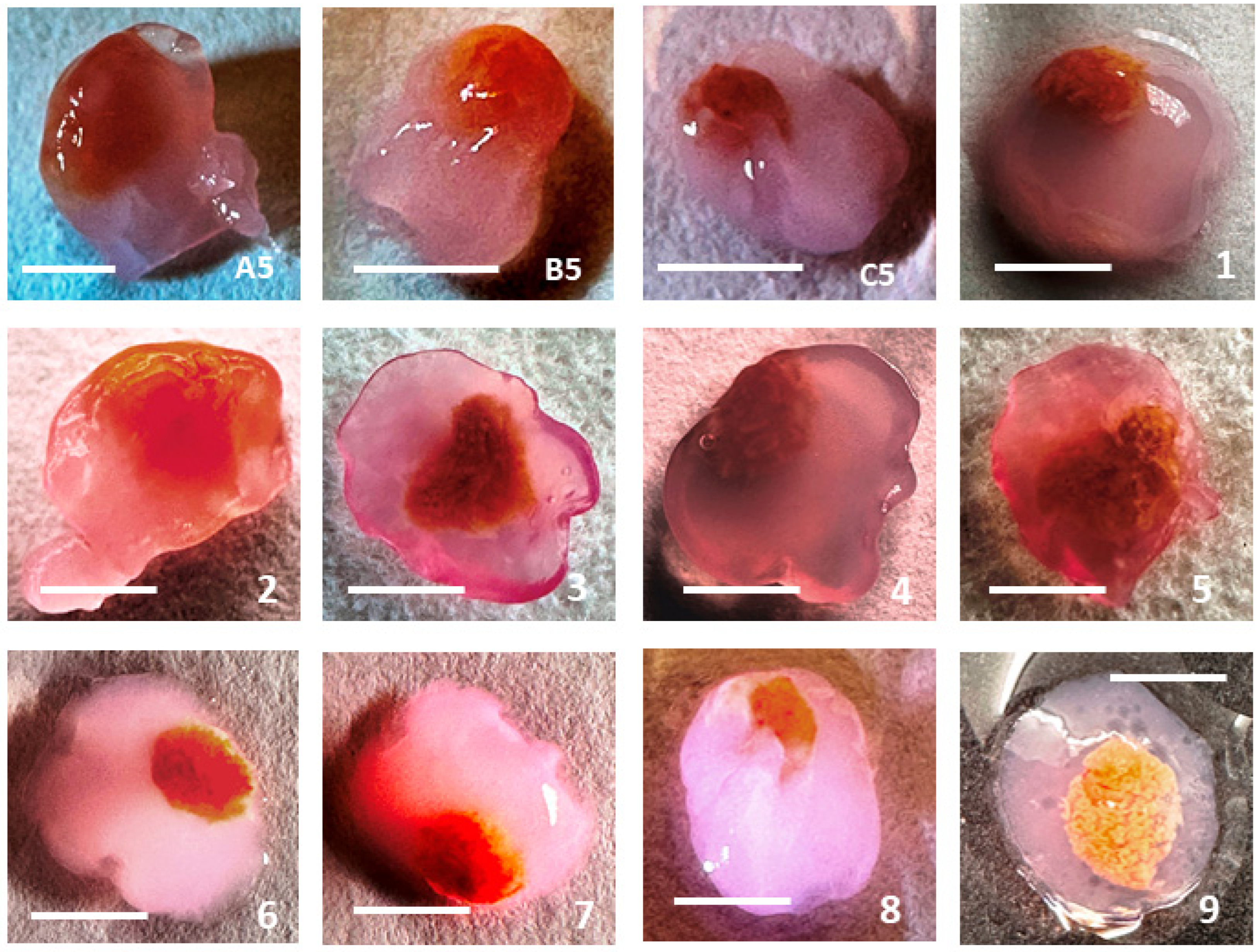
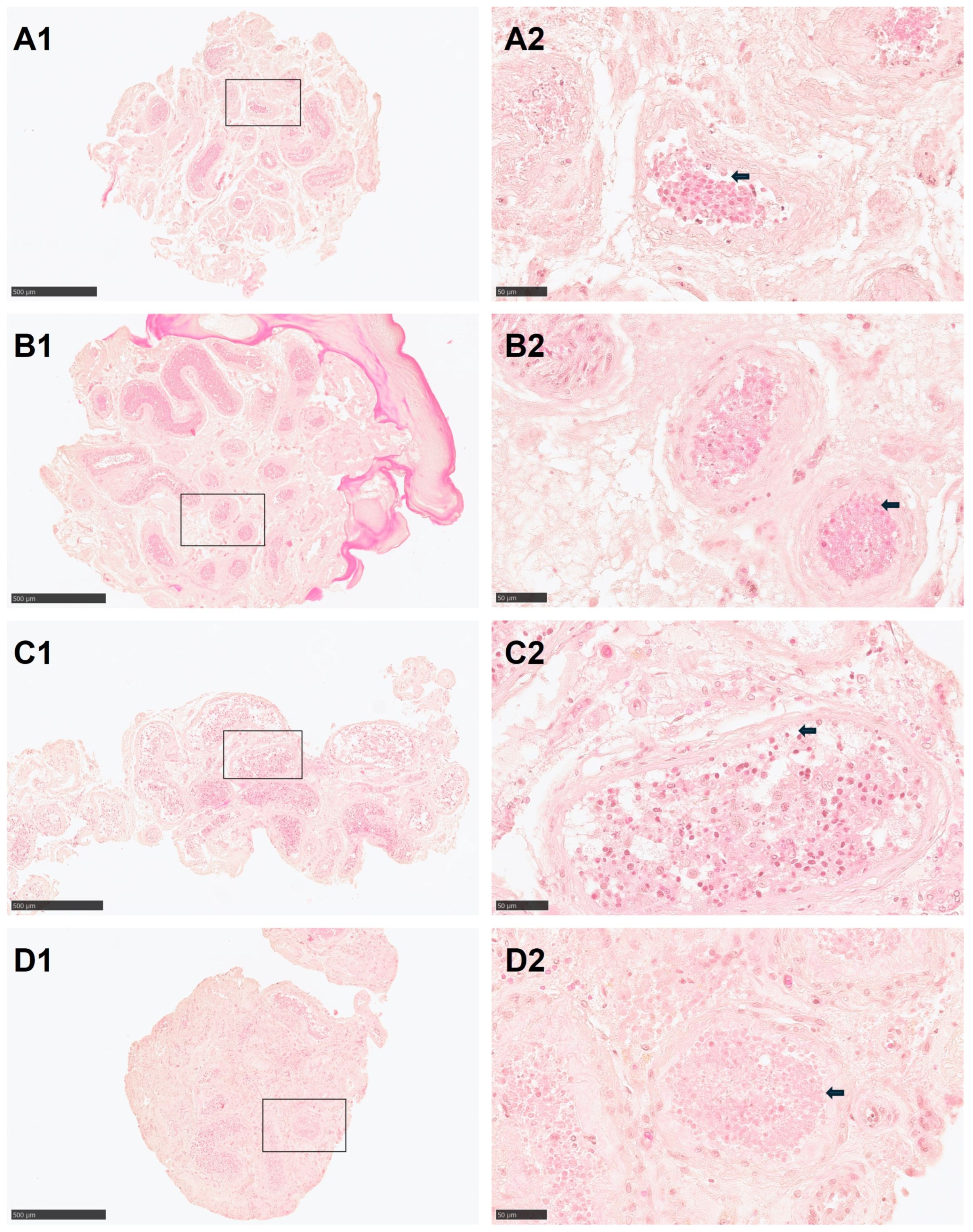
3.1. Morphology
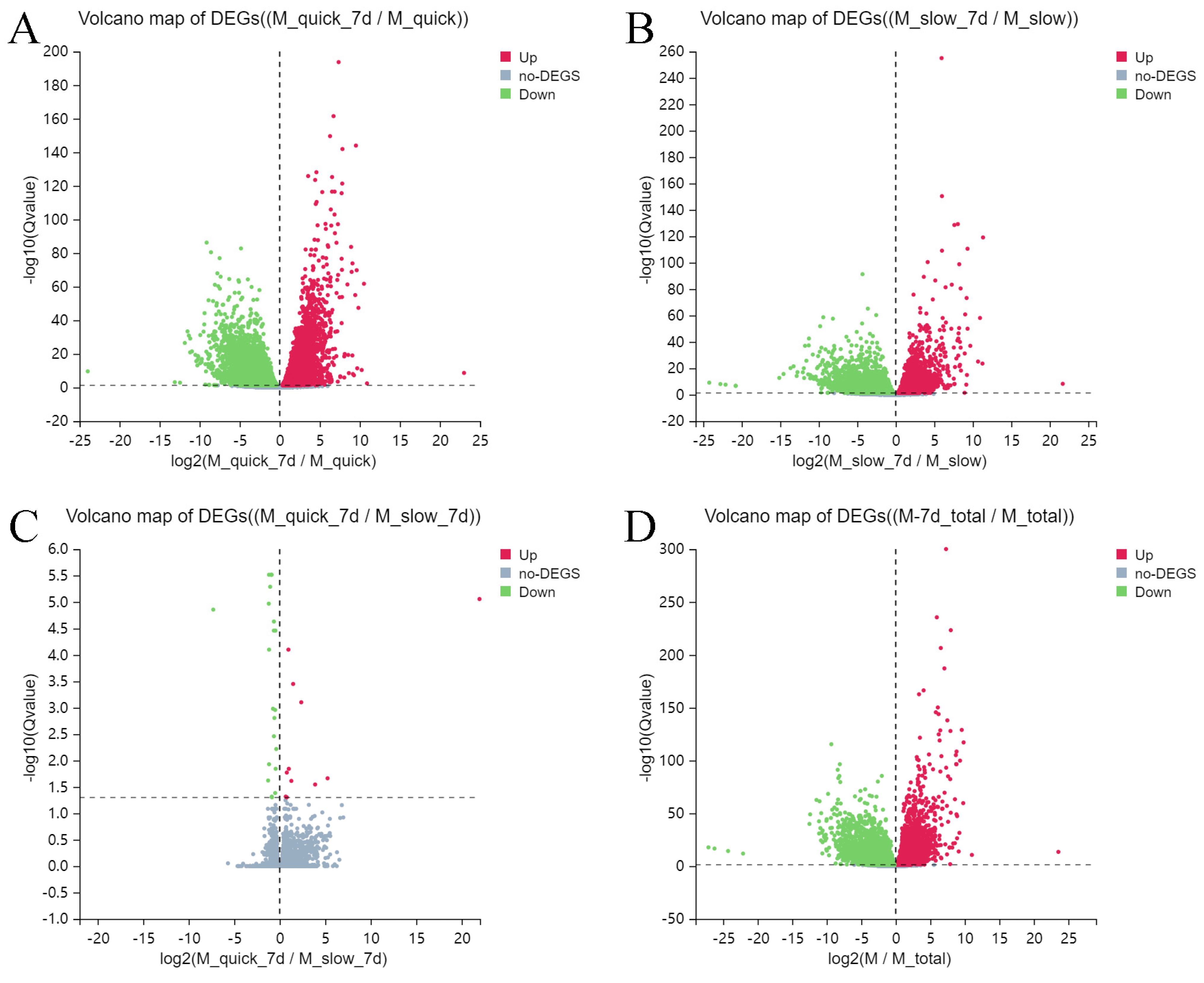
3.2. Differentially Expressed Genes
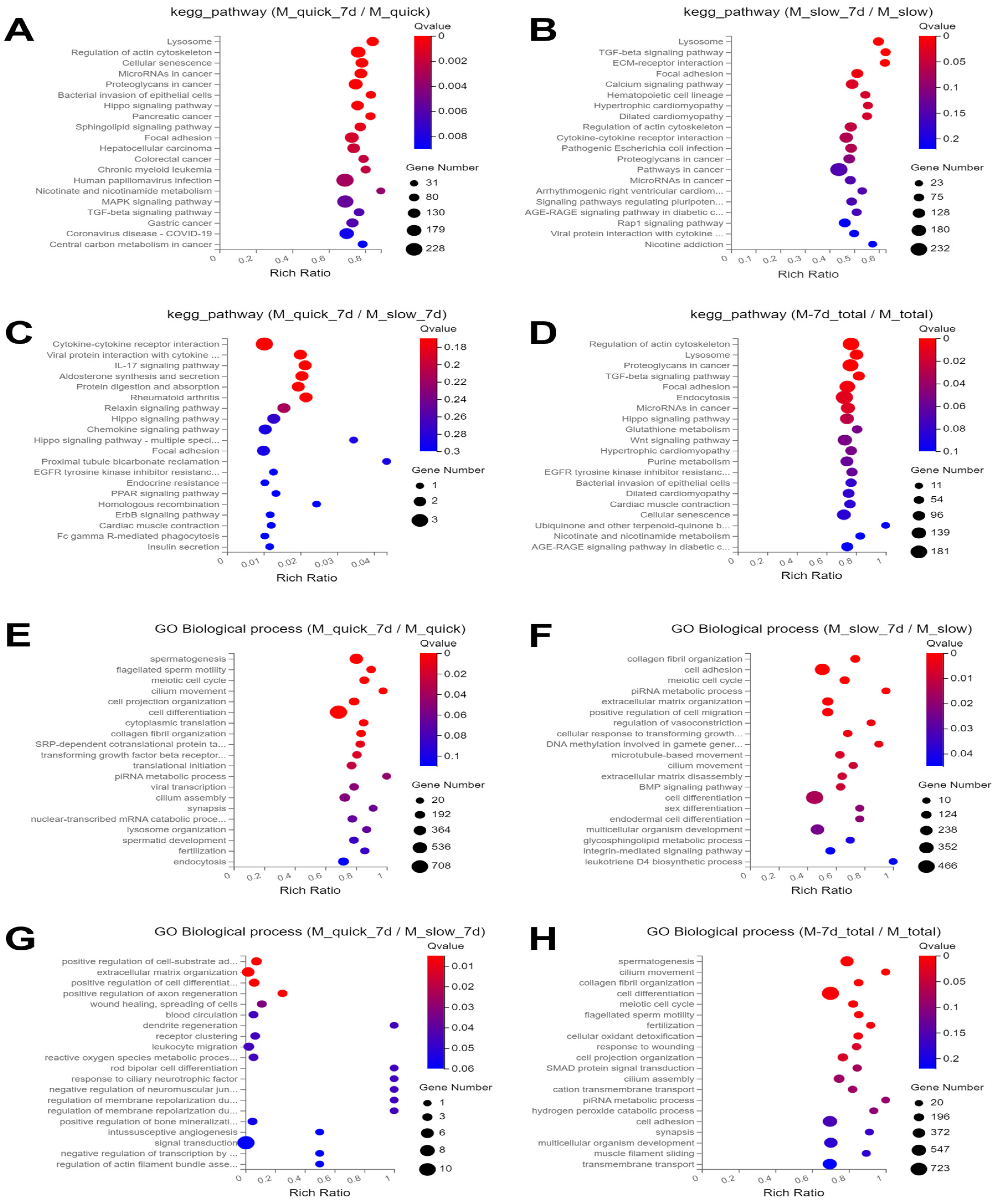
3.3. Enrichment Analysis of KEGG Pathways
3.4. GO Biological Process Enrichment Analysis
4. Discussion
4.1. Cryopreservation (Freezing and Thawing)
4.2. 3D In Vitro Culture
4.3. RNA Sequencing
4.3.1. Differential Expressed Genes
4.3.2. KEGG Pathways Analysis
4.3.3. GO Terms Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Trapphoff, T.; Dieterle, S. Cryopreservation of Ovarian and Testicular Tissue and the Influence on Epigenetic Pattern. Int. J. Mol. Sci. 2023, 24, 11061. [Google Scholar] [CrossRef] [PubMed]
- Onofre, J.; Kadam, P.; Baert, Y.; Goossens, E. Testicular tissue cryopreservation is the preferred method to preserve spermatogonial stem cells prior to transplantation. Reprod. BioMed. Online 2020, 40, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Patra, T.; Pathak, D.; Gupta, M.K. Strategies for cryopreservation of testicular cells and tissues in cancer and genetic diseases. Cell Tissue Res. 2021, 385, 1–19. [Google Scholar] [CrossRef]
- Hashimoto, K.; Odaka, H.; Ishikawa-Yamauchi, Y.; Nagata, S.; Nakamura, H.; Kimura, H.; Sato, T.; Makiyama, K.; Ogawa, T. Culture-space control is effective in promoting haploid cell formation and spermiogenesis in vitro in neonatal mice. Sci. Rep. 2023, 13, 12354. [Google Scholar] [CrossRef] [PubMed]
- Ibtisham, F.; Cham, T.C.; Fayaz, M.A.; Honaramooz, A. Effects of Growth Factors on In Vitro Culture of Neonatal Piglet Testicular Tissue Fragments. Cells 2023, 12, 2234. [Google Scholar] [CrossRef] [PubMed]
- Younis, N.; Caldeira-Brant, A.L.; Chu, T.; Abdalla, S.; Orwig, K.E. Human immature testicular tissue organ culture: A step towards fertility preservation and restoration. Front. Endocrinol. 2023, 14, 1242263. [Google Scholar] [CrossRef]
- Tan, J.; Li, J.; Zhang, Y.; Li, X.; Han, S.; Li, Z.; Zhou, X. Application of photocrosslinked gelatin, alginate and dextran hydrogels in the in vitro culture of testicular tissue. Int. J. Biol. Macromol. 2024, 260, 129498. [Google Scholar] [CrossRef]
- Önen, S.; Atik, A.C.; Gizer, M.; Köse, S.; Yaman, Ö.; Külah, H.; Korkusuz, P. A pumpless monolayer microfluidic device based on mesenchymal stem cell-conditioned medium promotes neonatal mouse in vitro spermatogenesis. Stem Cell Res. Ther. 2023, 14, 127. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, W.; Todorov, P.; Pei, C.; Isachenko, E.; Rahimi, G.; Mallmann, P.; Nawroth, F.; Isachenko, V. RNA Transcripts in Human Ovarian Cells: Two-Time Cryopreservation Does Not Affect Developmental Potential. Int. J. Mol. Sci. 2023, 24, 6880. [Google Scholar] [CrossRef]
- Dumont, L.; Lopez Maestre, H.; Chalmel, F.; Huber, L.; Rives-Feraille, A.; Moutard, L.; Bateux, F.; Rondanino, C.; Rives, N. Throughout in vitro first spermatogenic wave: Next-generation sequencing gene expression patterns of fresh and cryopreserved prepubertal mice testicular tissue explants. Front. Endocrinol. 2023, 14, 1112834. [Google Scholar] [CrossRef]
- Wang, W.; Todorov, P.; Isachenko, E.; Rahimi, G.; Merzenich, M.; Mallmann-Gottschalk, N.; Zhou, Y.; Yao, J.; Li, X.; Isachenko, V. Assessment of molecular and morphological dynamics during long-time in vitro cultivation of cryopreserved human ovarian tissue: Risk of genetic alterations. Front. Endocrinol. 2024, in press. [Google Scholar]
- Isachenko, V.; Morgenstern, B.; Todorov, P.; Isachenko, E.; Mallmann, P.; Hanstein, B.; Rahimi, G. Patient with Ovarian Insufficiency: Baby Born after Anticancer Therapy and Re-Transplantation of Cryopreserved Ovarian Tissue. J. Ovarian Res. 2020, 13, 118. [Google Scholar] [CrossRef] [PubMed]
- Isachenko, V.; Todorov, P.; Isachenko, E.; Rahimi, G.; Hanstein, B.; Salama, M.; Mallmann, P.; Tchorbanov, A.; Hardiman, P.; Getreu, N.; et al. Cryopreservation and xenografting of human ovarian fragments: Medulla decreases the phosphatidylserine translocation rate. Reprod. Biol. Endocrinol. 2016, 14, 79. [Google Scholar] [CrossRef] [PubMed]
- Pei, C.; Todorov, P.; Cao, M.; Kong, Q.; Isachenko, E.; Rahimi, G.; Mallmann-Gottschalk, N.; Uribe, P.; Sanchez, R.; Isachenko, V. Comparative Transcriptomic Analyses for the Optimization of Thawing Regimes during Conventional Cryopreservation of Mature and Immature Human Testicular Tissue. Int. J. Mol. Sci. 2023, 25, 214. [Google Scholar] [CrossRef]
- Isachenko, V.; Lapidus, I.; Isachenko, E.; Krivokharchenko, A.; Kreienberg, R.; Woriedh, M.; Bader, M.; Weiss, J.M. Human ovarian tissue vitrification versus conventional freezing: Morphological, endocrinological, and molecular biological evaluation. Reproduction 2009, 138, 319–327. [Google Scholar] [CrossRef]
- Ramasamy, R.; Yagan, N.; Schlegel, P.N. Structural and functional changes to the testis after conventional versus microdissection testicular sperm extraction. Urology 2005, 65, 1190–1194. [Google Scholar] [CrossRef]
- Ishikawa, T. Surgical recovery of sperm in non-obstructive azoospermia. Asian J. Androl. 2012, 14, 109–115. [Google Scholar] [CrossRef]
- Schiewe, M.C.; Rothman, C.; Spitz, A.; Werthman, P.E.; Zeitlin, S.I.; Anderson, R.E. Validation-verification of a highly effective, practical human testicular tissue in vitro culture-cryopreservation procedure aimed to optimize pre-freeze and post-thaw motility. J. Assist. Reprod. Genet. 2016, 33, 519–528. [Google Scholar] [CrossRef]
- Amelkina, O.; Silva, A.M.D.; Silva, A.R.; Comizzoli, P. Transcriptome dynamics in developing testes of domestic cats and impact of age on tissue resilience to cryopreservation. BMC Genom. 2021, 22, 847. [Google Scholar] [CrossRef]
- Isachenko, V.; Montag, M.; Isachenko, E.; van der Ven, K.; Dorn, C.; Roesing, B.; Braun, F.; Sadek, F.; van der Ven, H. Effective method for in-vitro culture of cryopreserved human ovarian tissue. Reprod. BioMed. Online 2006, 13, 228–234. [Google Scholar] [CrossRef]
- Djureinovic, D.; Fagerberg, L.; Hallström, B.; Danielsson, A.; Lindskog, C.; Uhlén, M.; Pontén, F. The human testis-specific proteome defined by transcriptomics and antibody-based profiling. Mol. Hum. Reprod. 2014, 20, 476–488. [Google Scholar] [CrossRef] [PubMed]
- Ji, C.; Wang, Y.; Wei, X.; Zhang, X.; Cong, R.; Yao, L.; Qin, C.; Song, N. Potential of testis-derived circular RNAs in seminal plasma to predict the outcome of microdissection testicular sperm extraction in patients with idiopathic non-obstructive azoospermia. Hum. Reprod. 2021, 36, 2649–2660. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Lv, X.; Chen, T.; Chen, Q.; Tian, H.; Yang, C.; Guo, W.; Liu, C. Construction and external validation of a 5-gene random forest model to diagnose non-obstructive azoospermia based on the single-cell RNA sequencing of testicular tissue. Aging 2021, 13, 24219–24235. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Zhang, S.; Jiang, S.; Huang, Z.; Chen, X.; Guo, H.; Li, M.; Zheng, S. Evaluation of immune status in testis and macrophage polarization associated with testicular damage in patients with nonobstructive azoospermia. Am. J. Reprod. Immunol. 2021, 86, e13481. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Luo, C.; Hu, L.; Chen, X.; Chen, Y.; Fan, J.; Cheng, C.Y.; Sun, F. Unraveling epigenomic abnormality in azoospermic human males by WGBS, RNA-Seq, and transcriptome profiling analyses. J. Assist. Reprod. Genet. 2020, 37, 789–802. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, Y.; Wang, Z.; Wang, Y.; Luo, Y.; Sun, N.; Zheng, S.; Yan, W.; Xiao, X.; Liu, S.; et al. CHST15 gene germline mutation is associated with the development of familial myeloproliferative neoplasms and higher transformation risk. Cell Death Dis. 2022, 13, 586. [Google Scholar] [CrossRef]
- Yang, Y.; Chung, E.K.; Wu, Y.L.; Savelli, S.L.; Nagaraja, H.N.; Zhou, B.; Hebert, M.; Jones, K.N.; Shu, Y.; Kitzmiller, K.; et al. Gene copy-number variation and associated polymorphisms of complement component C4 in human systemic lupus erythematosus (SLE): Low copy number is a risk factor for and high copy number is a protective factor against SLE susceptibility in European Americans. Am. J. Hum. Genet. 2007, 80, 1037–1054. [Google Scholar]
- Pereira, K.M.; Faria, A.G.; Liphaus, B.L.; Jesus, A.A.; Silva, C.A.; Carneiro-Sampaio, M.; Andrade, L.E. Low C4, C4A and C4B gene copy numbers are stronger risk factors for juvenile-onset than for adult-onset systemic lupus erythematosus. Rheumatology 2016, 55, 869–873. [Google Scholar] [CrossRef]
- Bairam, A.F.; Rasool, M.I.; Alherz, F.A.; Abunnaja, M.S.; El Daibani, A.A.; Gohal, S.A.; Kurogi, K.; Sakakibara, Y.; Suiko, M.; Liu, M.C. Sulfation of catecholamines and serotonin by SULT1A3 allozymes. Biochem. Pharmacol. 2018, 151, 104–113. [Google Scholar]
- Butcher, N.J.; Horne, M.K.; Mellick, G.D.; Fowler, C.J.; Masters, C.L.; Minchin, R.F. Sulfotransferase 1A3/4 copy number variation is associated with neurodegenerative disease. Pharmacogenom. J. 2018, 18, 209–214. [Google Scholar]
- Nenadov, D.S.; Pogrmic-Majkic, K.; Tesic, B.; Kokai, D.; Fa Nedeljkovic, S.; Stanic, B.; Andric, N. Impact of In Vitro Long-Term Low-Level DEHP Exposure on Gene Expression Profile in Human Granulosa Cells. Cells 2022, 11, 2304. [Google Scholar] [CrossRef] [PubMed]
- Rong, C.; Weng, L.; Li, M.; Zhou, L.; Li, Y. Serum interleukin-38 and -41 levels as candidate biomarkers in male infertility. Immunol. Lett. 2023, 255, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Sadia, K.; Leonel Javeres, M.N.; Tahir, F.; Shah, S.T.A.; Habib, R.; Muneer, Z.; Hussain, S.; Nurulain, S.M. Acetylcholinesterase, pro-inflammatory cytokines, and association of ACHE SNP rs 17228602 with male infertility. PLoS ONE 2023, 18, e0282579. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Zhang, S.; Gu, Y.; Sun, X.; Luo, C.; Zhou, J.; Li, Z.; Lin, H.; Zhang, W. PM2.5 activates IL-17 signaling pathway in human nasal mucosa-derived fibroblasts. Int. Immunopharmacol. 2024, 128, 111484. [Google Scholar] [CrossRef]
- Pérez, C.V.; Pellizzari, E.H.; Cigorraga, S.B.; Galardo, M.N.; Naito, M.; Lustig, L.; Jacobo, P.V. IL17A impairs blood-testis barrier integrity and induces testicular inflammation. Cell Tissue Res. 2014, 358, 885–898. [Google Scholar] [CrossRef]
- Wu, P.; Yang, K.; Sun, Z.; Zhao, Y.; Manthari, R.K.; Wang, J.; Cao, J. Interleukin-17A knockout or self-recovery alleviated autoimmune reaction induced by fluoride in mouse testis. Sci. Total Environ. 2023, 884, 163616. [Google Scholar] [CrossRef]
- Zhang, Y.; Hou, B.; Liu, T.; Wu, Y.; Wang, Z. Probiotics improve polystyrene microplastics-induced male reproductive toxicity in mice by alleviating inflammatory response. Ecotoxicol. Environ. Saf. 2023, 263, 115248. [Google Scholar] [CrossRef]
- Trivedi, P.C.; Bartlett, J.J.; Pulinilkunnil, T. Lysosomal Biology and Function: Modern View of Cellular Debris Bin. Cells 2020, 9, 1131. [Google Scholar] [CrossRef]
- Xu, H.; Kongmanas, K.; Kadunganattil, S.; Smith, C.E.; Rupar, T.; Goto-Inoue, N.; Hermo, L.; Faull, K.F.; Tanphaichitr, N. Arylsulfatase A deficiency causes seminolipid accumulation and a lysosomal storage disorder in Sertoli cells. J. Lipid Res. 2011, 52, 2187–2197. [Google Scholar] [CrossRef]
- Ali, W.; Deng, K.; Sun, J.; Ma, Y.; Liu, Z.; Zou, H. A new insight of cadmium-induced cellular evidence of autophagic-associated spermiophagy during spermatogenesis. Environ. Sci. Pollut. Res. Int. 2023, 30, 101064–101074. [Google Scholar] [CrossRef]
- Su, W.; Mruk, D.D.; Cheng, C.Y. Regulation of actin dynamics and protein trafficking during spermatogenesis--insights into a complex process. Crit. Rev. Biochem. Mol. Biol. 2013, 48, 153–172. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.H.; Chou, C.H.; Wu, J.C.; Liao, K.M.; Luo, W.J.; Hsu, W.L.; Chen, X.R.; Yu, S.L.; Pan, S.H.; Yang, P.C.; et al. LCRMP-1 is required for spermatogenesis and stabilises spermatid F-actin organization via the PI3K-Akt pathway. Commun. Biol. 2023, 6, 389. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yao, M.; Zeng, J.; Zheng, D.; Li, Q.; Ni, Y.; Xiao, X. FYN regulates cell adhesion at the blood-testis barrier and the apical ectoplasmic specialization via its effect on Arp3 in the mouse testis. Front. Immunol. 2022, 13, 915274. [Google Scholar] [CrossRef] [PubMed]
- Beardsley, A.; O’Donnell, L. Characterization of normal spermiation and spermiation failure induced by hormone suppression in adult rats. Biol. Reprod. 2003, 68, 1299–1307. [Google Scholar] [CrossRef]
- Beardsley, A.; Robertson, D.M.; O’Donnell, L. A complex containing alpha6beta1-integrin and phosphorylated focal adhesion kinase between Sertoli cells and elongated spermatids during spermatid release from the seminiferous epithelium. J. Endocrinol. 2006, 190, 759–770. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pei, C.; Todorov, P.; Kong, Q.; Cao, M.; Isachenko, E.; Rahimi, G.; Nawroth, F.; Mallmann-Gottschalk, N.; Liu, W.; Isachenko, V. Transcriptomic Differences by RNA Sequencing for Evaluation of New Method for Long-Time In Vitro Culture of Cryopreserved Testicular Tissue for Oncologic Patients. Cells 2024, 13, 1539. https://doi.org/10.3390/cells13181539
Pei C, Todorov P, Kong Q, Cao M, Isachenko E, Rahimi G, Nawroth F, Mallmann-Gottschalk N, Liu W, Isachenko V. Transcriptomic Differences by RNA Sequencing for Evaluation of New Method for Long-Time In Vitro Culture of Cryopreserved Testicular Tissue for Oncologic Patients. Cells. 2024; 13(18):1539. https://doi.org/10.3390/cells13181539
Chicago/Turabian StylePei, Cheng, Plamen Todorov, Qingduo Kong, Mengyang Cao, Evgenia Isachenko, Gohar Rahimi, Frank Nawroth, Nina Mallmann-Gottschalk, Wensheng Liu, and Volodimir Isachenko. 2024. "Transcriptomic Differences by RNA Sequencing for Evaluation of New Method for Long-Time In Vitro Culture of Cryopreserved Testicular Tissue for Oncologic Patients" Cells 13, no. 18: 1539. https://doi.org/10.3390/cells13181539