
Sequence Analysis of the Complete Mitochondrial Genome of a Medicinal Plant, Vitex rotundifolia Linnaeus f. (Lamiales: Lamiaceae)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and DNA Extraction
2.2. DNA Sequencing and Mitochondrial Genome Assembly
2.3. Mitochondrial Genome Annotation and Genome Alignments of Lamiales
2.4. Analysis of Repeat Sequences and Repeat-Mediated Homologous Recombinations
2.5. Identification of Plastid Integration
2.6. Phylogenetic Analysis of the 10 Lamiales Species Based on Shared Mitochondrial Protein Sequences
2.7. Estimation of Nucleotide Substitution Rates and RNA Editing Predicting
3. Results
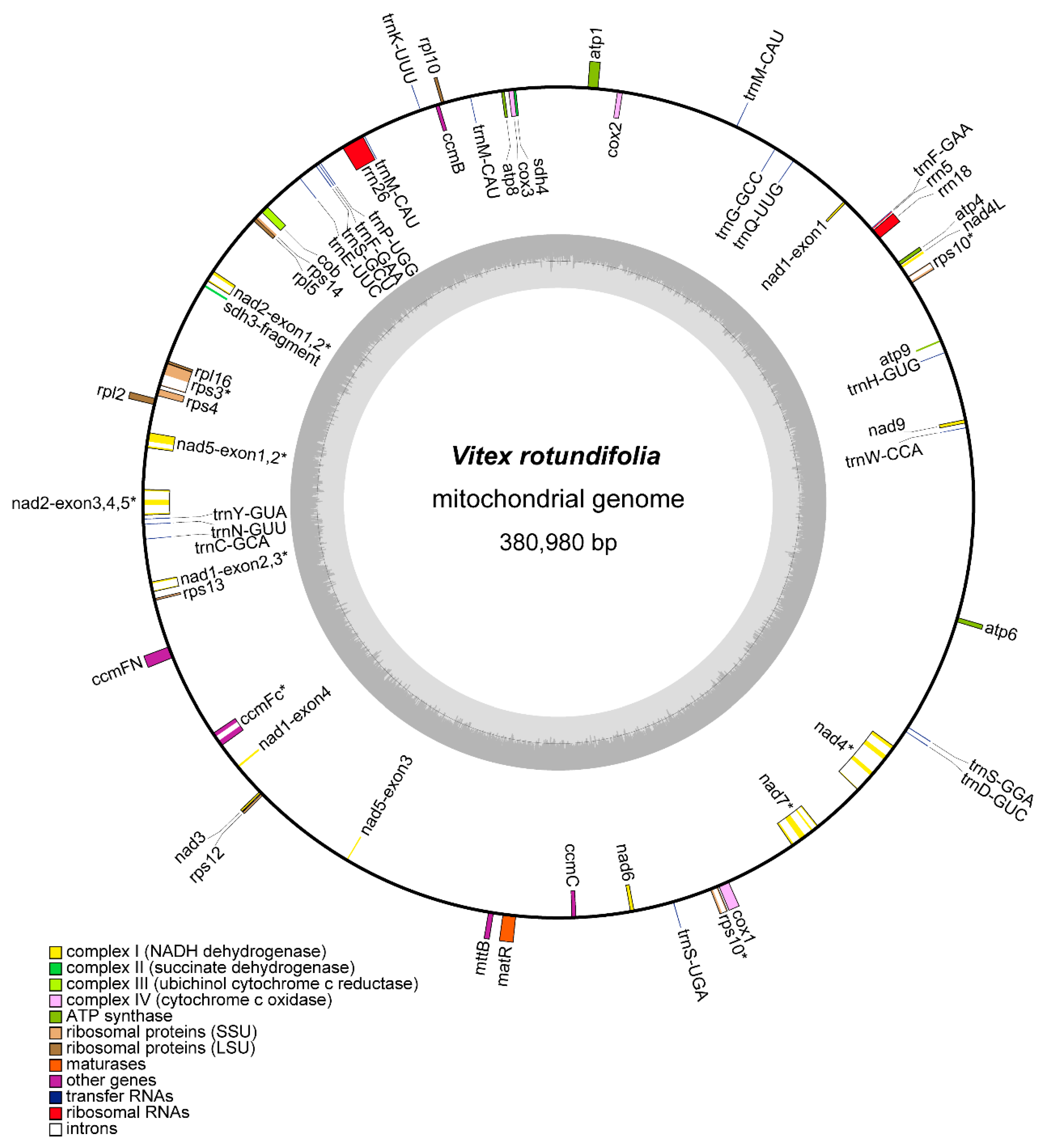
3.1. Organization and Rearrangements of the V. rotundifolia Mitochondrial Genome
3.2. Repeat Sequences and Repeat-Mediated Homologous Recombinations in the V. rotundifolia Mitochondrial Genomes
3.3. Identification of Mitochondrial Plastid Sequences (MTPT)
3.4. Phylogenetic Analysis
3.5. Identification of Genes under Selection and RNA Editing Events
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, X.-Q.; Zhang, T.; Zheng, B.; Xie, W.-D.; Shen, T. Labdane-type Diterpenoids from the Fruits of Vitex rotundifolia. Bull. Korean Chem. Soc. 2014, 35, 672–674. [Google Scholar] [CrossRef] [Green Version]
- Azizul, N.; Ahmad, W.; Rosli, N.; Azmi, M.; Liang, C.; Mazlan, N.; Assaw, S.J.T.M.R. The coastal medicinal plant Vitex rotundifolia: A mini-review on its bioactive compounds and pharmacological activity. Tradit. Med. Res. 2021, 6, 11. [Google Scholar] [CrossRef]
- Zhao, Y.-Y.; Guo, L.; Cao, L.-J.; Zhang, J.; Yin, Z.-Q. A new iridoid glycoside from the fruits of Vitex rotundifolia. Nat. Prod. Res. 2017, 31, 2491–2496. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.; Kim, H.-W.; Kim, Y.-K.; Cheon, S.-H.; Joo, M.-J.; Hong, J.-R.; Kwak, M.; Kim, K.-J. Three complete plastome sequences from the families of Lamiaceae, Mazaceae, and Phrymaceae (Lamiales). Mitochondrial DNA Part B 2021, 6, 224–226. [Google Scholar] [CrossRef]
- Terasawa, K.; Odahara, M.; Kabeya, Y.; Kikugawa, T.; Sekine, Y.; Fujiwara, M.; Sato, N. The Mitochondrial Genome of the Moss Physcomitrella patens Sheds New Light on Mitochondrial Evolution in Land Plants. Mol. Biol. Evol. 2007, 24, 699–709. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Moreno, L.; González, V.M.; Benjak, A.; Martí, M.C.; Puigdomènech, P.; Aranda, M.A.; Garcia-Mas, J. Determination of the melon chloroplast and mitochondrial genome sequences reveals that the largest reported mitochondrial genome in plants contains a significant amount of DNA having a nuclear origin. BMC Genom. 2011, 12, 424. [Google Scholar] [CrossRef] [Green Version]
- Skippington, E.; Barkman, T.J.; Rice, D.W.; Palmer, J.D. Miniaturized mitogenome of the parasitic plant Viscum scurruloideum is extremely divergent and dynamic and has lost all nad genes. Proc. Natl. Acad. Sci. USA 2015, 112, E3515-24. [Google Scholar] [CrossRef] [Green Version]
- Sloan, D.B. One ring to rule them all? Genome sequencing provides new insights into the ‘master circle’ model of plant mitochondrial DNA structure. New Phytol. 2013, 200, 978–985. [Google Scholar] [CrossRef]
- Mower, J.P.; Case, A.L.; Floro, E.R.; Willis, J.H. Evidence against Equimolarity of Large Repeat Arrangements and a Predominant Master Circle Structure of the Mitochondrial Genome from a Monkeyflower (Mimulus guttatus) Lineage with Cryptic CMS. Genome Biol. Evol. 2012, 4, 670–686. [Google Scholar] [CrossRef] [Green Version]
- Alverson, A.; Rice, D.W.; Dickinson, S.; Barry, K.; Palmer, J.D. Origins and Recombination of the Bacterial-Sized Multichromosomal Mitochondrial Genome of Cucumber. Plant Cell 2011, 23, 2499–2513. [Google Scholar] [CrossRef] [Green Version]
- Sloan, D.B.; Alverson, A.J.; Chuckalovcak, J.P.; Wu, M.; McCauley, D.E.; Palmer, J.D.; Taylor, D.R. Rapid Evolution of Enormous, Multichromosomal Genomes in Flowering Plant Mitochondria with Exceptionally High Mutation Rates. PLoS Biol. 2012, 10, e1001241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; McGinnis, S.; Madden, T.L. NCBI BLAST: A better web interface. Nucleic Acids Res. 2008, 36 (Suppl. S2), W5–W9. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.-J.; Yu, W.-B.; Yang, J.-B.; Song, Y.; Depamphilis, C.W.; Yi, T.-S.; Li, D.-Z. GetOrganelle: A fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef] [PubMed]
- Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997. [Google Scholar]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The sequence alignment/map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.; Harris, N.; Gibson, M.; Chetty, R.; Lewis, S. Apollo: A community resource for genome annotation editing. Bioinformatics 2009, 25, 1836–1837. [Google Scholar] [CrossRef] [Green Version]
- Lowe, T.M.; Eddy, S.R. tRNAscan-SE: A program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res. 1997, 25, 955–964. [Google Scholar] [CrossRef]
- Lohse, M.; Drechsel, O.; Kahlau, S.; Bock, R. OrganellarGenomeDRAW—A suite of tools for generating physical maps of plastid and mitochondrial genomes and visualizing expression data sets. Nucleic Acids Res. 2013, 41, W575–W581. [Google Scholar] [CrossRef]
- Darling, A.C.E.; Mau, B.; Blattner, F.R.; Perna, N.T. Mauve: Multiple Alignment of Conserved Genomic Sequence With Rearrangements. Genome Res. 2004, 14, 1394–1403. [Google Scholar] [CrossRef] [Green Version]
- Benson, G. Tandem repeats finder: A program to analyze DNA sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Xu, Y.; Shan, Y.; Pei, X.; Yong, S.; Liu, C.; Yu, J. Assembly of the complete mitochondrial genome of an endemic plant, Scutellaria tsinyunensis, revealed the existence of two conformations generated by a repeat-mediated recombination. Planta 2021, 254, 36. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. The CIPRES science gateway: A community resource for phylogenetic analyses. In Proceedings of the 2011 TeraGrid Conference: Extreme Digital Discovery, Online, 18 July 2011; pp. 1–8. [Google Scholar]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772–777. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z. PAML 4: Phylogenetic Analysis by Maximum Likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. ggplot2. Wiley Interdiscip. Rev. Comput. Stat. 2011, 3, 180–185. [Google Scholar] [CrossRef]
- Mower, J.P. The PREP suite: Predictive RNA editors for plant mitochondrial genes, chloroplast genes and user-defined alignments. Nucleic Acids Res. 2009, 37 (Suppl. S2), W253–W259. [Google Scholar] [CrossRef]
- Hepburn, N.J.; Schmidt, D.W.; Mower, J.P. Loss of Two Introns from the Magnolia tripetala Mitochondrial cox2 Gene Implicates Horizontal Gene Transfer and Gene Conversion as a Novel Mechanism of Intron Loss. Mol. Biol. Evol. 2012, 29, 3111–3120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, W.; Grewe, F.; Fan, W.; Young, G.J.; Knoop, V.; Palmer, J.D.; Mower, J.P. Ginkgo and Welwitschia Mitogenomes Reveal Extreme Contrasts in Gymnosperm Mitochondrial Evolution. Mol. Biol. Evol. 2016, 33, 1448–1460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Zhang, R.; Yun, Q.; Xu, Y.; Zhao, G.; Liu, J.; Shi, S.; Chen, Z.; Jia, L. Comprehensive analysis of complete mitochondrial genome of Sapindus mukorossi Gaertn.: An important industrial oil tree species in China. Ind. Crops Prod. 2021, 174, 114210. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group of Genes | CM1 | CM2 |
|---|---|---|
| Complex I | nad1 *, nad2 *, nad3, nad4L, nad5, nad6 | nad4 #, nad7 #, nad9 |
| Complex II | - | - |
| Complex III | cob | - |
| Complex IV | cox2, cox3 | cox1 |
| Complex V | atp1, atp4, atp8 | atp6, atp9 |
| Cytochrome-c biogenesis | ccmB, ccmC, ccmFC #, ccmFN | - |
| SecY-independent transport | mttB | - |
| Ribosomal protein small subunit | rps3 #, rps4, rps10 #, rps12, rps13, rps14 | rps10 # |
| Ribosomal protein large subunit | rpl2, rpl5, rpl10, rpl16 | - |
| Intron maturase | matR | - |
| Ribosomal RNAs | rrn5, rrn18, rrn26 | - |
| Transfer RNA | trnC-GCA, trnE-UUC, trnF-GAA, trnG-GCC, trnK-UUU, trnM-CAU, trnM-CAU, trnM-CAU, trnN-GUU, trnP-UGG, trnS-GCU, trnS-UGA, trnY-GUA, trnK-UUU, trnQ-UUG | trnH-GUG, trnW-CCA, trnS-GGA, trnD-GUC |
| Succinate dehydrogenase | sdh3 *, sdh4 | - |
| Repeat | Length (bp) | Direction | Position | Reads Support Master Circle Conformation | Reads Support Master Circle Conformation |
|---|---|---|---|---|---|
| R1 | 5616 | + | 1–5616 | 40 | 48 |
| + | 265,249–270,864 | 45.45% | 54.55% | ||
| R2 | 4148 | + | 30,846–34,992 | 66 | 64 |
| + | 305,624–309,771 | 50.77% | 49.23% | ||
| R3 | 283 | + | 264,966–265,248 | 10 | 16 |
| + | 380,698–380,980 | 38.46% | 62% | ||
| R4 | 211 | + | 36,020–36,229 | 184 | 6 |
| + | 130,730–130,940 | 96.84% | 3% | ||
| R5 | 102 | + | 143,734–143,835 | 120 | 2 |
| + | 233,534–233,634 | 98.36% | 2% | ||
| R6 | 110 | + | 49,216–49,324 | 78 | 1 |
| + | 205,946–206,055 | 98.73% | 1.27% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, X.; Duan, Z.; Wang, Y.; Zhang, Q.; Li, W. Sequence Analysis of the Complete Mitochondrial Genome of a Medicinal Plant, Vitex rotundifolia Linnaeus f. (Lamiales: Lamiaceae). Genes 2022, 13, 839. https://doi.org/10.3390/genes13050839
Yu X, Duan Z, Wang Y, Zhang Q, Li W. Sequence Analysis of the Complete Mitochondrial Genome of a Medicinal Plant, Vitex rotundifolia Linnaeus f. (Lamiales: Lamiaceae). Genes. 2022; 13(5):839. https://doi.org/10.3390/genes13050839
Chicago/Turabian StyleYu, Xiaoli, Zhonggang Duan, Yanjun Wang, Qingxin Zhang, and Wei Li. 2022. "Sequence Analysis of the Complete Mitochondrial Genome of a Medicinal Plant, Vitex rotundifolia Linnaeus f. (Lamiales: Lamiaceae)" Genes 13, no. 5: 839. https://doi.org/10.3390/genes13050839