Equine Penile Squamous Cell Carcinomas as a Model for Human Disease: A Preliminary Investigation on Tumor Immune Microenvironment

 ,
,  ,
, 
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Case Selection
- histological diagnosis of equine papilloma, carcinoma in situ, and squamous cell carcinoma, assessed by two boarded pathologists (IP, CB), as per recently suggested diagnostic criteria [17];
- confirmed penile localization of the lesions; and,
- availability of >0.5 cm2 of FFPE tumor tissue evaluated on section.
2.2. Histopathological and Immunohistochemical Characterization of Tumors and of the Immune Infiltrate
2.3. DNA Extraction and Evaluation of EcPV-2 Viral Load
2.4. RNA Extraction and Evaluation of Gene Expression
2.5. Statistical Analysis
3. Results
3.1. Case Selection, Histological Characterization of Tumors and of the Immune Infiltrate and Ki-67 Index
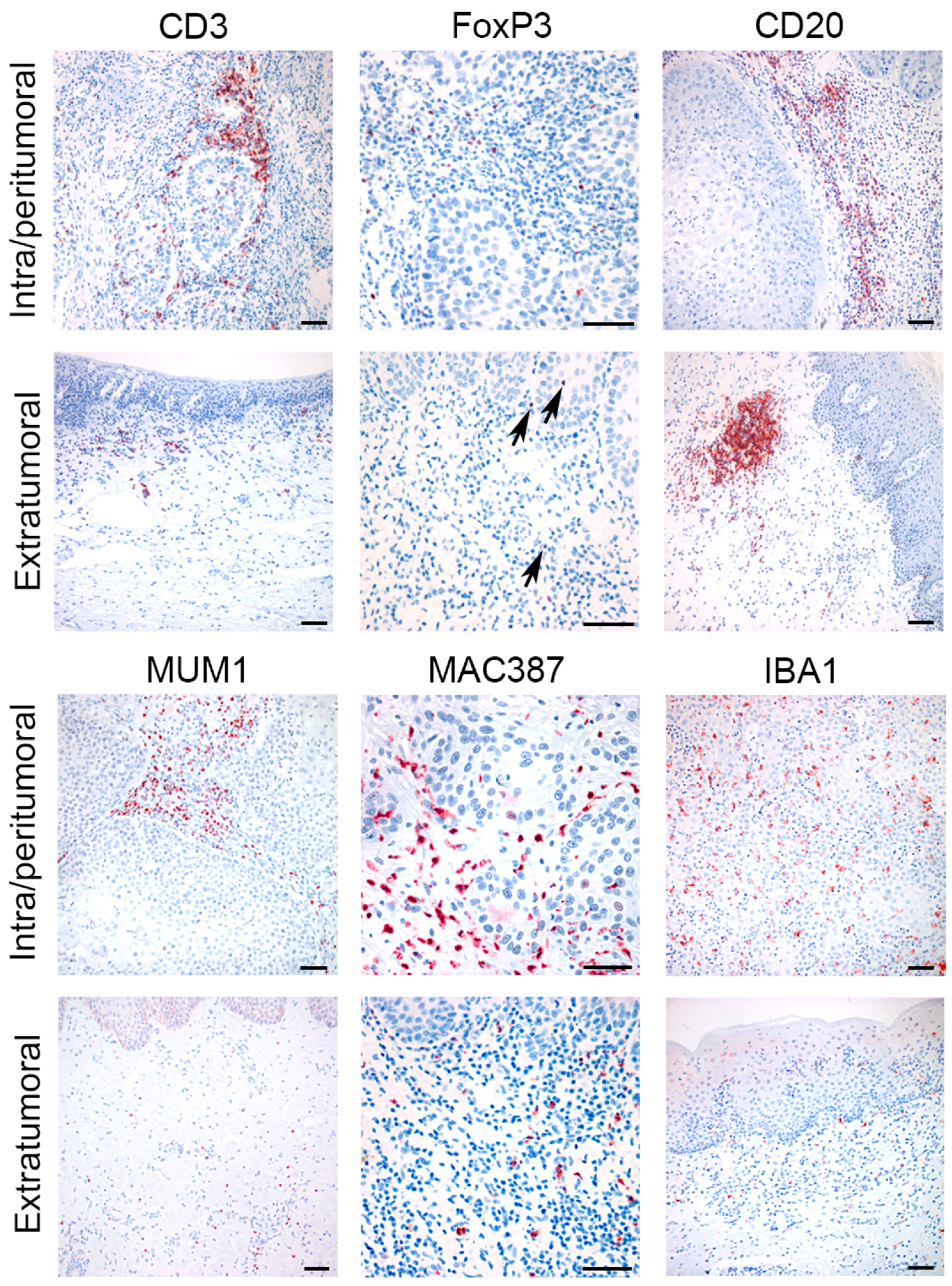
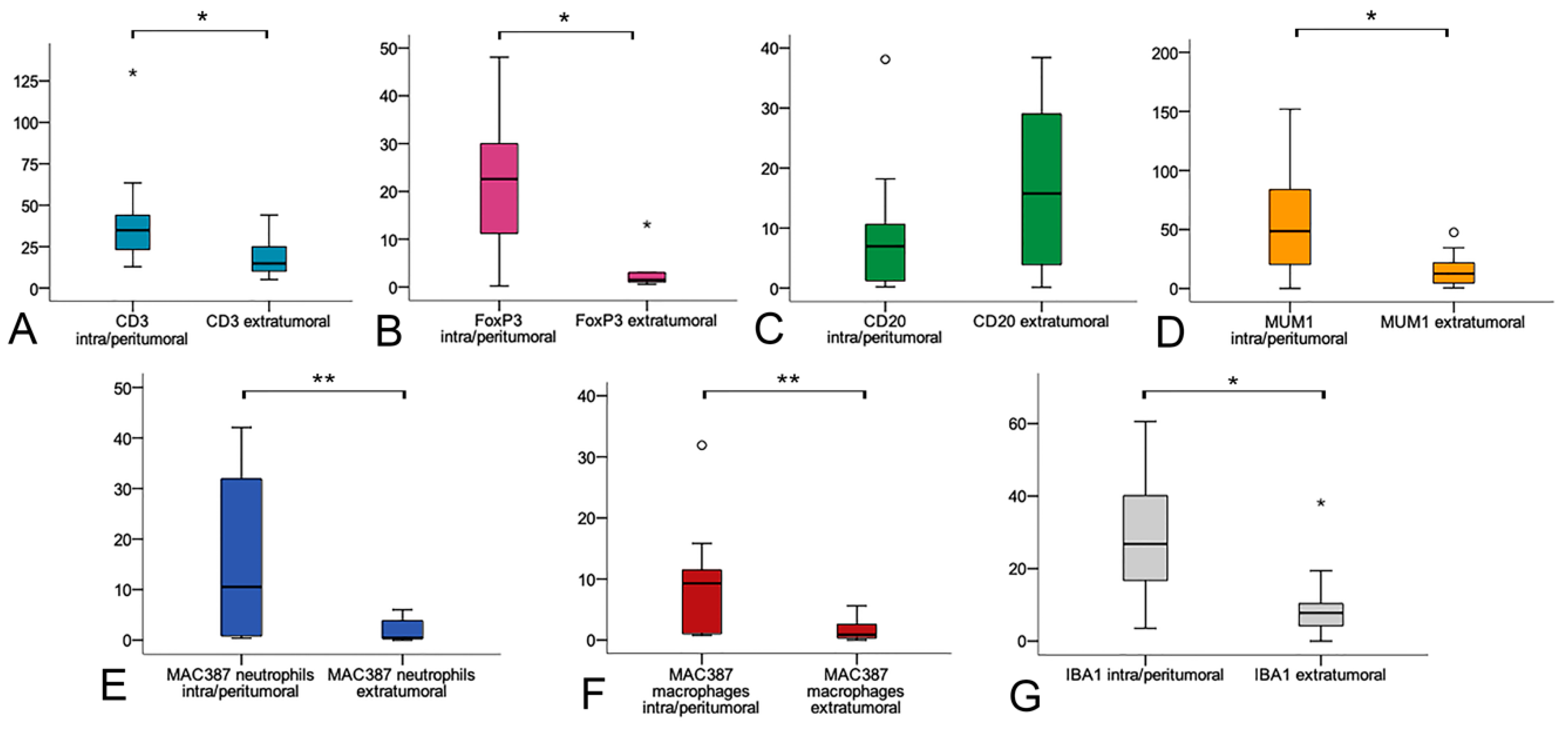
3.2. Immunohistochemical Characterization of the Intra/Peritumoral and Extratumoral Immune Infiltrate
3.3. Relationship between Immunohistochemical Expression of Markers of Immune Infiltrate, Histopathological Diagnosis and Features, and EcPV-2 Viral Load
3.4. EcPV-2 Detection and EcPV-2-L1 Gene Expression
3.5. Relationship between Immunohistochemical Markers of Immune Infiltrate and RT-qPCR Expression of Cytokines
3.6. Relationship between Immunohistochemical Markers of Immune Infiltrate and rt-qPCR Expression of Cytokines
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- IARC CancerBases / Cancer Incidence in Five Continents, Vol. XI. Available online: https://publications.iarc.fr/Databases/Iarc-Cancerbases/Cancer-Incidence-In-Five-Continents-Vol.-XI-2017 (accessed on 27 October 2020).
- Guan, P.; Howell-Jones, R.; Li, N.; Bruni, L.; De Sanjosé, S.; Franceschi, S.; Clifford, G.M. Human papillomavirus types in 115,789 HPV-positive women: A meta-analysis from cervical infection to cancer. Int. J. Cancer 2012, 131, 2349–2359. [Google Scholar] [CrossRef] [PubMed]
- Canete-Portillo, S.; Sanchez, D.F.; Cubilla, A.L. Pathology of Invasive and Intraepithelial Penile Neoplasia. Eur. Urol. Focus 2019, 5, 713–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alemany, L.; Cubilla, A.; Halec, G.; Kasamatsu, E.; Quirós, B.; Masferrer, E.; Tous, S.; Lloveras, B.; Hernández-Suarez, G.; Lonsdale, R.; et al. Role of Human Papillomavirus in Penile Carcinomas Worldwide. Eur. Urol. 2016, 69, 953–961. [Google Scholar] [CrossRef] [PubMed]
- Iorga, L.; Marcu, R.; Diaconu, C.; Stanescu, A.; Stoian, A.; Mischianu, D.; Surcel, M.; Bungau, S.; Constantin, T.; Boda, D.; et al. Penile carcinoma and HPV infection (Review). Exp. Med. 2019, 20, 91. [Google Scholar] [CrossRef] [Green Version]
- Djajadiningrat, R.S.; Jordanova, E.S.; Kroon, B.K.; Van Werkhoven, E.; De Jong, J.; Pronk, D.T.M.; Snijders, P.J.F.; Horenblas, S.; Heideman, D.A.M. Human papillomavirus prevalence in invasive penile cancer and association with clinical outcome. J. Urol. 2015, 193, 526–531. [Google Scholar] [CrossRef] [PubMed]
- Ottenhof, S.R.; Djajadiningrat, R.S.; Thygesen, H.H.; Jakobs, P.J.; Józwiak, K.; Heeren, A.M.; de Jong, J.; Sanders, J.; Horenblas, S.; Jordanova, E.S. The prognostic value of immune factors in the tumor microenvironment of penile squamous cell carcinoma. Front. Immunol. 2018, 9, 1253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doorbar, J. Host control of human papillomavirus infection and disease. Best Pr. Res. Clin. Obs. Gynaecol. 2018, 47, 27–41. [Google Scholar] [CrossRef] [Green Version]
- Béziat, V. Human genetic dissection of papillomavirus-driven diseases: New insight into their pathogenesis. Hum. Genet. 2020, 139, 919–939. [Google Scholar] [CrossRef]
- Rector, A.; Van Ranst, M. Animal papillomaviruses. Virology 2013, 445, 213–223. [Google Scholar] [CrossRef] [Green Version]
- Munday, J.S.; Thomson, N.A.; Luff, J.A. Papillomaviruses in dogs and cats. Vet. J. 2017, 225, 23–31. [Google Scholar] [CrossRef]
- Munday, J.S. Bovine and Human Papillomaviruses: A Comparative Review. Vet. Pathol. 2014, 51, 1063–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sykora, S.; Brandt, S. Papillomavirus infection and squamous cell carcinoma in horses. Vet. J. 2017, 223, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Lange, C.E.; Zollinger, S.; Tobler, K.; Ackermann, M.; Favrot, C. Clinically healthy skin of dogs is a potential reservoir for canine papillomaviruses. J. Clin. Microbiol. 2011, 49, 707–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doorbar, J.; Quint, W.; Banks, L.; Bravo, I.G.; Stoler, M.; Broker, T.R.; Stanley, M.A. The biology and life-cycle of human papillomaviruses. Vaccine 2012, 30, F55–F70. [Google Scholar] [CrossRef]
- Doorbar, J.; Egawa, N.; Griffin, H.; Kranjec, C.; Murakami, I. Human papillomavirus molecular biology and disease association. Rev. Med. Virol. 2015, 25, 2–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramsauer, A.S.; Wachoski-Dark, G.L.; Fraefel, C.; Tobler, K.; Brandt, S.; Knight, C.G.; Favrot, C.; Grest, P. Paving the way for more precise diagnosis of EcPV2-associated equine penile lesions. Bmc Vet. Res. 2019, 15, 356. [Google Scholar] [CrossRef] [Green Version]
- Arthurs, C.; Suarez-Bonnet, A.; Willis, C.; Xie, B.; Machulla, N.; Mair, T.S.; Cao, K.; Millar, M.; Thrasivoulou, C.; Priestnall, S.L.; et al. Equine penile squamous cell carcinoma: Expression of biomarker proteins and EcPV2. Sci. Rep. 2020, 10, 7863. [Google Scholar] [CrossRef]
- Van Den Top, J.G.B.; De Heer, N.; Klein, W.R.; Ensink, J.M. Penile and preputial squamous cell carcinoma in the horse: A retrospective study of treatment of 77 affected horses. Equine Vet. J. 2008, 40, 533–537. [Google Scholar] [CrossRef]
- Suárez-Bonnet, A.; Willis, C.; Pittaway, R.; Smith, K.; Mair, T.; Priestnall, S.L. Molecular carcinogenesis in equine penile cancer: A potential animal model for human penile cancer. Urol. Oncol. Semin. Orig. Investig. 2018, 36, 532.e9–532.e18. [Google Scholar] [CrossRef] [Green Version]
- Smola, S. Immunopathogenesis of HPV-Associated Cancers and Prospects for Immunotherapy. Viruses 2017, 9, 254. [Google Scholar] [CrossRef]
- Che, Y.; Yang, Y.; Suo, J.; An, Y.; Wang, X. Induction of systemic immune responses and reversion of immunosuppression in the tumor microenvironment by a therapeutic vaccine for cervical cancer. Cancer Immunol. Immunother. 2020. [Google Scholar] [CrossRef]
- Chu, C.; Yao, K.; Lu, J.; Zhang, Y.; Chen, K.; Lu, J.; Zhang, C.Z.; Cao, Y. Immunophenotypes Based on the Tumor Immune Microenvironment Allow for Unsupervised Penile Cancer Patient Stratification. Cancers 2020, 12, 1796. [Google Scholar] [CrossRef] [PubMed]
- Vassallo, J.; Rodrigues, A.F.F.; Campos, A.H.J.F.M.; Rocha, R.M.; da Cunha, I.W.; Zequi, S.C.; Guimarães, G.C.; da Fonseca, F.P.; Lopes, A.; Cubilla, A.; et al. Pathologic and imunohistochemical characterization of tumoral inflammatory cell infiltrate in invasive penile squamous cell carcinomas: Fox-P3 expression is an independent predictor of recurrence. Tumor Biol. 2015, 36, 2509–2516. [Google Scholar] [CrossRef]
- Pérez, J.; Mozos, E.; Martín, M.P.; Day, M.J. Immunohistochemical study of the inflammatory infiltrate associated with equine squamous cell carcinoma. J. Comp. Pathol. 1999, 121, 385–397. [Google Scholar] [CrossRef]
- Porcellato, I.; Modesto, P.; Cappelli, K.; Varello, K.; Peletto, S.; Brachelente, C.; Martini, I.; Mechelli, L.; Ferrari, A.; Ghelardi, A.; et al. Equus caballus papillomavirus type 2 (EcPV2) in co-occurring vulvar and gastric lesions of a pony. Res. Vet. Sci. 2020, 132, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Veyer, D.; Wack, M.; Grard, O.; Bonfils, P.; Hans, S.; Bélec, L.; Badoual, C.; Péré, H. HPV detection and genotyping of head and neck cancer biopsies by molecular testing with regard to the new oropharyngeal squamous cell carcinoma classification based on HPV status. Pathology 2019, 51, 421–425. [Google Scholar] [CrossRef] [PubMed]
- Miglio, A.; Cappelli, K.; Capomaccio, S.; Mecocci, S.; Silvestrelli, M.; Antognoni, M.T. Metabolic and biomolecular changes induced by incremental long-term training in young thoroughbred racehorses during first workout season. Animals 2020, 10, 317. [Google Scholar] [CrossRef] [Green Version]
- Cappelli, K.; Felicetti, M.; Capomaccio, S.; Nocelli, C.; Silvestrelli, M.; Verini-Supplizi, A. Effect of training status on immune defence related gene expression in Thoroughbred: Are genes ready for the sprint? Vet. J. 2013, 195, 373–376. [Google Scholar] [CrossRef]
- Crociati, M.; Capomaccio, S.; Mandara, M.T.; Stradaioli, G.; Sylla, L.; Monaci, M.; Cappelli, K. Different expression of Defensin-B gene in the endometrium of mares of different age during the breeding season. BMC Vet. Res. 2019, 15, 465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cappelli, K.; Amadori, M.; Mecocci, S.; Miglio, A.; Antognoni, M.T.; Razzuoli, E. Immune Response in Young Thoroughbred Racehorses under Training. Animals 2020, 10, 1809. [Google Scholar] [CrossRef]
- Razzuoli, E.; Villa, R.; Ferrari, A.; Amadori, M. A pig tonsil cell culture model for evaluating oral, low-dose IFN-α treatments. Vet. Immunol. Immunopathol. 2014, 160, 244–254. [Google Scholar] [CrossRef]
- Bosch, F.X.; Broker, T.R.; Forman, D.; Moscicki, A.-B.; Gillison, M.L.; Doorbar, J.; Stern, P.L.; Stanley, M.; Arbyn, M.; Poljak, M.; et al. Comprehensive Control of Human Papillomavirus Infections and Related Diseases. Vaccine 2013, 31, H1–H31. [Google Scholar] [CrossRef] [Green Version]
- Hibi, H.; Hatama, S.; Obata, A.; Shibahara, T.; Kadota, K. Laryngeal squamous cell carcinoma and papilloma associated with Equus caballus papillomavirus 2 in a horse. J. Vet. Med. Sci. 2019, 81, 1029–1033. [Google Scholar] [CrossRef] [Green Version]
- Alloway, E.; Linder, K.; May, S.; Rose, T.; DeLay, J.; Bender, S.; Tucker, A.; Luff, J. A Subset of Equine Gastric Squamous Cell Carcinomas Is Associated With Equus Caballus Papillomavirus–2 Infection. Vet. Pathol. 2020, 57, 427–431. [Google Scholar] [CrossRef]
- Muñoz, N.; Castellsagué, X.; de González, A.B.; Gissmann, L. Chapter 1: HPV in the etiology of human cancer. Vaccine 2006, 24. [Google Scholar] [CrossRef]
- Lange, C.E.; Tobler, K.; Lehner, A.; Grest, P.; Welle, M.M.; Schwarzwald, C.C.; Favrot, C. EcPV2 DNA in equine papillomas and in situ and invasive squamous cell carcinomas supports papillomavirus etiology. Vet. Pathol. 2013, 50, 686–692. [Google Scholar] [CrossRef]
- Massa, C.; Seliger, B. The tumor microenvironment: Thousand obstacles for effector T cells. Cell. Immunol. 2017, 343. [Google Scholar] [CrossRef]
- van den Top, J.G.B.; Harkema, L.; Lange, C.; Ensink, J.M.; van de Lest, C.H.A.; Barneveld, A.; van Weeren, P.R.; Gröne, A.; Martens, A. Expression of p53, Ki67, EcPV2- and EcPV3 DNA, and viral genes in relation to metastasis and outcome in equine penile and preputial squamous cell carcinoma. Equine Vet. J. 2015, 47, 188–195. [Google Scholar] [CrossRef]
- Teh, J.; O’Connor, E.; O’Brien, J.; Lim, W.M.; Taylor, M.; Heriot, A.; Ramsay, R.; Lawrentschuk, N. Future directions in advanced penile cancer–mechanisms of carcinogenesis and a search for targeted therapy. Futur. Oncol. 2020, 16, 29. [Google Scholar] [CrossRef]
- Wu, S.; Wu, Y.; Lu, Y.; Yue, Y.; Cui, C.; Yu, M.; Wang, S.; Liu, M.; Zhao, Y.; Sun, Z. STAT1 expression and HPV16 viral load predict cervical lesion progression. Oncol. Lett. 2020, 20. [Google Scholar] [CrossRef]
- Li, C.X.; Chang, W.S.; Mitsakos, K.; Rodger, J.; Holmes, E.C.; Hudson, B.J. Identification of a novel equine papillomavirus in semen from a thoroughbred stallion with a penile lesion. Viruses 2019, 11, 713. [Google Scholar] [CrossRef] [Green Version]
- Falini, B.; Fizzotti, M.; Pucciarini, A.; Bigerna, B.; Marafioti, T.; Gambacorta, M.; Pacini, R.; Alunni, C.; Natali-Tanci, L.; Ugolini, B.; et al. A monoclonal antibody (MUM1p) detects expression of the MUM1/IRF4 protein in a subset of germinal center B cells, plasma cells, and activated T cells. Blood 2000, 95, 2084–2092. [Google Scholar] [CrossRef]
- Lee, H.E.; Luo, L.; Kroneman, T.; Passow, M.R.; Del Rosario, K.M.; Christensen, M.R.; Francis, M.E.; O’Shaughnessy, J.W.; Blahnik, A.J.; Yang, P.; et al. Increased Plasma Cells and Decreased B-cells in Tumor Infiltrating Lymphocytes are Associated with Worse Survival in Lung Adenocarcinomas. J. Clin. Cell. Immunol. 2020, 11, 584. [Google Scholar]
- Shalapour, S.; Font-Burgada, J.; Di Caro, G.; Zhong, Z.; Sanchez-Lopez, E.; Dhar, D.; Willimsky, G.; Ammirante, M.; Strasner, A.; Hansel, D.E.; et al. Immunosuppressive plasma cells impede T-cell-dependent immunogenic chemotherapy. Nature 2015, 521, 94–98. [Google Scholar] [CrossRef]
- Wang, L.; Fu, Y.; Chu, Y. Regulatory B Cells. Adv. Exp. Med. Biol. 2020, 1254, 87–103. [Google Scholar]
- Attias, M.; Al--Aubodah, T.; Piccirillo, C.A. Mechanisms of human FoxP3 + T reg cell development and function in health and disease. Clin. Exp. Immunol. 2019, 197, 36–51. [Google Scholar] [CrossRef] [Green Version]
- Shang, B.; Liu, Y.; Jiang, S.; Liu, Y. Prognostic value of tumor-infiltrating FoxP3+ regulatory T cells in cancers: A systematic review and meta-analysis. Sci. Rep. 2015, 5, 15179. [Google Scholar] [CrossRef] [Green Version]
- Porcellato, I.; Brachelente, C.; De Paolis, L.; Menchetti, L.; Silvestri, S.; Sforna, M.; Vichi, G.; Iussich, S.; Mechelli, L. FoxP3 and IDO in canine melanocytic tumors. Vet. Pathol. 2019, 56, 189–199. [Google Scholar] [CrossRef]
- Sikora, M.; Kopeć, B.; Piotrowska, K.; Pawlik, A. Role of allograft inflammatory factor-1 in pathogenesis of diseases. Immunol. Lett. 2020, 218, 1–4. [Google Scholar] [CrossRef]
- Legradi, A.; Dulka, K.; Jancsó, G.; Gulya, K. Orofacial skin inflammation increases the number of macrophages in the maxillary subregion of the rat trigeminal ganglion in a corticosteroid-reversible manner. Cell Tissue Res. 2020. [Google Scholar] [CrossRef]
- Karagianni, A.E.; Lisowski, Z.M.; Hume, D.A.; Scott Pirie, R. The equine mononuclear phagocyte system: The relevance of the horse as a model for understanding human innate immunity. Equine Vet. J. 2020. [Google Scholar] [CrossRef]
- Lechien, J.R.; Descamps, G.; Seminerio, I.; Furgiuele, S.; Dequanter, D.; Mouawad, F.; Badoual, C.; Journe, F.; Saussez, S. HPV involvement in the tumor microenvironment and immune treatment in head and neck squamous cell carcinomas. Cancers 2020, 12, 1060. [Google Scholar] [CrossRef]
- Cai, X.; Zhang, L.; Wei, W. Regulatory B cells in inflammatory diseases and tumor. Int. Immunopharmacol. 2019, 67, 281–286. [Google Scholar] [CrossRef]
- Yoshimoto, T.; Xu, M.; Mizoguchi, I.; Morishima, N.; Chiba, Y.; Mizuguchi, J. Regulation of antitumor immune responses by the IL-12 family cytokines, IL-12, IL-23, and IL-27. Clin. Dev. Immunol. 2010, 2010, 832454. [Google Scholar]
- Zijlmans, H.J.M.A.A.; Punt, S.; Fleuren, G.J.; Trimbos, J.B.; Kenter, G.G.; Gorter, A. Role of IL-12p40 in cervical carcinoma. Br. J. Cancer 2012, 107, 1956–1962. [Google Scholar] [CrossRef] [Green Version]
- Gosmann, C.; Mattarollo, S.R.; Bridge, J.A.; Frazer, I.H.; Blumenthal, A. IL-17 Suppresses Immune Effector Functions in Human Papillomavirus-Associated Epithelial Hyperplasia. J. Immunol. 2014, 193, 2248–2257. [Google Scholar] [CrossRef]
- Lin, W.; Niu, Z.; Zhang, H.; Kong, Y.; Wang, Z.; Yang, X.; Yuan, F. Imbalance of Th1/Th2 and Th17/Treg during the development of uterine cervical cancer. Int. J. Clin. Exp. Pathol. 2019, 12, 3604–3612. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Antibody | Source | Working Dilution; Incubation | Antigen Retrieval | Reaction Pattern |
|---|---|---|---|---|
| CD3 | Rabbit polyclonal, Dako | 1:200; 1h RT | HIER, ph 9.0, TRIS-Edta | Membrane |
| CD20 | Rabbit polyclonal, Dako | 1:300; 1h RT | No AR | Membrane |
| FOXP3 | Rat monoclonal, eBioscience | 1:50; ON 4 °C | HIER, ph 9.0, TRIS-Edta | Nuclear |
| MUM1 | Mouse monoclonal, Dako | 1:200; 2h RT | HIER, ph 9.0, TRIS-Edta | Nuclear and cytoplasmic |
| IBA1 | Goat polyclonal, LSBio | 1:50; 2h RT | HIER, ph 9.0, TRIS-Edta | Cytoplasmic |
| MAC387 | Mouse monoclonal, Dako | 1:100; 2h RT | HIER, ph 9.0, TRIS-Edta | Cytoplasmic |
| Ki-67 | Mouse monoclonal, Dako | 1:50; 2h RT | HIER, ph 9.0, TRIS-Edta | Nuclear |
| Gene | Sequences | Accession Number | Bp |
|---|---|---|---|
| Ec-PV2-L1 | F-5′-TTGTCCAGGAGAGGGGTTAG-3′ | NC_012123.1 | 81 |
| R-5′-TGCCTTCCTTTTCTTGGTGG-3′ | |||
| pEc-PV2-L1 | FAM-CGTCCAGCACCTTCGACCACCA-TAMRA | NC_012123.1 | 22 |
| Ec-B2M DNA detection | F-5′-CTGATGTTCTCCAGGTGTTCC-3′ | NM_001082502.3 | 114 |
| R-5′-TCAATCTCAGGCGGATGGAA-3′ | |||
| Ec-B2M cDNA expression | F-5′-GGCTACTCTCCCTGACTGG-3′ | NM_001082502.3 | 136 |
| R-5′-TCAATCTCAGGCGGATGGAA-3′ | |||
| pEc-B2M | FAM-ACTCACGTCACCCAGCAGAGA-TAMRA | NM_001082502.3 | 21 |
| Gene | Primer Pairs Sequences | mRNA Position | Genomic Position | Amplicon Length | Accession |
|---|---|---|---|---|---|
| B2M | F-5′-GGCTACTCTCCCTGACTGG-3′ | 32-50 | chr1:145961271-145964672 | 136 | NM_001082502.3 |
| R-5′-TCAATCTCAGGCGGATGGAA-3′ | 147-167 | ||||
| TGFB1 | F-5′-CGGAATGGCTGTCCTTTGATG-3′ | 577-597 | chr10:12028778-12030603 | 127 | NM_001081849.1 |
| R-5′-CCCACGCGGAGTGTGT-TAT-3′ | 685-703 | ||||
| IL5 | F-5′-ACCTGATGATTCCTACTCCTGA-3′ | 145-166 | chr14:42325437+42326396 | 99 | NM_001082499.1 |
| R-5′-CCCCTTGGACAGTTTGATTCT-3′ | 223-243 | ||||
| IL8 | F-5′-CTGGCTGTGGCTCTCTTG-3′ | 13-30 | chr3:63719744-63720834 | 133 | NM_001083951.2 |
| R-5′-CAGTTTGGGATTGAAAGGTTTG-3′ | 122-143 | ||||
| IL10 | F-5′-TTCAGCAGGGTGAAGACTTTCT-3′ | 141-162 | chr5:2996651-2997849 | 107 | NM_001082490.1 |
| R-5′-AAGGCTTGGCAACCCAGGTA-3′ | 228-247 | ||||
| IL12/p35 | F-5′-CTGAGGACCGTCAGCAACAC-3′ | 243-262 | chr19:3775251-3777993 | 147 | NM_001082511.2 |
| R-5′-GTTCGGGGCGAGTTCCAG-3′ | 372-389 | ||||
| IL12/p40 | F-5′-GATCGTGGTGGATGCTGTTC-3′ | 629-648 | chr14:19332153+19333691 | 132 | NM_001082516.1 |
| R-5′-TCCACCTGCCGAGAATTCTT-3′ | 741-760 | ||||
| IL2 | F-5′-GAAGAAGAACTCAAACCTCTG-3′ | 237-257 | chr2:105984416+105986321 | 148 | NM_001085433.2 |
| R-5′-TTCCTGTCTCATCATCATATTC-3′ | 363-384 | ||||
| IFNG | F-5′-GCTGTGTGCGATTTTGGGT-3′ | 33-51 | chr6:84511911-84513199 | 130 | NM_001081949.1 |
| R-5′-ATCCAGGAAAAGAGGCCCAC-3′ | 142-161 |
| Histopathological Features | Number of Cases | Percentage (%) | |
|---|---|---|---|
| Histological diagnosis | SCC | 17 | 85.0 |
| in situ carcinoma | 2 | 10.0 | |
| Papilloma | 1 | 5.0 | |
| Differentiation grade | Poorly differentiation | 3 | 17.6 |
| Moderately differentiated | 12 | 70.6 | |
| Well differentiated | 2 | 11.8 | |
| Vascular invasion | Present | 4 | 23.5 |
| Absent | 16 | 76.5 | |
| Ulceration | Present | 15 | 75.0 |
| Absent | 5 | 25.0 | |
| Necrosis | Present | 6 | 30.0 |
| Absent | 14 | 60.0 | |
| Stroma | Scant | 5 | 25.0 |
| Moderate | 10 | 50.0 | |
| Severe | 5 | 25.0 | |
| Inflammation | Mild | 4 | 20.0 |
| Moderate | 13 | 65.0 | |
| Severe | 3 | 15.0 | |
| Absent | 0 | 0.0 | |
| Lymphocytes | Mild | 6 | 30.0 |
| Moderate | 6 | 30.0 | |
| Severe | 8 | 40.0 | |
| Absent | 0 | 0.0 | |
| Plasma cells | Mild | 9 | 45.0 |
| Moderate | 5 | 25.0 | |
| Severe | 5 | 25.0 | |
| Absent | 1 | 5.0 | |
| Macrophages | Mild | 14 | 70.0 |
| Moderate | 6 | 30.0 | |
| Severe | 0 | 0.0 | |
| Absent | 0 | 0.0 | |
| Neutrophils | Mild | 5 | 25.0 |
| Moderate | 12 | 60.0 | |
| Severe | 2 | 10.0 | |
| Absent | 1 | 5.0 | |
| Eosinophils | Mild | 8 | 40.0 |
| Moderate | 5 | 25.0 | |
| Severe | 1 | 5.0 | |
| Absent | 6 | 30.0 | |
| CD3 | FoxP3 | CD20 | MUM1 | MAC387 (n) | MAC387 (m) | IBA1 | Mitotic Count | Ki-67 Index | |
|---|---|---|---|---|---|---|---|---|---|
| CD3 | 1.000 | 0.321 | 0.272 | 0.472 * | 0.341 | −0.221 | 0.399 | 0.335 | 0.213 |
| FoxP3 | 1.000 | 0.494 * | 0.598 * | 0.564 * | 0.047 | 0.411 | 0.422 | 0.257 | |
| CD20 | 1.000 | 0.692 ** | 0.564 ** | 0.175 | 0.742 ** | 0.107 | 0.218 | ||
| MUM1 | 1.000 | 0.490 * | 0.039 | 0.606 ** | 0.150 | 0.480 | |||
| MAC387 (n) | 1.000 | 0.198 | 0.359 | 0.026 | 0.314 | ||||
| MAC387 (m) | 1.000 | 0.014 | 0.430 | -0.010 | |||||
| IBA1 | 1.000 | 0.393 | -0.028 | ||||||
| Mitotic count | 1.000 | -0.172 | |||||||
| KI67 index | 1.000 |
| ID | Histological Diagnosis | DNA | cDNA | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| B2M | L1 | Viral Load | L1 | B2M | IL5 | IL8 | IL10 | IL12/p35 | IL12/p40 | ||
| 1 | SCC | + | +++ | High | 33.5 ± 0.2 | 33.0 ± 0.2 | 37.4 ± 0.5 | 35.8 ± 1.5 | >48 | >48 | >48 |
| 2 | SCC | + | + | Low | >48 | 31.2 ± 0.1 | >48 | 35.4 ± 0.3 | >48 | 36.7 ± 1.5 | 37.2 ± 0.7 |
| 3 | CIS | + | +++ | High | 34.3 ± 0.3 | 33.2 ± 0.3 | >48 | 31.2 ± 1.6 | >48 | >48 | 35.4 ± 0.2 |
| 4 | SCC | + | ++++ | High | 35.1 ± 0.6 | 35.5 ± 0.4 | >48 | >48 | 37.8±0.9 | >48 | 36.5 ± 0.7 |
| 5 | P | + | + | Low | >48 | 32.5 ± 0.3 | 39.8 ± 2.3 | >48 | >48 | 35.0 ± 1.2 | 36.8 ± 0.1 |
| 6 | SCC | + | + | Low | >48 | 33.4 ± 1.0 | >48 | >48 | >48 | 45.0 ± 2.1 | 42.3 ± 7.7 |
| 7 | SCC | + | ++++ | High | 35.9 ± 0.7 | 34.5 ± 0.3 | >48 | >48 | >48 | >48 | 36.8 ± 0.1 |
| 8 | SCC | + | ++++ | High | 32.3 ± 0.3 | 33.0 ± 0.4 | >48 | 34.2 ± 0.2 | >48 | >48 | 36.9 ± 0.1 |
| 9 | SCC | + | ++++ | High | 34.9 ± 0.9 | 33.0 ± 0.5 | >48 | 36.0 ± 0.5 | >48 | >48 | 35.9 ± 0.2 |
| 10 | SCC | + | + | Low | >48 | 37.9 ± 0.9 | >48 | 34.9 ± 0.7 | >48 | 41.2 ± 2.5 | 36.5 ± 0.1 |
| 11 | SCC | + | ++++ | High | 33.0 ± 0.3 | 34.9 ± 0.8 | >48 | 35.9 ± 0.6 | >48 | >48 | 36.5 ± 0.5 |
| 12 | SCC | + | + | Low | >48 | 34.5 ± 0.3 | >48 | 37.2 ± 0.7 | >48 | 45.0 ± 3.4 | 37.6 ± 1.3 |
| 13 | SCC | + | ++ | Low | 36.6 ± 0.1 | 34.1 ± 1.3 | >48 | 36.7 ± 0.8 | >48 | >48 | >48 |
| 14 | SCC | + | ++++ | High | 31.3 ± 2.7 | 33.6 ± 0.5 | >48 | 36.7 ± 0.3 | >48 | >48 | 36.7 ± 0.6 |
| 15 | SCC | + | ++++ | High | 35.6 ± 0.1 | 34.2 ± 0.2 | >48 | 34.2 ± 0.1 | 39.7±4.6 | >48 | >48 |
| 16 | SCC | + | ++ | Low | 37.8 ± 1.5 | 27.9 ± 0.1 | 35.0 ± 1.1 | 26.1 ± 0.1 | 35.5 ± 1.7 | 38.2 ± 3.3 | 34.8 ± 0.1 |
| 17 | SCC | + | - | - | ND | 29.4 ± 0.2 | 42.6 ± 6.7 | 33.0 ± 0.7 | 36.5 ± 0.9 | 36.5 ± 1.7 | 35.9 ± 0.6 |
| 18 | CIS | + | +++ | High | 31.3 ± 2.7 | 32.6 ± 0.3 | 37.5 ± 0.9 | 30.5 ± 0.2 | >48 | >48 | 36.1 ± 0.6 |
| 19 | SCC | + | +++ | High | 35.3 ± 0.1 | 31.9 ± 0.5 | >48 | 33.4 ± 0.5 | 35.3 ± 1.5 | >48 | 36.0 ± 0.1 |
| 20 | SCC | + | - | - | ND | 32.1 ± 0.9 | >48 | 36.5 ± 0.7 | >48 | 45.6 ± 2.3 | 35.5 ± 0.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Porcellato, I.; Mecocci, S.; Mechelli, L.; Cappelli, K.; Brachelente, C.; Pepe, M.; Orlandi, M.; Gialletti, R.; Passeri, B.; Ferrari, A.; et al. Equine Penile Squamous Cell Carcinomas as a Model for Human Disease: A Preliminary Investigation on Tumor Immune Microenvironment. Cells 2020, 9, 2364. https://doi.org/10.3390/cells9112364
Porcellato I, Mecocci S, Mechelli L, Cappelli K, Brachelente C, Pepe M, Orlandi M, Gialletti R, Passeri B, Ferrari A, et al. Equine Penile Squamous Cell Carcinomas as a Model for Human Disease: A Preliminary Investigation on Tumor Immune Microenvironment. Cells. 2020; 9(11):2364. https://doi.org/10.3390/cells9112364
Chicago/Turabian StylePorcellato, Ilaria, Samanta Mecocci, Luca Mechelli, Katia Cappelli, Chiara Brachelente, Marco Pepe, Margherita Orlandi, Rodolfo Gialletti, Benedetta Passeri, Angelo Ferrari, and et al. 2020. "Equine Penile Squamous Cell Carcinomas as a Model for Human Disease: A Preliminary Investigation on Tumor Immune Microenvironment" Cells 9, no. 11: 2364. https://doi.org/10.3390/cells9112364
APA StylePorcellato, I., Mecocci, S., Mechelli, L., Cappelli, K., Brachelente, C., Pepe, M., Orlandi, M., Gialletti, R., Passeri, B., Ferrari, A., Modesto, P., Ghelardi, A., & Razzuoli, E. (2020). Equine Penile Squamous Cell Carcinomas as a Model for Human Disease: A Preliminary Investigation on Tumor Immune Microenvironment. Cells, 9(11), 2364. https://doi.org/10.3390/cells9112364