
The First Complete Chloroplast Genome of Campanula carpatica: Genome Characterization and Phylogenetic Diversity

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Chloroplast Genome Sequencing
2.2. Chloroplast Genome Assembly and Annotation
2.3. Repeat Sequence
2.4. Comparison of Campanula Chloroplast Genomes
2.5. Phylogenetic Analyses
3. Results and Discussion
3.1. Chloroplast Genome Sequencing and Assembly
3.2. Repeat Sequence Analysis
3.3. Comparison of Chloroplast Genomes among Campanula Species
3.4. Contraction and Expansion of Border Regions
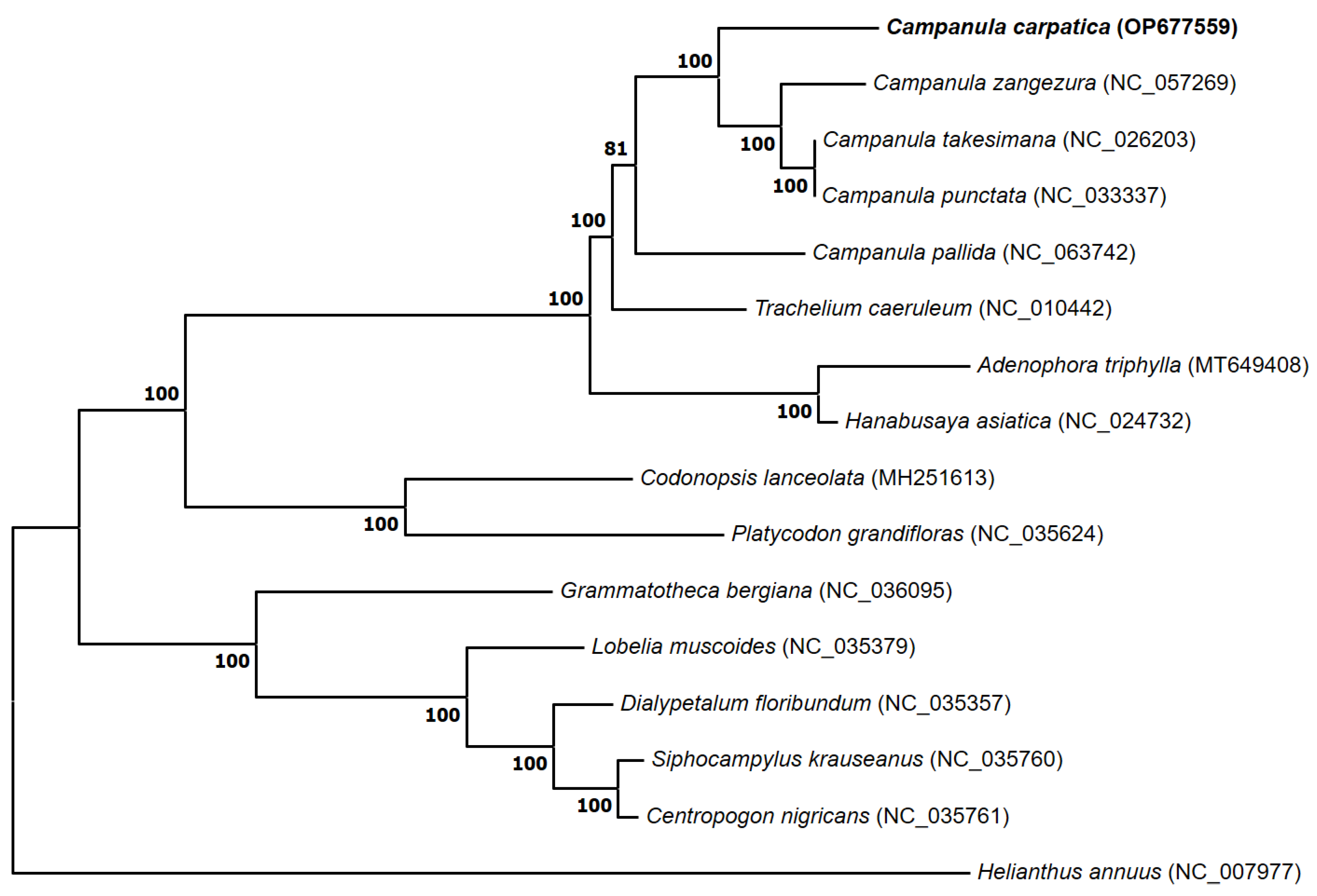
3.5. Phylogenetic Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sriskandarajah, S.; Mibus, H.; Serek, M. Transgenic Campanula carpatica plants with reduced ethylene sensitivity. Plant Cell Rep. 2007, 26, 805–813. [Google Scholar] [CrossRef]
- Frello, S.; Stummann, B.M.; Serek, M. Shoot regeneration of Campanula carpatica Jacq. (Campanulaceae) via callus phase. Sci. Hortic. 2002, 93, 85–90. [Google Scholar] [CrossRef]
- Mork, E.K.; Sriskandarajah, S.; Serek, M. Influence of seed germination conditions on regenerative ability in Campanula carpatica. Europ. J. Hortic. Sci. 2005, 70, 173–176. [Google Scholar]
- Sriskandarajah, S.; Frello, S.; Serek, M. Induction of adventitious shoots in vitro in Campanula carpatica. Plant Cell Tissue Organ. Cult. 2001, 67, 295–298. [Google Scholar] [CrossRef]
- Liveri, E.; Crowl, A.A.; Cellinese, N. Past, present, and future of Campanula (Campanulaceae) systematics—A review. Bot. Chron. 2019, 22, 209–222. [Google Scholar]
- Boutigny, A.L.; Dohin, N.; Pornin, D.; Polland, M. Overview and detectability of the genetic modifications in ornamental plants. Hortic. Res. 2020, 7, 11. [Google Scholar] [CrossRef]
- Song, F.; Li, T.; Burgess, K.S.; Feng, Y.; Ge, X.J. Complete plastome sequencing resolves taxonomic relationships among species of Calligonum L. (Polygonaceae) in China. BMC Plant Biol. 2020, 20, 261. [Google Scholar] [CrossRef]
- Daniell, H.; Jin, S.; Zhu, X.-G.; Gitzendanner, M.A.; Soltis, D.E.; Soltis, P.S. Green giant—A tiny chloroplast genome with mighty power to produce high-value proteins: History and phylogeny. Plant Biotechnol. J. 2021, 19, 430–447. [Google Scholar] [CrossRef]
- Tonti-Filippini, J.; Nevill, P.G.; Dixon, K.; Small, I. What Can We Do with 1000 Plastid Genomes? Plant J. 2017, 90, 808–818. [Google Scholar] [CrossRef] [Green Version]
- Avila, E.M.; Gisby, M.F.; Day, A. Seamless editing of the chloroplast genome in plants. BMC Plant Biol. 2016, 16, 168. [Google Scholar]
- Bendich, A.J. DNA abandonment and the mechanisms of uniparental inheritance of mitochondria and chloroplasts. Chromosome Res. 2013, 21, 287–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, W.; Duan, X.; Zhang, R.; Guo, C.; Li, L.; Xu, G.; Shan, H.; Kong, H.; Ren, Y. Chloroplast genomic data provide new and robust insights into the phylogeny and evolution of the Ranunculaceae. Mol. Phylogenet. Evol. 2019, 135, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.R. Mutation rates in plastid genomes: They are lower than you might think. Genome Biol. Evol. 2015, 7, 1227–1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobrogojski, J.; Adamiec, M.; Luciński, R. The chloroplast genome: A review. Acta Physiol. Plant. 2020, 42, 98. [Google Scholar] [CrossRef]
- Lei, W.; Ni, D.; Wang, Y.; Shao, J.; Wang, X.; Yang, D.; Wang, J.; Chen, H.; Liu, C. Intraspecific and heteroplasmic variations, gene losses and inversions in the chloroplast genome of Astragalus membranaceus. Sci. Rep. 2016, 6, 21669. [Google Scholar] [CrossRef] [Green Version]
- Henriquez, C.L.; Abdullah; Ahmed, I.A.; Carlsen, M.M.; Zuluaga, A.; Croat, T.B.; Mckain, M.R. Molecular evolution of chloroplast genomes in Monsteroideae (Araceae). Planta 2020, 251, 72. [Google Scholar] [CrossRef]
- Civáň, P.; Foster, P.G.; Embley, M.T.; Séneca, A.; Cox, C.J. Analyses of charophyte chloroplast genomes help characterize the ancestral chloroplast genome of land plants. Genome Biol. Evol. 2014, 6, 897–911. [Google Scholar] [CrossRef] [Green Version]
- Koren, S.; Walenz, B.P.; Berlin, K.; Miller, J.R.; Bergman, N.H.; Phillippy, A.M. Canu: Scalable and accurate long-read assembly via adaptive k-mer weighting and repeat separation. Genome Res. 2017, 27, 722–736. [Google Scholar] [CrossRef] [Green Version]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef]
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq—Versatile and accurate annotation of organelle genomes. Nucleic Acids Res. 2017, 45, W6–W11. [Google Scholar] [CrossRef] [Green Version]
- Rutherford, K.; Parkhill, J.; Crook, J.; Horsnell, T.; Rice, P.; Rajandream, M.A.; Barrell, B. Artemis: Sequence visualization and annotation. Bioinformatics 2000, 16, 944–945. [Google Scholar] [CrossRef] [Green Version]
- Lohse, M.; Drechsel, O.; Bock, R. OrganellarGenomeDRAW (OGDRAW): A tool for the easy generation of high-quality custom graphical maps of plastid and mitochondrial genomes. Curr. Genet. 2007, 52, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Ni, Y.; Li, J.; Zhang, X.; Yang, H.; Chen, H.; Liu, C. CPGView: A package for visualizing detailed chloroplast genome structures. Mol. Ecol. Resour. 2023, 23, 694–704. [Google Scholar] [CrossRef] [PubMed]
- Frazer, K.A.; Pachter, L.; Poliakov, A.; Rubin, E.M.; Dubchak, I. VISTA: Computational tools for comparative genomics. Nucleic Acids Res. 2004, 32, W273–W279. [Google Scholar] [CrossRef] [Green Version]
- Darling, A.C.E.; Mau, B.; Blattner, F.R.; Perna, N.T. Mauve: Multiple alignment of conserved genomic sequence with rearrangements. Genome Res. 2004, 14, 1394–1403. [Google Scholar] [CrossRef] [Green Version]
- Amiryousefi, A.; Hyvönen, J.; Poczai, P. Irscope: An online program to visualize the junction sites of chloroplast genomes. Bioinformatics 2018, 34, 3030–3031. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Jansen, R.K.; Raubeson, L.A.; Boore, J.L.; Depamphilis, C.W.; Chumley, T.W.; Haberle, R.C.; Wyman, S.K.; Alverson, A.J.; Peery, R.; Herman, S.J.; et al. Methods for obtaining and analyzing whole chloroplast genome sequences. Methods Enzymol. 2005, 395, 348–384. [Google Scholar]
- Knox, E.B. The dynamic history of plastid genomes in the Campanulaceae sensu lato is unique among angiosperms. Proc. Natl. Acad. Sci. USA. 2014, 111, 11097–11102. [Google Scholar] [CrossRef]
- Weng, M.L.; Blazier, J.C.; Govindu, M.; Jansen, R.K. Reconstruction of the ancestral plastid genome in Geraniaceae reveals a correlation between genome rearrangements, repeats, and nucleotide substitution rates. Mol. Biol. Evol. 2014, 31, 645–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taheri, S.; Lee Abdullah, T.; Yusop, M.R.; Hanafi, M.M.; Sahebi, M.; Azizi, P.; Shamshiri, R.R. Mining and development of novel SSR markers using next generation sequencing (NGS) data in plants. Molecules 2018, 23, 399. [Google Scholar] [CrossRef] [Green Version]
- Ledenyova, M.L.; Tkachenko, G.A.; Shpak, I.M. Imperfect and compound microsatellites in the genomes of Burkholderia pseudomallei strains. Mol. Biol. 2019, 53, 127–137. [Google Scholar] [CrossRef]
- Provan, J.; Powell, W.; Hollingsworth, P.M. Chloroplast microsatellites: New tools for studies in plant ecology and evolution. Trends Ecol. Evol. 2001, 16, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Nie, X.; Lv, S.; Zhang, Y.; Du, X.; Wang, L.; Biradar, S.S.; Tan, X.; Wan, F.; Weining, S. Complete chloroplast genome sequence of a major invasive species, Crofton weed (Ageratina adenophora). PLoS ONE 2012, 7, e36869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tangphatsornruang, S.; Uthaipaisanwong, P.; Sangsrakru, D.; Chanprasert, J.; Yoocha, T.; Jomchai, N.; Tragoonrung, S. Characterization of the complete chloroplast genome of Hevea brasiliensis reveals genome rearrangement, RNA editing sites and phylogenetic relationships. Gene 2011, 475, 104–112. [Google Scholar] [CrossRef]
- Timme, R.E.; Kuehl, J.V.; Boore, J.L.; Jansen, R.K. A comparative analysis of the Lactuca and Helianthus (Asteraceae) plastid genomes: Identification of divergent regions and categorization of shared repeats. Am. J. Bot. 2007, 94, 302–312. [Google Scholar] [CrossRef] [PubMed]
- Claude, S.J.; Park, S.; Park, S. Gene loss, genome rearrangement, and accelerated substitution rates in plastid genome of Hypericum ascyron (Hypericaceae). BMC Plant Biol. 2022, 22, 135. [Google Scholar] [CrossRef]
- Wang, L.; He, N.; Li, Y.; Fang, Y.; Zhang, F. Complete chloroplast genome sequence of Chinese lacquer tree (Toxicodendron vernicifluum, Anacardiaceae) and its phylogenetic significance. BioMed Res. Int. 2020, 1, 9014873. [Google Scholar] [CrossRef] [Green Version]
- Daniell, H.; Lin, C.S.; Yu, M.; Chang, W.J. Chloroplast genomes: Diversity, evolution, and applications in genetic engineering. Genome Biol. 2016, 17, 134. [Google Scholar] [CrossRef] [Green Version]
- Shi, W.; Song, W.; Chen, Z.; Cai, H.; Gong, Q.; Liu, J.; Shi, C.; Wang, S. Comparative chloroplast genome analyses of diverse Phoebe (Lauraceae) species endemic to China provide insight into their phylogeographical origin. PeerJ 2023, 11, e14573. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Wang, Y.; Liu, Y.; Xu, C.; Yuan, Q.; Guo, L.; Huang, L. Evolutionary and phylogenetic aspects of the chloroplast genome of Chaenomeles species. Sci. Rep. 2020, 10, 11466. [Google Scholar] [CrossRef] [PubMed]
- Lian, C.; Yang, H.; Lan, J.; Zhang, X.; Zhang, F.; Yang, J.; Chen, S. Comparative analysis of chloroplast genomes reveals phylogenetic relationships and intraspecific variation in the medicinal plant Isodon rubescens. PLoS ONE 2022, 17, e0266546. [Google Scholar] [CrossRef] [PubMed]
- Cheong, W.Y.; Kim, S.J.; Yang, J.; Lee, W.; Pak, J.H.; Kim, S.C. Insights from the chloroplast DNA into the progenitor-derivative relationship between Campanula punctata and C. takesimana (Campanulaceae) in Korea. J. Plant Biol. 2020, 63, 431–444. [Google Scholar] [CrossRef]
- Kim, K.A.; Cheon, K.S. Complete chloroplast genome sequence of Adenophora racemosa (Campanulaceae): Comparative analysis with congeneric species. PLoS ONE 2021, 16, e0248788. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Group of Genes | Name of Genes |
|---|---|---|
| Self-replication | Ribosome (LSU 1) | rpl2, rpl14, rpl16, rpl20, rpl32, rpl33, rpl36 |
| Ribosome (SSU 2) | rps2, rps3, rps4, rps7, rps8, rps11, rps12 *, rps14, rps16, rps18, rps19 | |
| RNA polymerase | rpoA, rpoB, rpoC1, rpoC2 | |
| rRNA genes | rrn4.5S *, rrn5S *, rrn16S3 *, rrn23S * | |
| tRNA genes | trnH-GUG, trnK-UUU, trnQ-UUG *, trnS-GCU, trnS-CGA, trnR-UCU, trnV-GAC, trnP-UGG, trnW-CCA, trnT-UGU, trnL-UAA, trnD-GUC, trnC-GCA, trnF-GAA, trnG-GCC, trnS-UGA, trnE-UUC, trnY-GUA, trnS-GGA, trnL-CAA *, trnA-UGC *, trnR-ACG *, trnI *, trnN-GUU *, trnV-UAC, trnL-UAG, trnM-CAU ** | |
| Photosynthesis | Photosystem I | psaA, psaB, psaC, psaJ |
| Photosystem II | psbA, psbB, psbC, psbD, psbE, psbF, psbH, psbI, psbJ *, psbK, psbM, psbN, psbT, psbZ, ycf3 | |
| Cytochrome 3 | petA, petB, petD, petG, petL, petN | |
| ATP synthase | atpA, atpB, atpE, atpF, atpH, atpI | |
| NADH 4 | ndhA *, ndhB, ndhC, ndhD, ndhE *, ndhF, ndhG *, ndhH *, ndhI *, ndhJ, ndhK | |
| Subunit of rubisco | rbcL | |
| Other genes | c-type cytochrom 5 | ccsA, |
| Envelop membrane protein | cemA, | |
| Maturase | matK | |
| Conserved 6 | ycf4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoon, W.-S.; Kim, C.-K.; Kim, Y.-K. The First Complete Chloroplast Genome of Campanula carpatica: Genome Characterization and Phylogenetic Diversity. Genes 2023, 14, 1597. https://doi.org/10.3390/genes14081597
Yoon W-S, Kim C-K, Kim Y-K. The First Complete Chloroplast Genome of Campanula carpatica: Genome Characterization and Phylogenetic Diversity. Genes. 2023; 14(8):1597. https://doi.org/10.3390/genes14081597
Chicago/Turabian StyleYoon, Won-Sub, Chang-Kug Kim, and Yong-Kab Kim. 2023. "The First Complete Chloroplast Genome of Campanula carpatica: Genome Characterization and Phylogenetic Diversity" Genes 14, no. 8: 1597. https://doi.org/10.3390/genes14081597
APA StyleYoon, W.-S., Kim, C.-K., & Kim, Y.-K. (2023). The First Complete Chloroplast Genome of Campanula carpatica: Genome Characterization and Phylogenetic Diversity. Genes, 14(8), 1597. https://doi.org/10.3390/genes14081597