
miR-377 Inhibits Proliferation and Differentiation of Bovine Skeletal Muscle Satellite Cells by Targeting FHL2

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. RNA Oligonucleotides, Vectors, and Transfection
2.3. RNA Isolation and Real-Time Quantitative PCR (qRT-PCR)
2.4. Western Blotting (WB) and Immunoprecipitation (IP) Analysis
2.5. Cell Counting Kit-8 (CCK-8) Assay
2.6. 5-Ethynyl-20-Deoxyuridine (EdU) Assay
2.7. Luciferase Reporter Assay
2.8. Statistical Analysis
3. Results
3.1. Expression Pattern of miR-377 in Bovine
3.2. miR-377 Inhibit the Proliferation of Bovine SMSCs
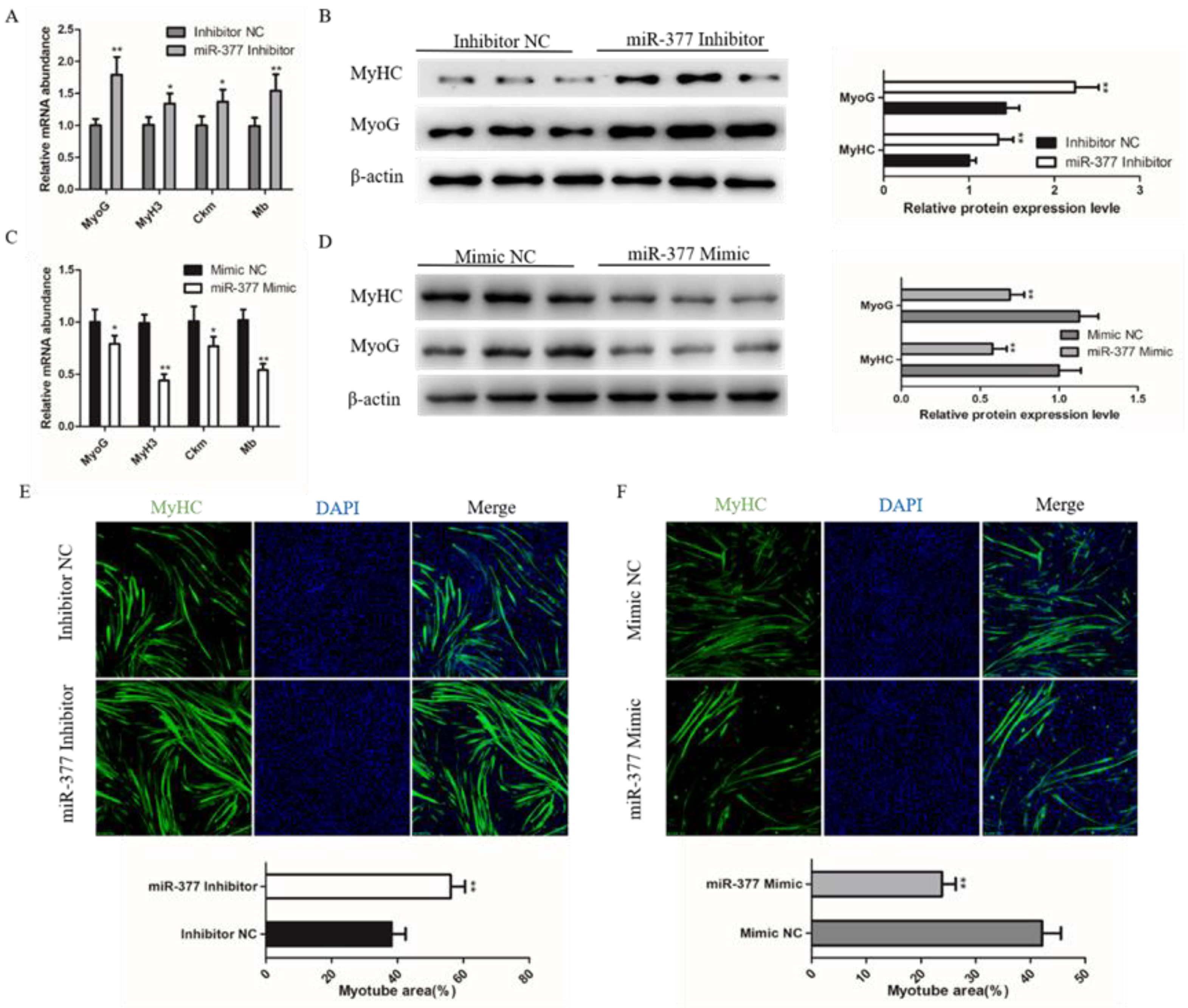
3.3. miR-377 Inhibits the Differentiation of Bovine SMSCs
3.4. miR-377 Directly Target FHL2 and Inhibit Its Expression
3.5. FHL2 Knockdown Inhibited the Proliferation of SMSCs
3.6. FHL2 Knockdown Inhibited Differentiation of SMSCs
3.7. miR-377 Disrupted the Dvl-2-Mediated Wnt Signaling Pathway in Bovine SMSCs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mogensen, M.; Sahlin, K.; Fernstroem, M.; Glintborg, D.; Vind, B.F.; Beck-Nielsen, H.; Hojlund, K. Mitochondrial Respiration Is Decreased in Skeletal Muscle of Patients with Type 2 Diabetes. Diabetes 2007, 56, 1592–1599. [Google Scholar] [CrossRef] [Green Version]
- Braun, T.; Gautel, M. Transcriptional mechanisms regulating skeletal muscle differentiation, growth and homeostasis. Nat. Rev. Mol. Cell Biol. 2011, 12, 349–361. [Google Scholar] [CrossRef]
- Lepper, C.; Conway, S.J.; Fan, C.M. Adult satellite cells and embryonic muscle progenitors have distinct genetic requirements. Nature 2009, 460, 627–631. [Google Scholar] [CrossRef] [Green Version]
- Cossu, G.; Biressi, S. Satellite cells, myoblasts and other occasional myogenic progenitors: Possible origin, phenotypic features and role in muscle regeneration. Semin. Cell Dev. Biol. 2005, 16, 606–631. [Google Scholar] [CrossRef]
- Ke, X.; He, L.; Wang, R.; Shen, J.; Wang, Z.; Shen, Y.; Fan, L.; Shao, J.; Qi, H. miR-377-3p-Mediated EGR1 Downregulation Promotes B[a]P-Induced Lung Tumorigenesis by Wnt/Beta-Catenin Transduction. Front. Oncol. 2021, 11, 699004. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Dong, X.M.; Wang, F.P. MiR-377-3p inhibits cell metastasis and epithelial-mesenchymal transition in cervical carcinoma through targeting SGK3. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 4687–4696. [Google Scholar]
- Huang, L.; Liu, Z.; Hu, J.; Luo, Z.; Zhang, C.; Wang, L.; Wang, Z. MiR-377-3p suppresses colorectal cancer through negative regulation on Wnt/β-catenin signaling by targeting XIAP and ZEB2. Pharmacol. Res. 2020, 156, 104774. [Google Scholar] [CrossRef]
- Yu, R.; Cai, L.; Chi, Y.; Ding, X.; Wu, X. miR-377 targets CUL4A and regulates metastatic capability in ovarian cancer. Int. J. Mol. Med. 2018, 41, 3147–3156. [Google Scholar] [CrossRef] [Green Version]
- Friedrich, F.W.; Reischmann, S.; Schwalm, A.; Unger, A.; Ramanujam, D.; Münch, J.; Müller, O.J.; Hengstenberg, C.; Galve, E.; Charron, P. FHL2 expression and variants in hypertrophic cardiomyopathy. Basic Res. Cardiol. 2014, 109, 451. [Google Scholar] [CrossRef] [Green Version]
- McLoughlin, P. The LIM-only Protein DRAL/FHL2 Interacts with and Is a Corepressor for the Promyelocytic Leukemia Zinc Finger Protein. J. Biol. Chem. 2002, 277, 37045–37053. [Google Scholar] [CrossRef] [Green Version]
- Arimura, T.; Hayashi, T.; Matsumoto, Y.; Shibata, H.; Hiroi, S.; Nakamura, T.; Inagaki, N.; Hinohara, K.; Takahashi, M.; Manatsu, S.I. Structural analysis of four and half LIM protein-2 in dilated cardiomyopathy. Biochem. Biophys. Res. Commun. 2007, 357, 162–167. [Google Scholar] [CrossRef]
- Hojayev, B.; Rothermel, B.A.; Gillette, T.G.; Hill, J.A. FHL2 Binds Calcineurin and Represses Pathological Cardiac Growth. Mol. Cell. Biol. 2012, 32, 4025–4034. [Google Scholar] [CrossRef] [Green Version]
- Lai, C.F.; Bai, S.; Uthgenannt, B.A.; Halstead, L.R.; Mcloughlin, P.; Schafer, B.W.; Chu, P.H.; Chen, J.; Otey, C.A.; Cao, X. Four and Half Lim Protein 2 (FHL2) Stimulates Osteoblast Differentiation. J. Bone Miner. Res. 2006, 21, 17–28. [Google Scholar] [CrossRef]
- Wei, Y.; Fabre, M.; Branchereau, S.; Gauthier, F.; Perilongo, G. Activation of β-catenin in epithelial and mesenchymal hepatoblastomas. Oncogene 2000, 19, 498–504. [Google Scholar] [CrossRef] [Green Version]
- Martin, B.; Schneider, R.; Janetzky, S.; Waibler, Z.; Pandur, P.; Kühl, M.; Behrens, J.; von der Mark, K.; Starzinski-Powitz, A.; Wixler, V. The LIM-Only Protein FHL2 Interacts with β-Catenin and Promotes Differentiation of Mouse Myoblasts. J. Cell Biol. 2002, 159, 113–122. [Google Scholar] [CrossRef]
- Algaber, A.; Madhi, R.; Hawez, A.; Rönnow, C.F.; Rahman, M. Targeting FHL2-E-cadherin axis by miR-340-5p attenuates colon cancer cell migration and invasion. Oncol. Lett. 2021, 22, 637. [Google Scholar] [CrossRef]
- Kong, L.; Zhang, C. LncRNA DLX6-AS1 aggravates the development of ovarian cancer via modulating FHL2 by sponging miR-195-5p. Cancer Cell Int. 2020, 20, 370. [Google Scholar] [CrossRef]
- Lyu, P.; Qi, Y.; Tu, Z.J.; Jiang, H. Single-cell RNA Sequencing Reveals Heterogeneity of Cultured Bovine Satellite Cells. Front. Genet. 2021, 12, 742077. [Google Scholar] [CrossRef]
- Wong, C.H.; Mak, W.Y.; Li, M.S.; Tsui, K.W. The LIM-only protein FHL2 regulates interleukin-6 expression through p38 MAPK mediated NF-κB pathway in muscle cells. Cytokine 2012, 59, 286–293. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402. [Google Scholar] [CrossRef]
- Wang, H.; Wei, Z.; Li, H.; Guan, Y.; Han, Z.; Wang, H.; Liu, B. MiR-377-3p inhibits atherosclerosis-associated vascular smooth muscle cell proliferation and migration via targeting neuropilin2. Biosci. Rep. 2020, 40, BSR20193425. [Google Scholar] [CrossRef]
- Zhang, P.; Wang, W.; Li, M. Circ_0010283/miR-377-3p/Cyclin D1 Axis Is Associated with Proliferation, Apoptosis, Migration, and Inflammation of Oxidized Low-density Lipoprotein-Stimulated Vascular Smooth Muscle Cells. J. Cardiovasc. Pharmacol. 2021, 78, 437–447. [Google Scholar] [CrossRef]
- Bachman, J.F.; Klose, A.; Liu, W.; Paris, N.D.; Blanc, R.S.; Schmalz, M.; Knapp, E.; Chakkalakal, J.V. Prepubertal skeletal muscle growth requires Pax7-expressing satellite cell-derived myonuclear contribution. Development 2018, 145, dev167197. [Google Scholar] [CrossRef] [Green Version]
- Dumont, N.A.; Bentzinger, C.F.; Sincennes, M.C.; Rudnicki, M.A. Satellite Cells and Skeletal Muscle Regeneration. Compr. Physiol. 2015, 5, 1027–1059. [Google Scholar]
- Liu, Z.; Han, S.; Wang, Y.; Cui, C.; Zhu, Q.; Jiang, X.; Yang, C.; Du, H.; Yu, C.; Li, Q.; et al. The LIM-Only Protein FHL2 is involved in Autophagy to Regulate the Development of Skeletal Muscle Cell. Int. J. Biol. Sci. 2019, 15, 838–846. [Google Scholar] [CrossRef]
- Shi, X.; Bowlin, K.M.; Garry, D.J. Fhl2 interacts with Foxk1 and corepresses Foxo4 activity in myogenic progenitors. Stem Cells 2010, 28, 462–469. [Google Scholar] [CrossRef]
- Han, X.H.; Jin, Y.R.; Seto, M.; Yoon, J.K. A WNT/β-Catenin Signaling Activator, R-spondin, Plays Positive Regulatory Roles during Skeletal Myogenesis. J. Biol. Chem. 2012, 286, 10649. [Google Scholar] [CrossRef] [Green Version]
- Brun, J.; Dieudonné, F.X.; Marty, C.; Müller, J.; Schüle, R.; Patiño-García, A.; Lecanda, F.; Fromigué, O.; Marie, P.J. FHL2 Silencing Reduces Wnt Signaling and Osteosarcoma Tumorigenesis In Vitro and In Vivo. PLoS ONE 2013, 8, e55034. [Google Scholar] [CrossRef] [Green Version]
- Cy, L.; Wang, L.; Zj, L.; Gao, P.; Af, H.; Yuan, Z. Plumbagin Shows Anti-proliferative Effect in Human Osteosarcoma Cells by Down Regulating FHL2 and Interfering with WNT/Β-Catenin Signalling. West Indian Med. J. 2016, 12, 1095–1100. [Google Scholar]
- Duan, Y.; Qiu, Y.; Huang, X.; Dai, C.; He, W. Deletion of FHL2 in fibroblasts attenuates fibroblasts activation and kidney fibrosis via restraining TGF-β1-induced Wnt/β-catenin signaling. J. Mol. Med. 2020, 98, 291–307. [Google Scholar] [CrossRef]
- Li, S.-Y.; Huang, P.-H.; Tarng, D.-C.; Lin, T.-P.; Yang, W.-C. Four-and-a-Half LIM Domains Protein 2 Is a Coactivator of Wnt Signaling in Diabetic Kidney Disease. J. Am. Soc. Nephrol. JASN 2015, 26, 3072–3084. [Google Scholar] [CrossRef] [Green Version]
- Pulvirenti, T.; Van Der Heijden, M.; Droms, L.A.; Huse, J.T.; Tabar, V.; Hall, A. Dishevelled 2 Signaling Promotes Self-Renewal and Tumorigenicity in Human Gliomas. Cancer Res. 2011, 71, 7280–7290. [Google Scholar] [CrossRef] [Green Version]
- Mcloon, L.K.; Rowe, J.; Wirtschafter, J.; Mccormick, K.M. Continuous myofiber remodeling in uninjured extraocular myofibers: Myonuclear turnover and evidence for apoptosis. Muscle Nerve 2004, 29, 707–715. [Google Scholar] [CrossRef] [Green Version]
- Ozbek, E.; Adas, G.; Otunctemur, A.; Duruksu, G.; Karaoz, E. Role of Mesenchymal Stem Cells Transfected with Vascular Endothelial Growth Factor in Maintaining Renal Structure and Function in Rats with Unilateral Ureteral Obstruction. Exp. Clin. Transplant. 2014, 13, 262. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Forward Primer (5′–3′) |
|---|---|
| FHL2-shRNA | GCAAGGACUUGUCUUAUAATT |
| UUAUAAGACAAGUCCUUGCTT | |
| MiR-377 mimic | AUCACACAAAGGCAACUUUUGU |
| Mimic NC | UUGUACUACACAAAAGUACUG |
| MiR-377 inhibitor | ACAAAAGUUGCCUUUGUGUGAU |
| Inhibitor NC | CAGUACUUUUGUGUAGUACAA |
| Genes | Forward Primer (5′–3′) | Size (bp) | Annealing Temperature (°C) | Accession Number |
|---|---|---|---|---|
| FHL2 | F: CTCTGCGCTTCTCAGCGATA | 128 | 61 | NM_001046046.2 |
| R: GGCAGGAAGTTACACCGGAA | ||||
| Ckm | F: CCTGACGGGTGAGTTCAAGG | 187 | 60 | NM_174773.4 |
| R: TGATCCTCCTCGTTCACCCA | ||||
| MyoG | F: TGGGCGTGTAAGGTGTGTAA | 78 | 60 | NM_001111325.1 |
| R: TATGGGAGCTGCATTCACTG | ||||
| MyH7 | F: CTTCAACCACCACATGTTCG | 178 | 58 | NM_174727.1 |
| R: GCTTCTGGAAGTTGCTGGAC | ||||
| Wnt5a | F: CAACTGGCAGGACTTTCTCAA | 127 | 61 | NM_174727.1 |
| R: CATCTCCGATGCCGGAACT | ||||
| Wnt10b | F: CTCTGCCACAGCCAAACTCT | 106 | 60 | XM_010805029.3 |
| R: ATCGAACTTGCCTGGCTTGA | ||||
| c-Myc | F: GTAATTCCAGCGAGAGGCAGA | 213 | 60 | NM_001046074.2 |
| R: CTAGGCTAGCTCGGCTCTTC | ||||
| Lef1 | F: CCCTGTGTTGTTCGGCCTC | 271 | 59 | NM_001192856.1 |
| R: ATTGGAAGGATGCGTCAGGG | ||||
| CyclinD1 | F: ATGAAGGAGACCATCCCCCT | 117 | 61 | NM_001046273.2 |
| R: CGCCAGGTTCCACTTGAGTT | ||||
| CDK2 | F: AGGGAACGTACGGAGTTGTG | 78 | 58 | NM_001014934.1 |
| R: GACATCCAGCAGCTTGACAAT | ||||
| PCNA | F: TCCAGAACAAGAGTATAGC | 162 | 60 | NM_001034494.1 |
| R: TACAACAGCATCTCCAAT | ||||
| miR-377 | F: ATCACACAAAGGCAACTTTTGT | / | 60 | / |
| R: CAGGTCCAGTTTTTTTTTTTTTT | ||||
| MB | F: ACTGACCTGCACCTTTACCC | 210 | 62 | NM_173881.2 |
| R: CTCAGGGCAAGCAAGACACT | ||||
| β-actin | F: CATCCTGACCCTCAAGTA | 146 | 56–62 | NM_173979.3 |
| R: CTCGTTGTAGAAGGTGTG | ||||
| U6 | F: GCTTCGGCAGCACATATACTAAAAT | / | 60 | / |
| R: CGCTTCACGAATTTGCGTGTCAT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Y.; Li, P.; Dan, X.; Kang, X.; Ma, Y.; Shi, Y. miR-377 Inhibits Proliferation and Differentiation of Bovine Skeletal Muscle Satellite Cells by Targeting FHL2. Genes 2022, 13, 947. https://doi.org/10.3390/genes13060947
Zhu Y, Li P, Dan X, Kang X, Ma Y, Shi Y. miR-377 Inhibits Proliferation and Differentiation of Bovine Skeletal Muscle Satellite Cells by Targeting FHL2. Genes. 2022; 13(6):947. https://doi.org/10.3390/genes13060947
Chicago/Turabian StyleZhu, Yun, Peng Li, Xingang Dan, Xiaolong Kang, Yun Ma, and Yuangang Shi. 2022. "miR-377 Inhibits Proliferation and Differentiation of Bovine Skeletal Muscle Satellite Cells by Targeting FHL2" Genes 13, no. 6: 947. https://doi.org/10.3390/genes13060947
APA StyleZhu, Y., Li, P., Dan, X., Kang, X., Ma, Y., & Shi, Y. (2022). miR-377 Inhibits Proliferation and Differentiation of Bovine Skeletal Muscle Satellite Cells by Targeting FHL2. Genes, 13(6), 947. https://doi.org/10.3390/genes13060947