
Chromosomal Evolution of the Talpinae

 ,
,  , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
3. Results
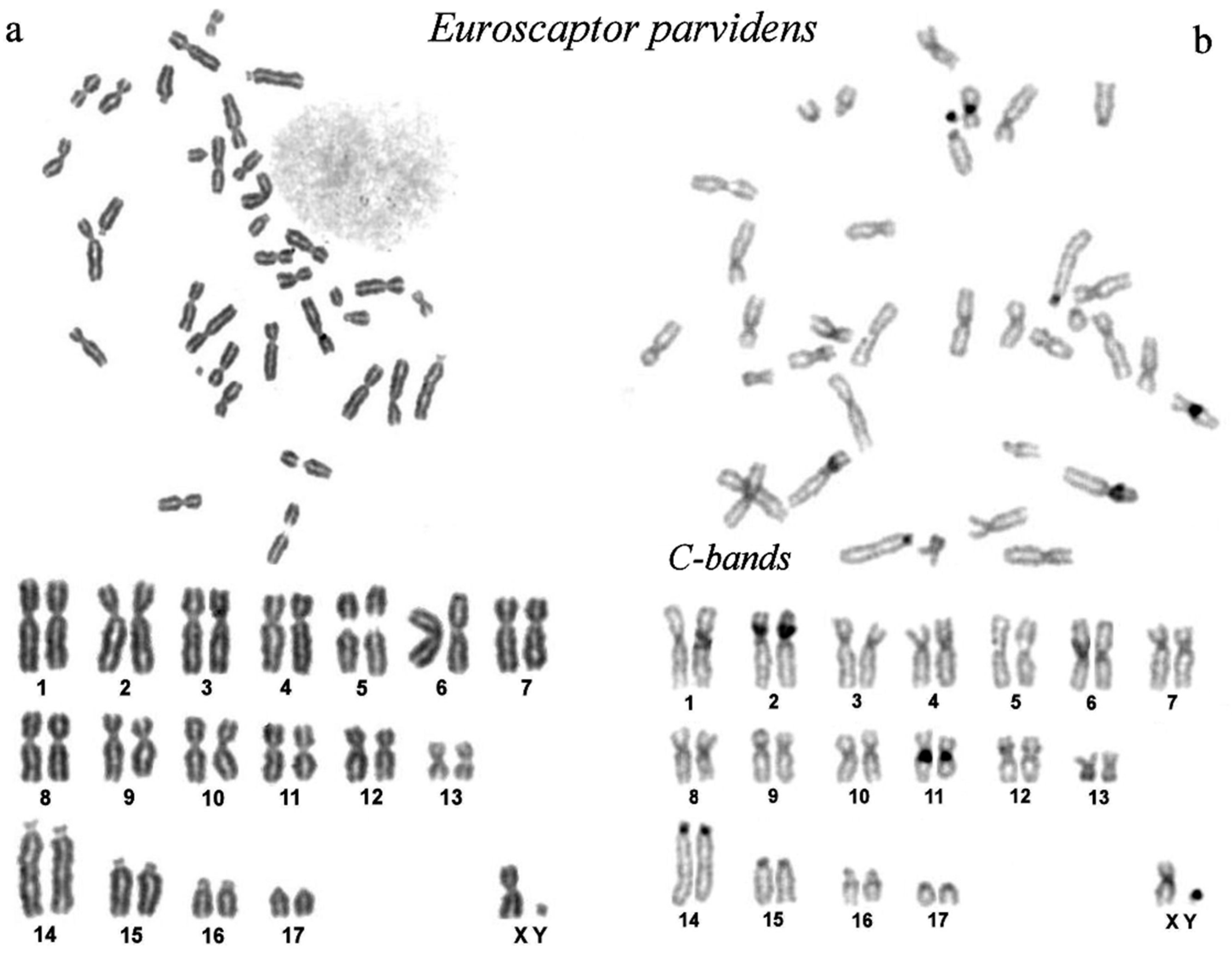
3.1. Description of the Karyotype of the Small-Toothed Mole (Euroscaptor parvidens) from Vietnam
3.2. Comparison of G-Banded Chromosomes of Euroscaptor Species
- -
- Seven autosomal pairs of the small-toothed mole (chromosomes 5, 8, 10–13, and 15) are homologous to chromosomes 2–7, 10, and 15, respectively, in the all compared species, as well as their X chromosomes;
- -
- Accumulation of repeats in the short arm changed the morphology of the submetacentric chromosome 2 of E. parvidens compared to the homologous chromosome 8 of other species;
- -
- Four metacentric chromosomes of the small-toothed mole (chromosomes 3, 4, 7, and 9) underwent pericentric inversions in contrast to the homologous acrocentric 11, 12, 13, and 14, respectively, of other species;
- -
- A small inversion of the pericentromeric heterochromatin distinguishes the subtelocentric chromosomes 14 of E. parvidens, 10 of E. klossi (EKLO), and the acrocentric chromosome 10 of both O. mizura (OMIZ) and E. malayana (EMAL), respectively;
- -
- Centric fusions/fissions separate chromosomes/chromosomal arms:
- (a)
- OMIZ 1 and EPAR 6 with EKLO 16+17 and EMAL 1p+16;
- (b)
- OMIZ 4, EKLO 4, and EMAL 4 with EPAR 16+17;
- (c)
- OMIZ 17+16 and EMIC 17+14 with EKLO 1 and EPAR 1.
3.3. Chromosome Painting
3.3.1. Assignment of the Peaks of the Flow-Sorted Karyotype
3.3.2. Cross-Species Chromosome Painting
Painting of the E. parvidens Karyotype with T. altaica Probes
- Chromosome painting of Mogera imaizumii with T. altaica probes
- 2.
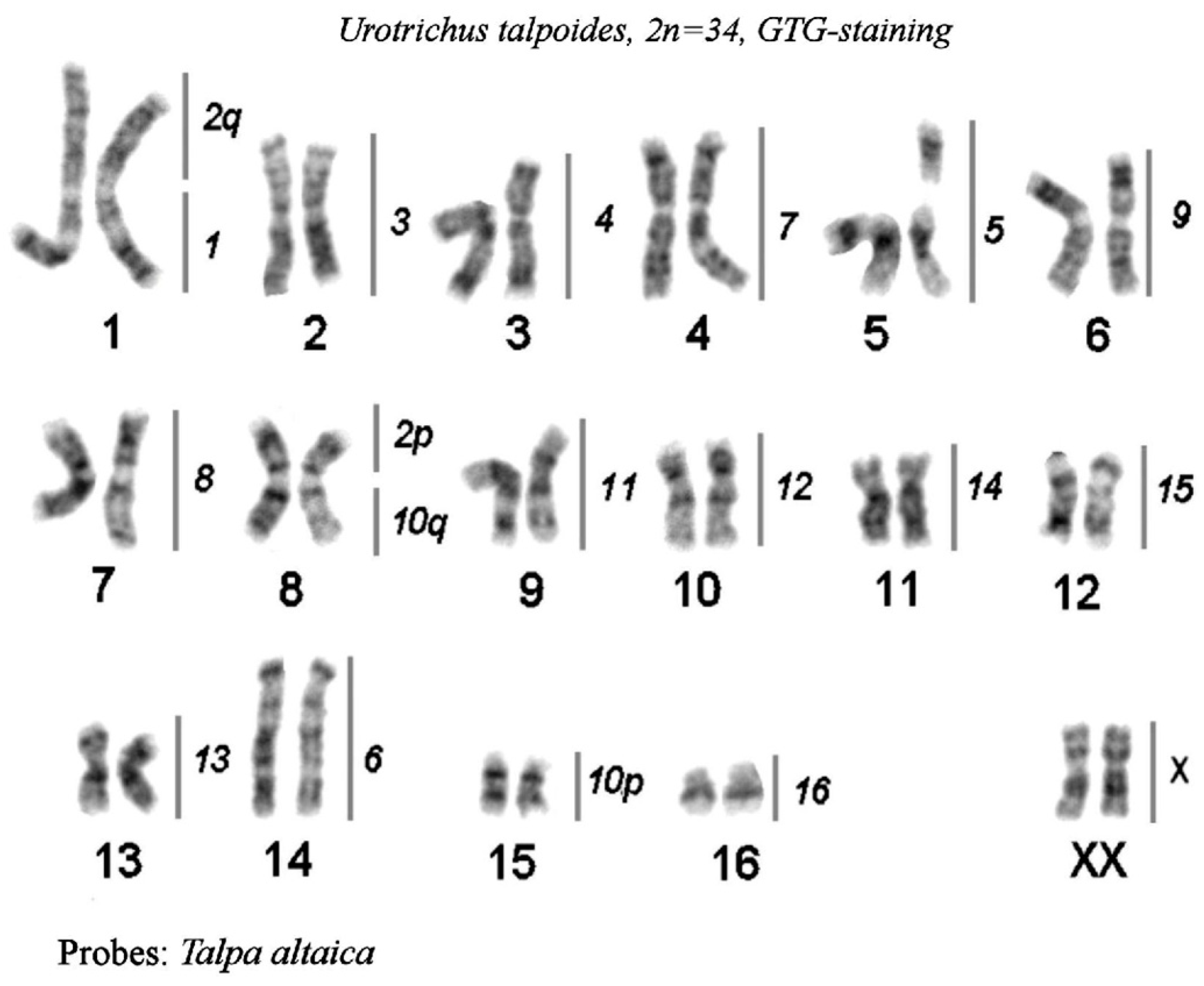
- Chromosome painting between Urotrichus talpoides and T. altaica
3.4. Comparison of Karyotypes of Four Species from the Talpinae Subfamily
- -
- Six pairs of autosomes homologous to chromosomes 3, 5, 12, and 14–16 of the Siberian mole remained conserved, as did the X chromosomes in all species;
- -
- Amplification of additional heterochromatin occurred in the p-arm of EPAR 2. A small block of centromeric heterochromatin is present in homologs from all compared species;
- -
- Five pairs of autosomes homologous to chromosomes 4, 7–9, and 13 of the Siberian mole differ by pericentric inversions and/or centromeric shifts;
- -
- Four pairs of autosomes homologous to chromosomes 1, 2, 10, and 11 of the Siberian mole are involved in chromosome fusion/fission.
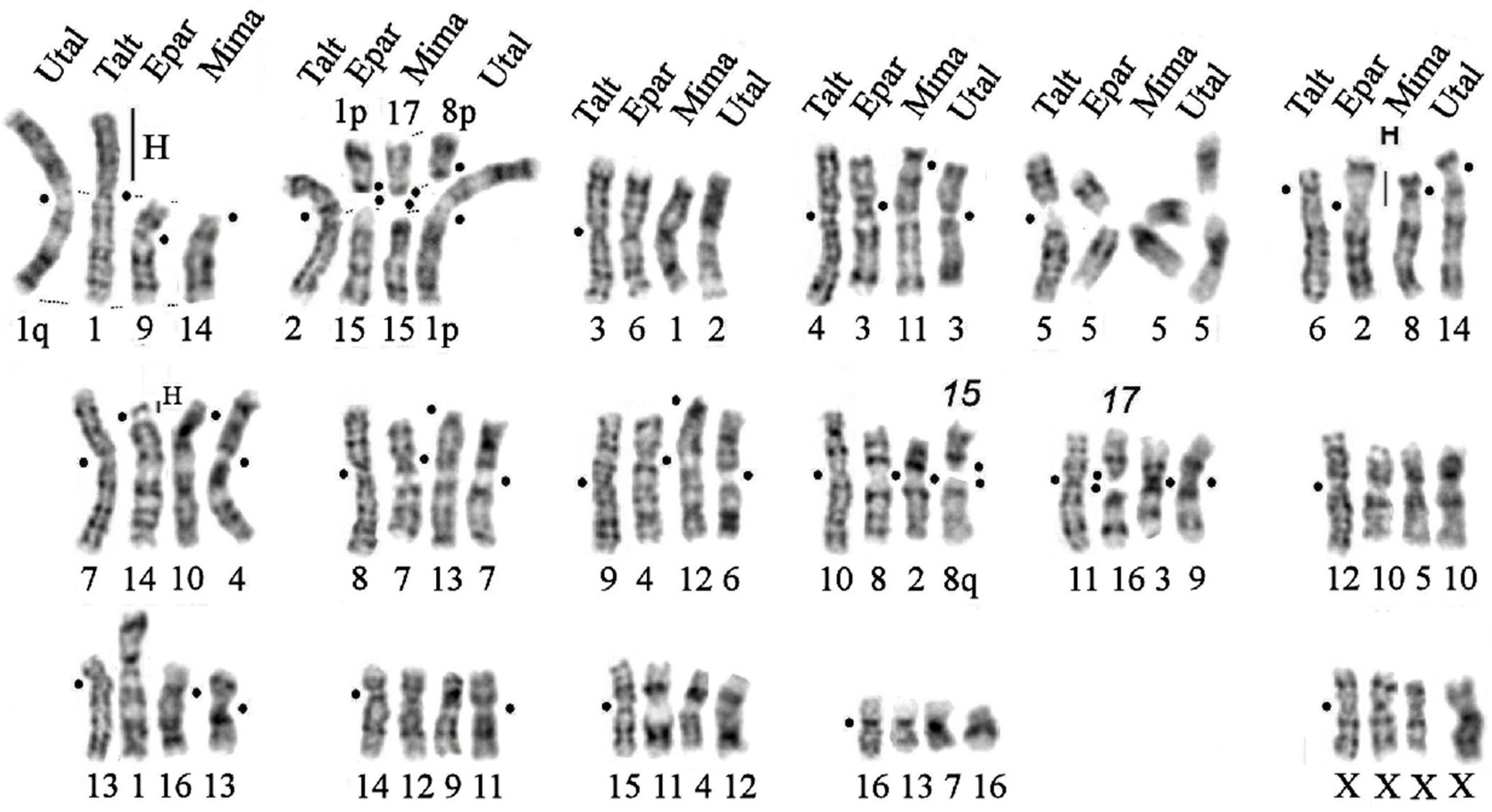
3.5. Comparison of Karyotypes of Eight Species from the Subfamily Talpinae
- -
- Five autosomal pairs homologous to chromosomes 5, 12, and 14–16 and the X chromosome of the Siberian mole remained unchanged in all eight mole species;
- -
- Chromosomal pairs homologous to TALT 6 are characterized by the amplification of heterochromatic blocks in the centromeric regions of chromosomes TEUR 1p and EPAR 2p (Figure 8a). Small blocks of heterochromatin are present in the centromeric region of all studied species;
- -
- Whole arm homology is generally preserved:
- -
- Inversions/centromeric shifts occurred on six chromosomes:
- (a)
- The chromosomal arms TALT 1q, TEUR 9q, and UTAL 1q are homologous to the acrocentrics EKLO 14, EMAL 13, MIMA 14, and OMIZ 14, respectively. Pericentric inversions of the proximal part of the q-arms produced submetacentric chromosomes EPAR 9 and MIMI 14. The short arms of TALT 1 and TEUR 9 are composed of heterochromatic blocks. UTAL1 resulted from a centric fusion of two ancestral acrocentrics (Figure 8f).
- (b)
- Chromosomes TALT 4, TEUR 3, and UTAL 3 are homologous; pericentric inversions of their p-arms led to the appearance of acrocentric chromosomes 11 in EKLO, OMIZ, and EMAL. A subsequent pericentric inversion of the subcentromeric region led to the appearance of the submetacentric chromosome MIMA 11. A centromeric shift probably led to the appearance of the submetacentric chromosome EPAR 3 (Figure 8g).
- (c)
- Chromosomes TALT 7, TEUR 4, and UTAL 4 are homologous. Pericentric inversions of the p-arms led to the appearance of acrocentric chromosomes OMIZ 10, EMAL10, and MIMA 10. The appearance of additional heterochromatic arms on EKLO 10 and EPAR 14 explains their subtelocentric morphology (Figure 8h).
- (d)
- Chromosomes TALT 8, TEUR 7, and UTAL 7 are homologous; pericentric inversions of the p-arm lead to the appearance of acrocentric chromosomes MIMA 13, OMIZ 13, EKLO 13, and q-arm EMAL1. The appearance of the submetacentric EPAR 7 can be explained by the centromeric shift followed by an inversion (Figure 8i).
- (e)
- Chromosomes TALT 9, TEUR 6, and UTAL 6 are homologous; pericentric inversions of the p-arms led to the appearance of acrocentric chromosomes MIMA 12, OMIZ 12, EKLO12, and EMAL12. A subsequent pericentric inversion of the proximal part of an ancestral acrocentric led to the appearance of a submetacentric EPAR 4 (Figure 8j).
- (f)
- The most confusing scenario of rearrangements relates to TALT 13, TEUR 13, UTAL 13, MIMA 16, and EPAR 1: the homology of these elements was shown only by the TALT 13 painting probes, and the difference in the GTG pattern of these chromosomes can be explained by a series of inversions. For example, the chromosomes TALT 13 and TEUR 13, according to human painting probe localization and GTG-banding, differ by a pericentric inversion on TEUR 13q and the proximal part of TALT 13q. In Euroscaptor species, chromosomes EPAR 1 and EKLO 1 are similar, and both have resulted from fusions of ancestral acrocentrics, whereas their q-arms are homologous to TALT 13 and OMIZ 16. EMAL 1 is also a result of the ancestral centric fusion of two acrocentrics. Thus, TALT 13 homologs in all species have undergone multiple inversions and fusions.
4. Discussion
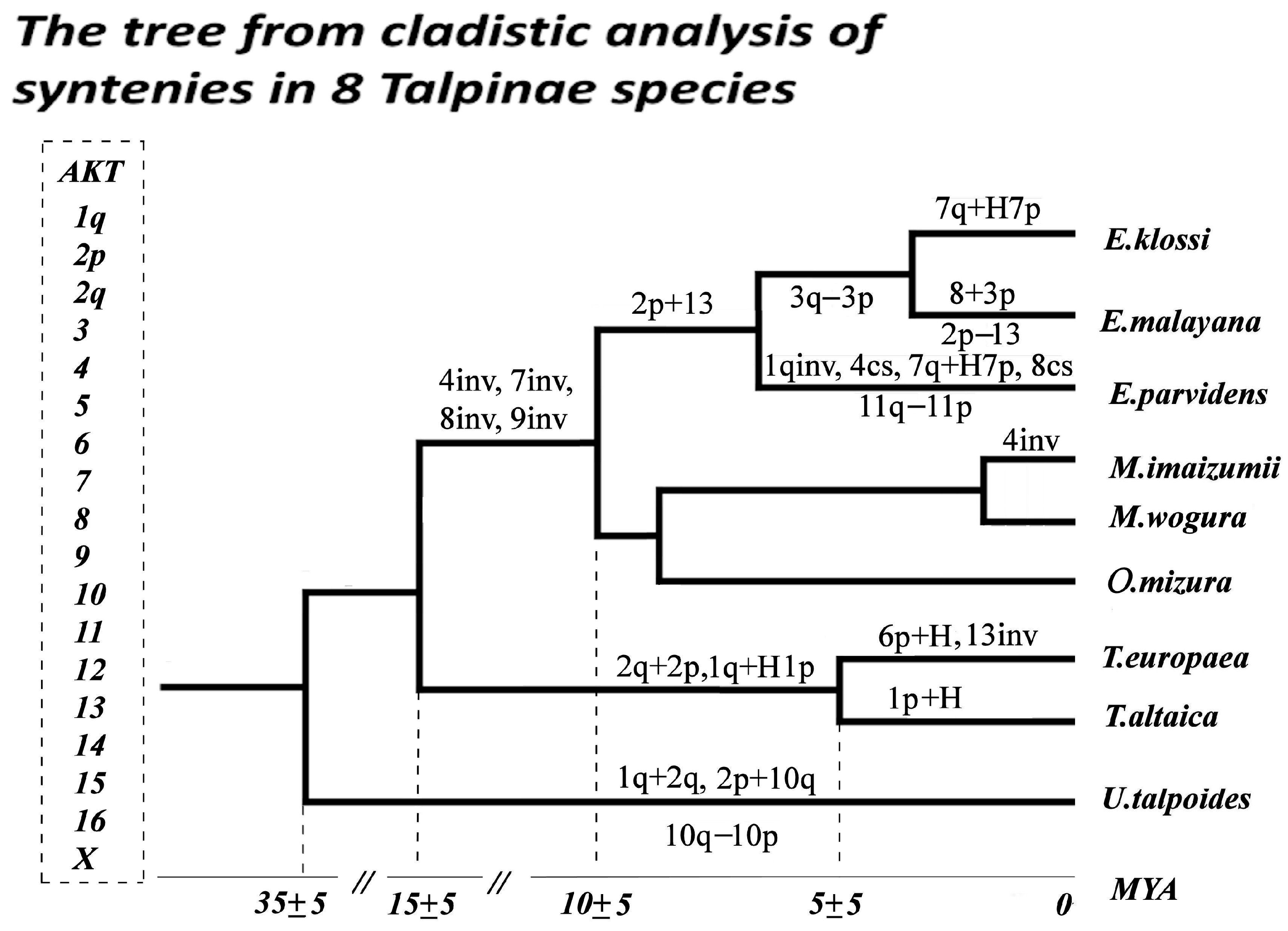
4.1. Chromosomal Rearrangements in the Genus Euroscaptor
4.2. Chromosomal Rearrangements in the Talpini and Urotrichini Tribes
4.3. Reconstruction of the Ancestral Karyotype of the Talpini and Urotrichini Tribes
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nevo, E. Adaptive Convergence and Divergence of Subterranean Mammals. Annu. Rev. Ecol. Syst. 1979, 10, 269–308. [Google Scholar] [CrossRef]
- He, K.; Shinohara, A.; Helgen, K.M.; Springer, M.S.; Jiang, X.L.; Campbell, K.L. Talpid Mole Phylogeny Unites Shrew Moles and Illuminates Overlooked Cryptic Species Diversity. Mol. Biol. Evol. 2017, 34, 78–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinohara, A.; Kawada, S.I.; Son, N.T.; Koshimoto, C.; Endo, H.; Can, D.N.; Suzuki, H. Molecular Phylogeny of East and Southeast Asian Fossorial Moles (Lipotyphla, Talpidae). J. Mammal. 2014, 95, 455–466. [Google Scholar] [CrossRef]
- Zemlemerova, E.D.; Bannikova, A.A.; Lebedev, V.S.; Rozhnov, V.V.; Abramov, A.V. Secrets of the Underground Vietnam: An Underestimated Species Diversity of Asian Moles (Lipotyphla: Talpidae: Euroscaptor). Proc. Zool. Inst. RAS 2016, 320, 193–2200. [Google Scholar] [CrossRef] [Green Version]
- Zemlemerova, E.; Abramov, A.; Kryukov, A.; Lebedev, V.; Min, M.S.; Lee, S.J.; Bannikova, A. Genetic and Morphologic Diversity of the Moles (Talpomorpha, Talpidae, Mogera) from the Continental Far East. J. Zool. Syst. Evol. Res. 2019, 57, 662–678. [Google Scholar] [CrossRef]
- Shinohara, A.; Suzuki, H.; Tsuchiya, K.; Zhang, Y.P.; Luo, J.; Jiang, X.L.; Wang, Y.X.; Campbell, K.L. Evolution and Biogeography of Talpid Moles from Continental East Asia and the Japanese Islands Inferred from Mitochondrial and Nuclear Gene Sequences. Zool. Sci. 2004, 21, 1177–1185. [Google Scholar] [CrossRef] [Green Version]
- Bannikova, A.A.; Zemlemerova, E.D.; Colangelo, P.; Sözen, M.; Sevindik, M.; Kidov, A.A.; Dzuev, R.I.; Kryštufek, B.; Lebedev, V.S. An Underground Burst of Diversity—A New Look at the Phylogeny and Taxonomy of the Genus Talpa Linnaeus, 1758 (Mammalia: Talpidae) as Revealed by Nuclear and Mitochondrial Genes. Zool. J. Linn. Soc. 2015, 175, 930–948. [Google Scholar] [CrossRef] [Green Version]
- Colangelo, P.; Bannikova, A.A.; Kryštufek, B.; Lebedev, V.S.; Annesi, F.; Capanna, E.; Loy, A. Molecular Systematics and Evolutionary Biogeography of the Genus Talpa (Soricomorpha: Talpidae). Mol. Phylogenet. Evol. 2010, 55, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Yudin, B.S. Insectivorous Mammals of Siberia; Nauka: Novosibirsk, Russian, 1989. [Google Scholar]
- Zemlemerova, E.D.; Bannikova, A.A.; Abramov, A.V.; Lebedev, V.S.; Rozhnov, V.V. New Data on Molecular Phylogeny of the East Asian Moles. Dokl. Biol. Sci. 2013, 451, 257–260. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Shinohara, A.; Jiang, X.L.; Campbell, K.L. Multilocus Phylogeny of Talpine Moles (Talpini, Talpidae, Eulipotyphla) and Its Implications for Systematics. Mol. Phylogenet. Evol. 2014, 70, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, A.; Kawada, S.I.; Son, N.T.; Can, D.N.; Sakamoto, S.H.; Koshimoto, C. Molecular Phylogenetic Relationships and Intra-Species Diversities of Three Euroscaptor Spp. (Talpidae: Lipotyphla: Mammalia) from Vietnam. Raffles Bull. Zool. 2015, 63, 366–375. [Google Scholar]
- Hai, B.T.; Motokawa, M.; Kawada, S.I.; Abramov, A.V.; Son, N.T. Skull Variation in Asian Moles of the Genus Euroscaptor (Eulipotyphla: Talpidae) in Vietnam. Mammal. Study 2020, 45, 265–280. [Google Scholar] [CrossRef]
- Kawada, S.I. Morphological Review of the Japanese Mountain Mole (Eulipotyphla, Talpidae) with the Proposal of a New Genus. Mammal. Study 2016, 41, 191–205. [Google Scholar] [CrossRef]
- Kawada, S.; Son, N.T.; Can, D.N. Moles (Insectivora, Talpidae, Talpinae) of Vietnam. CiNii Res. 2009, 35, 89–101. [Google Scholar]
- Kawada, S.I.; Harada, M.; Obara, Y.; Kobayashi, S.; Koyasu, K.; Oda, S. ichi Karyosystematic Analysis of Japanese Talpine Moles in the Genera Euroscaptor and Mogera (Insectivora, Talpidae). Zool. Sci. 2001, 18, 1003–1010. [Google Scholar] [CrossRef] [Green Version]
- Kawada, S.I.; Shinohara, A.; Kobayashi, S.; Harada, M.; Oda, S.I.; Lin, L.K. Revision of the Mole Genus Mogera (Mammalia: Lipotyphla: Talpidae) from Taiwan. Syst. Biodivers. 2007, 5, 223–240. [Google Scholar] [CrossRef] [Green Version]
- Zima, J. The Karyotype of Talpa Europaea Kratochvili (Talpidae, Insectivora). Folia Zool. 1983, 32, 131–136. [Google Scholar]
- Graphodatsky, A.; Radjabli, S.; Zaitsev, M.S.A. The Levels of Chromosome Conservatism in the Different Groups of Insectivores (Mammalia, Insectivora). In Questions of Systematics, Faunistics and Pa_laeontology of Small Mammals; Zoological Institute USSR: St. Petersburg, Russia, 1991; pp. 47–57. (In Russian) [Google Scholar]
- Kawada, S.; Harada, M.; Grafodatsky, A.S.; Oda, S. Cytogenetic Study of the Siberian Mole, Talpa Altaica (Insectivora: Talpidae) and Karyological Relationships within the Genus Talpa. Mammalia 2002, 66, 53–62. [Google Scholar] [CrossRef]
- Gornung, E.; Volleth, M.; Capanna, E.; Castiglia, R. Comparative Cytogenetics of Moles (Eulipotyphla, Talpidae): Chromosomal Differences in Talpa Romana and T. Europaea. Cytogenet. Genome Res. 2008, 121, 249–254. [Google Scholar] [CrossRef]
- Kawada, S. The Historical Notes and Taxonomic Problems of East Asian Moles, Euroscaptor, Parascaptor and Scaptochirus, of Continental Asia (Insectivora, Talpidae). Mammal. Study 2005, 30, S5–S11. [Google Scholar] [CrossRef]
- Kawada, S.; Kobayashi, S.; Endo, H.; Rerkamnuaychoke, W.; Oda, S. Karyological Study on Kloss’s Mole Euroscaptor Klossi (Insectivora, Talpidae) Collected in Chiang Rai Province, Thailand. Mammal. Study 2006, 31, 105–109. [Google Scholar] [CrossRef]
- Harada, M.; Ando, A.; Tsuchiya, K.; Koyasu, K. Geographical Variations in Chromosomes of the Greater Japanese Shrew-Mole, Urotrichus Talpoides (Mammalia: Insectivora). Zool. Sci. 2001, 18, 433–442. [Google Scholar] [CrossRef] [Green Version]
- Kawada, S.I.; Obara, Y. Reconsideration of the Karyological Relationship between Two Japanese Species of Shrew-Moles, Dymecodon Pilirostris and Urotrichus Talpoides. Zool. Sci. 1999, 16, 167–174. [Google Scholar] [CrossRef]
- Volleth, M.; Müller, S. Zoo-FISH in the European Mole (Talpa Europaea) Detects All Ancestral Boreo-Eutherian Human Homologous Chromosome Associations. Cytogenet. Genome Res. 2006, 115, 154–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biltueva, L.; Vorobieva, N. Chromosome Evolution in Eulipotyphla. Cytogenet. Genome Res. 2012, 137, 154–164. [Google Scholar] [CrossRef]
- Graphodatsky, A.S.; Yang, F.; O’Brien, P.C.M.; Perelman, P.; Milne, B.S.; Serdukova, N.; Kawada, S.I.; Ferguson-Smith, M.A. Phylogenetic Implications of the 38 Putative Ancestral Chromosome Segments for Four Canid Species. Cytogenet. Cell Genet. 2001, 92, 243–247. [Google Scholar] [CrossRef]
- Seabright, M. The Use of Proteolytic Enzymes for the Mapping of Structural Rearrangements in the Chromosomes of Man. Chromosoma 1972, 36, 204–210. [Google Scholar] [CrossRef]
- Sumner, A.T. 304 Preliminary Notes 11. Exptl. Cell Res. 1972, 75, 304–306. [Google Scholar] [CrossRef] [PubMed]
- Telenius, H.; Ponder, B.A.J.; Tunnacliffe, A.; Pelmear, A.H.; Carter, N.P.; Ferguson-Smith, M.A.; Behmel, A.; Nordenskjöld, M.; Pfragner, R. Cytogenetic Analysis by Chromosome Painting Using Dop-pcr Amplified Flow-sorted Chromosomes. Genes Chromosom. Cancer 1992, 4, 257–263. [Google Scholar] [CrossRef]
- Yang, F.; Carter, N.P.; Shi, L.; Ferguson-Smith, M.A. A Comparative Study of Karyotypes of Muntjacs by Chromosome Painting. Chromosoma 1995, 103, 642–652. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Brien, P.C.M.O.; Milne, B.S.; Graphodatsky, A.S.; Solanky, N.; Trifonov, V.; Rens, W.; Sargan, D.; Fox, R. A Complete Comparative Chromosome Map for the Dog, Red Fox, and Human and Its Integration with Canine Genetic Maps. Genomics 1999, 202, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Kawada, S.I.; Li, S.; Wang, Y.X.; Mock, O.B.; Oda, S.I.; Campbell, K.L. Karyotype Evolution of Shrew Moles (Soricomorpha: Talpidae). J. Mammal. 2008, 89, 1428–1434. [Google Scholar] [CrossRef]
- Kawada, S.; Shinohara, A.; Yasuda, M.; Oda, S.; Liat, L.B. Karyological Study of the Malaysian Mole, Euroscaptor Micrura Malayana (Insectivora, Talpidae) from Cameron Highlands, Peninsular Malaysia. Mammal. Study 2005, 30, 109–115. [Google Scholar] [CrossRef]
- Satou, M.; Kaiya, H.; Nishi, Y.; Shinohara, A.; Kawada, S.I.; Miyazato, M.; Kangawa, K.; Sugimoto, H. Mole Ghrelin: CDNA Cloning, Gene Expression, and Diverse Molecular Forms in Mogera Imaizumii. Gen. Comp. Endocrinol. 2016, 232, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, R.; Burgos, M.; de la Guardia, R.D. Karyotype and Chromosome Banding in the Mole (Talpa Occidentalis) from the South-East of the Iberian Peninsula. Implications on Its Taxonomic Position. Caryologia 1984, 37, 253–258. [Google Scholar] [CrossRef]
- Iannucci, A.; Makunin, A.; Lisachov, A.; Ciofi, C.; Stanyon, R.; Svartman, M.; Trifonov, V. Bridging the Gap between Vertebrate Cytogenetics and Genomics with Single-Chromosome Sequencing (ChromSeq). Genes 2021, 12, 124. [Google Scholar] [CrossRef]
- Lewin, H.; Graves, J.; Ryder, O.; Graphodatsky, A.; O’Brien, S. Precision nomenclature for the new genomics. GigaScience 2019, 8, giz086. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| O. mizura | E. parvidens | E. klossi | E. malayana |
|---|---|---|---|
| 1 | 6 | 16+17 | 1p+16 |
| 2 | 5 | 2 | 2 |
| 3 | 8 | 3 | 3 |
| 4 | 16+17 | 4 | 4 |
| 5 | 10 | 5 | 5 |
| 6 q H | 11+q H | 6+H | 6+H |
| 7 | 13 | 7 | 7 |
| 8 | 2+p H | 8 | 8 |
| 9 | 12 | 9 | 9 |
| 10 q H | 14+p H | 10+p H | 10+q H |
| 11 | 3 inv | 11 | 11 |
| 12 | 4 inv | 12 | 12 |
| 13 | 7 cs | 13 | 1q |
| 14 | 9inv | 14 | 13 |
| 15 | 15 | 15 | 15 |
| 16 | 1q | 1q | 14 |
| 17 | 1p | 1p | 17 |
| T. altaica | T. europaea | U. talpoides | O. mizura | M. imaizumii | E. klossi | E. malayana | E. parvidens |
|---|---|---|---|---|---|---|---|
| 1q (+p H) | 9q (+p H) | 1q | 14 | 14 | 14 | 13 | 9 inv |
| 2 | 2 | 1p+8p | 17+15 | 15+17 | 1p+15 | 17+15 | 1p+15 |
| 3 | 5 | 2 | 1 | 1 | 16+17 | 1p+16 | 6 |
| 4 | 3 | 3 | 11 inv | 11 inv+inv | 11 inv | 11 inv | 3 inv+cs |
| 5 | 8 | 5 | 2 | 5 | 2 | 2 | 5 |
| 6q | 1+p H | 14 | 8q | 8q | 8q | 8q | 2+p H |
| 7 | 4 | 4 | 10 inv | 10 inv | 10 inv+p H | 10 inv | 14 inv+p H |
| 8 | 7 | 7 | 13 inv | 13 inv | 13 inv | 1q inv (+1p) | 7 inv+cs |
| 9 | 6 | 6 | 12 inv | 12 inv | 12 inv | 12 inv | 4 inv+inv |
| 10 | 10 | 8q+15 | 3 | 2 | 3 | 3 | 8 |
| 11 | 12 | 9 | 4 | 3 | 4 | 4 | 16+17 |
| 12 | 11 | 10 | 5 | 5 | 5 | 5 | 10 |
| 13 | 13 inv | 13 | 16 | 16 | 1q | 14 | 1q |
| 14 | 14 | 11 | 9 | 9 | 9 | 9 | 12 |
| 15 | 15 | 12 | 6 | 4 | 6 | 6 | 11 |
| 16 | 16 | 16 | 7 | 7 | 7 | 7 | 13 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biltueva, L.S.; Vorobieva, N.V.; Lemskya, N.A.; Perelman, P.L.; Trifonov, V.A.; Panov, V.V.; Abramov, A.V.; Kawada, S.-i.; Serdukova, N.A.; Graphodatsky, A.S. Chromosomal Evolution of the Talpinae. Genes 2023, 14, 1472. https://doi.org/10.3390/genes14071472
Biltueva LS, Vorobieva NV, Lemskya NA, Perelman PL, Trifonov VA, Panov VV, Abramov AV, Kawada S-i, Serdukova NA, Graphodatsky AS. Chromosomal Evolution of the Talpinae. Genes. 2023; 14(7):1472. https://doi.org/10.3390/genes14071472
Chicago/Turabian StyleBiltueva, Larisa S., Nadezhda V. Vorobieva, Natalya A. Lemskya, Polina L. Perelman, Vladimir A. Trifonov, Victor V. Panov, Alexey V. Abramov, Shin-ichiro Kawada, Natalya A. Serdukova, and Alexandr S. Graphodatsky. 2023. "Chromosomal Evolution of the Talpinae" Genes 14, no. 7: 1472. https://doi.org/10.3390/genes14071472
APA StyleBiltueva, L. S., Vorobieva, N. V., Lemskya, N. A., Perelman, P. L., Trifonov, V. A., Panov, V. V., Abramov, A. V., Kawada, S.-i., Serdukova, N. A., & Graphodatsky, A. S. (2023). Chromosomal Evolution of the Talpinae. Genes, 14(7), 1472. https://doi.org/10.3390/genes14071472