
Genome-Wide Identification and Analysis of the GRAS Transcription Factor Gene Family in Theobroma cacao
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of GRAS Gene Family in T. cacao
2.2. Physicochemical Properties and Subcellular Localization Analyses of TcGRAS Genes
2.3. Phylogenetic Analysis and Classification of TcGRAS Genes
2.4. Gene Structure and Conserved Motif Analyses of TcGRAS Genes
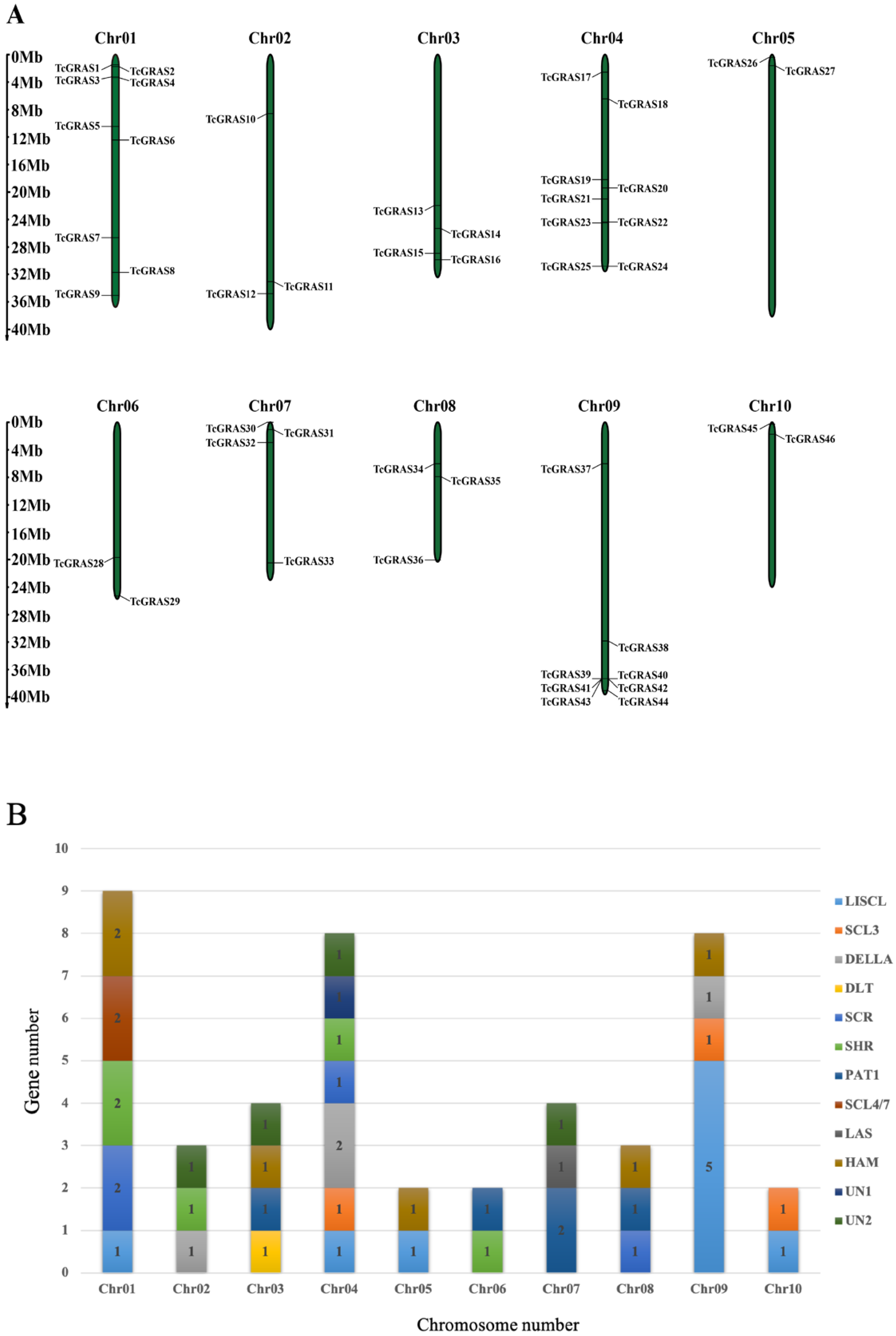
2.5. Chromosomal Mapping and Cis-Acting Regulatory Analyses of TcGRAS Genes
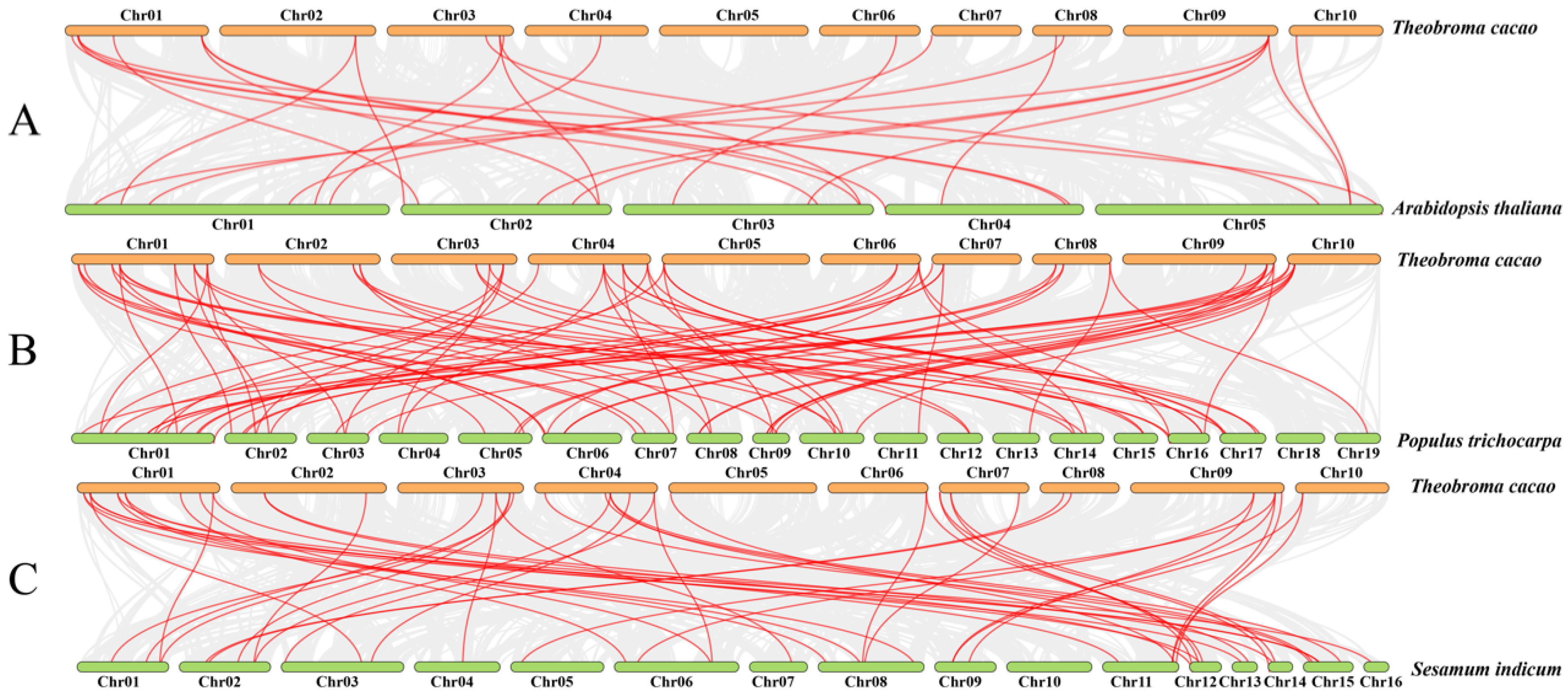
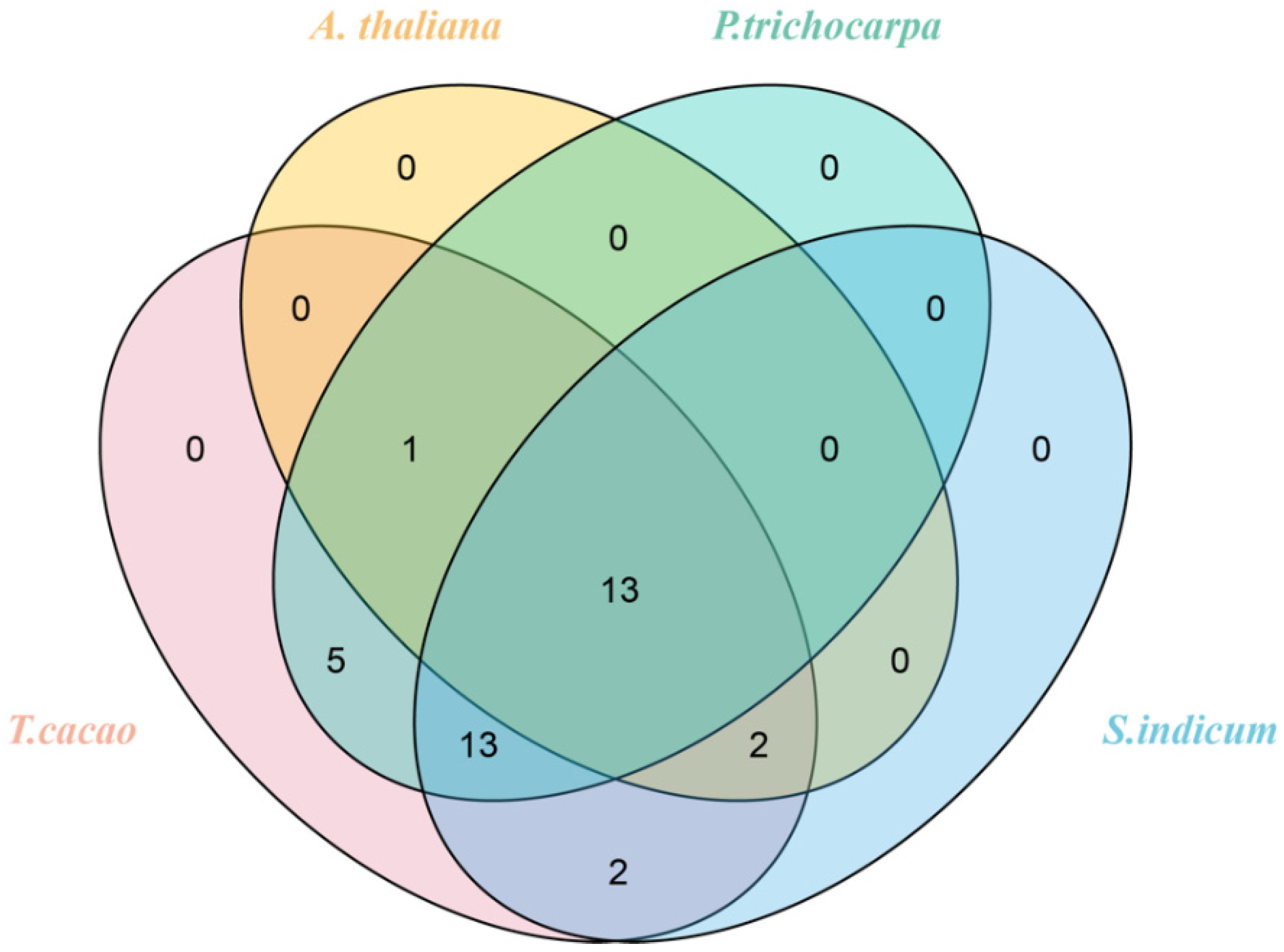
2.6. Gene Duplication and Synteny Analyses of TcGRAS Genes
2.7. GO Annotation Analyses of T. cacao TcGRAS Genes
3. Results
3.1. Identification of GRAS Members in T. cacao
3.2. Phylogenetic Analysis of TcGRAS and AtGRAS
3.3. Gene Structure, Conserved Motifs, and Domain Analyses of TcGRAS Genes
3.4. Chromosomal Mapping and Cis-Acting Regulatory Analyses of TcGRAS Genes
3.5. Gene Duplication and Syntenic Analysis of TcGRAS Genes
3.6. GO Annotation of T.cacao TcGRAS Proteins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Riano-Pachon, D.M.; Ruzicic, S.; Dreyer, I.; Mueller-Roeber, B. PlnTFDB: An integrative plant transcription factor database. BMC Bioinform. 2007, 8, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Sun, Y.; Xue, J.; Jia, X.; Li, R. Genome-wide characterization and expression analysis of GRAS gene family in pepper (Capsicum annuum L.). PeerJ 2018, 6, e4796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.; Xuan, A.; Bu, C.; Ci, D.; Tian, M.; Zhang, D. Osmotic stress-responsive promoter upstream transcripts (PROMPTs) act as carriers of MYB transcription factors to induce the expression of target genes in Populus simonii. Plant Biotechnol. J. 2019, 17, 164–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laurenzio, L.D.; Wysocka-Diller, J.; Malamy, J.E.; Pysh, L.; Helariutta, Y.; Freshour, K.; Hahn, M.G.; Feldmann, K.A.; Benfey, P.N. The SCARECROW gene regulates an asymmetric cell division that is essential for generating the radial organization of the Arabidopsis root. Cell 1996, 86, 423–433. [Google Scholar] [CrossRef] [Green Version]
- Peng, J.; Carol, P.; Richards, D.E.; King, K.E.; Cowling, R.J.; Murphy, G.P.; Harberd, P.H. The Arabidopsis GAI gene defines a signaling pathway that negatively regulates gibberellin responses. Genes Dev. 1997, 11, 3194–3205. [Google Scholar] [CrossRef] [Green Version]
- Silverstone, A.L.; Ciampaglio, C.N.; Sun, T. The Arabidopsis RGA gene encodes a transcriptional regulator repressing the gibberellin signal transduction pathway. Plant Cell 1998, 10, 155–169. [Google Scholar] [CrossRef] [Green Version]
- Bolle, C. The role of GRAS proteins in plant signal transduction and development. Planta 2004, 218, 683–692. [Google Scholar] [CrossRef]
- Tian, C.; Wan, P.; Sun, S.; Li, J.; Chen, M. Genome-wide analysis of the GRAS gene family in Rice and Arabidopsis. Plant Mol. Biol. 2004, 54, 519–532. [Google Scholar] [CrossRef]
- Liu, X.; Widmer, A. Genome-wide comparative analysis of the GRAS gene family in Populus, Arabidopsis and Rice. Plant Mol. Biol. Rep. 2014, 32, 1129–1145. [Google Scholar] [CrossRef]
- Wang, T.; Yu, T.; Fu, J.; Su, H.; Chen, J.; Zhou, Y.; Chen, M.; Guo, J.; Ma, Y.; Wei, W.; et al. Genome-wide analysis of the GRAS gene family and functional identification of GmGRAS37 in drought and salt tolerance. Front. Plant Sci. 2020, 11, 604690. [Google Scholar] [CrossRef]
- Huang, W.; Xian, Z.; Kang, X.; Tang, N.; Li, Z. Genome-wide identification, phylogeny and expression analysis of GRAS gene family in tomato. BMC Plant Biol. 2015, 15, 209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Z.J.; Song, N.; Peng, W.Y.; Yang, Y.; Qiu, T.; Huang, C.T.; Dai, L.Y.; Wang, B. Genome identification and expression analysis of GRAS family related to development, hormone and pathogen stress in Brachypodium distachyon. Front. Sustain. Food Syst. 2021, 5, 675177. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, N.; Zhu, X.; Yang, J.; Li, S.; Che, Y.; Liu, W.; Si, H. Identification and expression analysis of StGRAS gene family in potato (Solanum tuberosum L.). Comput. Biol. Chem. 2019, 80, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Duan, X.; Jiang, H.; Sun, Y.; Tang, Y.; Yuan, Z.; Guo, J.; Liang, W.; Chen, L.; Yin, J.; et al. Genome- wide analysis of basic/helix-loop-helix transcription factor family in rice and Arabidopsis. Plant Physiol. 2006, 141, 1167–1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirsch, S.; Oldroyd, G.E. GRAS-domain transcription factors that regulate plant development. Plant Signal. Behav. 2009, 4, 698–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Jones, W.T.; Rikkerink, E.H. GRAS proteins: The versatile roles of intrinsically disordered proteins in plant signalling. Biochem. J. 2012, 442, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Stuurman, J.; Jäggi, F.; Kuhlemeier, C. Shoot meristem maintenance is controlled by a GRAS-gene mediated signal from differentiating cells. Genes Dev. 2002, 16, 2213–2218. [Google Scholar] [CrossRef] [Green Version]
- Morohashi, K.; Minami, M.; Takase, H.; Hotta, Y.; Hiratsuka, K. Isolation and characterization of a novel GRAS gene that regulates meiosis-associated gene expression. J. Biol. Chem. 2003, 278, 20865–20873. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, K.; Schmitt, T.; Rossberg, M.; Schmitz, G.; Theres, K. The Lateral suppressor (Ls) gene of tomato encodes a new member of the VHIID protein family. Proc. Natl. Acad. Sci. USA 1999, 96, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.; Liang, D.; Shuai, P.; Xia, X.; Yin, W. The salt- and drought-inducible poplar GRAS protein SCL7 confers salt and drought tolerance in Arabidopsis thaliana. J. Exp. Bot. 2010, 61, 4011–4019. [Google Scholar] [CrossRef] [PubMed]
- Helariutta, Y.; Fukaki, H.; Wysocka-Diller, J.; Nakajima, K.; Jung, J.; Sena, G.; Hauser, M.T.; Benfey, P.N. The SHORT-ROOT gene controls radial patterning of the Arabidopsis root through radial signaling. Cell 2000, 101, 555–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Andersson-Gunneras, S.; Gaboreanu, I.; Hertzberg, M.; Tucker, M.R.; Zheng, B.; Lesniewska, J.; Mellerowicz, E.J.; Laux, T.; Sandberg, G.; et al. Reduced expression of the SHORT-ROOT gene increases the rates of growth and development in hybrid poplar and Arabidopsis. PLoS ONE 2011, 6, e28878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, H.; Jin, Y.; Liu, W.; Li, F.; Fang, J.; Yin, Y.; Qian, Q.; Zhu, L.; Chu, C. DWARF AND LOW-TILLERING, a new member of the GRAS family, plays positive roles in brassinosteroid signaling in rice. Plant J. 2009, 58, 803–816. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.O.; Chang, K.S.; Kim, I.A.; Lee, M.H.; Lee, S.A.; Song, S.K.; Lee, M.M.; Lim, J. Funneling of gibberellin signaling by the GRAS transcription regulator SCARECROW-LIKE 3 in the Arabidopsis root. Proc. Natl. Acad. Sci. USA 2011, 108, 2166–2171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, X.; Lee, L.; Xia, K.; Yan, Y.; Yu, H. DELLAs modulate jasmonate signaling via competitive binding to JAZs. Dev. Cell 2010, 19, 884–894. [Google Scholar]
- Bolle, C.; Koncz, C.; Chua, N.H. PAT1, a new member of the GRAS family, is involved in phytochrome A signal transduction. Genes Dev. 2000, 14, 1269–1278. [Google Scholar] [CrossRef]
- Mustiga, G.M.; Gezan, S.A.; Phillips-Mora, W.; Arciniegas-Leal, A.; Mata-Quirós, A.; Motamayor, J.C. Phenotypic description of Theobroma cacao L. for yield and vigor traits from 34 hybrid families in Costa Rica based on the genetic basis of the parental population. Front. Plant Sci. 2018, 9, 808. [Google Scholar] [CrossRef] [Green Version]
- Corti, R.; Flammer, A.J.; Hollenberg, N.K.; Lüscher, T.F. Cocoa and cardiovascular health. Circulation 2009, 119, 1433–1441. [Google Scholar] [CrossRef] [Green Version]
- Argout, X.; Salse, J.; Aury, J.M.; Guiltinan, M.J.; Droc, G.; Gouzy, J.; Allegre, M.; Chaparro, C.; Legavre, T.; Maximova, S.N. The genome of Theobroma cacao. Nat. Genet. 2011, 3, 101–108. [Google Scholar] [CrossRef]
- Shen, S.; Zhang, Q.; Shi, Y.; Sun, Z.; Zhang, Q.; Hou, S.; Wu, R.; Jiang, L.; Zhao, X.; Guo, Y. Genome-wide analysis of the NAC Domain transcription factor gene family in Theobroma cacao. Genes 2019, 11, 35. [Google Scholar] [CrossRef] [Green Version]
- Silva Monteiro de Almeida, D.; Oliveira Jordão do Amaral, D.; Del-Bem, L.E.; Bronze Dos Santos, E.; Santana Silva, R.J.; Peres Gramacho, K.; Vincentz, M.; Micheli, F. Genome-wide identification and characterization of cacao WRKY transcription factors and analysis of their expression in response to witches’ broom disease. PLoS ONE 2017, 12, e0187346. [Google Scholar] [CrossRef] [PubMed]
- Martins Alves, A.M.; Pereira Menezes Reis, S.; Peres Gramacho, K.; Micheli, F. The glutathione peroxidase family of Theobroma cacao: Involvement in the oxidative stress during witches’ broom disease. Int. J. Biol. Macromol. 2020, 164, 3698–3708. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Wu, B.; Yan, L.; Qin, X.; Lai, J. Metabolome and transcriptome profiling of Theobroma cacao provides insights into the molecular basis of pod color variation. J. Plant Res. 2021, 134, 1323–1334. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Wang, T.; Xu, Z.; Sun, L.; Zhang, Q. Genome-wide analysis of the GRAS gene family in Prunus mume. Mol. Gen. Genomics 2015, 290, 303–317. [Google Scholar] [CrossRef] [PubMed]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A. The Pfam protein families database in 2019. Nucleic Acids Res. 2019, 47, D427–D432. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Copley, R.R.; Schmidt, S.; Ciccarelli, F.D.; Doerks, T.; Schultz, J.; Ponting, C.P.; Bork, P. SMART 4.0: Towards genomic data integration. Nucleic Acids Res. 2004, 32, D142–D144. [Google Scholar] [CrossRef] [Green Version]
- Wilkins, M.R.; Gasteiger, E.; Bairoch, A.; Sanchez, J.C.; Hochstrasser, D.F. Protein identification and analysis tools in the ExPASy server. Methods Mol. Biol. 1999, 112, 531–552. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [Green Version]
- Pysh, L.D.; Wysocka-Diller, J.W.; Camilleri, C.; Bouchez, D.; Benfey, P.N. The GRAS gene family in Arabidopsis: Sequence characterization and basic expression analysis of the SCARECROW-LIKE genes. Plant J. 1999, 18, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Letunic, B. Interactive Tree of Life (iTOL): An online tool for phylogenetic tree display and annotation. Bioinformatics 2007, 23, 127–128. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.Y.; Li, W.F.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.P.; Zhang, Y.B.; Zhang, Z.; Zhu, J.; Yu, J. KaKs_Calculator 2.0: A Toolkit Incorporating Gamma-Series Methods and Sliding Window Strategies. Genom. Proteom. Bioinf. 2010, 8, 77–80. [Google Scholar] [CrossRef] [Green Version]
- Yu, G. Using ggtree to visualize data on tree-like structures. Curr. Protoc. Bioinform. 2020, 69, e96. [Google Scholar] [CrossRef]
- Roy, S.W.; Penny, D. Patterns of intron loss and gain in plants: Intron loss-dominated evolution and genome-wide comparison of O. sativa and A. thaliana. Mol. Biol. Evol. 2007, 24, 171–181. [Google Scholar] [CrossRef] [Green Version]
- Rogozin, I.B.; Carmel, L.; Csuros, M.; Koonin, E.V. Origin and evolution of spliceosomal introns. Biol. Direct. 2012, 7, 11. [Google Scholar] [CrossRef] [Green Version]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef] [Green Version]
- Hanada, K.; Zou, C.; Lehti-Shiu, M.D.; Shinozaki, K.; Shiu, S.H. Importance of lineage-specific expansion of plant tandem duplicates in the adaptive response to environmental stimuli. Plant Physiol. 2008, 148, 993–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, S.Y.; Ma, Z.; Ramachandran, S. Evolutionary history and stress regulation of the lectin superfamily in higher plants. BMC Evol. Biol. 2010, 10, 79. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.L.; Geisler, M. Genome-wide computational identification of biologically significant cis-regulatory elements and associated transcription factors from rice. Plants 2019, 8, 441. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Subfamily | Function | Species | Classified Subfamilies | Reference |
|---|---|---|---|---|---|
| HAM | HAM | Maintenance of stems | Petunia hybrida | unclassified | [17] |
| LISCL | LISCL | Regulating the transcription process during microsporogenesis | Lilium longiflorum | unclassified | [18] |
| Ls | LAS | Initiation of the axillary meristem | tomato | HAM, LAS, SCL4/7, SCR, SCL9, SCL28, DELLA, SHR, PAT1, Os4, Os19, GRAS37, and Pt20 | [19] |
| PeSCL7 | SCL4/7 | Enhanced drought tolerance and salt tolerance of transgenic Arabidopsis plants | Populus euphratica | unclassified | [20] |
| AtSCR | SCR | Involved in radial root morphology and growth | A. thaliana | HAM, LISCL, LAS, SCL4/7, SCR, DLT, SCL3, DELLA, SHR, and PAT1 | [4] |
| AtSHR | SHR | Involved in radial root morphology and growth | A. thaliana | HAM, LISCL, LAS, SCL4/7, SCR, DLT, SCL3, DELLA, SHR, and PAT1 | [21] |
| PtSHR1 | SHR | Increased growth rates | Populus tomentosa | Os19, HAM, Os4, Pt20, DLT, AtSCl3, AtSHR, AtPAT1, AtSCR, AtSCL4/7, AtLAS, DELLA, and LISCL | [22] |
| DLT | DLT | Involved in brassinoltone signaling | rice | LISCL, SHR, DELLA, SCL3, PAT1, SCR, SCL4/7, LAS, Os19, HAM, Os4, and DLT | [23] |
| AtSCL3 | SCL3 | Integrated multiple signals during Arabidopsis root cell elongation | A. thaliana | HAM, LISCL, LAS, SCL4/7, SCR, DLT, SCL3, DELLA, SHR, and PAT1 | [24] |
| AtRGA | DELLA | Modulated jasmonate signaling via competitive binding to JAZs | A. thaliana | HAM, LISCL, LAS, SCL4/7, SCR, DLT, SCL3, DELLA, SHR, and PAT1 | [25] |
| BdSLR1 BdSLRL1 | DELLA | Play a role in plant growth via the GA signal pathway | Brachypodium distachyon | HAM, PAT1, SHR, DELLA, SCL3, SCL4/7, LAS, SCR, DLT, and LISCL | [12] |
| AtPAT1 | PAT1 | Involved in signaling in Arabidopsis photochromes | A. thaliana | HAM, LISCL, LAS, SCL4/7, SCR, DLT, SCL3, DELLA, SHR, and PAT1 | [26] |
| GmGRAS37 | PAT1 | Improved resistance to drought and salt stresses | soybean | AtSCL4/7, Os19, Os4, HAM, DELLA, DLT, AtPAT1, LISCL, AtSCR, AtSCL3, and AtSHR | [10] |
| StGRAS9 | PAT1 | Responded to plant hormones IAA, ABA, and GA3 treatment | potato | DELLA, LAS, HAM, PATI, SCR, LISCL, SHR, and SCL3 | [13] |
| Gene Name | Gene ID | Physicochemical Characteristics | SL | ORF | ||||
|---|---|---|---|---|---|---|---|---|
| PI | MW (Da) | Length (aa) | Instability Index | Aliphatic Index | ||||
| TcGRAS1 | TCM_000399 | 5.00 | 67,286.55 | 608 | 54.24 | 81.73 | endomembrane system | 1827 |
| TcGRAS2 | TCM_000435 | 6.80 | 77,476.49 | 684 | 50.67 | 84.71 | nucleus | 2055 |
| TcGRAS3 | TCM_000764 | 5.46 | 50,905.28 | 456 | 41.68 | 92.43 | chloroplast | 1371 |
| TcGRAS4 | TCM_000801 | 5.59 | 57,001.87 | 505 | 52.26 | 70.32 | chloroplast | 1518 |
| TcGRAS5 | TCM_002021 | 5.42 | 48,046.37 | 441 | 56.70 | 91.54 | nucleus | 1326 |
| TcGRAS6 | TCM_002319 | 5.57 | 63,858.30 | 569 | 55.85 | 79.17 | nucleus | 1710 |
| TcGRAS7 | TCM_003984 | 5.84 | 57,336.13 | 511 | 44.83 | 83.99 | nucleus | 1536 |
| TcGRAS8 | TCM_004818 | 6.15 | 63,796.67 | 565 | 31.39 | 85.91 | nucleus | 1698 |
| TcGRAS9 | TCM_005571 | 5.41 | 79,449.91 | 730 | 62.24 | 84.47 | nucleus | 2193 |
| TcGRAS10 | TCM_007806 | 5.31 | 49,737.63 | 445 | 43.06 | 88.97 | chloroplast | 1338 |
| TcGRAS11 | TCM_010708 | 6.38 | 56,277.83 | 499 | 47.40 | 92.06 | nucleus | 1500 |
| TcGRAS12 | TCM_010965 | 5.66 | 67,186.67 | 615 | 39.21 | 80.93 | nucleus | 1848 |
| TcGRAS13 | TCM_014574 | 6.04 | 57,510.78 | 519 | 52.58 | 90.02 | nucleus | 1560 |
| TcGRAS14 | TCM_015228 | 5.76 | 67,975.59 | 628 | 45.58 | 77.82 | nucleus | 1887 |
| TcGRAS15 | TCM_015991 | 5.69 | 87,113.44 | 795 | 55.54 | 80.34 | nucleus | 2388 |
| TcGRAS16 | TCM_016186 | 5.68 | 74,493.64 | 671 | 52.33 | 79.79 | nucleus | 2016 |
| TcGRAS17 | TCM_017269 | 5.67 | 72,460.97 | 654 | 53.85 | 78.35 | nucleus | 1965 |
| TcGRAS18 | TCM_017746 | 5.88 | 86,321.23 | 795 | 53.09 | 81.66 | nucleus | 2388 |
| TcGRAS19 | TCM_018964 | 6.92 | 46,686.93 | 418 | 44.25 | 94.07 | nucleus | 1257 |
| TcGRAS20 | TCM_019165 | 5.27 | 71,951.43 | 655 | 49.47 | 81.62 | nucleus | 1968 |
| TcGRAS21 | TCM_019414 | 5.43 | 39,709.90 | 347 | 49.00 | 94.96 | nucleus | 1044 |
| TcGRAS22 | TCM_019956 | 6.08 | 191,183.51 | 1659 | 47.01 | 80.49 | nucleus | 4980 |
| TcGRAS23 | TCM_019978 | 5.82 | 51,125.89 | 457 | 46.55 | 79.63 | chloroplast | 1374 |
| TcGRAS24 | TCM_021350 | 5.67 | 50,849.34 | 457 | 33.62 | 93.09 | nucleus | 1374 |
| TcGRAS25 | TCM_021351 | 5.08 | 67,410.79 | 600 | 47.40 | 83.42 | nucleus | 1803 |
| TcGRAS26 | TCM_021618 | 4.86 | 61,003.74 | 538 | 49.81 | 70.91 | nucleus | 1617 |
| TcGRAS27 | TCM_021920 | 6.17 | 85,337.10 | 755 | 52.92 | 73.09 | nucleus | 2268 |
| TcGRAS28 | TCM_029136 | 6.30 | 64,717.65 | 582 | 46.26 | 74.60 | nucleus | 1749 |
| TcGRAS29 | TCM_030393 | 5.89 | 61,290.47 | 548 | 52.83 | 79.00 | nucleus | 1647 |
| TcGRAS30 | TCM_030498 | 6.19 | 49,455.27 | 438 | 55.76 | 96.74 | nucleus | 1317 |
| TcGRAS31 | TCM_030733 | 5.56 | 60,140.03 | 540 | 58.44 | 79.35 | nucleus | 1623 |
| TcGRAS32 | TCM_031132 | 6.13 | 66,239.71 | 596 | 52.40 | 80.54 | nucleus | 1791 |
| TcGRAS33 | TCM_033446 | 4.94 | 58,582.29 | 521 | 39.07 | 86.91 | nucleus | 1566 |
| TcGRAS34 | TCM_035069 | 5.28 | 53,980.68 | 487 | 40.15 | 85.15 | nucleus | 1464 |
| TcGRAS35 | TCM_035362 | 5.06 | 63,856.24 | 570 | 49.25 | 82.14 | nucleus | 1713 |
| TcGRAS36 | TCM_036707 | 6.31 | 52,208.63 | 457 | 43.57 | 86.81 | nucleus | 1374 |
| TcGRAS37 | TCM_037975 | 5.62 | 60,414.53 | 548 | 49.30 | 87.08 | nucleus | 1647 |
| TcGRAS38 | TCM_040833 | 5.61 | 52,722.63 | 470 | 50.42 | 98.81 | nucleus | 1413 |
| TcGRAS39 | TCM_041810 | 6.33 | 79,445.42 | 705 | 46.24 | 71.65 | nucleus | 2118 |
| TcGRAS40 | TCM_041812 | 6.17 | 78,589.84 | 690 | 51.06 | 80.39 | nucleus | 2073 |
| TcGRAS41 | TCM_041813 | 5.40 | 75,260.51 | 666 | 53.55 | 81.14 | nucleus | 2001 |
| TcGRAS42 | TCM_041814 | 5.16 | 89,125.20 | 790 | 45.83 | 71.96 | nucleus | 2373 |
| TcGRAS43 | TCM_041815 | 6.30 | 92,905.70 | 829 | 46.81 | 70.95 | nucleus | 2490 |
| TcGRAS44 | TCM_042194 | 4.63 | 60,211.98 | 537 | 45.66 | 85.74 | nucleus | 1614 |
| TcGRAS45 | TCM_042392 | 5.80 | 54,285.57 | 483 | 62.33 | 96.15 | nucleus | 1452 |
| TcGRAS46 | TCM_042705 | 5.62 | 83,092.65 | 737 | 52.65 | 76.73 | nucleus | 2214 |
| Tandem Duplication | Chromosome Name | Ka | Ks | Ka/Ks |
|---|---|---|---|---|
| TcGRAS24 and TcGRAS25 | Chr04 | 5.21 | 2.66 | 1.96 |
| TcGRAS42 and TcGRAS43 | Chr09 | 5.62 | 1.41 | 3.99 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, S.; Zhang, Q.; Chen, J.; Meng, J.; Wang, C.; Du, J.; Guo, Y. Genome-Wide Identification and Analysis of the GRAS Transcription Factor Gene Family in Theobroma cacao. Genes 2023, 14, 57. https://doi.org/10.3390/genes14010057
Hou S, Zhang Q, Chen J, Meng J, Wang C, Du J, Guo Y. Genome-Wide Identification and Analysis of the GRAS Transcription Factor Gene Family in Theobroma cacao. Genes. 2023; 14(1):57. https://doi.org/10.3390/genes14010057
Chicago/Turabian StyleHou, Sijia, Qianqian Zhang, Jing Chen, Jianqiao Meng, Cong Wang, Junhong Du, and Yunqian Guo. 2023. "Genome-Wide Identification and Analysis of the GRAS Transcription Factor Gene Family in Theobroma cacao" Genes 14, no. 1: 57. https://doi.org/10.3390/genes14010057