
Aliens on Boats? The Eastern and Western Expansion of the African House Gecko



Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
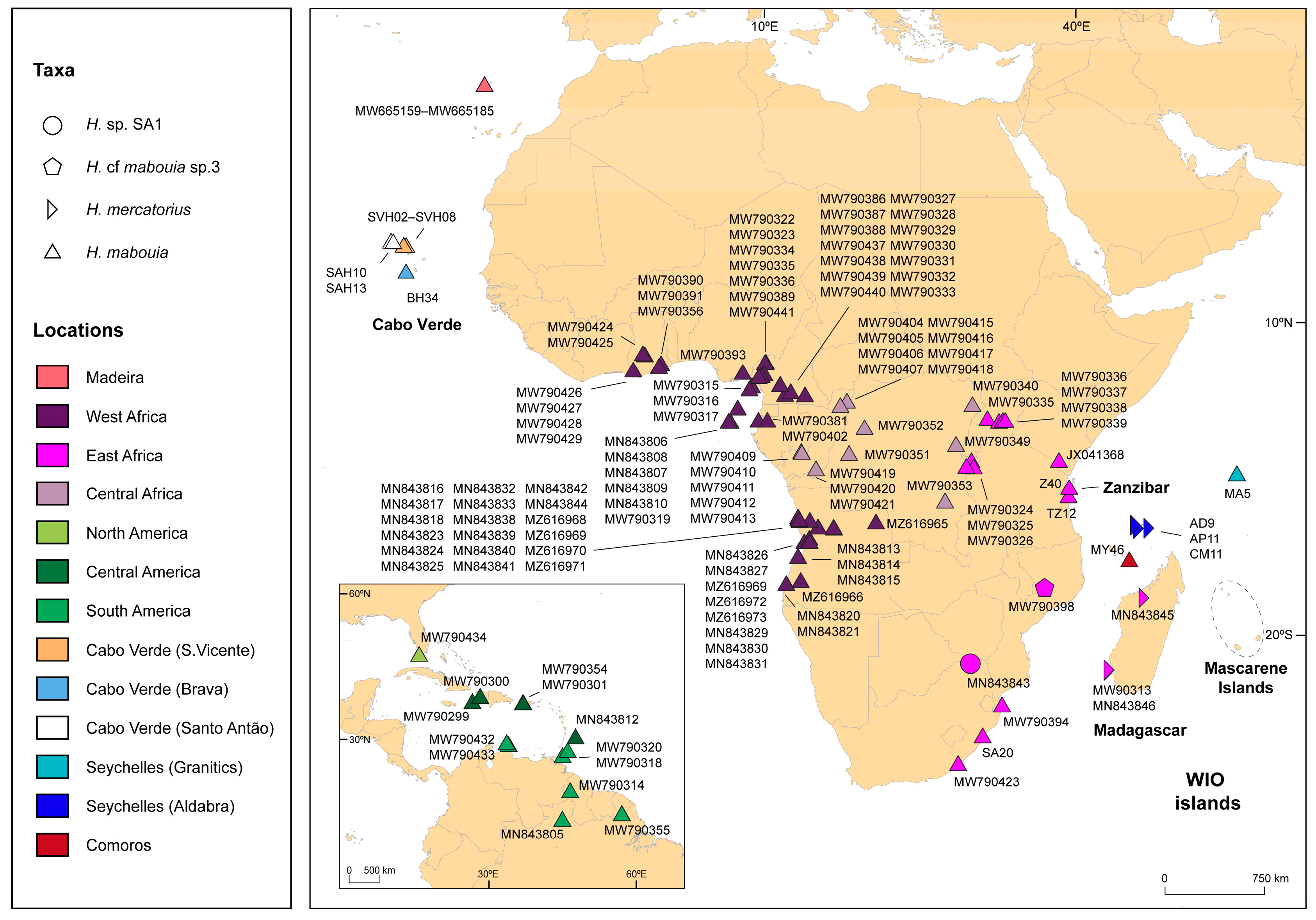
2.1. Data Collection
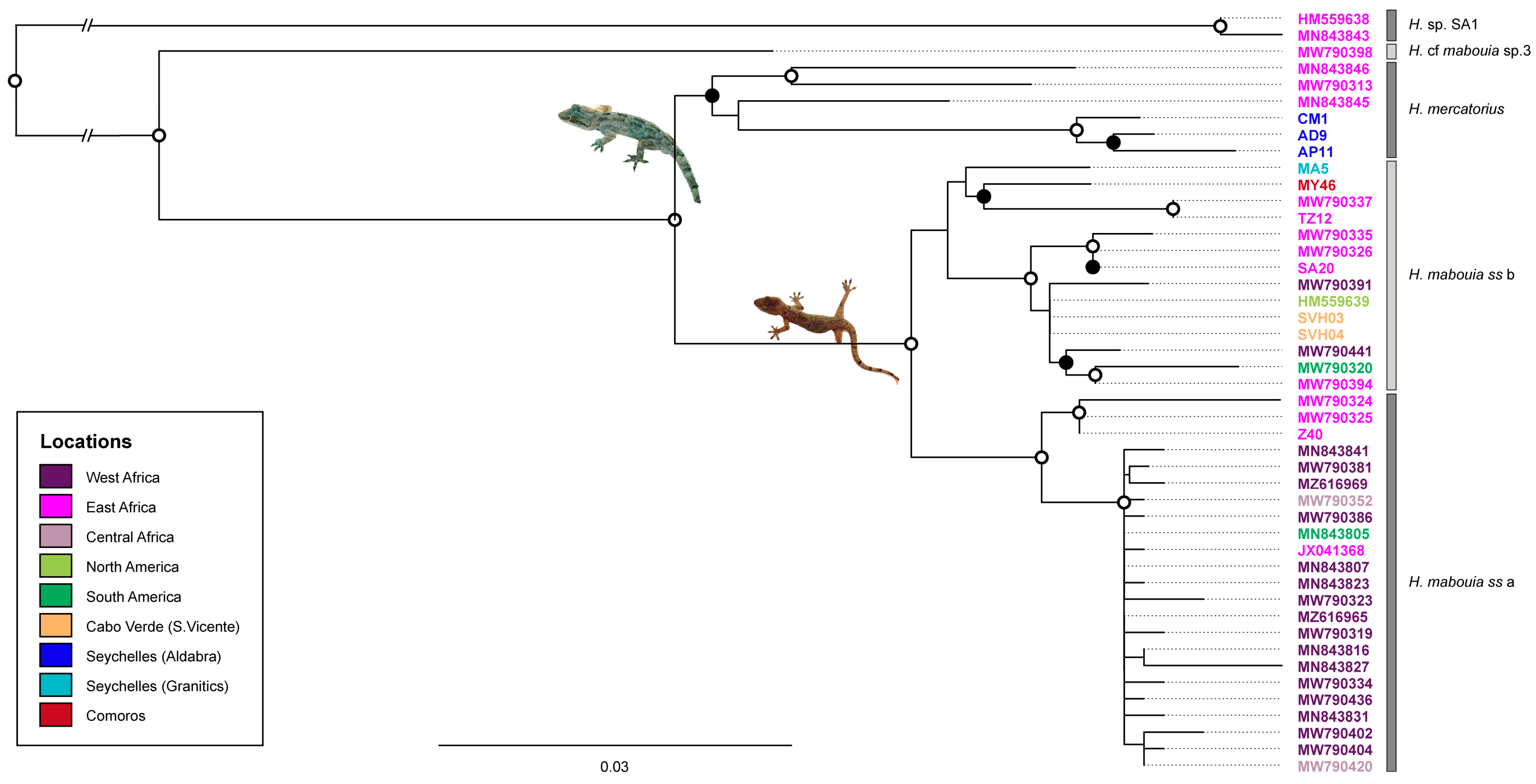
2.2. Alignment and Phylogenetic Inference
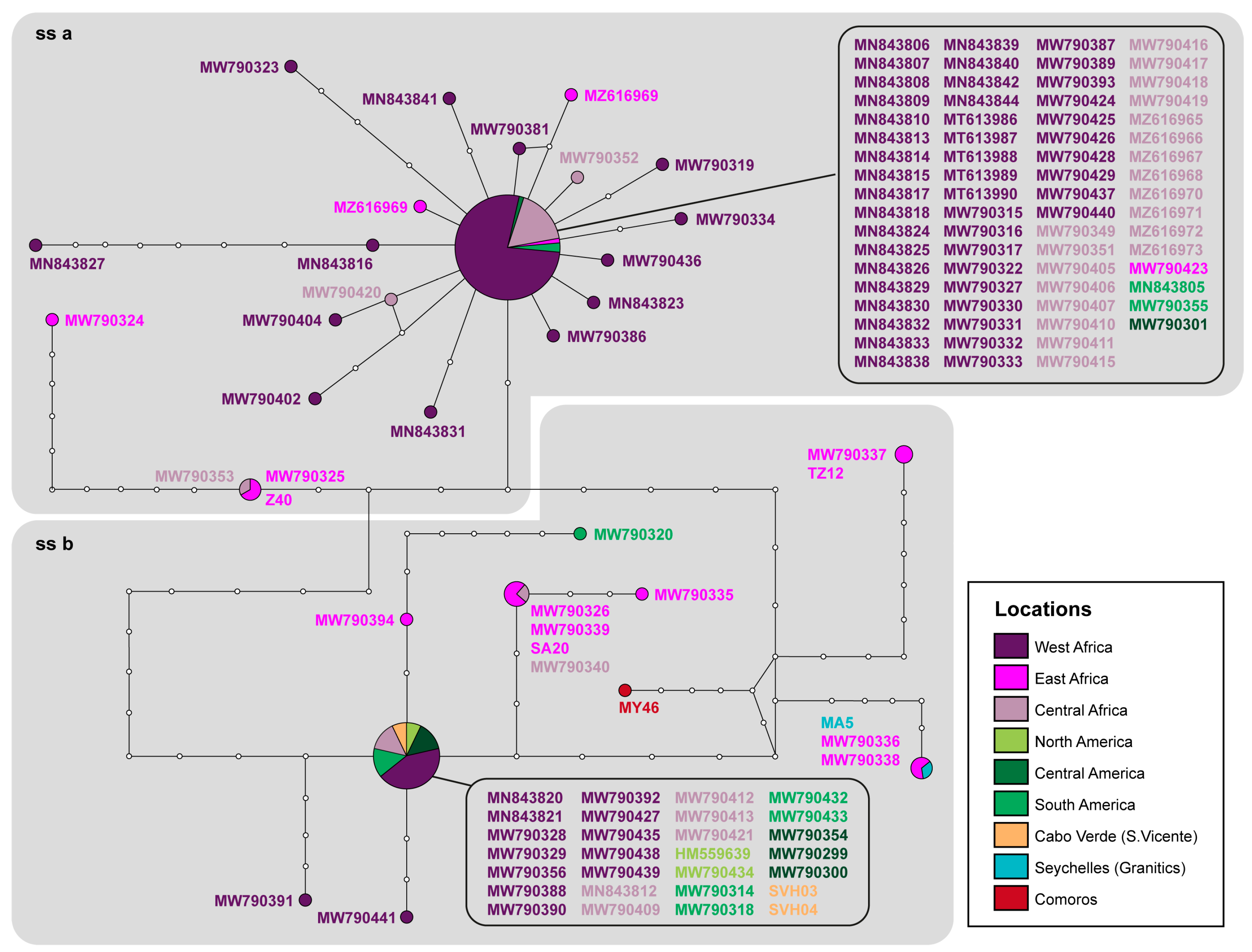
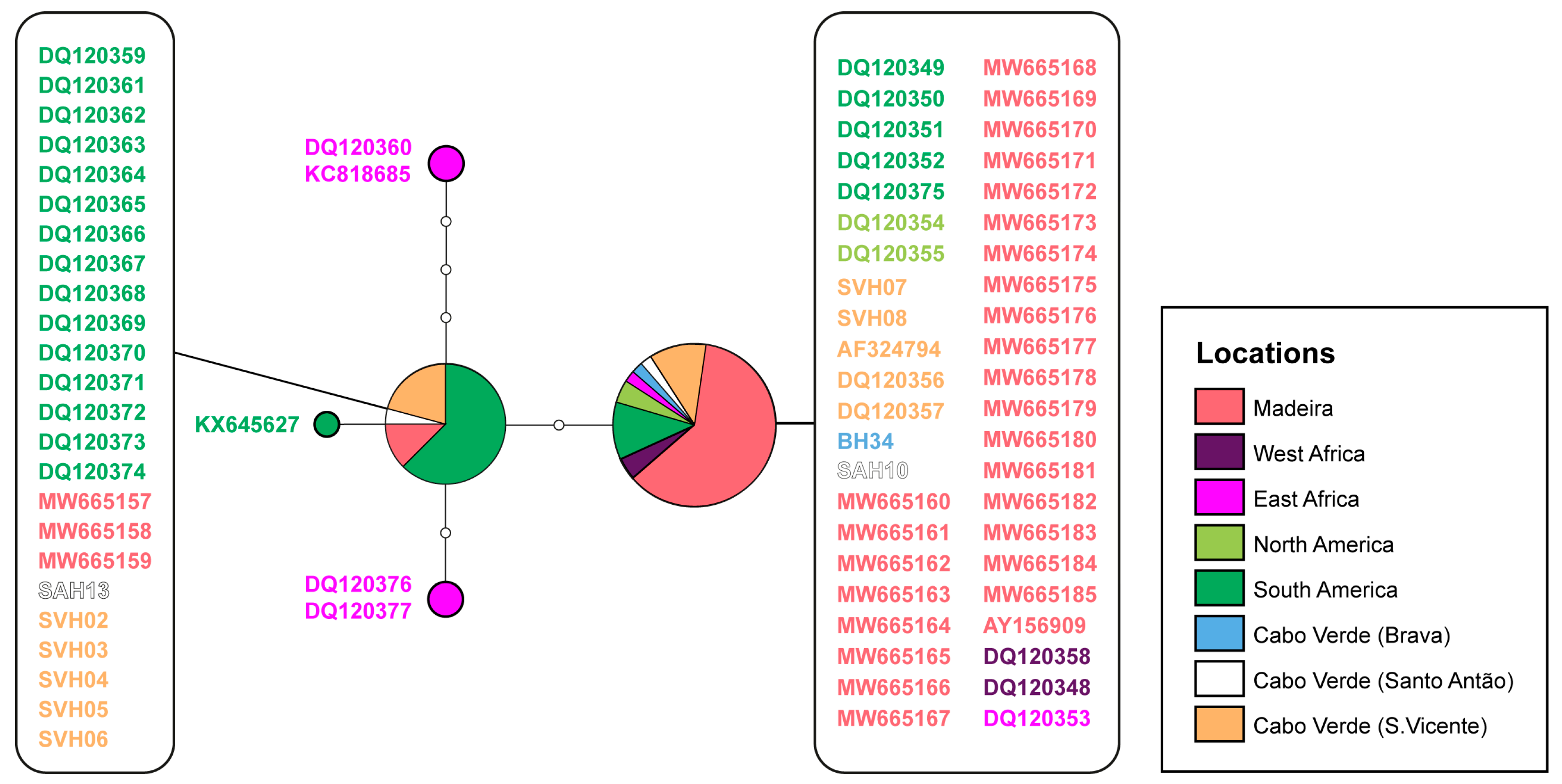
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Paini, D.R.; Sheppard, A.W.; Cook, D.C.; De Barro, P.J.; Worner, S.P.; Thomas, M.B. Global threat to agriculture from invasive species. Proc. Natl. Acad. Sci. USA 2016, 113, 7575–7579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz, G.M.; Carlton, J.T. Invasive species: Vectors and management strategies. Divers. Distrib. 2003, 10, 508. [Google Scholar] [CrossRef]
- Gaiotto, J.V.; Abrahão, C.R.; Dias, R.A.; Bugoni, L. Diet of invasive cats, rats and tegu lizards reveals impact over threatened species in a tropical island. Perspect. Ecol. Conserv. 2020, 18, 294–303. [Google Scholar] [CrossRef]
- Willson, J.D. Indirect effects of invasive Burmese pythons on ecosystems in southern Florida. J. Appl. Ecol. 2017, 54, 1251–1258. [Google Scholar] [CrossRef] [Green Version]
- Alpert, P. The advantages and disadvantages of being introduced. Biol. Invasions 2006, 8, 1523–1534. [Google Scholar] [CrossRef]
- Reed, R.; Kraus, F. Invasive reptiles and amphibians: Global perspectives and local solutions. Anim. Conserv. 2010, 13, 3–4. [Google Scholar] [CrossRef]
- Novosolov, M.; Raia, P.; Meiri, S. The island syndrome in lizards. Glob. Ecol. Biogeogr. 2013, 22, 184–191. [Google Scholar] [CrossRef]
- Cooper, W.E., Jr.; Pyron, R.A.; Garland, T., Jr. Island tameness: Living on islands reduces flight initiation distance. Proc. R. Soc. B Boil. Sci. 2014, 281, 20133019. [Google Scholar] [CrossRef] [Green Version]
- Soares, N.; Gonçalves, J.F.; Vasconcelos, R.; Ribeiro, R.P. Combining multiple data sources to predict IUCN conservation status of reptiles. In Advances in Intelligent Data Analysis XX, Proceedings of the 20th International Symposium on Intelligent Data Analysis, IDA 2022, Rennes, France, 20–22 April 2022; Springer: Cham, Switzerland, 2022; pp. 302–314. [Google Scholar]
- Agarwal, I.; Ceríaco, L.M.; Metallinou, M.; Jackman, T.R.; Bauer, A.M. How the African house gecko (Hemidactylus mabouia) conquered the world. R. Soc. Open Sci. 2021, 8, 210749. [Google Scholar] [CrossRef]
- Bansal, R.; Karanth, K.P. Phylogenetic analysis and molecular dating suggest that Hemidactylus anamallensis is not a member of the Hemidactylus radiation and has an ancient late Cretaceous origin. PLoS ONE 2013, 8, e60615. [Google Scholar] [CrossRef] [Green Version]
- Cole, N.C.; Jones, C.G.; Harris, S. The need for enemy-free space: The impact of an invasive gecko on island endemics. Biol. Conserv. 2005, 125, 467–474. [Google Scholar] [CrossRef]
- Weterings, R.; Vetter, K.C. Invasive house geckos (Hemidactylus spp.): Their current, potential and future distribution. Curr. Zool. 2018, 64, 559–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, D.F.; Meshaka, W.E., Jr.; van Buurt, G. The superior colonizing gecko Hemidactylus mabouia on Curaçao: Conservation implications for the native gecko Phyllodactylus martini. J. Herpetol 2015, 49, 60–63. [Google Scholar] [CrossRef]
- Buurt, G.V. Field Guide to the Reptiles and Amphibians of Aruba, Curaçao and Bonaire; Edition Chimaira: Frankfurt am Main, Germany, 2004. [Google Scholar]
- Williams, R.; Pernetta, A.P.; Horrocks, J.A. Outcompeted by an invader? Interference and exploitative competition between tropical house gecko (Hemidactylus mabouia) and Barbados leaf-toed gecko (Phyllodactylus pulcher) for diurnal refuges in anthropogenic coastal habitats. Integr. Zool. 2016, 11, 229–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Short, K.H.; Petren, K. Rapid species displacement during the invasion of Florida by the tropical house gecko Hemidactylus mabouia. Biol. Invasions 2012, 14, 1177–1186. [Google Scholar] [CrossRef]
- Dornburg, A.; Lippi, C.; Federman, S.; Moore, J.A.; Warren, D.L.; Iglesias, T.L.; Brandley, M.C.; Watkins-Colwell, G.J.; Lamb, A.D.; Jones, A. Disentangling the influence of urbanization and invasion on endemic geckos in tropical biodiversity hot spots: A case study of Phyllodactylus martini (Squamata: Phyllodactylidae) along an urban gradient in Curaçao. Bull. Peabody Mus. Nat. Hist. 2016, 57, 147–164. [Google Scholar] [CrossRef] [Green Version]
- Zaaf, A.; Van Damme, R. Limb proportions in climbing and ground-dwelling geckos (Lepidosauria, Gekkonidae): A phylogenetically informed analysis. Zoomorphology 2001, 121, 45–53. [Google Scholar] [CrossRef]
- Rocha, S.; Carretero, M.A.; Harris, D.J. On the diversity, colonization patterns and status of Hemidactylus spp. (Reptilia: Gekkonidae) from the Western Indian Ocean islands. Herpetol. J. 2010, 20, 83–89. [Google Scholar]
- Vasconcelos, R.; Brito, J.C.; Carranza, S.; Harris, D.J. Review of the distribution and conservation status of the terrestrial reptiles of the Cape Verde Islands. Oryx 2013, 47, 77–87. [Google Scholar] [CrossRef] [Green Version]
- Simberloff, D. Invasive Species: What Everyone Needs to Know; Oxford University Press: Oxford, UK, 2013. [Google Scholar]
- Rato, C.; Martins, B.; Rocha, R.; Silva-Rocha, I. Uncovered genetic diversity in Hemidactylus mabouia (Reptilia: Gekkonidae) from Madeira Island reveals uncertain sources of introduction. Amphibia-Reptilia 2021, 42, 369–375. [Google Scholar] [CrossRef]
- Jesus, J.; Brehm, A.; Pinheiro, M.; Harris, D.J. Relationships of Hemidactylus (Reptilia: Gekkonidae) from the Cape Verde Islands: What mitochondrial DNA data indicate. J. Herpetol. 2001, 35, 672–675. [Google Scholar] [CrossRef]
- Koehler, G.; Vasconcelos, R.; Köhler, G.; Geniez, P.; Crochet, P.-A. A new endemic species of Hemidactylus (Squamata: Gekkonidae) from São Nicolau Island, Cabo Verde. Zootaxa 2020, 4878, 501–522. [Google Scholar] [CrossRef]
- Vences, M.; Wanke, S.; Vieites, D.R.; Branch, W.R.; Glaw, F.; Meyer, A. Natural colonization or introduction? Phylogeographical relationships and morphological differentiation of house geckos (Hemidactylus) from Madagascar. Biol. J. Linn. Soc. 2004, 83, 115–130. [Google Scholar] [CrossRef]
- Boumans, L.; Vieites, D.R.; Glaw, F.; Vences, M. Geographical patterns of deep mitochondrial differentiation in widespread Malagasy reptiles. Mol. Phylogenetics Evol. 2007, 45, 822–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha, S.; Carretero, M.A.; Harris, D.J. Diversity and phylogenetic relationships of Hemidactylus geckos from the Comoro islands. Mol. Phylogenetics Evol. 2005, 35, 292–299. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989. [Google Scholar]
- Arnold, E.; Vasconcelos, R.; Harris, D.; Mateo, J.; Carranza, S. Systematics, biogeography and evolution of the endemic Hemidactylus geckos (Reptilia, Squamata, Gekkonidae) of the Cape Verde Islands: Based on morphology and mitochondrial and nuclear DNA sequences. Zool. Scr. 2008, 37, 619–636. [Google Scholar] [CrossRef]
- Macey, J.R.; Larson, A.; Ananjeva, N.B.; Fang, Z.; Papenfuss, T.J. Two novel gene orders and the role of light-strand replication in rearrangement of the vertebrate mitochondrial genome. Mol. Biol. Evol. 1997, 14, 91–104. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organisation and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Glez-Peña, D.; Gomez-Blanco, D.; Reboiro-Jato, M.; Fdez-Riverola, F.; Posada, D. ALTER: Program-oriented conversion of DNA and protein alignments. Nucleic Acids Res. 2010, 38, W14–W18. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Crandall, M.C.D.P.K.; Clement, M.; Posada, D. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1660. [Google Scholar] [CrossRef] [Green Version]
- Murias dos Santos, A.; Cabezas, M.P.; Tavares, A.I.; Xavier, R.; Branco, M. tcsBU: A tool to extend TCS network layout and visualisation. Bioinformatics 2016, 32, 627–628. [Google Scholar] [CrossRef] [Green Version]
- Du Tertre, J.-B. Histoire Générale des Isles de S. Christophe, de la Guadeloupe, de la Martinique et Autres dans l’Amérique; Chez Jacques Langlois et Emmanuel Langlois: Paris, France, 1654. [Google Scholar]
- Sanchez, M.; Rocha, S.; Probst, J.-M. Un nouveau gecko nocturne naturalisé sur l’île de La Réunion: Hemidactylus mercatorius Gray, 1842 (Reptilia: Squamata: Gekkonidae). Bull. Soc. Herp. Fr. 2012, 142, 89–106. [Google Scholar]
- Santos, A.S.C.d. História da Colonização do Ratinho-Caseiro, Mus musculus domesticus, em Ilhas Atlânticas (Madeira, Açores e Cabo Verde): Uma Abordagem Multilocus. Ph.D. Thesis, Universidade de Lisboa, Lisboa, Portugal, 2017. [Google Scholar]
- Jesus, J.; Brehm, A.; Harris, D.J. Phylogenetic relationships of Hemidactylus geckos from the Gulf of Guinea islands: Patterns of natural colonisations and anthropogenic introductions estimated from mitochondrial and nuclear DNA sequences. Mol. Phylogenetics Evol. 2005, 34, 480–485. [Google Scholar] [CrossRef]
- López-Jurado, L.F.; Mateo, J.A.; Geniez, P. Los reptiles de la Isla de Boavista (archipiélago de Cabo Verde). Bol. Asoc. Herpetol. Esp. 1999, 10, 10–13. [Google Scholar]
- Cole, N. The new noisy neighbours: Impacts of alien house geckos on endemics in Mauritius. Aliens-Invasive Species Spec. Group Newsl. 2005, 22, 8–10. [Google Scholar]
- Fuenmayor, G.R.; Ugueto, G.; Bauer, A.M.; Barros, T.; Manzanilla, J. Expansion and natural history of a successful colonising gecko in Venezuela (Reptilia: Gekkonidae: Hemidactylus mabouia) and the discovery of H. frenatus in Venezuela. Herpetol. Rev. 2005, 36, 121. [Google Scholar]
- Rödder, D.; Solé, M.; Böhme, W. Predicting the potential distribution of two alien invasive Housegeckos (Gekkonidae: Hemidactylus frenatus, Hemidactylus mabouia). North-West. J. Zool. 2008, 4, 236–246. [Google Scholar]
- van Buur, G. Conservation of amphibians and reptiles in Aruba, Curaçao and Bonaire. Appl. Herpetol. 2006, 3, 307–321. [Google Scholar] [CrossRef] [Green Version]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinho, C.J.; Cardoso, L.; Rocha, S.; Vasconcelos, R. Aliens on Boats? The Eastern and Western Expansion of the African House Gecko. Genes 2023, 14, 381. https://doi.org/10.3390/genes14020381
Pinho CJ, Cardoso L, Rocha S, Vasconcelos R. Aliens on Boats? The Eastern and Western Expansion of the African House Gecko. Genes. 2023; 14(2):381. https://doi.org/10.3390/genes14020381
Chicago/Turabian StylePinho, Catarina Jesus, Luís Cardoso, Sara Rocha, and Raquel Vasconcelos. 2023. "Aliens on Boats? The Eastern and Western Expansion of the African House Gecko" Genes 14, no. 2: 381. https://doi.org/10.3390/genes14020381
APA StylePinho, C. J., Cardoso, L., Rocha, S., & Vasconcelos, R. (2023). Aliens on Boats? The Eastern and Western Expansion of the African House Gecko. Genes, 14(2), 381. https://doi.org/10.3390/genes14020381