
Ectopic Expression of MADS-Box Transcription Factor VvAGL12 from Grape Promotes Early Flowering, Plant Growth, and Production by Regulating Cell-Wall Architecture in Arabidopsis

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Homologous Alignment and Evolutionary Tree Analysis
2.3. Cloning and Vector Construction of Grape VvAGL12 Gene
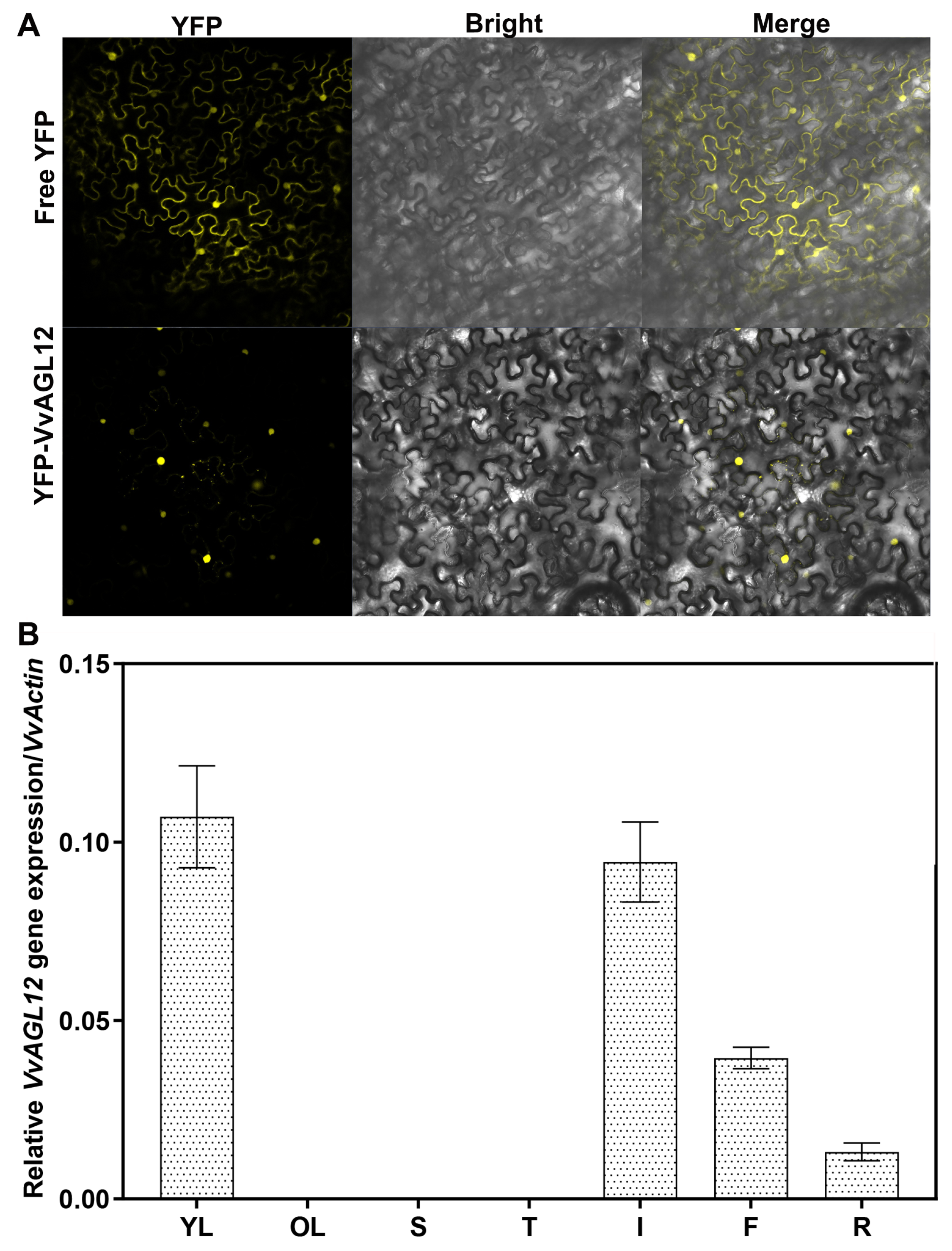
2.4. Subcellular Localization
2.5. Transformation and Phenotypic Identification of VvAGL12 Transgenic Arabidopsis
2.6. Transcriptome Sequencing
2.7. RNA Extraction and Real-Time Quantitative PCR Analysis
2.8. Statistical Analysis
3. Results
3.1. Identification and Expression Pattern Analysis of the AGL12 Gene in Grapevine
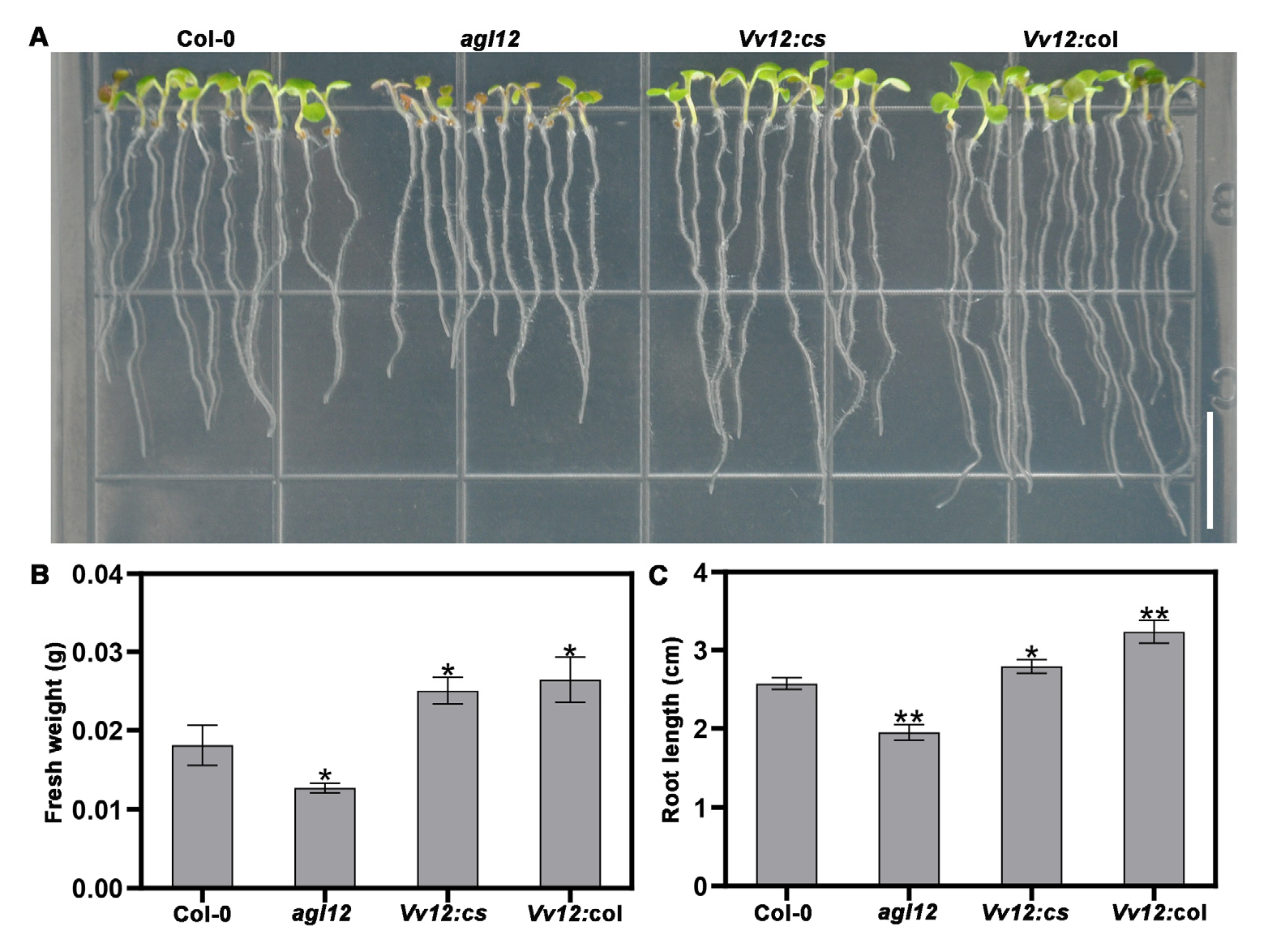
3.2. Ectopic Expression of VvAGL12 Enhanced Primary Root Development and Flowering Time in Transgenic Arabidopsis
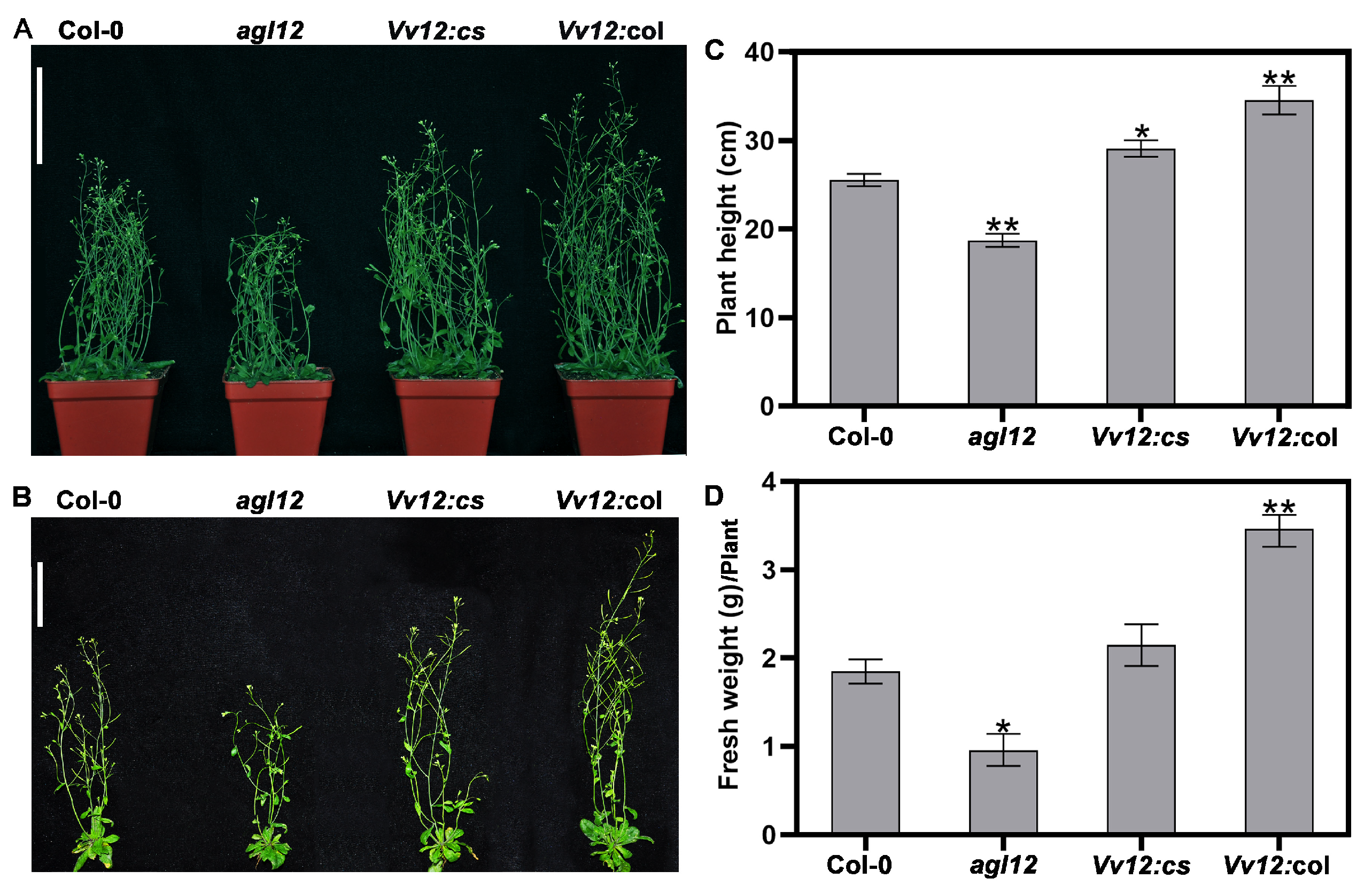
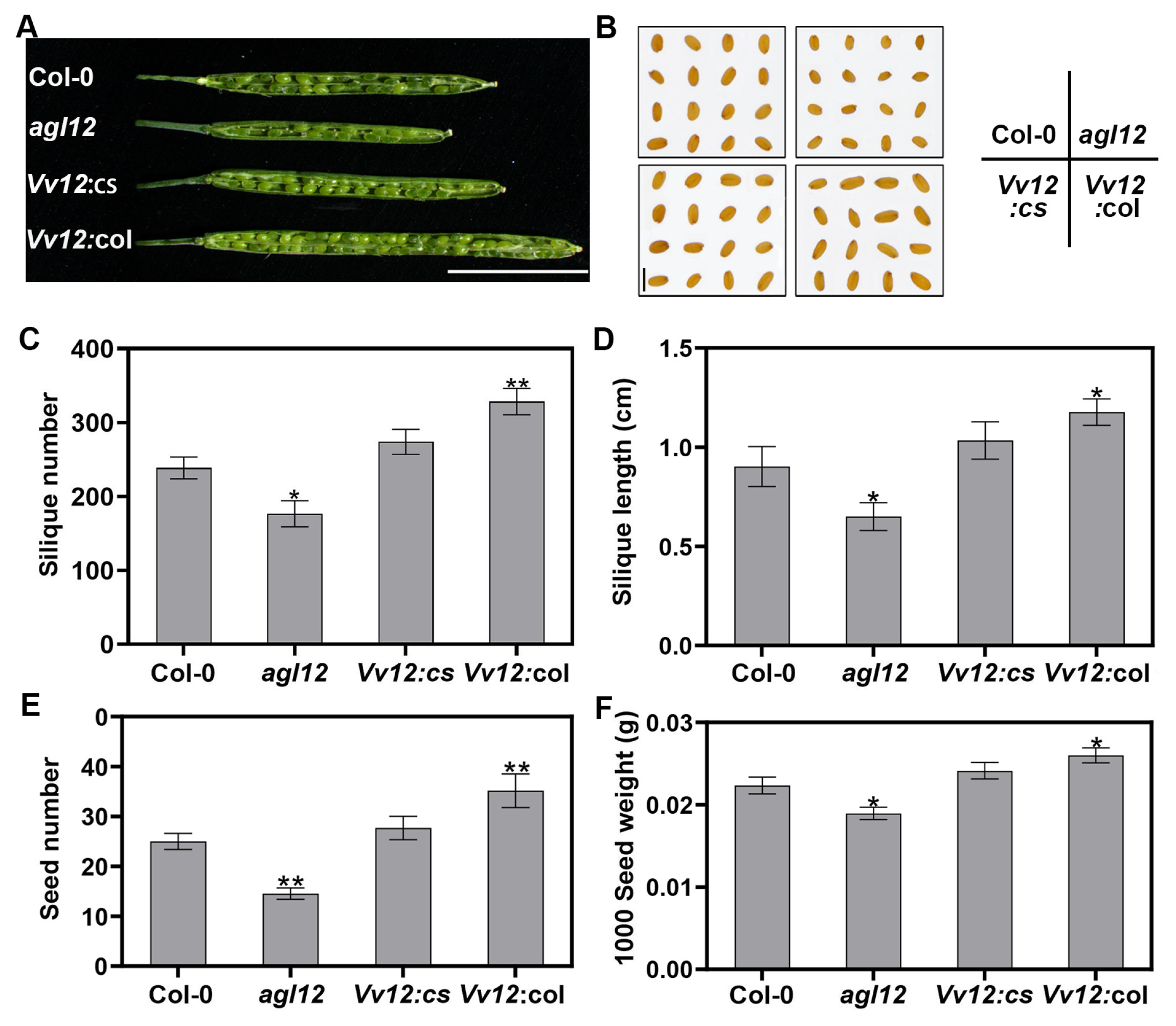
3.3. Ectopic Expression of VvAGL12 Improved the Biomass in Transgenic Arabidopsis
3.4. Ectopic Expression of VvAGL12 Enhanced the Expression Level of Cell-Wall-Related Genes in Arabidopsis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lakhwani, D.; Vikarm Dhar, Y.; Singh, S.; Pandey, A.; Kumar Trivedi, P.; Hasan Asif, M. Genome wide identification of MADS box gene family in Musa balbisiana and their divergence during evolution. Gene 2022, 836, 146666. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Ma, H. Identification and expression analysis of the MADS-box genes of Kentucky bluegrass during inflorescence development. Physiol. Mol. Biol. Plants 2022, 28, 1359–1374. [Google Scholar] [CrossRef]
- Hugouvieux, V.; Silva, C.S.; Jourdain, A.; Stigliani, A.; Charras, Q.; Conn, V.; Conn, S.J.; Carles, C.C.; Parcy, F.; Zubieta, C. Tetramerization of MADS family transcription factors SEPALLATA3 and AGAMOUS is required for floral meristem determinacy in Arabidopsis. Nucleic Acids Res. 2018, 46, 4966–4977. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Riquelme, J.; Lijavetzky, D.; Martínez-Zapater, J.M.; Carmona, M.J. Genome-wide analysis of MIKCC-Type MADS box genes in grapevine. Plant Physiol. 2009, 149, 354–369. [Google Scholar] [CrossRef] [PubMed]
- Messenguy, F.; Dubois, E. Role of MADS box proteins and their cofactors in combinatorial control of gene expression and cell development. Gene 2003, 316, 1–21. [Google Scholar] [CrossRef]
- Alvarez-Buylla, E.R.; Liljegren, S.J.; Pelaz, S.; Gold, S.E.; Burgeff, C.; Ditta, G.S.; Vergara-Silva, F.; Yanofsky, M.F. MADS-box gene evolution beyond flowers: Expression in pollen, endosperm, guard cells, roots and trichomes. Plant J. 2000, 24, 457–466. [Google Scholar]
- Burgeff, C.; Liljegren, S.; Tapia-López, R.; Yanofsky, M.F.; Alvarez-Buylla, E.R. MADS-box gene expression in lateral primordia, meristems and differentiated tissues of Arabidopsis thaliana roots. Planta 2002, 214, 365–372. [Google Scholar] [CrossRef]
- Zhang, S.; Yao, J.; Wang, L.; Wu, N.; van Nocker, S.; Li, Z.; Gao, M.; Wang, X. Role of grapevine SEPALLATA-related MADS-box gene VvMADS39 in flower and ovule development. Plant J. 2022, 111, 1565–1579. [Google Scholar] [CrossRef]
- Zhang, P.; Wang, R.; Wang, X.; Mysore, K.S.; Wen, J.; Meng, Y.; Gu, X.; Niu, L.; Lin, H. MtFULc controls inflorescence development by directly repressing MtTFL1 in Medicago truncatula. J. Plant Physiol. 2021, 256, 153329. [Google Scholar] [CrossRef]
- Yue, Y.; Sun, S.; Li, J.; Yu, H.; Wu, H.; Sun, B.; Li, T.; Han, T.; Jiang, B. GmFULa improves soybean yield by enhancing carbon assimilation without altering flowering time or maturity. Plant Cell Rep. 2021, 40, 1875–1888. [Google Scholar] [CrossRef]
- Shah, S.; Karunarathna, N.L.; Jung, C.; Emrani, N. An APETALA1 ortholog affects plant architecture and seed yield component in oilseed rape (Brassica napus L.). BMC Plant Biol. 2018, 18, 380. [Google Scholar] [CrossRef] [PubMed]
- Mou, Y.; Yuan, C.; Sun, Q.; Yan, C.; Zhao, X.; Wang, J.; Wang, Q.; Shan, S.; Li, C. MIKC-type MADS-box transcription factor gene family in peanut: Genome-wide characterization and expression analysis under abiotic stress. Front. Plant Sci. 2022, 13, 980933. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, T.; Chen, H.; Tang, H.; Mu, Y.; Gou, L.; Habib, A.; Lan, X.; Ma, J. The wheat (Triticum aestiveum L.) MADS-box transcription factor TaMADS32 plays a role in response to abiotic stresses. Biotechnol. Biotechnol. Equip. 2022, 36, 451–461. [Google Scholar] [CrossRef]
- Yuan, J.; Long, H.; Qiu, F.; Wang, Y.; Zhang, M.; Chao, Y.; Chen, L. MADS-box protein MtSOC1c regulates flowering and seed development in Medicago truncatula. Ind. Crops Prod. 2023, 193, 116125. [Google Scholar] [CrossRef]
- Montiel, G.; Gaudet, M.; Laurans, F.; Rozenberg, P.; Simon, M.; Gantet, P.; Jay-Allemand, C.; Breton, C. Overexpression of MADS-box gene AGAMOUS-LIKE 12 activates root development in Juglans sp and Arabidopsis thaliana. Plants 2020, 9, 444. [Google Scholar] [CrossRef]
- García-Cruz, K.V.; García-Ponce, B.; Garay-Arroyo, A.; Sanchez, M.P.; Ugartechea-Chirino, Y.; Desvoyes, B.; Pacheco-Escobedo, M.A.; Tapia-López, R.; Ransom-Rodríguez, I.; Gutierrez, C.; et al. The MADS-box XAANTAL1 increases proliferation at the Arabidopsis root stem-cell niche and participates in transition to differentiation by regulating cell-cycle components. Ann. Bot. 2016, 118, 787–796. [Google Scholar] [CrossRef]
- Tapia-López, R.; García-Ponce, B.; Dubrovsky, J.G.; Garay-Arroyo, A.; Pérez-Ruíz, R.V.; Kim, S.-H.; Acevedo, F.; Pelaz, S.; Alvarez-Buylla, E.R. An AGAMOUS-related MADS-box gene, XAL1 (AGL12), regulates root meristem cell proliferation and flowering transition in Arabidopsis. Plant Physiol. 2008, 146, 1182–1192. [Google Scholar] [CrossRef]
- Lee, S.; Woo, Y.-M.; Ryu, S.-I.; Shin, Y.-D.; Kim, W.T.; Park, K.Y.; Lee, I.-J.; An, G. Further characterization of a rice AGL12 group MADS-box gene, OsMADS26. Plant Physiol. 2008, 147, 156–168. [Google Scholar] [CrossRef]
- Zou, J.; Zhang, T.; Wen, G.; Song, B.; Jiang, S. First report of Penicillium olsonii causing postharvest fruit rot of grape (Vitis vinifera) in China. Plant Dis. 2022, 106, 1761. [Google Scholar] [CrossRef]
- Ji, L.; Zhou, A.; Yu, X.; Dong, Z.; Zhao, H.; Xue, H.; Wu, W. Differential expression analysis of the SRB1 gene in fluconazole-resistant and susceptible strains of Candida albicans. J. Antibiot. 2020, 73, 309–313. [Google Scholar] [CrossRef]
- Yu, S.; Wang, C.; Wang, Q.; Sun, Q.; Zhang, Y.; Dong, J.; Yin, Y.; Zhang, S.; Yu, G. Identification and analysis of SOD family genes in peanut (Arachis hypogaea L.) and their potential roles in stress responses. Agronomy 2023, 13, 1959. [Google Scholar] [CrossRef]
- Wang, Z.; Wong, D.C.J.; Wang, Y.; Xu, G.; Ren, C.; Liu, Y.; Kuang, Y.; Fan, P.; Li, S.; Xin, H.; et al. GRAS-domain transcription factor PAT1 regulates jasmonic acid biosynthesis in grape cold stress response. Plant Physiol. 2021, 186, 1660–1678. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.; Denyer, T.; Hadfield, J. Bioanalyzer chips can be used interchangeably for many analyses of DNA or RNA. Biotechniques 2016, 60, 197–199. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Chen, S.; Xie, L.; Wang, L.; Feng, Y.; Lv, T.; Fang, Y.; Ding, H. The complete chloroplast genomes of three Hamamelidaceae species: Comparative and phylogenetic analyses. Ecol. Evol. 2022, 12, e8637. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Jones, W.D.; Jensen, R.V.; Harris, S.C.; Perkins, R.G.; Goodsaid, F.M.; Guo, L.; Croner, L.J.; Boysen, C.; Fang, H.; et al. The balance of reproducibility, sensitivity, and specificity of lists of differentially expressed genes in microarray studies. BMC Bioinform. 2008, 9, S10. [Google Scholar] [CrossRef]
- Tchagang, A.B.; Gawronski, A.; Bérubé, H.; Phan, S.; Famili, F.; Pan, Y. GOAL: A software tool for assessing biological significance of genes groups. BMC Bioinform. 2010, 11, 229. [Google Scholar] [CrossRef]
- Hu, Q.; Jiang, B.; Wang, L.; Song, Y.; Tang, X.; Zhao, Y.; Fan, X.; Gu, Y.; Zheng, Q.; Cheng, J.; et al. Genome-wide analysis of growth-regulating factor genes in grape (Vitis vinifera L.): Identification, characterization and their responsive expression to osmotic stress. Plant Cell Rep. 2023, 42, 107–121. [Google Scholar] [CrossRef]
- Kudo, T.; Sasaki, Y.; Terashima, S.; Matsuda-Imai, N.; Takano, T.; Saito, M.; Kanno, M.; Ozaki, S.; Suwabe, K.; Suzuki, G.; et al. Identification of reference genes for quantitative expression analysis using large-scale RNA-seq data of Arabidopsis thaliana and model crop plants. Genes Genet. Syst. 2016, 91, 111–125. [Google Scholar] [CrossRef]
- De Bodt, S.; Theissen, G.; Van de Peer, Y. Promoter analysis of MADS-box genes in eudicots through phylogenetic footprinting. Mol. Biol. Evol. 2006, 23, 1293–1303. [Google Scholar] [CrossRef]
- Becker, A.; Theißen, G. The major clades of MADS-box genes and their role in the development and evolution of flowering plants. Mol. Phylogenet. Evol. 2003, 29, 464–489. [Google Scholar] [CrossRef]
- Alvarez-Buylla, E.R.; García-Ponce, B.; Sánchez, M.D.I.P.; Espinosa-Soto, C.; García-Gómez, M.L.; Piñeyro-Nelson, A.; Garay-Arroyo, A. MADS-box genes underground becoming mainstream: Plant root developmental mechanisms. New Phytol. 2019, 223, 1143–1158. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yin, X.; Cheng, C.; Wang, H.; Guo, R.; Xu, X.; Zhao, J.; Zheng, Y.; Wang, X. Evolutionary and expression analysis of a MADS-box gene superfamily involved in ovule development of seeded and seedless grapevines. Mol. Genet. Genom. 2015, 290, 825–846. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wang, L.; Yao, J.; Wu, N.; Ahmad, B.; Van Nocker, S.; Wu, J.; Abudureheman, R.; Li, Z.; Wang, X. Control of ovule development in Vitis vinifera by VvMADS28 and interacting genes. Hortic. Res. 2023, 10, 070. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, Z.; Wu, J.; Hong, L.; Liang, J.; Ren, Y.; Guan, P.; Hu, J. MADS-Box protein complex VvAG2, VvSEP3 and VvAGL11 regulates the formation of ovules in Vitis vinifera L. cv. ‘Xiangfei’. Genes 2021, 12, 647. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Liu, Y.; Zhang, A.; Su, S.; Yan, A.; Huang, L.; Ali, I.; Liu, Y.; Forde, B.G.; Gan, Y. MADS-box transcription factor OsMADS25 regulates root development through affection of nitrate accumulation in rice. PLoS ONE 2015, 10, e0135196. [Google Scholar] [CrossRef] [PubMed]
- Adal, A.M.; Binson, E.; Remedios, L.; Mahmoud, S.S. Expression of lavender AGAMOUS-like and SEPALLATA3-like genes promote early flowering and alter leaf morphology in Arabidopsis thaliana. Planta 2021, 254, 54. [Google Scholar] [CrossRef]
- Khong, G.N.; Pati, P.K.; Richaud, F.; Parizot, B.; Bidzinski, P.; Mai, C.D.; Bès, M.; Bourrié, I.; Meynard, D.; Beeckman, T.; et al. OsMADS26 negatively regulates resistance to pathogens and drought tolerance in rice. Plant Physiol. 2015, 169, 2935–2949. [Google Scholar] [CrossRef]
- Zhou, F.-Y.; Yu, Q.; Zhang, Y.; Yao, C.-C.; Han, Y.-J. StMADS11 subfamily gene PfMADS16 from Polypogon fugax regulates early flowering and seed development. Front. Plant Sci. 2020, 11, 525. [Google Scholar] [CrossRef]
- Paolo, D.; Rotasperti, L.; Schnittger, A.; Masiero, S.; Colombo, L.; Mizzotti, C. The Arabidopsis MADS-domain transcription factor SEEDSTICK controls seed size via direct activation of E2Fa. Plants 2021, 10, 192. [Google Scholar] [CrossRef]
- Tsuchiya, M.; Satoh, S.; Iwai, H. Distribution of XTH, expansin, and secondary-wall-related CesA in floral and fruit abscission zones during fruit development in tomato (Solanum lycopersicum). Front. Plant Sci. 2015, 6, 323. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, X.; Wang, C.; Hou, D.; Li, T.; Li, D.; Ma, C.; Sun, Z.; Tian, Y. Pear xyloglucan endotransglucosylase/hydrolases PcBRU1 promotes stem growth through regulating cell wall elongation. Plant Sci. 2021, 312, 111026. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; He, M.; Jiang, Z.; Liu, L.; Pu, J.; Zhang, W.; Wang, S.; Xu, F. The xyloglucan endotransglucosylase/hydrolase gene XTH22/TCH4 regulates plant growth by disrupting the cell wall homeostasis in Arabidopsis under boron deficiency. Int. J. Mol. Sci. 2022, 23, 1250. [Google Scholar] [CrossRef]
- Miedes, E.; Suslov, D.; Vandenbussche, F.; Kenobi, K.; Ivakov, A.; Van Der Straeten, D.; Lorences, E.P.; Mellerowicz, E.J.; Verbelen, J.-P.; Vissenberg, K. Xyloglucan endotransglucosylase/hydrolase (XTH) overexpression affects growth and cell wall mechanics in etiolated Arabidopsis hypocotyls. J. Exp. Bot. 2013, 64, 2481–2497. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Duan, H.; Chen, F.; Wang, Z.; Huang, X.; Deng, X.; Liu, Y. Identification of quantitative trait Loci controlling high calcium response in Arabidopsis thaliana. PLoS ONE 2014, 9, e112511. [Google Scholar] [CrossRef] [PubMed]
- Gilliham, M.; Dayod, M.; Hocking, B.J.; Xu, B.; Conn, S.J.; Kaiser, B.N.; Leigh, R.A.; Tyerman, S.D. Calcium delivery and storage in plant leaves: Exploring the link with water flow. J. Exp. Bot. 2011, 62, 2233–2250. [Google Scholar] [CrossRef]
- Dayod, M.; Tyerman, S.D.; Leigh, R.A.; Gilliham, M. Calcium storage in plants and the implications for calcium biofortification. Protoplasma 2010, 247, 215–231. [Google Scholar] [CrossRef]
- Chan, C.W.M.; Schorrak, L.M.; Smith, R.K., Jr.; Bent, A.F.; Sussman, M.R. A cyclic nucleotide-gated Ion channel, CNGC2, is crucial for plant development and adaptation to calcium stress. Plant Physiol. 2003, 132, 728–731. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Liu, H.; Yang, H.; Lai, S.; Cheng, X.; Xin, Y.; Yang, B.; Hou, H.; Yao, Y.; Zhang, S.; et al. Quality attributes and cell wall properties of strawberries (Fragaria annanassa Duch.) under calcium chloride treatment. Food Chem. 2011, 126, 450–459. [Google Scholar] [CrossRef]
- Imaizumi, C.; Tomatsu, H.; Kitazawa, K.; Yoshimi, Y.; Shibano, S.; Kikuchi, K.; Yamaguchi, M.; Kaneko, S.; Tsumuraya, Y.; Kotake, T. Heterologous expression and characterization of an Arabidopsis β-l-arabinopyranosidase and α-d-galactosidases acting on β-l-arabinopyranosyl residues. J. Exp. Bot. 2017, 68, 4651–4661. [Google Scholar] [CrossRef]
- Hoffmann, N.; Benske, A.; Betz, H.; Schuetz, M.; Samuels, A.L. Laccases and peroxidases Co-localize in lignified secondary cell walls throughout stem development. Plant Physiol. 2020, 184, 806–822. [Google Scholar] [CrossRef]
- Passardi, F.; Tognolli, M.; De Meyer, M.; Penel, C.; Dunand, C. Two cell wall associated peroxidases from Arabidopsis influence root elongation. Planta 2006, 223, 965–974. [Google Scholar] [CrossRef]
- Kurdyukov, S.; Faust, A.; Nawrath, C.; Bär, S.; Voisin, D.; Efremova, N.; Franke, R.; Schreiber, L.; Saedler, H.; Métraux, J.-P.; et al. The Epidermis-specific extracellular BODYGUARD controls cuticle development and morphogenesis in Arabidopsis. Plant Cell 2006, 18, 321–339. [Google Scholar] [CrossRef] [PubMed]
- An, J.-P.; Wang, X.-F.; Espley, R.V.; Lin-Wang, K.; Bi, S.-Q.; You, C.-X.; Hao, Y.-J. An apple B-Box protein MdBBX37 modulates anthocyanin biosynthesis and hypocotyl elongation synergistically with MdMYBs and MdHY5. Plant Cell Physiol. 2019, 61, 130–143. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Bian, Y.; Liu, J.; Sun, Y.; Xu, D. B-box proteins: Pivotal players in light-mediated development in plants. J. Integr. Plant Biol. 2020, 62, 1293–1309. [Google Scholar] [CrossRef]
- Andrés, F.; Coupland, G. The genetic basis of flowering responses to seasonal cues. Nat. Rev. Genet. 2012, 13, 627–639. [Google Scholar] [CrossRef]
- Kinoshita, A.; Richter, R. Genetic and molecular basis of floral induction in Arabidopsis thaliana. J. Exp. Bot. 2020, 71, 2490–2504. [Google Scholar] [CrossRef] [PubMed]
- Susila, H.; Nasim, Z.; Gawarecka, K.; Jung, J.-Y.; Jin, S.; Youn, G.; Ahn, J.H. Chloroplasts prevent precocious flowering through a GOLDEN2-LIKE–B-BOX DOMAIN PROTEIN module. Plant Commun. 2023, 4, 100515. [Google Scholar] [CrossRef]
- Xu, M.; Hu, T.; Zhao, J.; Park, M.-Y.; Earley, K.W.; Wu, G.; Yang, L.; Poethig, R.S. Developmental functions of miR156-Regulated SQUAMOSA PROMOTER BINDING PROTEIN-LIKE (SPL) genes in Arabidopsis thaliana. PLoS Genet. 2016, 12, e1006263. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mao, T.; Wang, X.; Gao, H.; Gong, Z.; Liu, R.; Jiang, N.; Zhang, Y.; Zhang, H.; Guo, X.; Yu, C. Ectopic Expression of MADS-Box Transcription Factor VvAGL12 from Grape Promotes Early Flowering, Plant Growth, and Production by Regulating Cell-Wall Architecture in Arabidopsis. Genes 2023, 14, 2078. https://doi.org/10.3390/genes14112078
Mao T, Wang X, Gao H, Gong Z, Liu R, Jiang N, Zhang Y, Zhang H, Guo X, Yu C. Ectopic Expression of MADS-Box Transcription Factor VvAGL12 from Grape Promotes Early Flowering, Plant Growth, and Production by Regulating Cell-Wall Architecture in Arabidopsis. Genes. 2023; 14(11):2078. https://doi.org/10.3390/genes14112078
Chicago/Turabian StyleMao, Tingting, Xueting Wang, Hongsheng Gao, Zijian Gong, Ruichao Liu, Ning Jiang, Yaru Zhang, Hongxia Zhang, Xiaotong Guo, and Chunyan Yu. 2023. "Ectopic Expression of MADS-Box Transcription Factor VvAGL12 from Grape Promotes Early Flowering, Plant Growth, and Production by Regulating Cell-Wall Architecture in Arabidopsis" Genes 14, no. 11: 2078. https://doi.org/10.3390/genes14112078