
Intraspecific Chloroplast Genome Variation and Domestication Origins of Major Cultivars of Styphnolobium japonicum

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Plant Total DNA Extraction, Genome Sequencing, and Assembly
2.3. Chloroplast Genome Comparative Analysis
2.4. Phylogenetic Analysis
3. Results
3.1. Characteristics of the Chloroplast Genome of S. japonicum
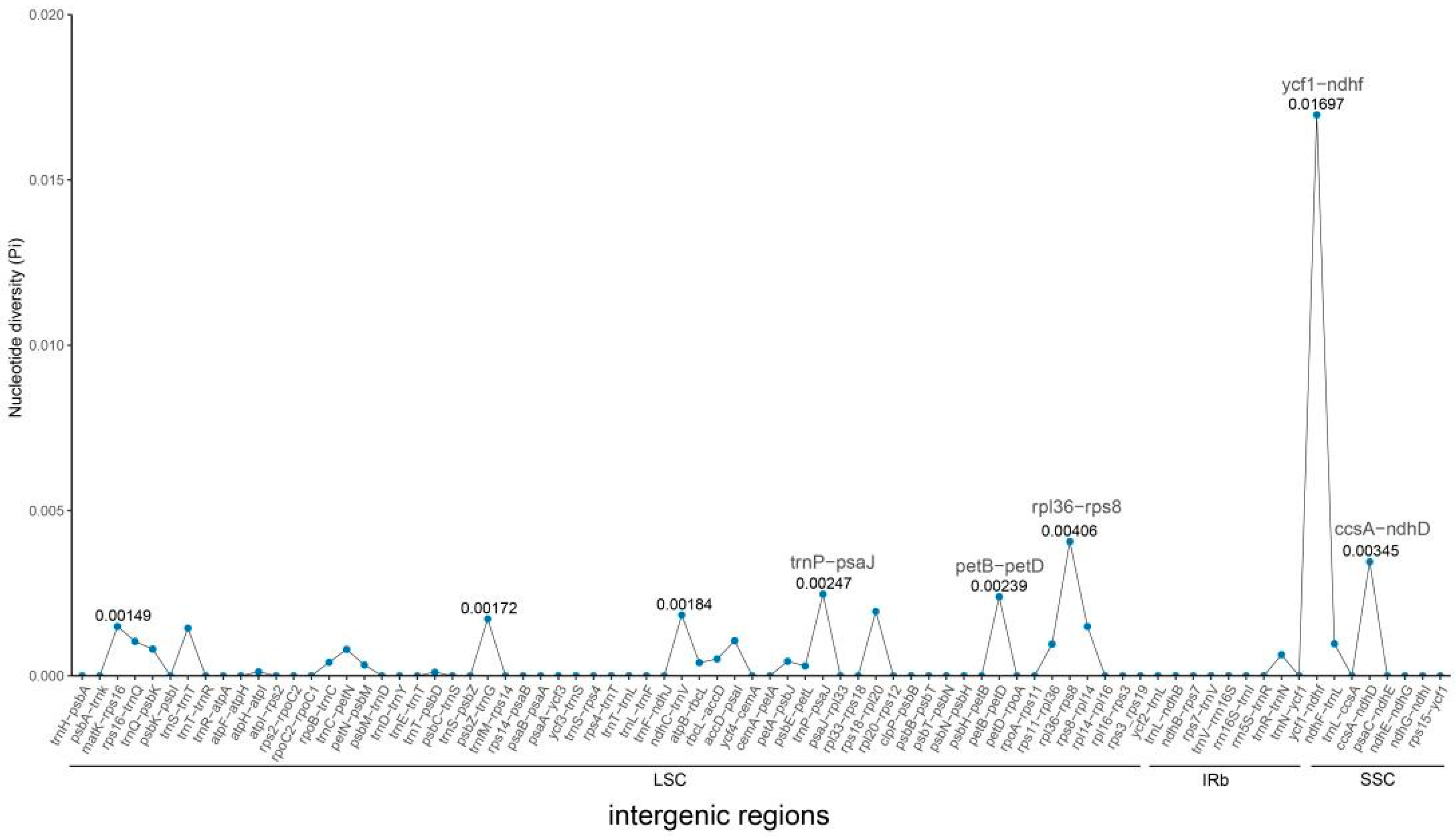
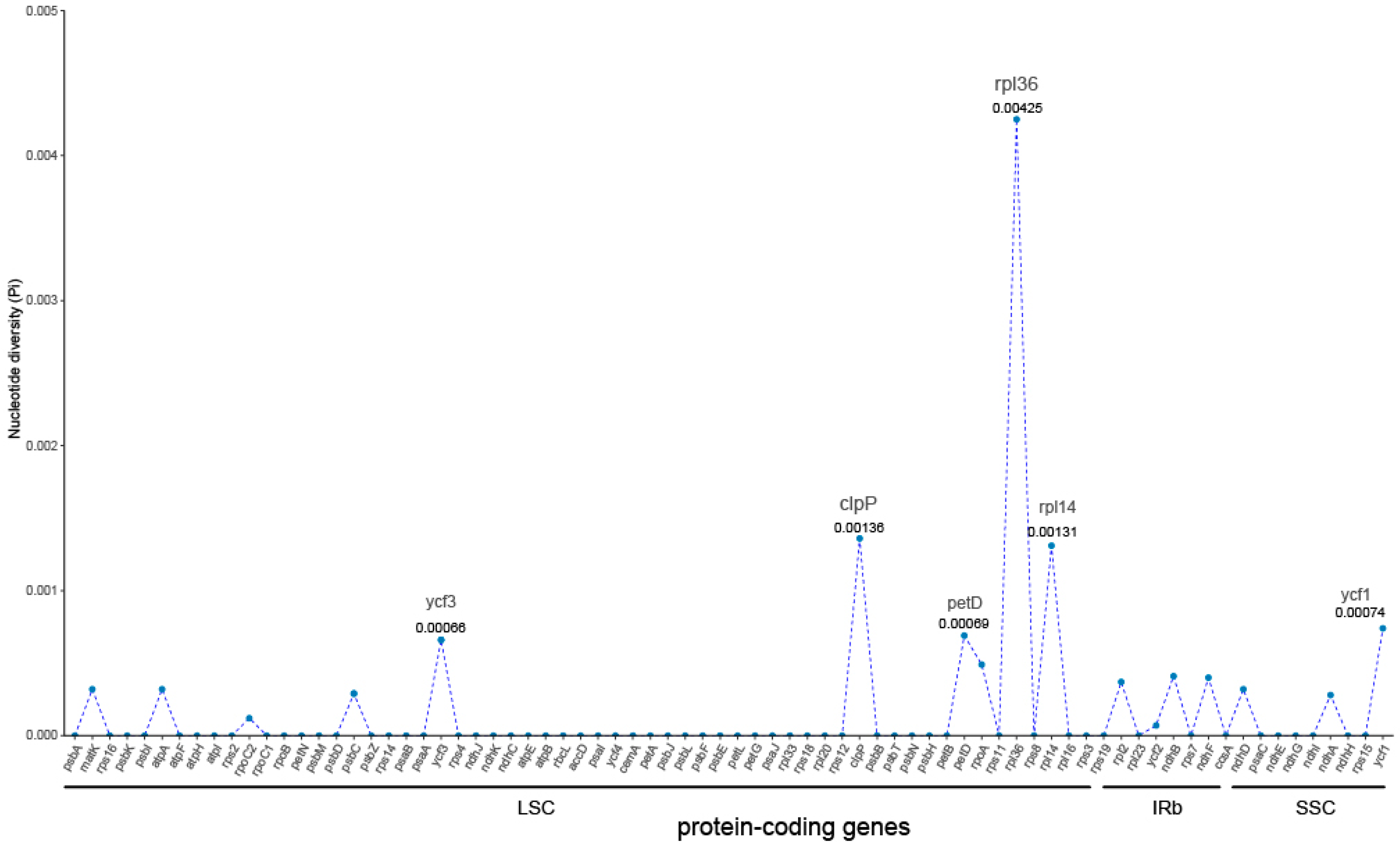
3.2. Chloroplast Genome Comparative Analysis
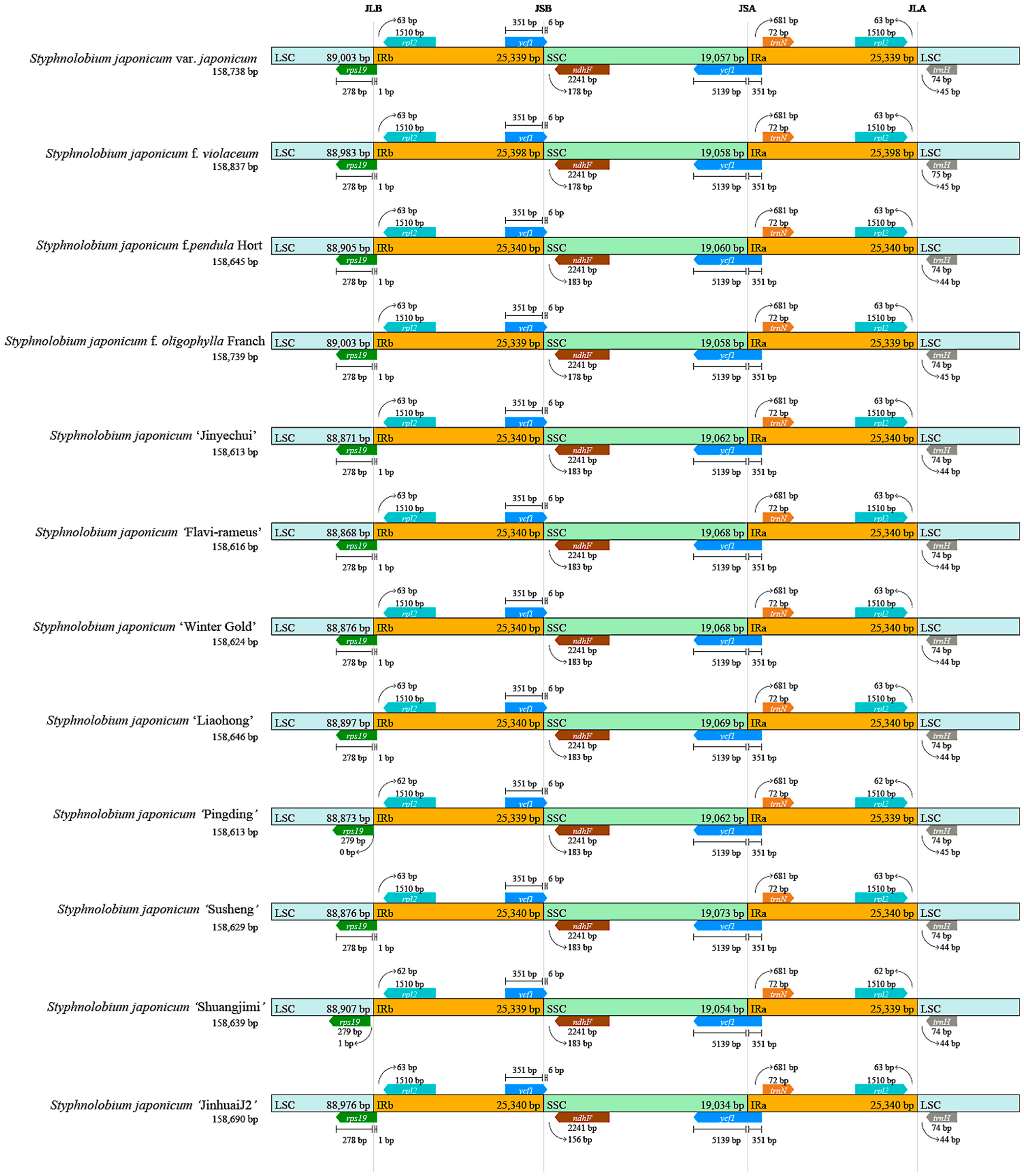
3.3. Chloroplast Genome Collinearity and Partition Boundary Analysis
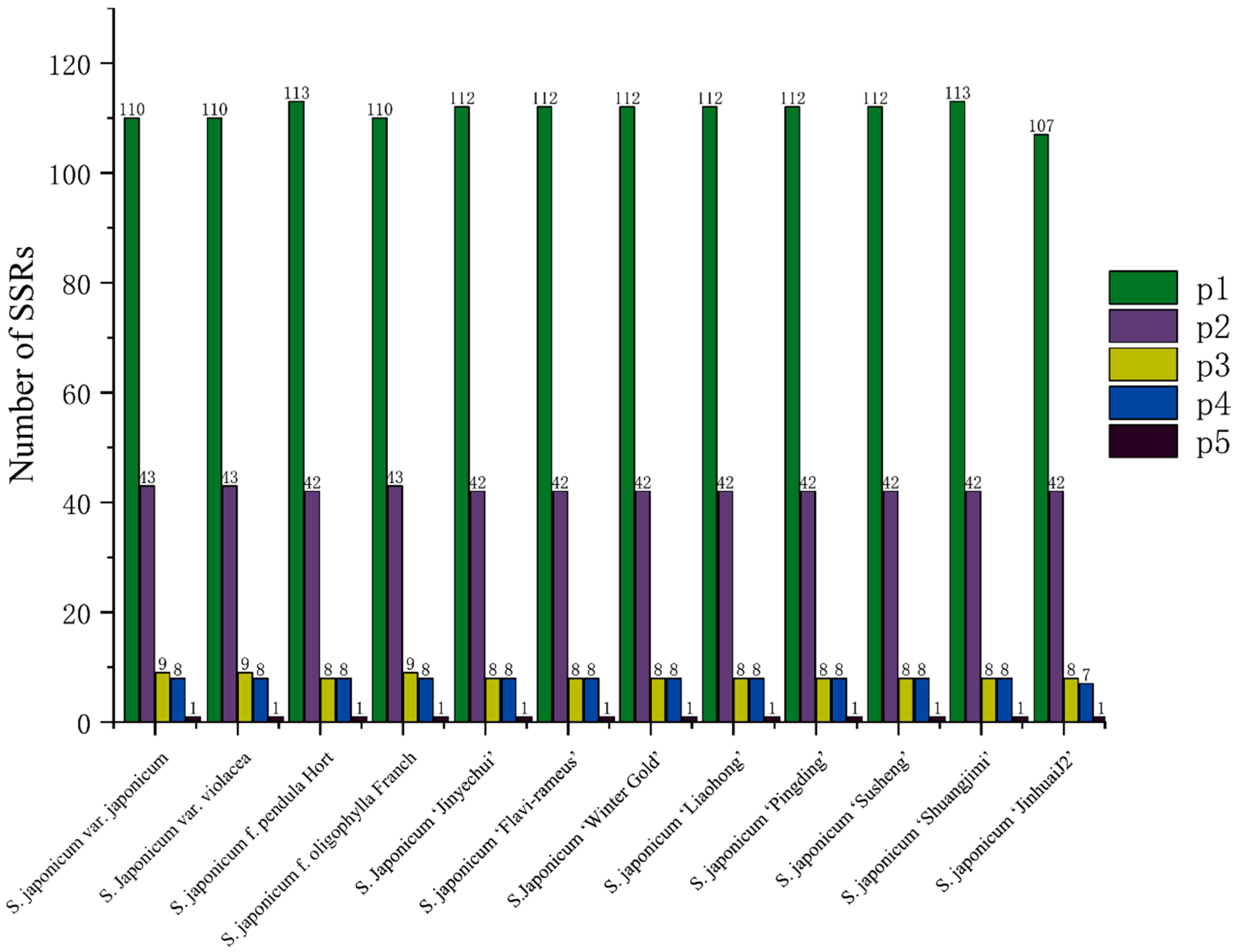
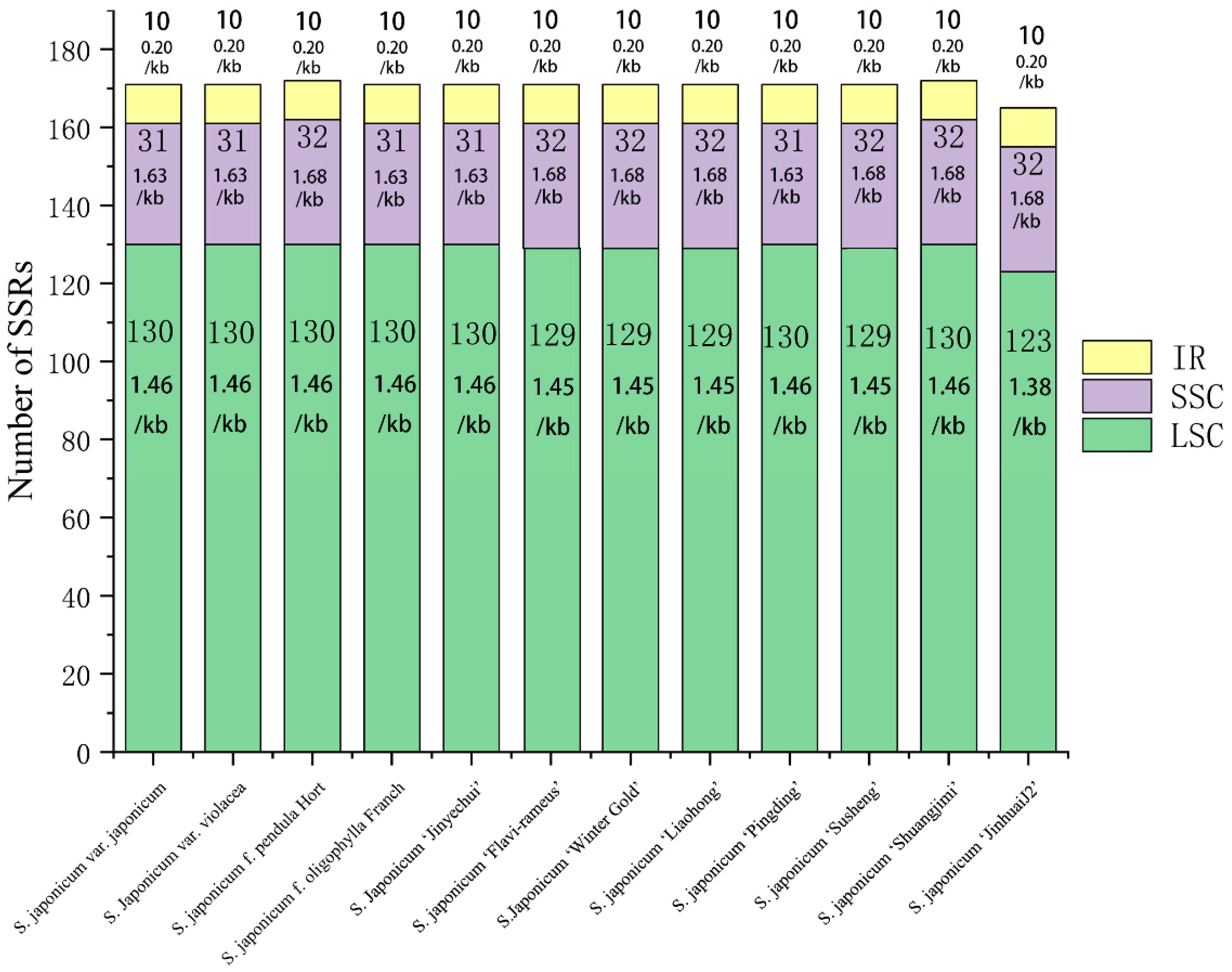
3.4. Chloroplast Genome Repetitive Sequence Analysis
3.5. Phylogenetic Analysis of the S. japonicum Species
4. Discussion
4.1. The Low Variation Level of the Chloroplast Genome in S. japonicum
4.2. Regional Differentiation of Genetic Polymorphisms in Chloroplast Genome of S. japonicum
4.3. Intraspecific Phylogenetic Relationship of S. japonicum
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, X.; Wang, Y.; Zhang, B.; Lin, Z. Research proaress on herbaceous, chemical constituents and pharmacoloaical effects of different medicinal parts of Styphnolobium japonicum. Chin. Tradit. Herb. Drugs 2018, 49, 4461–4467. [Google Scholar]
- Li, R.; Yuan, S.; Xiao, Y. Research progress on chemical constituents, pharmacological effects and processing of traditional Chinese medicine Flos Styphnolobium japonicum. Chin. J. Inf. Tradit. Chin. Med. 2002, 6, 77–82. [Google Scholar]
- Liu, L.; Li, X.; Chen, Y.; Shi, P.; Qu, L.; Zhang, Y. solation and identification of chemical constituents from Flos Styphnolobium japonicum. J. Tianjin Univ. Tradit. Chin. Med. 2014, 33, 230–233. [Google Scholar]
- Sun, L.; Sui, D.; Zhu, F. Development of Flos Styphnolobium japonicum and its clarified juice beverage. Non-Wood For. Res. 1998, 1, 47–49. [Google Scholar]
- Wang, X.; Wang, X. Development of Styphnolobium japonicum and Honey Healthy Compound Beverage. Hubei Agric. Sci. 2009, 48, 2827–2830. [Google Scholar]
- Abdelhady, M.I.S.; Kamal, A.M.; Othman, S.M.; Mubarak, M.S.; Hadda, T.B. Total Polyphenolic Content, Antioxidant, Cytotoxic, Antidiabetic Activities, and Polyphenolic Compounds of Sophora japonica Grown in Egypt. Med. Chem. Res. 2015, 24, 482–495. [Google Scholar] [CrossRef]
- Paniwnyk, L.; Beaufoy, E.; Lorimer, J.P.; Mason, T.J. The Extraction of Rutin from Flower Buds of Sophora japonica. Ultrason. Sonochem. 2001, 8, 299–301. [Google Scholar] [CrossRef]
- Wang, J.; Feng, L.; Wei, H.; Zheng, C.; Wang, J. Determination of sophoricoside in different habitats and different medicinal parts of Fructus sophoraein Shaanxi province. Northwest Pharm. J. 2011, 26, 412–415. [Google Scholar]
- Zhang, C.; Shen, Y.; Yi, W. Comparison Study on Photosynthesis of Pagoda Tree and Walnut Seedlings under Salinity. For. Res. 2002, 1, 41–46. [Google Scholar]
- Gao, M. Cultivation Technology of Ornamental Tree Species Sophora japonica Greening Seedlings. For. By-Prod. Spec. China 2023, 1, 37–38. [Google Scholar]
- Kite, G.C.; Pennington, R.T. Quinolizidine Alkaloid Status of Styphnolobium and Cladrastis (Leguminosae). Biochem. Syst. Ecol. 2003, 31, 1409–1416. [Google Scholar] [CrossRef]
- Palomino, G.; Martinez, P.; Bernal, C.; Sousa, M. Diferencias Cromosomicas Entre Algunas Especies de Los Generos Sophora L. Y Styphnolobium Schott. Ann. Mo. Bot. Gard. 1993, 80, 284–290. [Google Scholar] [CrossRef]
- Sousa, M.; Rudd, V.E. Revision Del Genero Styphnolobium (Leguminosae: Papilionoideae: Sophoreae). Ann. Mo. Bot. Gard. 1993, 80, 270–283. [Google Scholar] [CrossRef]
- Käss, E.; Wink, M. Phylogenetic Relationships in the Papilionoideae (Family Leguminosae) Based on Nucleotide Sequences of CpDNA (RbcL) and NcDNA (ITS 1 and 2). Mol. Phylogenet. Evol. 1997, 8, 65–88. [Google Scholar] [CrossRef]
- Duan, L.; Harris, A.; Ye, W.; Deng, S.-W.; Song, Z.-Q.; Chen, H.-F.; Wen, J. Untangling the Taxonomy of the Cladrastis Clade (Leguminosae: Papilionoideae) by Integrating Phylogenetics and Ecological Evidence. Taxon 2019, 68, 1189–1203. [Google Scholar] [CrossRef]
- Choi, I.-S.; Choi, B.-H. The Distinct Plastid Genome Structure of Maackia fauriei (Fabaceae: Papilionoideae) and Its Systematic Implications for Genistoids and Tribe Sophoreae. PLoS ONE 2017, 12, e0173766. [Google Scholar] [CrossRef]
- Li, X.; Cai, Q. Study of Isozyme Zymograms of Six Species of Pogada Trees. J. Southwest Univ. (Nat. Sci. Ed.) 2006, 6, 950–952. [Google Scholar]
- Heenan, P.B.; Dawson, M.I.; Wagstaff, S.J. The Relationship of Sophora Sect. Edwardsia (Fabaceae) to Sophora tomentosa, the Type Species of the Genus Sophora, Observed from DNA Sequence Data and Morphological Characters. Bot. J. Linn. Soc. 2004, 146, 439–446. [Google Scholar] [CrossRef]
- Lu, Y.; Li, W.; Xie, X.; Zheng, Y.; Li, B. The Complete Chloroplast Genome Sequence of Sophora japonica var. violacea: Gene Organization and Genomic Resources. Conserv. Genet. Resour. 2018, 10, 1–4. [Google Scholar] [CrossRef]
- Shi, Y.; Liu, B. Complete Chloroplast Genome Sequence of Sophora japonica ‘JinhuaiJ2′ (Papilionaceae), an Important Traditional Chinese Herb. Mitochondrial DNA Part B 2020, 5, 319–320. [Google Scholar] [CrossRef]
- Sun, R.X.; Zhang, C.H.; Zheng, Y.Q.; Zong, Y.C.; Yu, X.D.; Huang, P. Molecular Identification and Genetic Variation of Varieties of Styphnolobium japonicum (Fabaceae) Using SRAP Markers. Genet. Mol. Res. 2016, 15, gmr.15027837. [Google Scholar] [CrossRef] [PubMed]
- Chase, M.W.; Palmer, J.D. Chloroplast DNA Systematics of Lilioid Monocots: Resources, Feasibility, and an Example from the Orchidaceae. Am. J. Bot. 1989, 76, 1720–1730. [Google Scholar] [CrossRef]
- Daniell, H.; Lin, C.-S.; Yu, M.; Chang, W.-J. Chloroplast Genomes: Diversity, Evolution, and Applications in Genetic Engineering. Genome Biol. 2016, 17, 134. [Google Scholar] [CrossRef] [PubMed]
- Tonti-Filippini, J.; Nevill, P.G.; Dixon, K.; Small, I. What Can We Do with 1000 Plastid Genomes? Plant J. 2017, 90, 808–818. [Google Scholar] [CrossRef]
- Goremykin, V.V.; Holland, B.; Hirsch-Ernst, K.I.; Hellwig, F.H. Analysis of Acorus Calamus Chloroplast Genome and Its Phylogenetic Implications. Mol. Biol. Evol. 2005, 22, 1813–1822. [Google Scholar] [CrossRef]
- Yang, Q.; Jiang, M.; Wang, L.; Chen, H. Complete chloroplast genome of Allium chinense: Comparative genomic and phylogenetic analysis. Acta Pharm. Sin. 2019, 54, 173–181. [Google Scholar] [CrossRef]
- Daniell, H.; Khan, M.S.; Allison, L. Milestones in Chloroplast Genetic Engineering: An Environmentally Friendly Era in Biotechnology. Trends Plant Sci. 2002, 7, 84–91. [Google Scholar] [CrossRef]
- Boynton, J.E.; Gillham, N.W.; Harris, E.H.; Hosler, J.P.; Johnson, A.M.; Jones, A.R.; Randolph-Anderson, B.L.; Robertson, D.; Klein, T.M.; Shark, K.B.; et al. Chloroplast Transformation in Chlamydomonas with High Velocity Microprojectiles. Science 1988, 240, 1534–1538. [Google Scholar] [CrossRef]
- Olejniczak, S.A.; Łojewska, E.; Kowalczyk, T.; Sakowicz, T. Chloroplasts: State of Research and Practical Applications of Plastome Sequencing. Planta 2016, 244, 517–527. [Google Scholar] [CrossRef]
- Bock, R. Plastid Biotechnology: Prospects for Herbicide and Insect Resistance, Metabolic Engineering and Molecular Farming. Curr. Opin. Biotechnol. 2007, 18, 100–106. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An Integrated and Extendable Desktop Software Platform for the Organization and Analysis of Sequence Data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Greiner, S.; Le hwark, P.; Bock, R. Organellar Genome DRAW (OGDRAW) Version 1.3.1: Expanded Toolkit for the Graphical Visualization of Organellar Genomes. Nucleic Acids Res. 2019, 47, W59–W64. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Amiryousefi, A.; Hyvönen, J.; Poczai, P. IRscope: An Online Program to Visualize the Junction Sites of Chloroplast Genomes. Bioinformatics 2018, 34, 3030–3031. [Google Scholar] [CrossRef]
- Beier, S.; Thiel, T.; Münch, T.; Scholz, U.; Mascher, M. MISA-Web: A Web Server for Microsatellite Prediction. Bioinformatics 2017, 33, 2583–2585. [Google Scholar] [CrossRef]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An Integrated and Scalable Desktop Platform for Streamlined Molecular Sequence Data Management and Evolutionary Phylogenetics Studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Shimada, H.; Sugiura, M. Fine Structural Features of the Chloroplast Genome: Comparison of the Sequenced Chloroplast Genomes. Nucleic Acids Res. 1991, 19, 983–995. [Google Scholar] [CrossRef]
- Sinn, B.T.; Sedmak, D.D.; Kelly, L.M.; Freudenstein, J.V. Total Duplication of the Small Single Copy Region in the Angiosperm Plastome: Rearrangement and Inverted Repeat Instability in Asarum. Am. J. Bot. 2018, 105, 71–84. [Google Scholar] [CrossRef]
- Jiang, M.; Xu, Y.; Wang, L.; Liu, J.; Yu, J.; Chen, H. High Level of Intraspecific Divergence and Low Frequency of RNA Editing in the Chloroplast Genome Sequence of Tagetes erecta. Mitochondrial DNA B Resour. 2020, 5, 2948–2953. [Google Scholar] [CrossRef] [PubMed]
- Muraguri, S.; Xu, W.; Chapman, M.; Muchugi, A.; Oluwaniyi, A.; Oyebanji, O.; Liu, A. Intraspecific Variation within Castor Bean (Ricinus communis L.) Based on Chloroplast Genomes. Ind. Crops Prod. 2020, 155, 112779. [Google Scholar] [CrossRef]
- Zhang, R.-S.; Yang, J.; Hu, H.-L.; Xia, R.-X.; Li, Y.-P.; Su, J.-F.; Li, Q.; Liu, Y.-Q.; Qin, L. A High Level of Chloroplast Genome Sequence Variability in the Sawtooth Oak Quercus acutissima. Int. J. Biol. Macromol. 2020, 152, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.-H.; Kim, S.-C. Comparative Analysis of the Complete Chloroplast Genome Sequences of Three Closely Related East-Asian Wild Roses (Rosa Sect. Synstylae; Rosaceae). Genes 2019, 10, 23. [Google Scholar] [CrossRef] [PubMed]
- Pham, M.H.; Tran, T.H.; Le, T.D.; Le, T.L.; Hoang, H.; Chu, H.H. The Complete Chloroplast Genome of An Ophiorrhiza baviensis Drake Species Reveals Its Molecular Structure, Comparative, and Phylogenetic Relationships. Genes 2023, 14, 227. [Google Scholar] [CrossRef]
- Yu, J.; Fu, J.; Fang, Y.; Xiang, J.; Dong, H. Complete Chloroplast Genomes of Rubus Species (Rosaceae) and Comparative Analysis within the Genus. BMC Genom. 2022, 23, 32. [Google Scholar] [CrossRef]
- Saldaña, C.L.; Rodriguez-Grados, P.; Chávez-Galarza, J.C.; Feijoo, S.; Guerrero-Abad, J.C.; Vásquez, H.V.; Maicelo, J.L.; Jhoncon, J.H.; Arbizu, C.I. Unlocking the Complete Chloroplast Genome of a Native Tree Species from the Amazon Basin, Capirona (Calycophyllum spruceanum, Rubiaceae), and Its Comparative Analysis with Other Ixoroideae Species. Genes 2022, 13, 113. [Google Scholar] [CrossRef]
- Lin, Z.; Zhou, P.; Ma, X.; Deng, Y.; Liao, Z.; Li, R.; Ming, R. Comparative Analysis of Chloroplast Genomes in Vasconcellea pubescens A. DC. and Carica papaya L. Sci. Rep. 2020, 10, 15799. [Google Scholar] [CrossRef]
- Wang, Y.-C.; Zhou, H.-Y.; Liu, X.-Q. Chloroplast Genomic Variation in Euonymus maackii Rupr. and Its Differentiation Time in Euonymus. Forests 2022, 13, 265. [Google Scholar] [CrossRef]
- Su, Q.; Liu, L.; Zhao, M.; Zhang, C.; Zhang, D.; Li, Y.; Li, S. The Complete Chloroplast Genomes of Seventeen Aegilops tauschii: Genome Comparative Analysis and Phylogenetic Inference. PeerJ 2020, 8, e8678. [Google Scholar] [CrossRef]
- Li, P.; Lou, G.; Cai, X.; Zhang, B.; Cheng, Y.; Wang, H. Comparison of the Complete Plastomes and the Phylogenetic Analysis of Paulownia Species. Sci. Rep. 2020, 10, 2225. [Google Scholar] [CrossRef]
- Li, X.; Zuo, Y.; Zhu, X.; Liao, S.; Ma, J. Complete Chloroplast Genomes and Comparative Analysis of Sequences Evolution among Seven Aristolochia (Aristolochiaceae) Medicinal Species. Int. J. Mol. Sci. 2019, 20, 1045. [Google Scholar] [CrossRef]
- Dong, W.; Liu, J.; Yu, J.; Wang, L.; Zhou, S. Highly Variable Chloroplast Markers for Evaluating Plant Phylogeny at Low Taxonomic Levels and for DNA Barcoding. PLoS ONE 2012, 7, e35071. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, J.; Xia, T.; Zhou, S. DNA Barcoding: Species Delimitation in Tree Peonies. Sci. China Life Sci. 2009, 52, 568–578. [Google Scholar] [CrossRef]
- Park, J.; Xi, H.; Kim, Y. The Complete Chloroplast Genome of Arabidopsis Thaliana Isolated in Korea (Brassicaceae): An Investigation of Intraspecific Variations of the Chloroplast Genome of Korean A. thaliana. Int. J. Genom. 2020, 2020, 3236461. [Google Scholar] [CrossRef]
- Silva, S.R.; Pinheiro, D.G.; Penha, H.A.; Płachno, B.J.; Michael, T.P.; Meer, E.J.; Miranda, V.F.O.; Varani, A.M. Intraspecific Variation within the Utricularia amethystina Species Morphotypes Based on Chloroplast Genomes. Int. J. Mol. Sci. 2019, 20, 6130. [Google Scholar] [CrossRef]
- Liu, Y. Cultivation and Pruning Technology of S. japonicum f. pendula. Hebei Agric. 2018, 7, 39–40. [Google Scholar]
- Jiao, F.; Tan, R. Characteristics and Cultivation Techniques of New Variety of S. japonicum ‘Shuangjimi’. J. Shandong For. Sci. Technol. 2005, 6, 46–47. [Google Scholar]
- Zhao, P.; Zhang, H.; Li, Y. Study on Introduction and Cultivation of S. japonicum ‘Flavi-Rameus’. J. Beijing Vocat. Coll. Agric. 2015, 4, 19–24. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Names | GenBank | Size/bp | No. of Genes | GC Content% | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Genome | LSC | SSC | IR | Genome | PCG | tRNA | rRNA | Genome | LSC | SSC | IR | ||
| S. japonicum var. japonicum | MG784459 | 158,738 | 89,003 | 19,057 | 50,678 | 129 | 83 | 38 | 8 | 36.1% | 33.5 | 29.61 | 43.15 |
| S. Japonicum var. violacea | KY872756 | 158,837 | 88,983 | 19,058 | 50,796 | 129 | 83 | 38 | 8 | 36.1% | 33.51 | 29.61 | 43.17 |
| S. japonicum f. pendula | ON571617 | 158,645 | 88,905 | 19,060 | 50,680 | 129 | 83 | 38 | 8 | 36.1% | 33.53 | 29.6 | 43.15 |
| S. japonicum f. oligophylla | ON571618 | 158,739 | 89,003 | 19,058 | 50,678 | 129 | 83 | 38 | 8 | 36.1% | 33.5 | 29.61 | 43.15 |
| S. Japonicum ‘Jinyechui’ | ON571616 | 158,613 | 88,871 | 19,062 | 50,680 | 129 | 83 | 38 | 8 | 36.1% | 33.54 | 29.6 | 43.15 |
| S. japonicum ‘Flavi-rameus’ | N571615 | 158,616 | 88,868 | 19,068 | 50,680 | 129 | 83 | 38 | 8 | 36.1% | 33.54 | 29.59 | 43.15 |
| S. Japonicum ‘Winter Gold’ | ON571614 | 158,624 | 88,876 | 19,068 | 50,680 | 129 | 83 | 38 | 8 | 36.1% | 33.54 | 29.59 | 43.15 |
| S. japonicum ‘Liaohong’ | ON571620 | 158,646 | 88,897 | 19,069 | 50,680 | 129 | 83 | 38 | 8 | 36.1% | 33.53 | 29.59 | 43.15 |
| S. japonicum ‘Pingding’ | ON571621 | 158,613 | 88,873 | 19,062 | 50,678 | 129 | 83 | 38 | 8 | 36.1% | 33.54 | 29.6 | 43.15 |
| S. japonicum ‘Susheng’ | ON553276 | 158,629 | 88,876 | 19,073 | 50,680 | 129 | 83 | 38 | 8 | 36.1% | 33.54 | 29.58 | 43.15 |
| S. japonicum ‘Shuangjimi’ | ON571619 | 158,639 | 88,907 | 19,054 | 50,678 | 129 | 83 | 38 | 8 | 36.1% | 33.53 | 29.61 | 43.15 |
| S. japonicum ‘JinhuaiJ2’ | MN701078 | 158,690 | 88,977 | 19,034 | 50,680 | 129 | 83 | 38 | 8 | 36.1% | 33.51 | 29.65 | 43.15 |
| Category | Gene Group | Gene Name |
|---|---|---|
| Photosynthesis | Subunits of photosystem I | psaA, psaB, psaC, psaI, psaJ |
| Subunits of photosystem II | psbA, psbB, psbC, psbD, psbE, psbF, psbH, psbI, psbJ, psbK, psbL, psbM, psbN, psbT, psbZ | |
| Subunits of NADH dehydrogenase | ndhA *, ndhB *(2), ndhC, ndhD, ndhE, ndhF, ndhG, ndhH, ndhI, ndhJ, ndhK | |
| Subunits of cytochrome b/f complex | petA, petB *, petD *, petG, petL, petN | |
| Subunits of ATP synthase | atpA, atpB, atpE, atpF *, atpH, atpI | |
| Large subunit of rubisco | rbcL | |
| Self-replication | Proteins of large ribosomal subunit | rpl14, rpl16 *, rpl2 *(2), rpl20, rpl23 (2), rpl33, rpl36 |
| Proteins of small ribosomal subunit | rps11, rps12 *(2), rps14, rps15, rps16 *, rps18, rps19, rps2, rps3, rps4, rps7 (2), rps8 | |
| Subunits of RNA polymerase | rpoA, rpoB, rpoC1 *, rpoC2 | |
| Ribosomal RNAs | rrn16S (2), rrn23S (2), rrn4.5S (2), rrn5S (2) | |
| Transfer RNAs | trnA-UGC *(2), trnC-GCA, trnD-GUC, trnE-UUC, trnF-GAA, trnG-GCC, trnH-GUG, trnI-CAU (2), trnI-GAU *(2), trnK-UUU *, trnL-CAA (2), trnL-UAA *, trnL-UAG, trnM-CAU, trnN-GUU (2), trnP-GGG, trnP-UGG, trnQ-UUG, trnR-ACG (2), trnR-UCU, trnS-GCU, trnS-GGA, trnS-UGA, trnT-CGU *, trnT-GGU, trnT-UGU, trnV-GAC(2), trnV-UAC *, trnW-CCA, trnY-GUA, trnfM-CAU | |
| Other genes | Maturase | matK |
| Protease | clpP ** | |
| Envelope membrane protein | cemA | |
| Acetyl-CoA carboxylase | accD | |
| c-type cytochrome synthesis gene | ccsA | |
| Genes of unknown function | Conserved hypothetical chloroplast ORF | ycf1 (2), ycf2 (2), ycf3 **, ycf4 |
| Genome | LSC | IRa | SSC | Noncoding Region | Coding Region | |
|---|---|---|---|---|---|---|
| Total number of sites | 159,102 | 89,215 | 25,399 | 19,089 | 81,133 | 78,072 |
| Number of polymorphic sites | 132 | 59 | 18 | 38 | 87 | 45 |
| Pi | 0.00029 | 0.00026 | 0.00014 | 0.00085 | 0.00043 | 0.00019 |
| Theta-W: | 0.00028 | 0.00022 | 0.00024 | 0.00066 | 0.00043 | 0.00019 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mu, Z.; Zhang, Y.; Zhang, B.; Cheng, Y.; Shang, F.; Wang, H. Intraspecific Chloroplast Genome Variation and Domestication Origins of Major Cultivars of Styphnolobium japonicum. Genes 2023, 14, 1156. https://doi.org/10.3390/genes14061156
Mu Z, Zhang Y, Zhang B, Cheng Y, Shang F, Wang H. Intraspecific Chloroplast Genome Variation and Domestication Origins of Major Cultivars of Styphnolobium japonicum. Genes. 2023; 14(6):1156. https://doi.org/10.3390/genes14061156
Chicago/Turabian StyleMu, Zhiqiang, Yu Zhang, Bin Zhang, Yueqin Cheng, Fude Shang, and Hongwei Wang. 2023. "Intraspecific Chloroplast Genome Variation and Domestication Origins of Major Cultivars of Styphnolobium japonicum" Genes 14, no. 6: 1156. https://doi.org/10.3390/genes14061156