Shared Molecular Genetic Mechanisms Underlie Endometriosis and Migraine Comorbidity

 , , , , , ,
, , , , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Sources and Study Samples
2.1.1. IEC Endometriosis GWAS Data
2.1.2. IHGC Migraine GWAS Data
2.1.3. United Kingdom (UK) Biobank Data
2.2. SNP Effect Concordance Analysis (SECA)
2.3. Linkage Disequilibrium Score Regression (LDSC)
2.4. Cross-Disorder Meta-Analysis of Endometriosis and Migraine
2.5. Mendelian Randomisation (MR)
2.6. Gene-Based Association Study
2.7. Overlapping Genes and Statistics Tests
2.8. Pathway-Based Functional Enrichment Analyses
3. Results
3.1. SECA: Genetic Overlap between Endometriosis and Migraine
3.2. LD Score Regression Results for Endometriosis-Migraine
3.3. SNPs Associated with Endometriosis and Migraine
3.4. Mendelian Randomisation (MR)
3.5. Gene-Based Analysis for Endometriosis and Migraine
3.6. Functional Enrichment Analyses
4. Discussion
Strengths and Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
List of Members of Consortia and Institutions
References
- Giudice, L.C. Endometriosis. N. Engl. J. Med. 2010, 362, 2389–2398. [Google Scholar] [CrossRef] [PubMed]
- Zondervan, K.T.; Becker, C.M.; Koga, K.; Missmer, S.A.; Taylor, R.N.; Viganò, P. Endometriosis. Nat. Rev. Dis. Primers 2018, 4, 9. [Google Scholar] [CrossRef] [PubMed]
- Vos, T.; Barber, R.M.; Bell, B.; Bertozzi-Villa, A.; Biryukov, S.; Bolliger, I.; Charlson, F.; Davis, A.; Degenhardt, L.; Dicker, D. Global, regional, and national incidence, prevalence, and years lived with disability for 301 acute and chronic diseases and injuries in 188 countries, 1990–2013: A systematic analysis for the Global Burden of Disease Study 2013. Lancet 2015, 386, 743–800. [Google Scholar] [CrossRef] [Green Version]
- Yeh, W.Z.; Blizzard, L.; Taylor, B.V. What is the actual prevalence of migraine? Brain Behav. 2018, 8, e00950. [Google Scholar] [CrossRef] [PubMed]
- Maitrot-Mantelet, L.; Hugon-Rodin, J.; Vatel, M.; Marcellin, L.; Santulli, P.; Chapron, C.; Plu-Bureau, G. Migraine in relation with endometriosis phenotypes: Results from a French case-control study. Cephalalgia: Int. J. Head. 2019. [Google Scholar] [CrossRef]
- Tanos, V.; Raad, E.A.; Berry, K.E.; Toney, Z.A. Review of migraine incidence and management in obstetrics and gynaecology. Eur. J. Obstetr. Gynecol. Reprod. Biol. 2019, 240, 248–255. [Google Scholar] [CrossRef]
- Rothrock, J.F. What is migraine? Head.: J. Head Face Pain 2008, 48, 330. [Google Scholar] [CrossRef]
- Nnoaham, K.E.; Hummelshoj, L.; Webster, P.; d’Hooghe, T.; de Cicco, N.F.; de Cicco, N.C.; Jenkinson, C.; Kennedy, S.H.; Zondervan, K.T.; World Endometriosis Research Foundation Global Study of Women's Health consortium. Reprint of: Impact of endometriosis on quality of life and work productivity: A multicenter study across ten countries. Fertil. Steril. 2019, 112. [Google Scholar] [CrossRef]
- Agosti, R. Migraine burden of disease: From the patient’s experience to a socio-economic view. Head.: J. Head Face Pain 2018, 58, 17–32. [Google Scholar] [CrossRef] [Green Version]
- Buse, D.C.; Fanning, K.M.; Reed, M.L.; Murray, S.; Dumas, P.K.; Adams, A.M.; Lipton, R.B. Life with migraine: Effects on relationships, career, and finances from the chronic migraine epidemiology and outcomes (CaMEO) study. Head.: J. Head Face Pain 2019, 59, 1286–1299. [Google Scholar] [CrossRef] [Green Version]
- Hållstam, A.; Stålnacke, B.M.; Svensén, C.; Löfgren, M. Living with painful endometriosis – A struggle for coherence. A qualitative study. Sex. Reprod. Healthcare 2018, 17, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Rei, C.; Williams, T.; Feloney, M. Endometriosis in a man as a rare source of abdominal pain: A case report and review of the literature. Case Rep. Obstetr. Gynecol. 2018, 2018, 2083121. [Google Scholar] [CrossRef] [Green Version]
- Nyholt, D.R.; Gillespie, N.G.; Merikangas, K.R.; Treloar, S.A.; Martin, N.G.; Montgomery, G.W. Common genetic influences underlie comorbidity of migraine and endometriosis. Genet. Epidemiol. 2009, 33, 105–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmussen, B.K. Migraine and tension-type headache in a general population: Precipitating factors, female hormones, sleep pattern and relation to lifestyle. Pain 1993, 53, 65–72. [Google Scholar] [CrossRef]
- Wang, S.J.; Fuh, J.L.; Young, Y.H.; Lu, S.R.; Shia, B.C. Prevalence of migraine in Taipei, Taiwan: A population-based survey. Cephalalgia 2000, 20, 566–572. [Google Scholar] [CrossRef] [PubMed]
- Vetvik, K.G.; MacGregor, E.A. Sex differences in the epidemiology, clinical features, and pathophysiology of migraine. Lancet Neurol. 2017, 16, 76–87. [Google Scholar] [CrossRef]
- Aegidius, K.; Zwart, J.A.; Hagen, K.; Dyb, G.; Holmen, T.; Stovner, L. Increased headache prevalence in female adolescents and adult women with early menarche. The Head-HUNT Studies. Eur. J. Neurol. 2011, 18, 321–328. [Google Scholar] [CrossRef]
- Tietjen, G.E.; Conway, A.; Utley, C.; Gunning, W.T.; Herial, N.A. Migraine is associated with menorrhagia and endometriosis. Head.: J. Head Face Pain 2006, 46, 422–428. [Google Scholar] [CrossRef]
- James, A. More than menorrhagia: A review of the obstetric and gynaecological manifestations of bleeding disorders. Haemophilia 2005, 11, 295–307. [Google Scholar] [CrossRef]
- Vercellini, P.; Viganò, P.; Somigliana, E.; Fedele, L. Endometriosis: Pathogenesis and treatment. Nat. Rev. Endocrinol. 2014, 10, 261. [Google Scholar] [CrossRef]
- Matalliotakis, I.M.; Cakmak, H.; Fragouli, Y.G.; Goumenou, A.G.; Mahutte, N.G.; Arici, A. Epidemiological characteristics in women with and without endometriosis in the Yale series. Arch. Gynecol. Obstetr. 2008, 277, 389–393. [Google Scholar] [CrossRef] [PubMed]
- Missmer, S.A.; Hankinson, S.E.; Spiegelman, D.; Barbieri, R.L.; Marshall, L.M.; Hunter, D.J. Incidence of laparoscopically confirmed endometriosis by demographic, anthropometric, and lifestyle factors. Am. J. Epidemiol. 2004, 160, 784–796. [Google Scholar] [CrossRef] [PubMed]
- Kvisvik, E.V.; Stovner, L.J.; Helde, G.; Bovim, G.; Linde, M. Headache and migraine during pregnancy and puerperium: The MIGRA-study. J. Head. Pain 2011, 12, 443–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrovski, B.É.; Vetvik, K.G.; Lundqvist, C.; Eberhard-Gran, M. Characteristics of menstrual versus non-menstrual migraine during pregnancy: A longitudinal population-based study. J. Head. Pain 2018, 19, 27. [Google Scholar] [CrossRef] [Green Version]
- Lichten, E.M.; Bennett, R.S.; Whitty, A.J.; Daoud, Y. Efficacy of danazol in the control of hormonal migraine. J. Reprod. Med. 1991, 36, 419–424. [Google Scholar]
- Tervilä, L.; Marttila, P. Headache as a Symptom of Endometriosis Externa. Ann. Chirurgiae Gynaecol. Fenniae 1975, 64, 239–241. [Google Scholar]
- Ferrero, S.; Pretta, S.; Bertoldi, S.; Anserini, P.; Remorgida, V.; Del Sette, M.; Gandolfo, C.; Ragni, N. Increased frequency of migraine among women with endometriosis. Hum. Reprod. 2004, 19, 2927–2932. [Google Scholar] [CrossRef] [Green Version]
- Mirkin, D.; Murphy-Barron, C.; Iwasaki, K. Actuarial analysis of private payer administrative claims data for women with endometriosis. J. Manag. Care Pharm. 2007, 13, 262–272. [Google Scholar] [CrossRef]
- Karamustafaoglu Balci, B.; Kabakci, Z.; Guzey, D.Y.; Avci, B.; Guler, M.; Attar, E. Association between endometriosis, headache, and migraine. J. Endometr. Pelvic Pain Disord. 2019, 11, 19–24. [Google Scholar] [CrossRef]
- Miller, J.A.; Missmer, S.A.; Vitonis, A.F.; Sarda, V.; Laufer, M.R.; DiVasta, A.D. Prevalence of migraines in adolescents with endometriosis. Fertil. Steril. 2018, 109, 685–690. [Google Scholar] [CrossRef] [Green Version]
- Sapkota, Y.; Steinthorsdottir, V.; Morris, A.P.; Fassbender, A.; Rahmioglu, N.; De Vivo, I.; Buring, J.E.; Zhang, F.; Edwards, T.L.; Jones, S.; et al. Meta-analysis identifies five novel loci associated with endometriosis highlighting key genes involved in hormone metabolism. Nat. Commun. 2017, 8, 15539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gormley, P.; Anttila, V.; Winsvold, B.S.; Palta, P.; Esko, T.; Pers, T.H.; Farh, K.H.; Cuenca-Leon, E.; Muona, M.; Furlotte, N.A.; et al. Meta-analysis of 375,000 individuals identifies 38 susceptibility loci for migraine. Nat. Genet. 2016, 48, 856–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canis, M.; Donnez, J.; Guzick, D.; Halme, J.; Rock, J.; Schenken, R.; Vernon, M. Revised american society for reproductive medicine classification of endometriosis: 1996. Fertil. Steril. 1997, 67, 817–821. [Google Scholar]
- Headache Classification Committee of the International Headache Society (IHS). The International Classification of Headache Disorders, 3rd edition (beta version). Cephalalgia 2013, 33, 629–808. [Google Scholar] [CrossRef] [Green Version]
- 1000 Genomes Project Consortium; Abecasis, G.R.; Auton, A.; Brooks, L.D.; DePristo, M.A.; Durbin, R.M.; Handsaker, R.E.; Kang, H.M.; Marth, G.T.; McVean, G.A. An integrated map of genetic variation from 1092 human genomes. Nature 2012, 491, 56–65. [Google Scholar]
- Magi, R.; Morris, A.P. GWAMA: Software for genome-wide association meta-analysis. BMC Bioinform. 2010, 11, 288. [Google Scholar] [CrossRef] [Green Version]
- Sudlow, C.; Gallacher, J.; Allen, N.; Beral, V.; Burton, P.; Danesh, J.; Downey, P.; Elliott, P.; Green, J.; Landray, M. UK biobank: An open access resource for identifying the causes of a wide range of complex diseases of middle and old age. PLoS Med. 2015, 12, e1001779. [Google Scholar] [CrossRef] [Green Version]
- Nyholt, D.R. SECA: SNP effect concordance analysis using genome-wide association summary results. Bioinformatics 2014, 30, 2086–2088. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Zhao, H.; Boomsma, D.I.; Ligthart, L.; Belin, A.C.; Smith, G.D.; Esko, T.; Freilinger, T.M.; Hansen, T.F.; Ikram, M.A. Molecular genetic overlap between migraine and major depressive disorder. Eur. J. Hum. Genet. 2018, 26, 1202. [Google Scholar] [CrossRef] [Green Version]
- Sapkota, Y.; Low, S.-K.; Attia, J.; Gordon, S.D.; Henders, A.K.; Holliday, E.G.; MacGregor, S.; Martin, N.G.; McEvoy, M.; Morris, A.P.; et al. Association between endometriosis and the interleukin 1A (IL1A) locus. Hum. Reprod. 2015, 30, 239–248. [Google Scholar] [CrossRef] [Green Version]
- Han, B.; Eskin, E. Random-effects model aimed at discovering associations in meta-analysis of genome-wide association studies. Am. J. Hum. Genet. 2011, 88, 586–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemani, G.; Zheng, J.; Elsworth, B.; Wade, K.H.; Haberland, V.; Baird, D.; Laurin, C.; Burgess, S.; Bowden, J.; Langdon, R. The MR-Base platform supports systematic causal inference across the human phenome. Elife 2018, 7, e34408. [Google Scholar] [CrossRef] [PubMed]
- Stone, G.W.; Pocock, S.J. Randomized trials, statistics, and clinical inference. J. Am. Coll. Cardiol. 2010, 55, 428–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liberopoulos, G.; Trikalinos, N.A.; Ioannidis, J.P. The elderly were under-represented in osteoarthritis clinical trials. J. Clin. Epidemiol. 2009, 62, 1218–1223. [Google Scholar] [CrossRef]
- Zheng, J.; Baird, D.; Borges, M.C.; Bowden, J.; Hemani, G.; Haycock, P.; Evans, D.M.; Smith, G.D. Recent Developments in Mendelian Randomization Studies. Curr. Epidemiol. Rep. 2017, 4, 330–345. [Google Scholar] [CrossRef] [Green Version]
- Bowden, J.; Davey Smith, G.; Haycock, P.C.; Burgess, S. Consistent estimation in Mendelian randomization with some invalid instruments using a weighted median estimator. Genet. Epidemiol. 2016, 40, 304–314. [Google Scholar] [CrossRef] [Green Version]
- Bowden, J.; Davey Smith, G.; Burgess, S. Mendelian randomization with invalid instruments: Effect estimation and bias detection through Egger regression. Int. J. Epidemiol. 2015, 44, 512–525. [Google Scholar] [CrossRef] [Green Version]
- Davies, N.M.; Holmes, M.V.; Smith, G.D. Reading Mendelian randomisation studies: A guide, glossary, and checklist for clinicians. BMJ 2018, 362, k601. [Google Scholar] [CrossRef] [Green Version]
- Staley, J.R.; Blackshaw, J.; Kamat, M.A.; Ellis, S.; Surendran, P.; Sun, B.B.; Paul, D.S.; Freitag, D.; Burgess, S.; Danesh, J.; et al. PhenoScanner: A database of human genotype–phenotype associations. Bioinformatics 2016, 32, 3207–3209. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.Z.; McRae, A.F.; Nyholt, D.R.; Medland, S.E.; Wray, N.R.; Brown, K.M.; Hayward, N.K.; Montgomery, G.W.; Visscher, P.M.; Martin, N.G.; et al. A Versatile Gene-Based Test for Genome-wide Association Studies. Am. J. Hum. Genet. 2010, 87, 139–145. [Google Scholar] [CrossRef] [Green Version]
- Mishra, A.; Macgregor, S. VEGAS2: Software for More Flexible Gene-Based Testing. Twin Res. Hum. Genet. 2014, 18, 86–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, H.; Eising, E.; De Vries, B.; Vijfhuizen, L.S.; Consortium, I.H.G.; Anttila, V.; Winsvold, B.S.; Kurth, T.; Stefansson, H.; Kallela, M. Gene-based pleiotropy across migraine with aura and migraine without aura patient groups. Cephalalgia 2016, 36, 648–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.-X.; Yeung, J.M.; Cherny, S.S.; Sham, P.C. Evaluating the effective numbers of independent tests and significant p-value thresholds in commercial genotyping arrays and public imputation reference datasets. Hum. Genet. 2012, 131, 747–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reimand, J.; Isserlin, R.; Voisin, V.; Kucera, M.; Tannus-Lopes, C.; Rostamianfar, A.; Wadi, L.; Meyer, M.; Wong, J.; Xu, C. Pathway enrichment analysis and visualization of omics data using g: Profiler, GSEA, Cytoscape and EnrichmentMap. Nat. Protocols 2019, 1. [Google Scholar] [CrossRef] [PubMed]
- Reimand, J.; Arak, T.; Adler, P.; Kolberg, L.; Reisberg, S.; Peterson, H.; Vilo, J. g:Profiler-a web server for functional interpretation of gene lists (2016 update). Nucleic Acids Res. 2016, 44, W83–W89. [Google Scholar] [CrossRef] [PubMed]
- Merico, D.; Isserlin, R.; Stueker, O.; Emili, A.; Bader, G.D. Enrichment map: A network-based method for gene-set enrichment visualization and interpretation. PLoS ONE 2010, 5, e13984. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Burgess, S.; Butterworth, A.; Thompson, S.G. Mendelian randomization analysis with multiple genetic variants using summarized data. Genet. Epidemiol. 2013, 37, 658–665. [Google Scholar] [CrossRef] [Green Version]
- Wainschtein, P.; Jain, D.P.; Yengo, L.; Zheng, Z.; Cupples, L.A.; Shadyab, A.H.; McKnight, B.; Shoemaker, B.M.; Mitchell, B.D.; Psaty, B.M. Recovery of trait heritability from whole genome sequence data. BioRxiv 2019, 588020. [Google Scholar] [CrossRef]
- Mbarek, H.; Steinberg, S.; Nyholt, D.R.; Gordon, S.D.; Miller, M.B.; McRae, A.F.; Hottenga, J.J.; Day, F.R.; Willemsen, G.; de Geus, E.J.; et al. Identification of Common Genetic Variants Influencing Spontaneous Dizygotic Twinning and Female Fertility. Am. J. Hum. Genet. 2016, 98, 898–908. [Google Scholar] [CrossRef] [Green Version]
- Ruth, K.S.; Beaumont, R.N.; Tyrrell, J.; Jones, S.E.; Tuke, M.A.; Yaghootkar, H.; Wood, A.R.; Freathy, R.M.; Weedon, M.N.; Frayling, T.M. Genetic evidence that lower circulating FSH levels lengthen menstrual cycle, increase age at menopause and impact female reproductive health. Hum. Reprod. 2016, 31, 473–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruth, K.S.; Campbell, P.J.; Chew, S.; Lim, E.M.; Hadlow, N.; Stuckey, B.G.; Brown, S.J.; Feenstra, B.; Joseph, J.; Surdulescu, G.L. Genome-wide association study with 1000 genomes imputation identifies signals for nine sex hormone-related phenotypes. Eur. J. Hum. Genet. 2016, 24, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haycock, P.C.; Burgess, S.; Wade, K.H.; Bowden, J.; Relton, C.; Davey Smith, G. Best (but oft-forgotten) practices: The design, analysis, and interpretation of Mendelian randomization studies. Am. J. Clin. Nutr. 2016, 103, 965–978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, H.; Nyholt, D.R. Gene-based analyses reveal novel genetic overlap and allelic heterogeneity across five major psychiatric disorders. Hum. Genet. 2017, 136, 263–274. [Google Scholar] [CrossRef]
- Joazeiro, C.A.; Weissman, A.M. RING finger proteins: Mediators of ubiquitin ligase activity. Cell 2000, 102, 549–552. [Google Scholar] [CrossRef] [Green Version]
- Blacque, O.; Leroux, M. Bardet-Biedl syndrome: An emerging pathomechanism of intracellular transport. Cell. Mol. Life Sci. CMLS 2006, 63, 2145–2161. [Google Scholar] [CrossRef]
- Chiang, A.P.; Beck, J.S.; Yen, H.-J.; Tayeh, M.K.; Scheetz, T.E.; Swiderski, R.E.; Nishimura, D.Y.; Braun, T.A.; Kim, K.-Y.A.; Huang, J. Homozygosity mapping with SNP arrays identifies TRIM32, an E3 ubiquitin ligase, as a Bardet–Biedl syndrome gene (BBS11). Proc. Natl. Acad. Sci. USA 2006, 103, 6287–6292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neri, M.; Selvatici, R.; Scotton, C.; Trabanelli, C.; Armaroli, A.; De Grandis, D.; Levy, N.; Gualandi, F.; Ferlini, A. A patient with limb girdle muscular dystrophy carries a TRIM32 deletion, detected by a novel CGH array, in compound heterozygosis with a nonsense mutation. Neuromusc. Disord. 2013, 23, 478–482. [Google Scholar] [CrossRef] [Green Version]
- Buzzi, M.G.; Moskowitz, M.A. The pathophysiology of migraine: Year 2005. J. Head. Pain 2005, 6, 105. [Google Scholar] [CrossRef] [Green Version]
- Alpay, Z.; Saed, G.M.; Diamond, M.P. Female infertility and free radicals: Potential role in adhesions and endometriosis. J. Soc. Gynecol. Investig. 2006, 13, 390–398. [Google Scholar] [CrossRef]
- Makker, A.; Goel, M.M.; Das, V.; Agarwal, A. PI3K-Akt-mTOR and MAPK signaling pathways in polycystic ovarian syndrome, uterine leiomyomas and endometriosis: An update. Gynecol. Endocrinol. 2012, 28, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Tonks, N.K. Protein tyrosine phosphatases: From genes, to function, to disease. Nat. Rev. Mol. Cell Biol. 2006, 7, 833–846. [Google Scholar] [CrossRef] [PubMed]
- Danen, E.H. Integrins: An overview of structural and functional aspects. In Madame Curie Bioscience Database; Landes Bioscience: Austin, TX, USA, 2013. [Google Scholar]
- Karar, J.; Maity, A. PI3K/AKT/mTOR pathway in angiogenesis. Front Mol. Neurosci. 2011, 4, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.Y.; Jiao, Z.Y.; Li, W.; Tian, Q. PI3K/AKT signaling pathway activation in a rat model of migraine. Mol. Med. Rep. 2017, 16, 4849–4854. [Google Scholar] [CrossRef] [PubMed]
- Goadsby, P.J.; Reuter, U.; Hallström, Y.; Broessner, G.; Bonner, J.H.; Zhang, F.; Sapra, S.; Picard, H.; Mikol, D.D.; Lenz, R.A. A controlled trial of erenumab for episodic migraine. N. Engl. J. Med. 2017, 377, 2123–2132. [Google Scholar] [CrossRef]
- Silberstein, S.D.; Dodick, D.W.; Bigal, M.E.; Yeung, P.P.; Goadsby, P.J.; Blankenbiller, T.; Grozinski-Wolff, M.; Yang, R.; Ma, Y.; Aycardi, E. Fremanezumab for the preventive treatment of chronic migraine. N. Engl. J. Med. 2017, 377, 2113–2122. [Google Scholar] [CrossRef]
- Cady, R.; Lipton, R.B. Qualitative change in migraine prevention? Head.: J. Head Face Pain 2018, 58, 1092–1095. [Google Scholar] [CrossRef]
- Slomovitz, B.M.; Coleman, R.L. The PI3K/AKT/mTOR pathway as a therapeutic target in endometrial cancer. Clin. Cancer Res. 2012, 18, 5856–5864. [Google Scholar] [CrossRef] [Green Version]
- Shih, R.-H.; Wang, C.-Y.; Yang, C.-M. NF-kappaB Signaling Pathways in Neurological Inflammation: A Mini Review. Front. Mol. Neurosci. 2015, 8. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB signaling in inflammation. Sign. Transd. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.-M.; An, J. Cytokines, inflammation, and pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, S.H.; Monsanto, S.P.; Miller, C.; Singh, S.S.; Thomas, R.; Tayade, C. Pathophysiology and Immune Dysfunction in Endometriosis. BioMed. Res. Int. 2015, 2015, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, A.; Wasiliew, P.; Kracht, M. Interleukin-1 (IL-1) Pathway. Sci. Signal. 2010, 3. [Google Scholar] [CrossRef] [PubMed]
- Capobianco, A.; Rovere-Querini, P. Endometriosis, a disease of the macrophage. Front. Immunol. 2013, 4, 9. [Google Scholar] [CrossRef] [Green Version]
- Idriss, H.T.; Naismith, J.H. TNFα and the TNF receptor superfamily: Structure-function relationship (s). Microsc. Res. Tech. 2000, 50, 184–195. [Google Scholar] [CrossRef]
- Bouwmeester, T.; Bauch, A.; Ruffner, H.; Angrand, P.-O.; Bergamini, G.; Croughton, K.; Cruciat, C.; Eberhard, D.; Gagneur, J.; Ghidelli, S.; et al. A physical and functional map of the human TNF-α/NF-κB signal transduction pathway. Nat. Cell Biol. 2004, 6, 97–105. [Google Scholar] [CrossRef]
- Pizzo, A.; Salmeri, F.M.; Ardita, F.V.; Sofo, V.; Tripepi, M.; Marsico, S. Behaviour of cytokine levels in serum and peritoneal fluid of women with endometriosis. Gynecol. Obstetr. Investing. 2002, 54, 82–87. [Google Scholar] [CrossRef]
- Schürks, M.; Rist, P.M.; Zee, R.Y.; Chasman, D.I.; Kurth, T. Tumour necrosis factor gene polymorphisms and migraine: A systematic review and meta-analysis. Cephalalgia 2011, 31, 1381–1404. [Google Scholar] [CrossRef]
- Gu, L.; Yan, Y.; Long, J.; Su, L.; Hu, Y.; Chen, Q.; Xie, J.; Wu, G. The TNF-α-308G/A polymorphism is associated with migraine risk: A meta-analysis. Exp. Therapeut. Med. 2012, 3, 1082–1086. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| P1 | P2 | Concordant SNPs | Discordant SNPs | Total SNPs | Proportion of Concordance | OR | P |
|---|---|---|---|---|---|---|---|
| 1 | 1 | 30790 | 28398 | 59188 | 0.52 | 1.18 | 8.77 × 10−23 |
| 0.9 | 0.9 | 27540 | 25213 | 52753 | 0.52 | 1.19 | 4.11 × 10−24 |
| 0.8 | 0.8 | 24222 | 21971 | 46193 | 0.52 | 1.22 | 1.30 × 10−25 |
| 0.7 | 0.7 | 20842 | 18665 | 39507 | 0.53 | 1.25 | 6.60 × 10−28 |
| 0.6 | 0.6 | 17474 | 15458 | 32932 | 0.53 | 1.28 | 1.23 × 10−28 |
| 0.5 | 0.5 | 14218 | 12383 | 26601 | 0.53 | 1.32 | 2.69 × 10−29 |
| 0.4 | 0.4 | 10973 | 9415 | 20388 | 0.54 | 1.36 | 1.31 × 10−27 |
| 0.3 | 0.3 | 7804 | 6510 | 14314 | 0.55 | 1.44 | 3.21 × 10−27 |
| 0.2 | 0.2 | 4771 | 3855 | 8626 | 0.55 | 1.53 | 6.92 × 10−23 |
| 0.1 | 0.1 | 1946 | 1496 | 3442 | 0.57 | 1.69 | 2.06 × 10−14 |
| 0.05 | 0.05 | 804 | 579 | 1383 | 0.58 | 1.92 | 2.23 × 10−09 |
| 0.01 | 0.01 | 85 | 43 | 128 | 0.66 | 3.61 | 7.20 × 10−04 |
| SNP-based Heritability | ||||||
| Phentype | Dataset source | Valid SNPs in analysis | Liability scale h2SNP (95% CI) | Intercept (se) | ||
| Endometriosis | IEC | 1,157,235 | 11.44% (10.73–12.15%) | Constrained to 1 | ||
| Migraine | IHGC | 1,173,223 | 8.99% (8.23–9.75%) | 1.0232 (0.008) | ||
| Migraine | UKBB | 1,177,705 | 16.87% (15.07–18.67%) | 1.0122 (0.007) | ||
| SNP-based Genetic Correlation | ||||||
| Phenotype 1 (data source) | Phenotype 2 (data source) | SNPs with valid alleles | rG (se) [p-value] | Phenotype 1 Intercept | Phenotype 2 Intercept | Gencov Intercept (se) |
| Endometriosis (IEC) | Migraine (IHGC) | 1,154,255 | 0.38 (0.0364) [2.30 × 10−25] | Constrained to 1 | 1.0214 (specified) | Constrained to 0 |
| Endometriosis (IEC) | Migraine (UKBB) | 1,152,558 | 0.14 (0.0438) 1.60 × 10−°3 | Constrained to 1 | 1.0136 (specified) | Constrained to 0 |
| SNPs | EA | OA | Beta (endo) | SE (endo) | P(endo) | Beta (migr) | SE (migr) | P(migr) | Beta (endo-migr) | F-Stat * | SE (endo-migr) | P (endo-migr) |
| rs10167914 | A | G | −0.11 | 0.02 | 1.10 × 10−°9 | 0.01 | 0.01 | 0.27 | −0.11 | 37.27 | 0.10 | 0.27 |
| rs11674184 | T | G | 0.12 | 0.01 | 2.67 × 10−17 | −0.02 | 0.01 | 0.16 | −0.13 | 71.40 | 0.09 | 0.16 |
| rs12037376 | A | G | 0.15 | 0.02 | 8.87 × 10−17 | 0.01 | 0.01 | 0.35 | −0.09 | 68.97 | 0.10 | 0.35 |
| rs12700667 | A | G | 0.10 | 0.02 | 9.08 × 10−1° | 0.01 | 0.01 | 0.6 | −0.07 | 37.64 | 0.13 | 0.61 |
| rs1537377 | T | C | −0.09 | 0.01 | 1.33 × 10−1° | 0.00 | 0.01 | 0.83 | 0.03 | 41.53 | 0.12 | 0.83 |
| rs1903068 | A | G | 0.10 | 0.01 | 1.04 × 10−11 | 0.01 | 0.01 | 0.32 | 0.11 | 46.28 | 0.11 | 0.32 |
| rs4762326 | T | C | 0.08 | 0.01 | 2.20 × 10−°9 | −0.01 | 0.01 | 0.31 | 0.14 | 35.55 | 0.13 | 0.31 |
| rs6546324 | A | C | 0.08 | 0.01 | 3.01 × 10−°8 | −0.01 | 0.01 | 0.42 | 0.11 | 30.56 | 0.14 | 0.42 |
| rs71575922 | C | G | −0.11 | 0.02 | 2.02 × 10−°8 | −0.01 | 0.01 | 0.35 | −0.12 | 31.41 | 0.13 | 0.35 |
| rs74485684 | T | C | 0.11 | 0.02 | 2.00 × 10−°8 | 0.04 | 0.01 | 0.01 | 0.36 | 31.55 | 0.13 | 0.01 |
| rs760794 | T | C | 0.09 | 0.01 | 1.79 × 10−1° | −0.02 | 0.01 | 0.07 | −0.22 | 40.43 | 0.12 | 0.07 |
| Methods | Number of SNPs | Beta | SE | P | ||||||||
| All—IVW | 11 | −0.02 | 0.05 | 0.67 | ||||||||
| All—MR Egger | 11 | −0.25 | 0.27 | 0.38 | ||||||||
| All—Simple mode | 11 | 0.08 | 0.12 | 0.50 | ||||||||
| All—Weighted mode | 11 | −0.11 | 0.07 | 0.14 | ||||||||
| All—Weighted median | 11 | −0.09 | 0.05 | 0.10 | ||||||||
| S/N | Chro | Gene | Start Position | Stop Position | IEC Endometriosis | IHGC Migraine | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene p-Value | Top SNP | Top SNP p-Value | Gene p-Value | Top SNP | Top SNP p-Value | FCP | |||||
| 1 | 11 | ARL14EP | 30344648 | 30359165 | 1.00 × 10−06 | rs4071559 | 5.60 × 10−08 | 5.54 × 10−02 | rs4071559 | 5.97 × 10−03 | 9.81 × 10−07 |
| 2 | 9 | TRIM32 | 119449580 | 119463579 | 2.76 × 10−02 | rs11793648 | 3.14 × 10−03 | 5.00 × 10−06 | rs76973802 | 7.15 × 10−07 | 2.32 × 10−06 |
| 3 | 17 | SLC35G6 | 7384720 | 7386383 | 1.59 × 10−03 | rs9891297 | 3.09 × 10−04 | 9.40 × 10−05 | rs8065577 | 2.21 × 10−05 | 2.50 × 10−06 |
| The effective number of genes in Endometriosis and migraine | |||||||||||
| Disorder | Total genes | P value < 0.1 | P < 0.05 | P < 0.01 | |||||||
| Raw c | Effective d | Raw c | Effective d | Proportion e | Raw c | Effective d | Proportion e | Raw c | Effective d | Proportion e | |
| Endometriosis a | 20473 | 17104 | 2966 | 2433 | 0.142 | 1749 | 1430 | 0.084 | 481 | 386 | 0.023 |
| Migraine b | 20473 | 17046 | 3239 | 2579 | 0.151 | 1871 | 1467 | 0.086 | 587 | 450 | 0.026 |
| Number of overlapping genes and binomial test results for gene-based association | |||||||||||
| Discovery | Targets | Overlapping genes | Proportion of overlap | Binomial test P-value | |||||||
| Raw | Effective | Expected | Observed | ||||||||
| P value < 0.01 | |||||||||||
| Endometriosis | Migraine | 17 | 15 | 450/17046 = 0.026 | 15/386 = 0.039 | 0.08259 | |||||
| P value < 0.05 | |||||||||||
| Endometriosis | Migraine | 196 | 171 | 1467/17046 = 0.086 | 171/1430 = 0.120 | 9.83 × 10−06 | |||||
| P value < 0.1 | |||||||||||
| Endometriosis | Migraine | 493 | 420 | 2579/17046 = 0.151 | 420/2433 = 0.173 | 1.85 × 10−03 | |||||
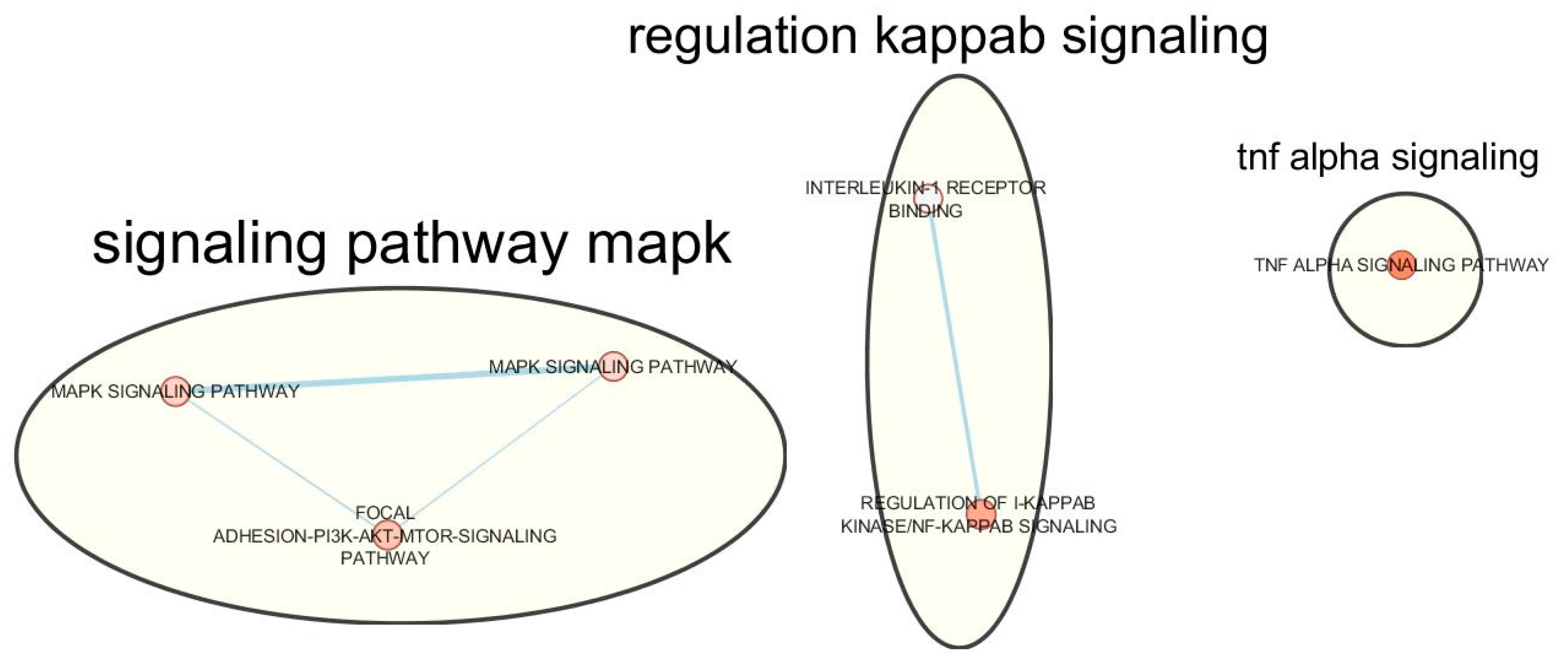
| Term ID for pathway | Pathway term name | Adjusted p-value | Genes |
| Source: Gene Ontology (Molecular function) | |||
| Interleukin-1 receptor binding | GO: 0005149 | 9.19 × 10−03 | IL36RN, IL37, IL36B, IL1B, IL1F10 |
| Source: Gene Ontology (Biological process) | |||
| Regulation of I-kappaB kinase and NF-kappaB signalling | GO: 0043122 | 1.90 × 10−02 | TRIM32, IL36RN, IL37, TMED4, IL36B, IL1B, RNF31, IKBKB, SHISA5, TANK, PARK2, IL1F10, ZDHHC17, GSTP1, DAB2IP, SLC35B2, TRIM13 |
| Source: Biological pathways (Kyoto Encyclopedia of Genes [KEGG]) | |||
| MAPK signalling pathway | KEGG: 04010 | 1.40 × 10−02 | IL1B, FGF18, NGF, IKBKB, MAP2K5, PTPN5, PDGFC, MAPK9, NRAS, PPP3CA, CACNA1E, FGF17, MAP2K6, FGF9, MET, RPS6KA4, FGFR4 |
| Source: Biological pathways (WikiPathways) | |||
| MAPK Signalling Pathway | WP: WP382 | 1.3 × 10−02 | FGF11, IL1B, FGF18, NGF, IKBKB, MAP2K5, PTPN5, MAPK9, NRAS, PPP3CA, CACNA1E, FGF17, MAP2K6, FGF9, RPS6KA4, FGFR4 |
| Focal Adhesion-PI3K-Akt-mTOR-signaling pathway | WP: WP3932 | 1.54 × 10−02 | FGF11, ITGB5, CREB5, PFKFB4, PPP2CA, DDIT4, FGF18, NGF, IKBKB, PTK2, PDGFC, SLC2A4, NRAS, CREB3L2, FGF17, FGF9, MET, FGFR4 |
| TNF alpha Signalling Pathway | WP: WP231 | 2.3 × 10−02 | PPP2CA, RFK, IKBKB, PTPRCAP, MAPK9, NSMAF, NRAS, TANK, MAP2K6 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adewuyi, E.O.; Sapkota, Y.; International Endogene Consortium (IEC); 23andMe Research Team; International Headache Genetics Consortium (IHGC); Auta, A.; Yoshihara, K.; Nyegaard, M.; Griffiths, L.R.; Montgomery, G.W.; et al. Shared Molecular Genetic Mechanisms Underlie Endometriosis and Migraine Comorbidity. Genes 2020, 11, 268. https://doi.org/10.3390/genes11030268
Adewuyi EO, Sapkota Y, International Endogene Consortium (IEC), 23andMe Research Team, International Headache Genetics Consortium (IHGC), Auta A, Yoshihara K, Nyegaard M, Griffiths LR, Montgomery GW, et al. Shared Molecular Genetic Mechanisms Underlie Endometriosis and Migraine Comorbidity. Genes. 2020; 11(3):268. https://doi.org/10.3390/genes11030268
Chicago/Turabian StyleAdewuyi, Emmanuel O., Yadav Sapkota, International Endogene Consortium (IEC), 23andMe Research Team, International Headache Genetics Consortium (IHGC), Asa Auta, Kosuke Yoshihara, Mette Nyegaard, Lyn R. Griffiths, Grant W. Montgomery, and et al. 2020. "Shared Molecular Genetic Mechanisms Underlie Endometriosis and Migraine Comorbidity" Genes 11, no. 3: 268. https://doi.org/10.3390/genes11030268
APA StyleAdewuyi, E. O., Sapkota, Y., International Endogene Consortium (IEC), 23andMe Research Team, International Headache Genetics Consortium (IHGC), Auta, A., Yoshihara, K., Nyegaard, M., Griffiths, L. R., Montgomery, G. W., Chasman, D. I., & Nyholt, D. R. (2020). Shared Molecular Genetic Mechanisms Underlie Endometriosis and Migraine Comorbidity. Genes, 11(3), 268. https://doi.org/10.3390/genes11030268