
Combined Analysis of Untargeted Metabolomics and Transcriptomics Revealed Seed Germination and Seedling Establishment in Zelkova schneideriana

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
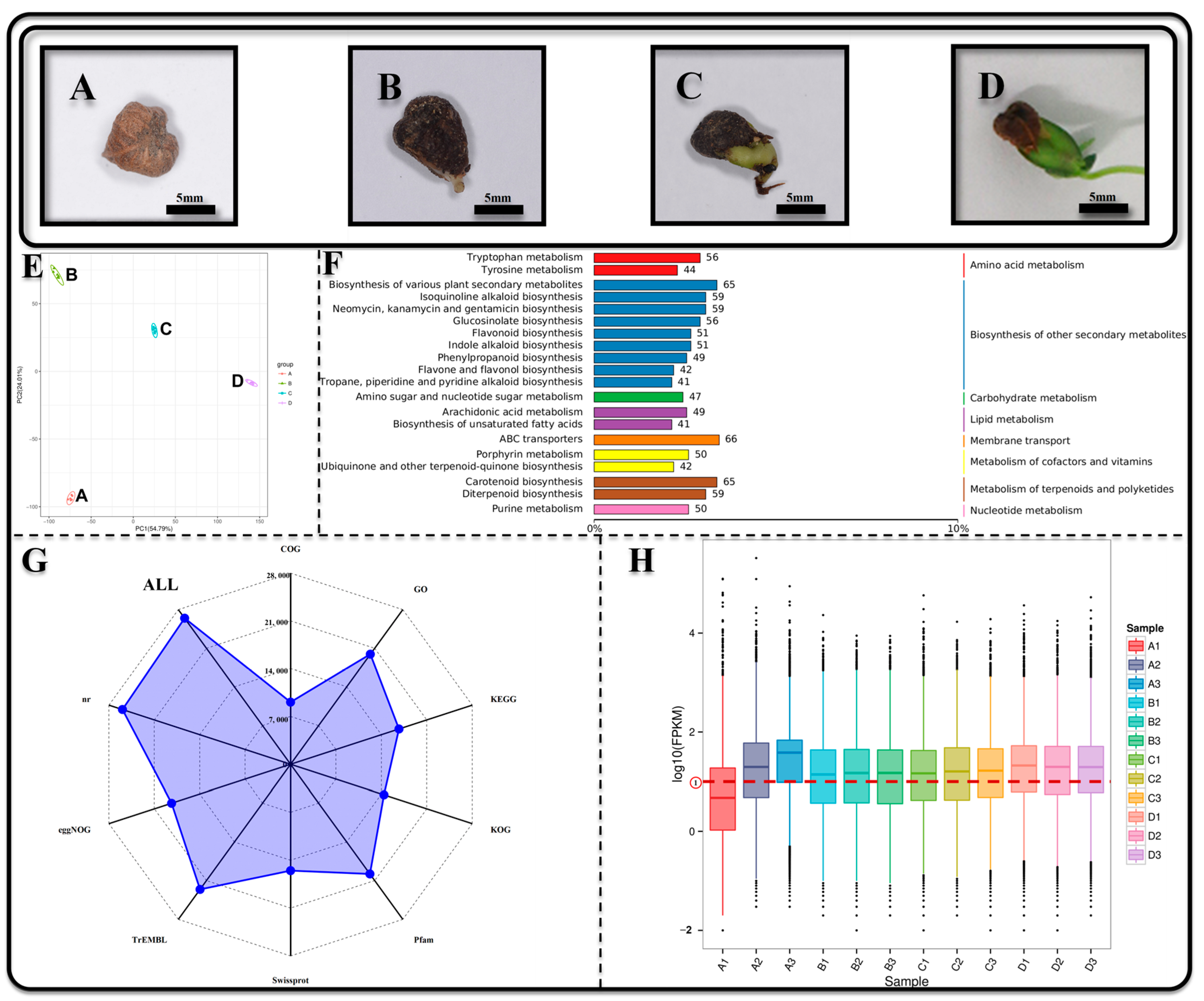
2.1. Plant Materials
2.2. Sample Metabolite Extraction
2.3. LC-MS Analysis
2.4. Data Preprocessing and Annotation and Metabolite Data Analysis
2.5. Transcriptome Sequencing and De Novo Assembly
2.6. Functional Annotation and Differential Expression Analysis of Differentially Expressed Genes (DEGs)
2.7. Validation of Gene Expression Profiles through RT-qPCR
2.8. Statistical Analysis
3. Results
3.1. Transcriptome and Metabolome Functional Annotation and Expression Profiling

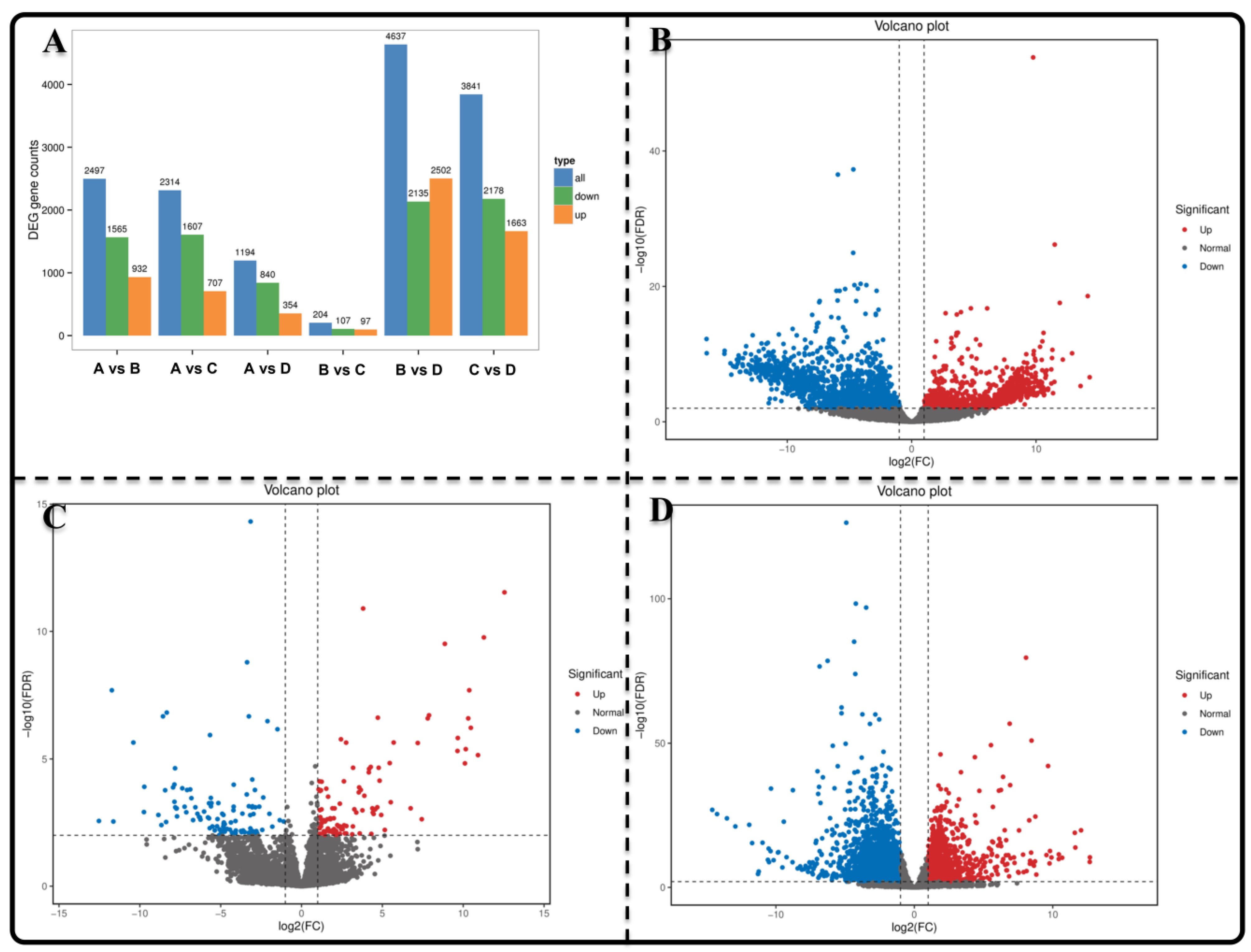
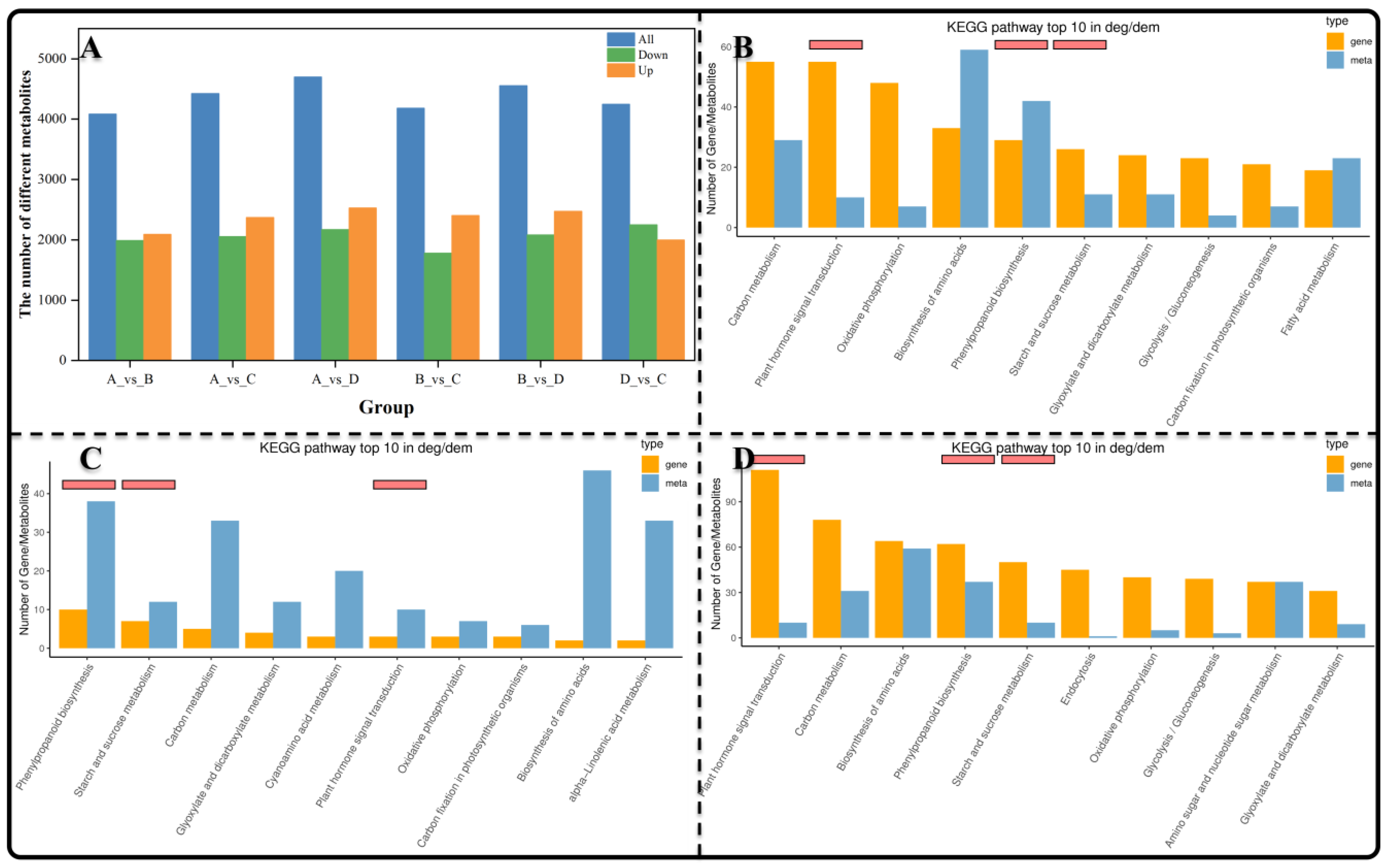
3.2. The Identification of Differentially Express Genes
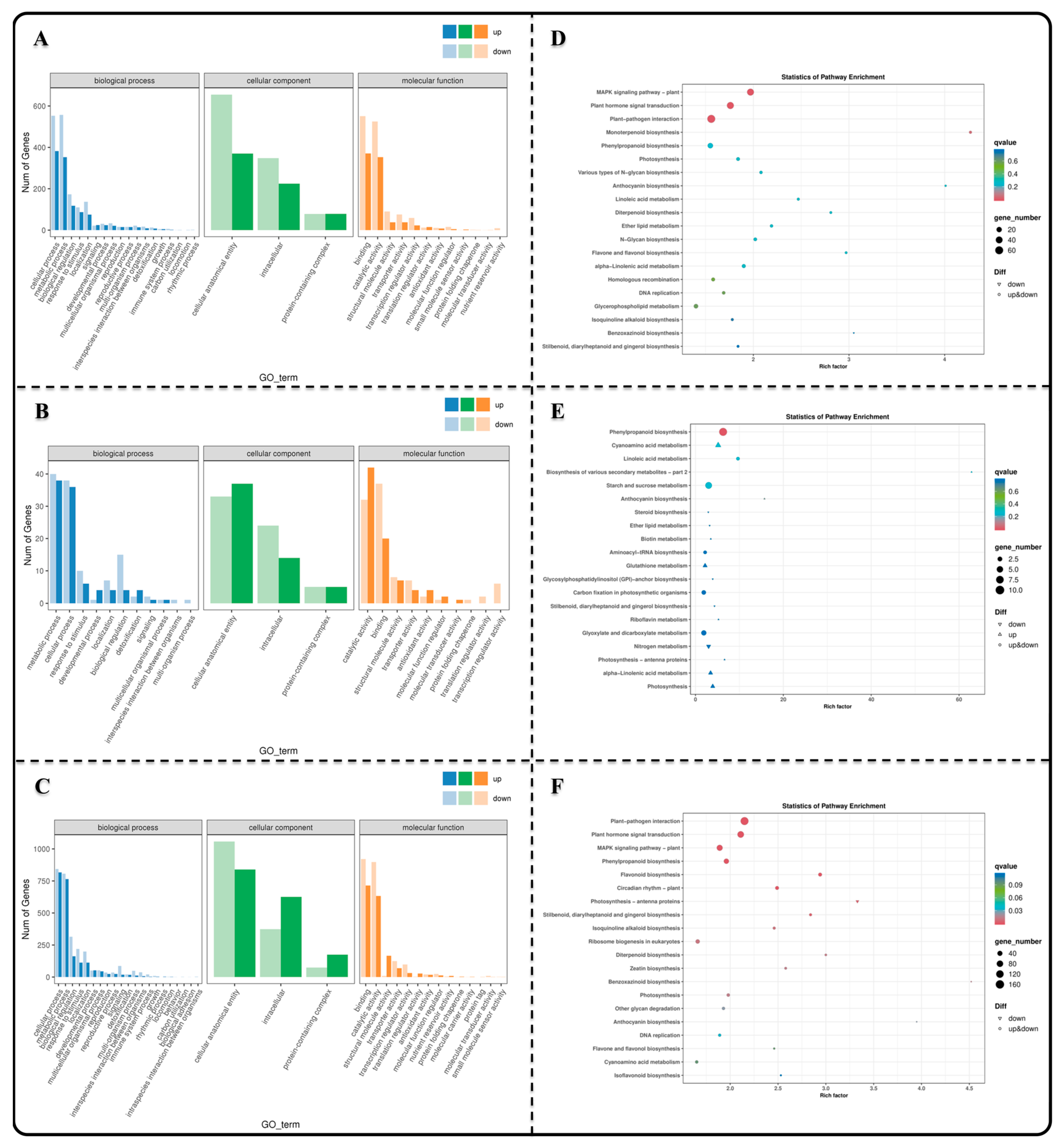
3.3. GO and KEGG Enrichment Analysis of Differentially Expressed Genes
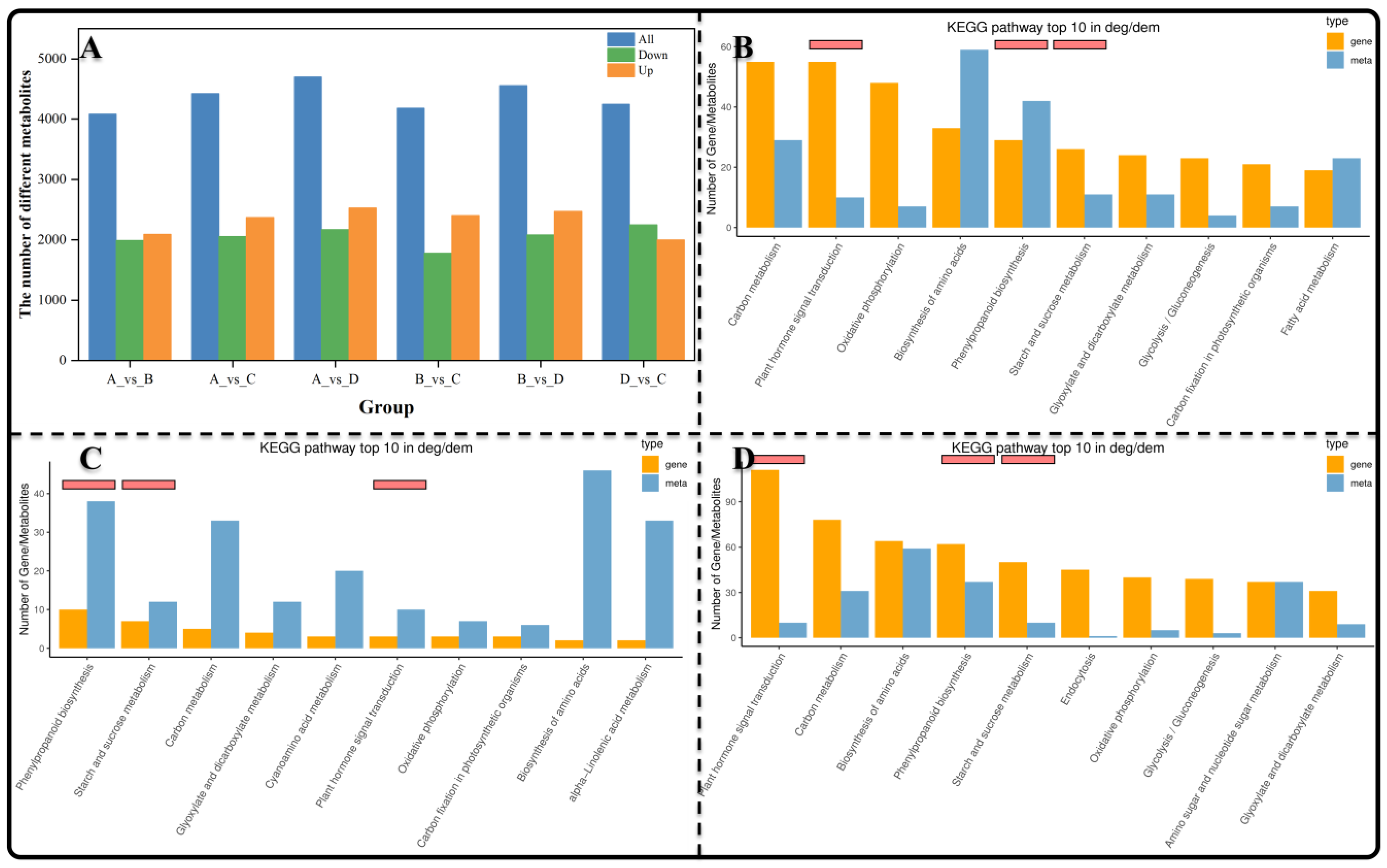
3.4. The Identification and KEGG Enrichment Analysis of Differentially Expressed Metabolites (DEMs)
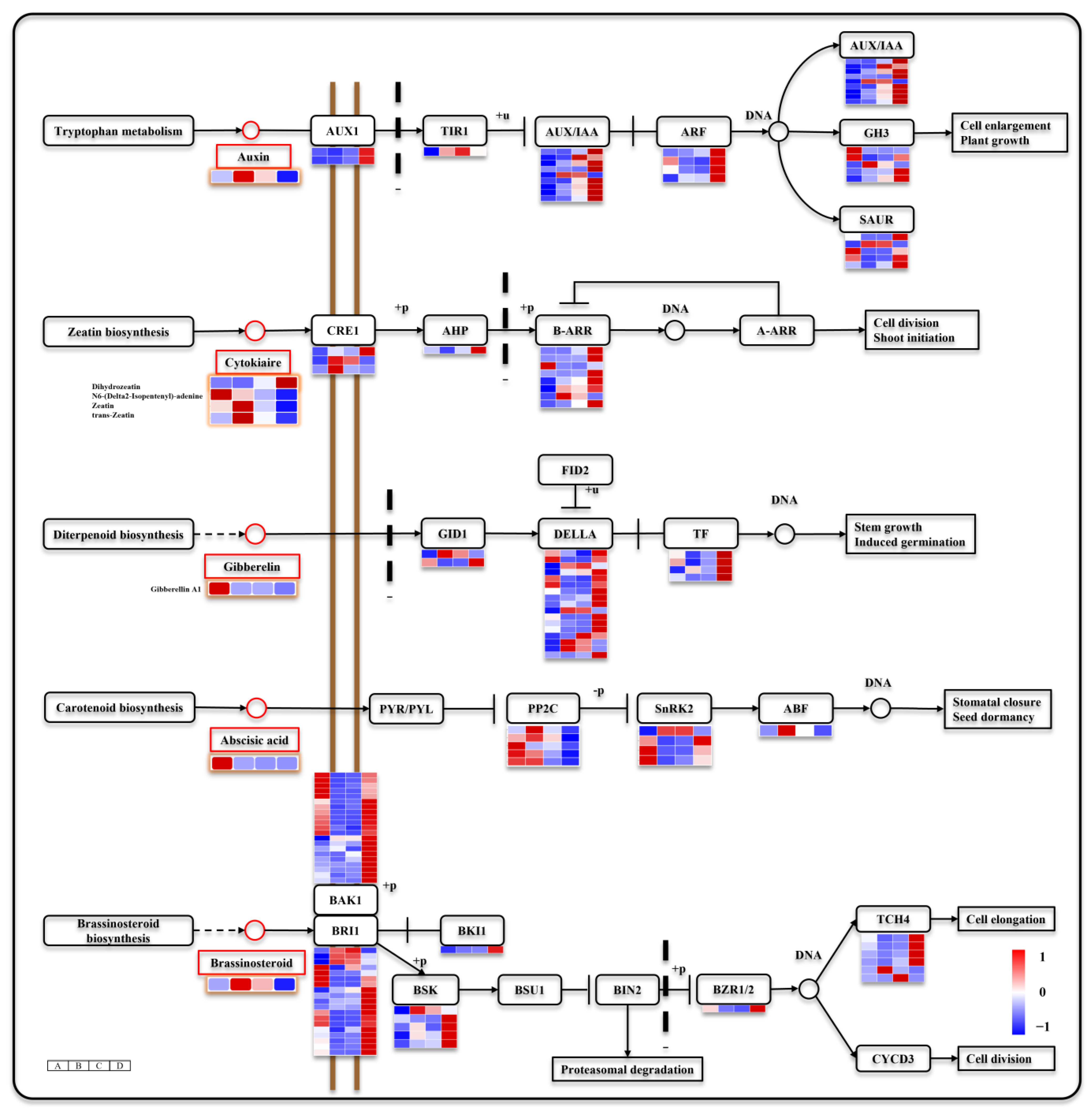
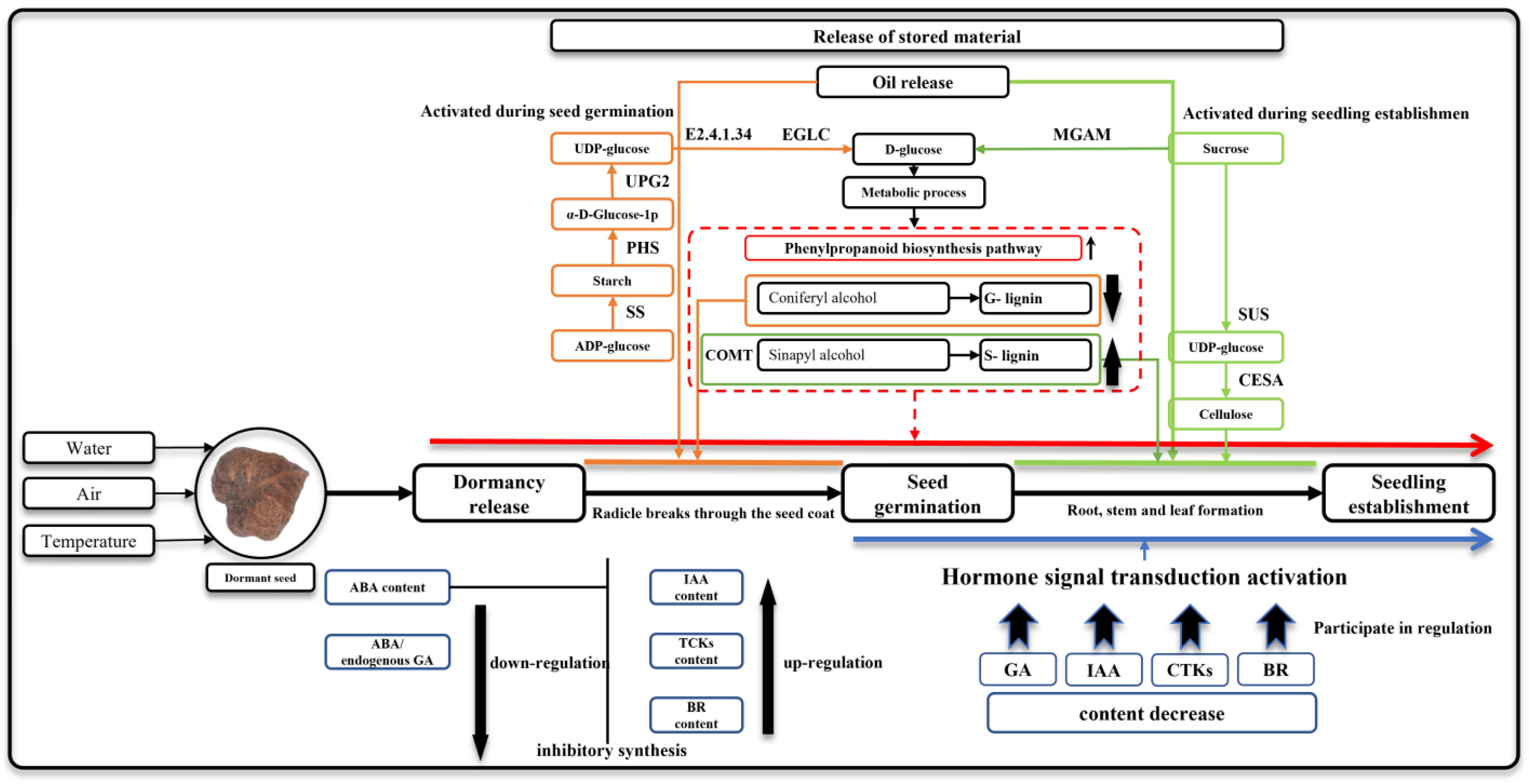
3.5. Seed Germination and Seeding Establishment Are Regulated by Hormone Signals
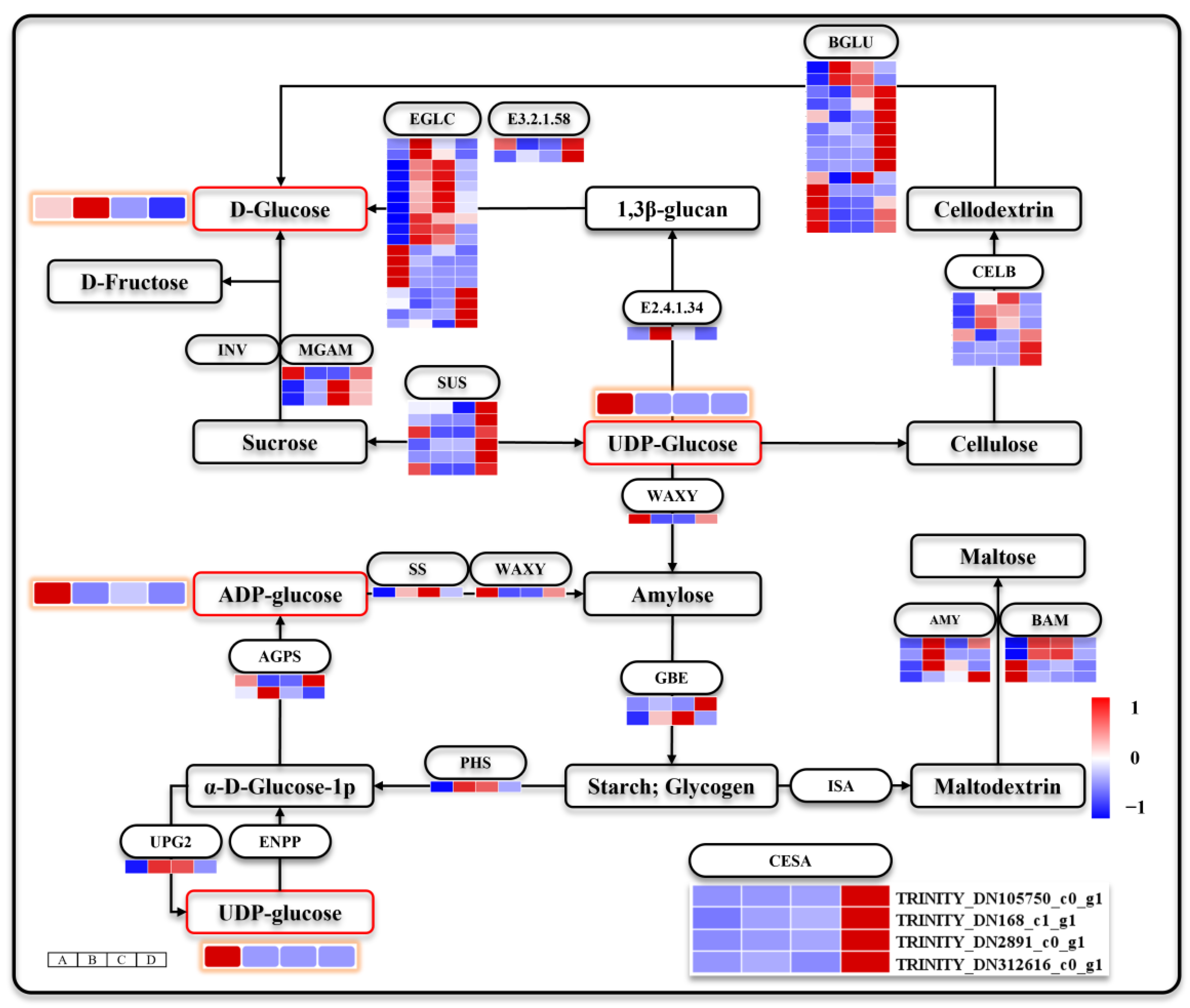
3.6. Releasing of Stored Carbohydrates during Seed Germination and Seeding Establishment
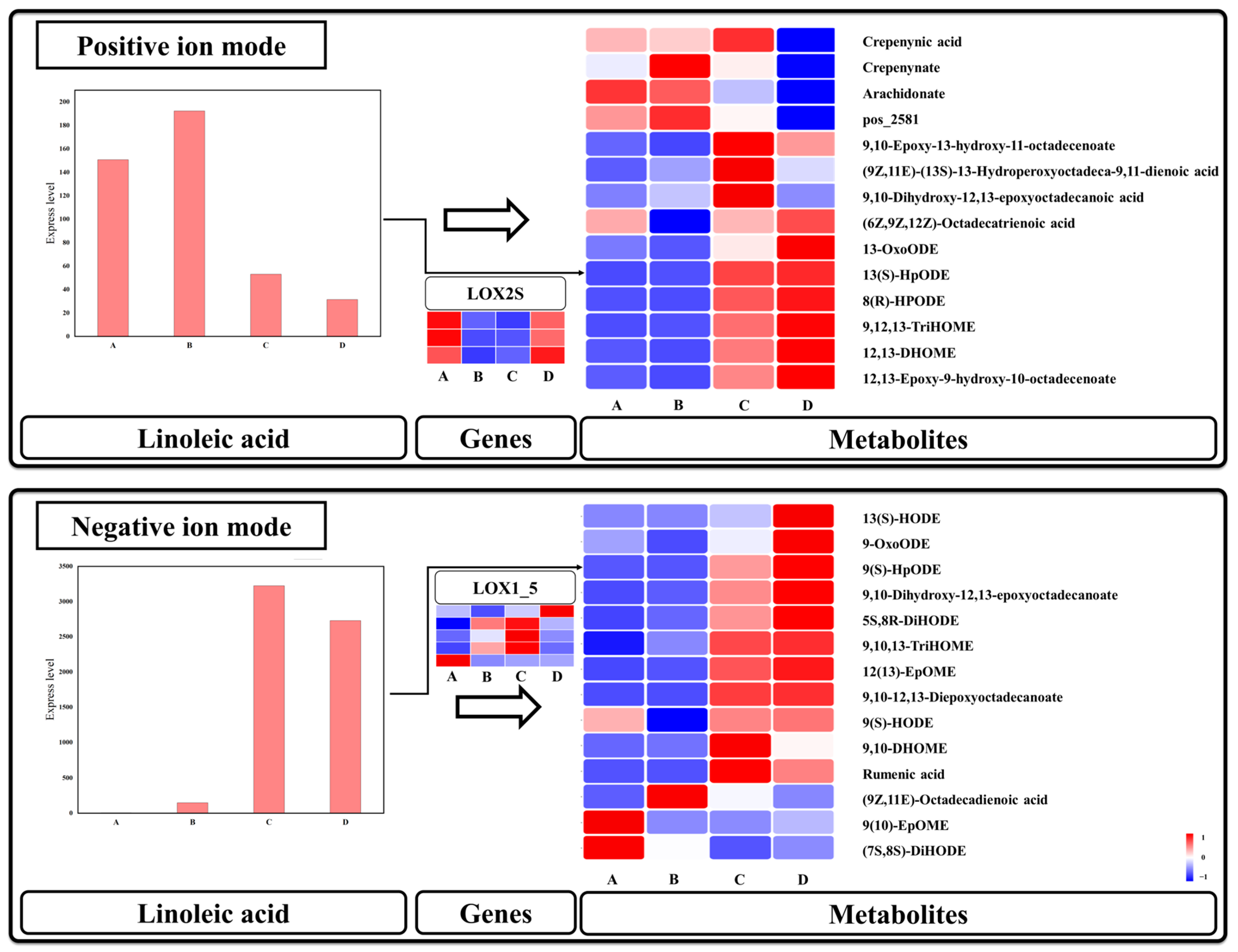
3.7. Linoleic Acid Metabolism Is Mainly Activated after Seed Germination

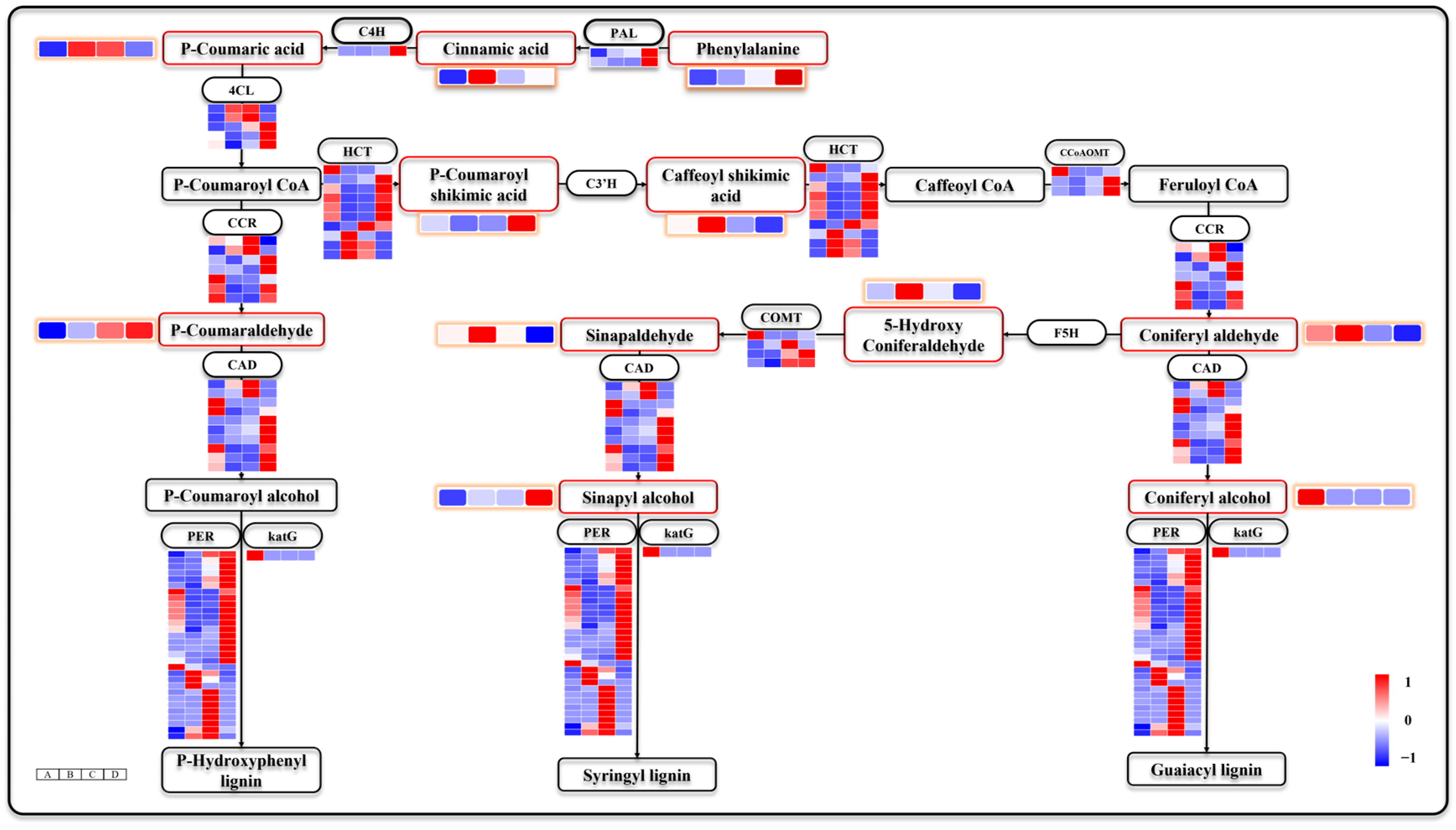
3.8. Phenylpropane Biosynthesis Is also Involved in Seed Germination and Seeding Establishment

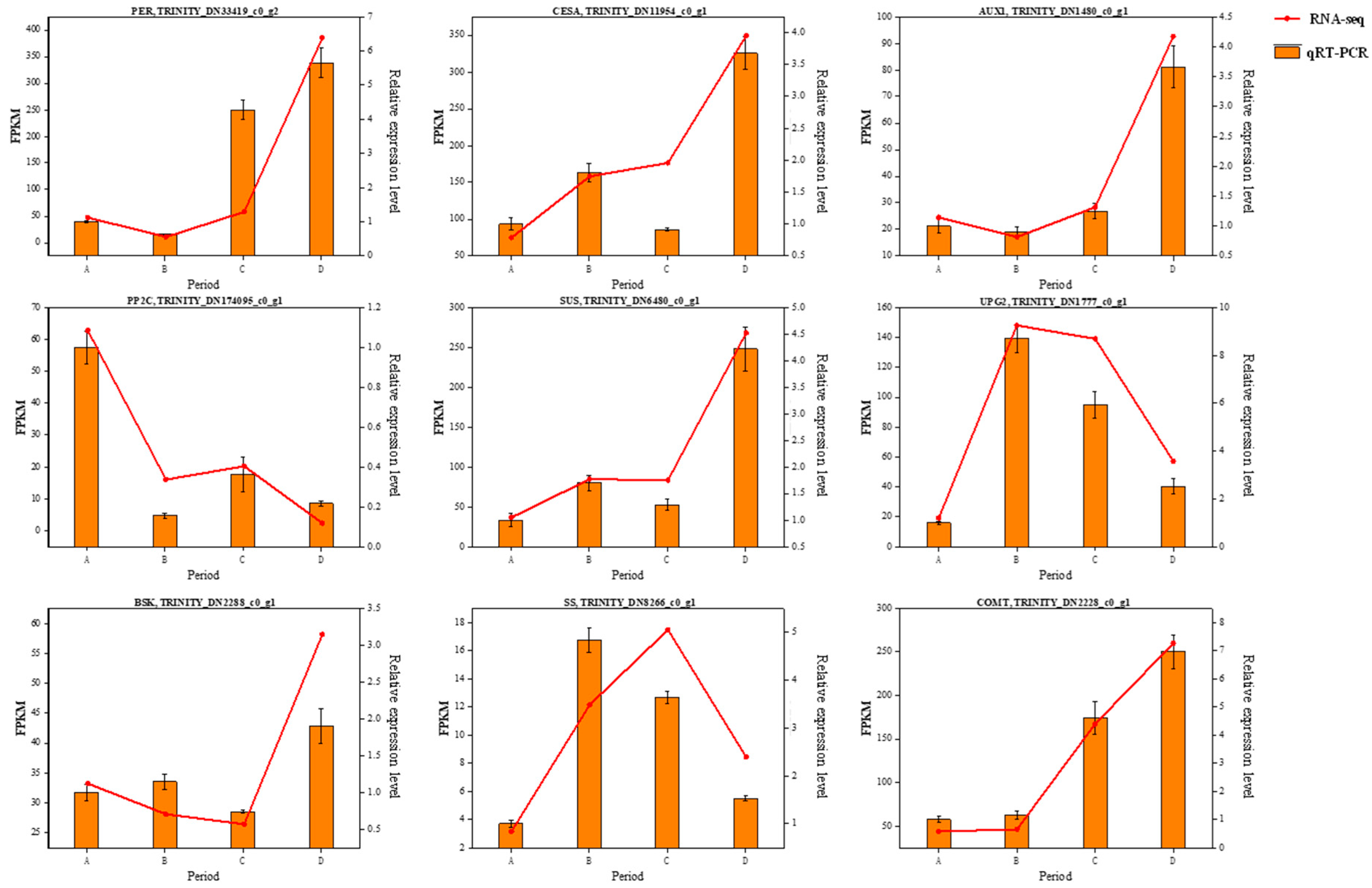
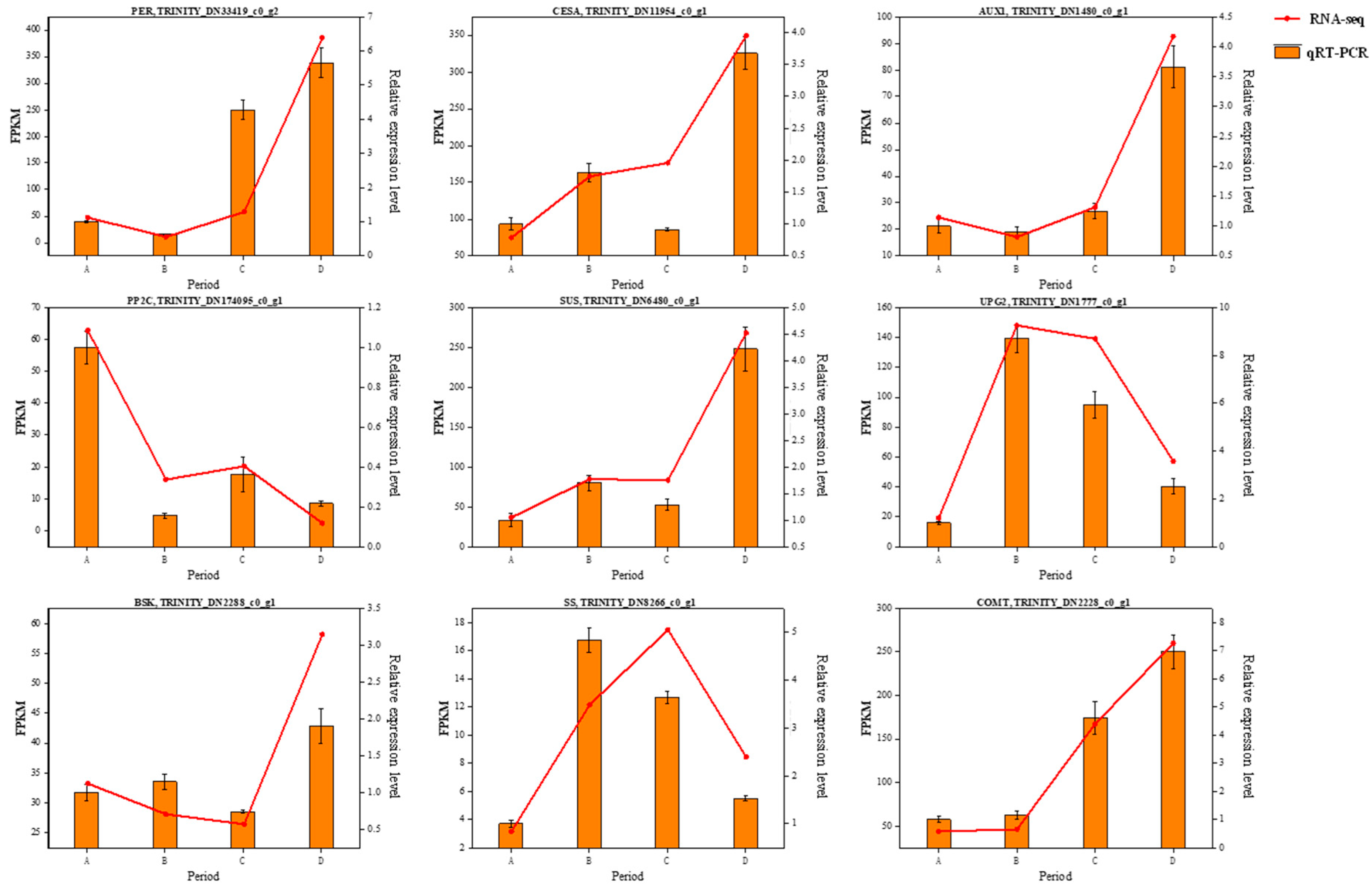
3.9. Quantitative Real-Time PCR (RT-qPCR) Validation
4. Discussion
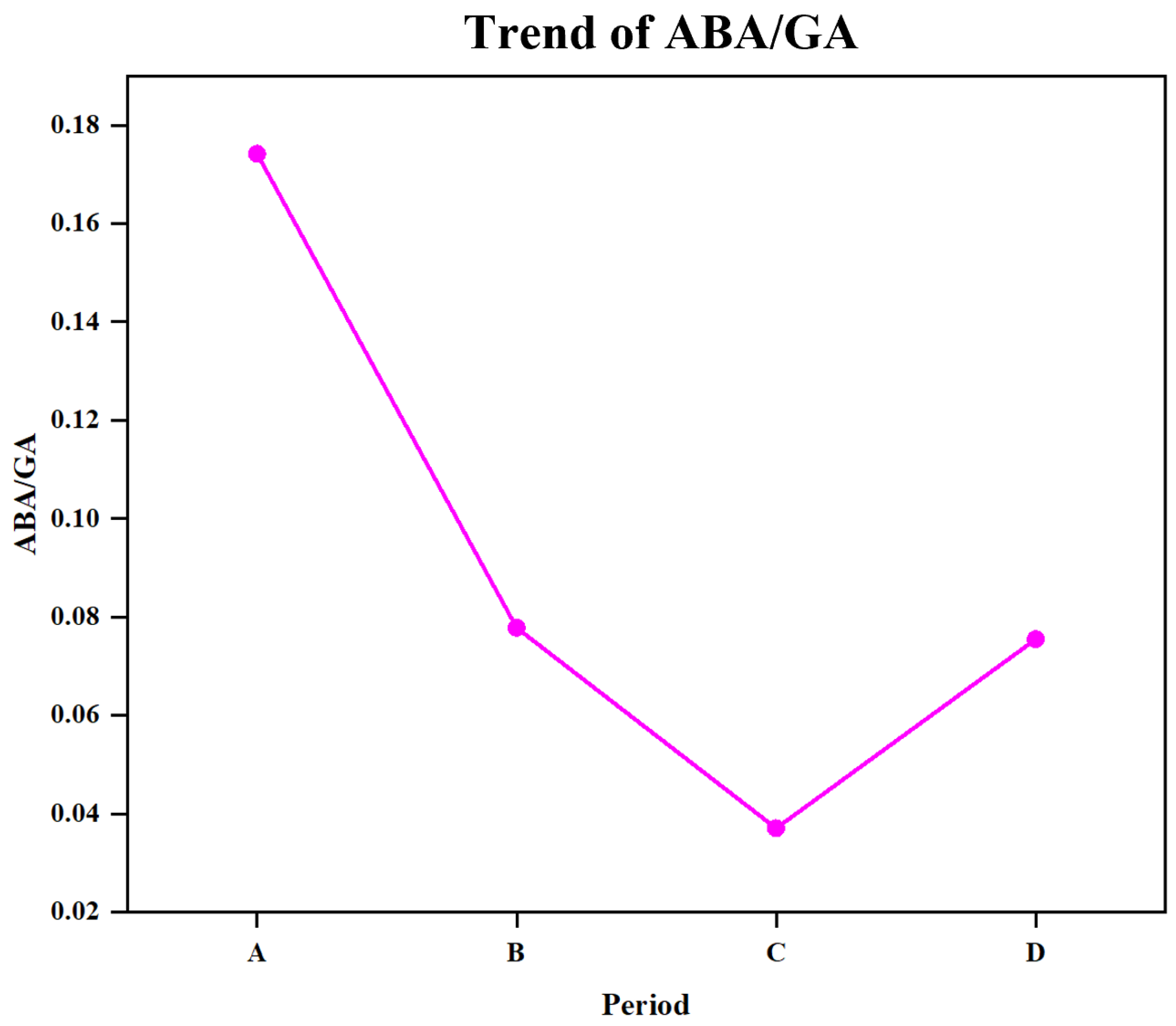
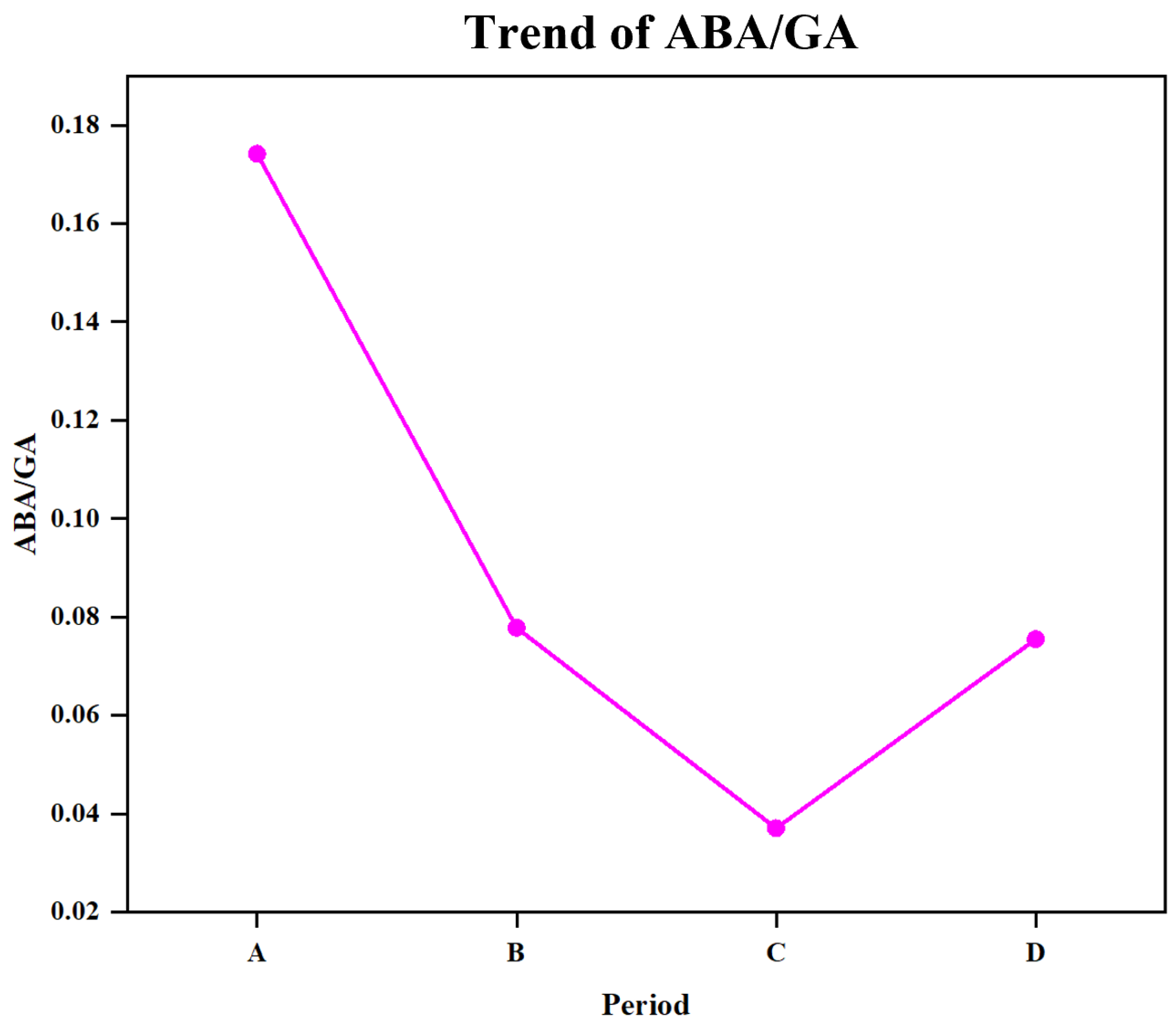
4.1. ABA and GA Are Key Hormones Involved in Seed Germination and Affect the Synthesis of Other Hormones
4.2. Seed Germination and Seeding Establishment Have Different Characteristics of Sugar Metabolism
4.3. Lipids Release May Serve as a Potential Supplement in the Case of Sugar Deficiency
4.4. The Formation of G-Lignin and S-Lignin Contributes Differently to Seed Germination and Seeding Establishment
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rubio de Casas, R.; Willis, C.G.; Pearse, W.D.; Baskin, C.C.; Baskin, J.M.; Cavender-Bares, J. Global Biogeography of Seed Dormancy Is Determined by Seasonality and Seed Size: A Case Study in the Legumes. New Phytol. 2017, 214, 1527–1536. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Yang, P. Studies on the Molecular Mechanisms of Seed Germination. Proteomics 2015, 15, 1671–1679. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Mahmood, A.; Römheld, V.; Neumann, G. Nutrient Seed Priming Improves Seedling Development of Maize Exposed to Low Root Zone Temperatures during Early Growth. Eur. J. Agron. 2013, 49, 141–148. [Google Scholar] [CrossRef]
- Chen, M.; Thelen, J.J. The Plastid Isoform of Triose Phosphate Isomerase Is Required for the Postgerminative Transition from Heterotrophic to Autotrophic Growth in Arabidopsis. Plant Cell 2010, 22, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Molina, L.; Mongrand, S.; Chua, N.-H. A Postgermination Developmental Arrest Checkpoint Is Mediated by Abscisic Acid and Requires the ABI5 Transcription Factor in Arabidopsis. Proc. Natl. Acad. Sci. USA 2001, 98, 4782–4787. [Google Scholar] [CrossRef] [PubMed]
- Hooley, R. Gibberellins: Perception, Transduction and Responses. In Signals and Signal Transduction Pathways in Plants; Springer: Berlin/Heidelberg, Germany, 1994; pp. 1529–1555. [Google Scholar] [CrossRef]
- Graeber, K.; Nakabayashi, K.; Miatton, E.; Leubner-Metzger, G.; Soppe, W.J.J. Molecular Mechanisms of Seed Dormancy. Plant Cell Environ. 2012, 1769–1786. [Google Scholar] [CrossRef]
- Miransari, M.; Smith, D.L. Plant Hormones and Seed Germination. Environ. Exp. Bot. 2014, 99, 110–121. [Google Scholar] [CrossRef]
- Guan, S.X.; Li, Y.S.; Zeng, W.Q.; Lin, J.W.; Zhan, H.; Han, X.Y.; Zhang, X.L.; Lu, X.J. A gibberellin, abscisic acid, and DELAY OF GERMINATION 1 interaction network regulates critical developmental transitions in model plant Arabidopsis thaliana—A review. Appl. Ecol. Environ. Res. 2021, 19, 4699–4720. [Google Scholar] [CrossRef]
- Nelson, S.K.; Kanno, Y.; Seo, M.; Steber, C.M. Seed Dormancy Loss from Dry After-Ripening Is Associated with Increasing Gibberellin Hormone Levels in Arabidopsis thaliana. Front. Plant Sci. 2023, 14, 1145414. [Google Scholar] [CrossRef]
- Bialek, K.; Michalczuk, L.; Cohen, J.D. Auxin Biosynthesis during Seed Germination in Phaseolus vulgaris. Plant Physiol. 1992, 100, 509–517. [Google Scholar] [CrossRef]
- Hentrich, M.; Böttcher, C.; Düchting, P.; Cheng, Y.; Zhao, Y.; Berkowitz, O.; Masle, J.; Medina, J.; Pollmann, S. The Jasmonic Acid Signaling Pathway Is Linked to Auxin Homeostasis through the Modulation of YUCCA8 and YUCCA9 Gene Expression. Plant J. 2013, 74, 626–637. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.W.; Chen, J.J.W.; Wu, C.S.; Chang, H.C.; Chen, H.Y.; Kuo, H.H.; Lee, Y.S.; Chang, Y.L.; Chang, H.C.; Shiuea, S.Y.; et al. Auxin Plays a Role in the Adaptation of Rice to Anaerobic Germination and Seedling Establishment. Plant Cell Environ. 2023, 46, 1157–1175. [Google Scholar] [CrossRef] [PubMed]
- Finkelstein, R.; Reeves, W.; Ariizumi, T.; Steber, C. Molecular Aspects of Seed Dormancy. Annu. Rev. Plant Biol. 2008, 59, 387–415. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Cai, Z.; Wang, X. The Primary Signaling Outputs of Brassinosteroids Are Regulated by Abscisic Acid Signaling. Proc. Natl. Acad. Sci. USA 2009, 106, 4543–4548. [Google Scholar] [CrossRef] [PubMed]
- Depuydt, S.; Hardtke, C.S. Hormone Signalling Crosstalk in Plant Growth Regulation. Curr. Biol. 2011, 21, R365–R373. [Google Scholar] [CrossRef] [PubMed]
- Chiwocha, S.D.S.; Cutler, A.J.; Abrams, S.R.; Ambrose, S.J.; Yang, J.; Ross, A.R.S.; Kermode, A.R. The Etr1-2 Mutation in Arabidopsis thaliana Affects the Abscisic Acid, Auxin, Cytokinin and Gibberellin Metabolic Pathways during Maintenance of Seed Dormancy, Moist-Chilling and Germination. Plant J. 2005, 42, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Nikolić, R.; Mitić, N.; Miletić, R.; Nešković, M. Effects of Cytokinins on In Vitro Seed Germination and Early Seedling Morphogenesis in Lotus corniculatus L. J. Plant Growth Regul. 2006, 25, 187–194. [Google Scholar] [CrossRef]
- Galland, M.; He, D.; Lounifi, I.; Arc, E.; Clément, G.; Balzergue, S.; Huguet, S.; Cueff, G.; Godin, B.; Collet, B.; et al. An Integrated “Multi-Omics” Comparison of Embryo and Endosperm Tissue-Specific Features and Their Impact on Rice Seed Quality. Front. Plant Sci. 2017, 8, 1984. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Li, X.; Wang, X.; Chen, H.; Chen, F.; Shen, S. Proteomic Analysis of Rice (Oryza sativa) Seeds during Germination. Proteomics 2007, 7, 3358–3368. [Google Scholar] [CrossRef]
- Sreenivasulu, N.; Usadel, B.; Winter, A.; Radchuk, V.; Scholz, U.; Stein, N.; Weschke, W.; Strickert, M.; Close, T.J.; Stitt, M.; et al. Barley Grain Maturation and Germination: Metabolic Pathway and Regulatory Network Commonalities and Differences Highlighted by New MapMan/PageMan Profiling Tools. Plant Physiol. 2008, 146, 1738–1758. [Google Scholar] [CrossRef]
- Soriano, D.; Huante, P.; Gamboa-deBuen, A.; Orozco-Segovia, A. Effects of Burial and Storage on Germination and Seed Reserves of 18 Tree Species in a Tropical Deciduous Forest in Mexico. Oecologia 2014, 174, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Sohn, S.-I.; Pandian, S.; Kumar, T.S.; Zoclanclounon, Y.A.B.; Muthuramalingam, P.; Shilpha, J.; Satish, L.; Ramesh, M. Seed Dormancy and Pre-Harvest Sprouting in Rice—An Updated Overview. Int. J. Mol. Sci. 2021, 22, 11804. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Tian, T.; Chen, J.; Wang, D.; Tong, B.; Liu, J. Transcriptome Analysis of Cinnamomum migao Seed Germination in Medicinal Plants of Southwest China. BMC Plant Biol. 2021, 21, 270. [Google Scholar] [CrossRef] [PubMed]
- MacGregor, D.R.; Kendall, S.L.; Florance, H.; Fedi, F.; Moore, K.; Paszkiewicz, K.; Smirnoff, N.; Penfield, S. Seed Production Temperature Regulation of Primary Dormancy Occurs through Control of Seed Coat Phenylpropanoid Metabolism. New Phytol. 2015, 205, 642–652. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.; Li, J.; Wang, X.; Zhao, X.; Yang, X.; Tang, Q.; He, G.; Zhou, G.; Kong, Y. Xylan Synthesized byIrregular Xylem 14(IRX14) Maintains the Structure of Seed Coat Mucilage in Arabidopsis. J. Exp. Bot. 2016, 67, 1243–1257. [Google Scholar] [CrossRef] [PubMed]
- Nonogaki, H. Seed Germination and Dormancy: The Classic Story, New Puzzles, and Evolution. J. Integr. Plant Biol. 2019, 61, 541–563. [Google Scholar] [CrossRef] [PubMed]
- Carrillo-Barral, N.; del Carmen Rodríguez-Gacio, M.; Matilla, A.J. Delay of Germination-1 (DOG1): A Key to Understanding Seed Dormancy. Plants 2020, 9, 480. [Google Scholar] [CrossRef] [PubMed]
- Penfield, S.; Josse, E.-M.; Kannangara, R.; Gilday, A.D.; Halliday, K.J.; Graham, I.A. Cold and Light Control Seed Germination through the bHLH Transcription Factor SPATULA. Curr. Biol. 2005, 15, 1998–2006. [Google Scholar] [CrossRef] [PubMed]
- Tatematsu, K.; Nakabayashi, K.; Kamiya, Y.; Nambara, E. Transcription Factor AtTCP14 Regulates Embryonic Growth Potential during Seed Germination in Arabidopsis thaliana. Plant J. 2008, 53, 42–52. [Google Scholar] [CrossRef]
- Jiang, W.; Yu, D. Arabidopsis WRKY2 Transcription Factor Mediates Seed Germination and Postgermination Arrest of Development by Abscisic Acid. BMC Plant Biol. 2009, 9, 96. [Google Scholar] [CrossRef]
- Lee, K.; Lee, H.G.; Yoon, S.; Kim, H.U.; Seo, P.J. The Arabidopsis MYB96 Transcription Factor Is a Positive Regulator of ABSCISIC ACID-INSENSITIVE4 in the Control of Seed Germination. Plant Physiol. 2015, 168, 677–689. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Feng, C.-Z.; Ye, Q.; Wu, W.-H.; Chen, Y.-F. Arabidopsis WRKY6 Transcription Factor Acts as a Positive Regulator of Abscisic Acid Signaling during Seed Germination and Early Seedling Development. PLoS Genet. 2016, 12, e1005833. [Google Scholar] [CrossRef] [PubMed]
- Jo, L.; Pelletier, J.M.; Harada, J.J. Central Role of the LEAFY COTYLEDON1 Transcription Factor in Seed Development. J. Integr. Plant Biol. 2019, 61, 564–580. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Huang, L.; Sun, H.; Lyu, Y. ‘Chongtian’: A New Zelkova schneideriana cultivar. HortScience 2023, 58, 465–466. [Google Scholar] [CrossRef]
- Zhou, Y.; Lu, X.; Zhang, G. Potentially Differential Impacts on Niche Overlap between Chinese Endangered Zelkova schneideriana and Its Associated Tree Species under Climate Change. Front. Ecol. Evol. 2023, 11, 1218149. [Google Scholar] [CrossRef]
- Song, Y.; Bétrisey, S.; Kozlowski, G. Zelkova schneideriana. The IUCN Red List of Threatened Species 2018. In 2018 Edn: IUCN Global Species Programme, the IUCN Species Survival Commission (SSC) and the IUCN Red List Partnership; World Headquarters: Gland, Switzerland, 2018; p. 13. [Google Scholar]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-Length Transcriptome Assembly from RNA-Seq Data without a Reference Genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and Sensitive Protein Alignment Using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate Transcript Quantification from RNA-Seq Data with or without a Reference Genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and Memory-Efficient Alignment of Short DNA Sequences to the Human Genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef]
- Ren, Y.; Shen, F.; Liu, J.A.; Liang, W.; Zhang, C.; Lian, T.; Jiang, L. Application of Methionine Increases the Germination Rate of Maize Seeds by Triggering Multiple Phenylpropanoid Biosynthetic Genes at Transcript Levels. Plants 2023, 12, 3802. [Google Scholar] [CrossRef]
- Mao, X.; Cai, T.; Olyarchuk, J.G.; Wei, L. Automated Genome Annotation and Pathway Identification Using the KEGG Orthology (KO) as a Controlled Vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef]
- Ni, Z.; Han, X.; Yang, Z.; Xu, M.; Feng, Y.; Chen, Y.; Xu, L. Integrative Analysis of Wood Biomass and Developing Xylem Transcriptome Provide Insights into Mechanisms of Lignin Biosynthesis in Wood Formation of Pinus massoniana. Int. J. Biol. Macromol. 2020, 163, 1926–1937. [Google Scholar] [CrossRef] [PubMed]
- McGinty, E.M.; Murphy, K.M.; Hauvermale, A.L. Seed Dormancy and Preharvest Sprouting in Quinoa (Chenopodium quinoa Willd). Plants 2021, 10, 458. [Google Scholar] [CrossRef] [PubMed]
- Koornneef, M.; Bentsink, L.; Hilhorst, H. Seed Dormancy and Germination. Curr. Opin. Plant Biol. 2002, 5, 33–36. [Google Scholar] [CrossRef]
- Nambara, E.; Okamoto, M.; Tatematsu, K.; Yano, R.; Seo, M.; Kamiya, Y. Abscisic Acid and the Control of Seed Dormancy and Germination. Seed Sci. Res. 2010, 20, 55–67. [Google Scholar] [CrossRef]
- Huang, X.; Hou, L.; Meng, J.; You, H.; Li, Z.; Gong, Z.; Yang, S.; Shi, Y. The Antagonistic Action of Abscisic Acid and Cytokinin Signaling Mediates Drought Stress Response in Arabidopsis. Mol. Plant 2018, 11, 970–982. [Google Scholar] [CrossRef]
- Wang, H.; Tang, J.; Liu, J.; Hu, J.; Liu, J.; Chen, Y.; Cai, Z.; Wang, X. Abscisic Acid Signaling Inhibits Brassinosteroid Signaling through Dampening the Dephosphorylation of BIN2 by ABI1 and ABI2. Mol. Plant 2018, 11, 315–325. [Google Scholar] [CrossRef]
- Xie, H.; Wang, D.; Qin, Y.; Ma, A.; Fu, J.; Qin, Y.; Hu, G.; Zhao, J. Genome-Wide Identification and Expression Analysis of SWEET Gene Family in Litchi chinensis Reveal the Involvement of LcSWEET2a/3b in Early Seed Development. BMC Plant Biol. 2019, 19, 499. [Google Scholar] [CrossRef] [PubMed]
- Née, G.; Kramer, K.; Nakabayashi, K.; Yuan, B.; Xiang, Y.; Miatton, E.; Finkemeier, I.; Soppe, W.J.J. Delay of Germination1 Requires PP2C Phosphatases of the ABA Signalling Pathway to Control Seed Dormancy. Nat. Commun. 2017, 8, 72. [Google Scholar] [CrossRef]
- Piskurewicz, U.; Jikumaru, Y.; Kinoshita, N.; Nambara, E.; Kamiya, Y.; Lopez-Molina, L. The Gibberellic Acid Signaling Repressor RGL2 Inhibits Arabidopsis Seed Germination by Stimulating Abscisic Acid Synthesis and ABI5 Activity. Plant Cell 2008, 20, 2729–2745. [Google Scholar] [CrossRef]
- Lee, K.P.; Piskurewicz, U.; Turečková, V.; Strnad, M.; Lopez-Molina, L. A Seed Coat Bedding Assay Shows That RGL2-Dependent Release of Abscisic Acid by the Endosperm Controls Embryo Growth in Arabidopsis Dormant Seeds. Proc. Natl. Acad. Sci. USA 2010, 107, 19108–19113. [Google Scholar] [CrossRef] [PubMed]
- Kleczkowski, L.A.; Kunz, S.; Wilczynska, M. Mechanisms of UDP-Glucose Synthesis in Plants. Crit. Rev. Plant Sci. 2010, 29, 191–203. [Google Scholar] [CrossRef]
- Figueroa, C.M.; Asencion Diez, M.D.; Ballicora, M.A.; Iglesias, A.A. Structure, Function, and Evolution of Plant ADP-Glucose Pyrophosphorylase. Plant Mol. Biol. 2022, 108, 307–323. [Google Scholar] [CrossRef] [PubMed]
- Li, C.Y.; Li, C.; Lu, Z.X.; Li, W.H.; Cao, L.P. Morphological Changes of Starch Granules during Grain Filling and Seed Germination in Wheat. Starch-Stärke 2012, 64, 166–170. [Google Scholar] [CrossRef]
- Shendurse, A.M.; Khedkar, C.D. Glucose: Properties and Analysis. In Encyclopedia of Food and Health; Oxford University Press: Oxford, UK, 2016; Volume 3, pp. 239–247. [Google Scholar] [CrossRef]
- Kühn, C.; Barker, L.; Bürkle, L.; Frommer, W.-B. Update on Sucrose Transport in Higher Plants. J. Exp. Bot. 1999, 50, 935–953. [Google Scholar] [CrossRef]
- Baroja-Fernández, E.; Muñoz, F.J.; Li, J.; Bahaji, A.; Almagro, G.; Montero, M.; Etxeberria, E.; Hidalgo, M.; Sesma, M.T.; Pozueta-Romero, J. Sucrose Synthase Activity in the Sus1/Sus2/Sus3/Sus4 Arabidopsis Mutant Is Sufficient to Support Normal Cellulose and Starch Production. Proc. Natl. Acad. Sci. USA 2012, 109, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Cai, G.; Kim, S.-C.; Li, J.; Zhou, Y.; Wang, X. Transcriptional Regulation of Lipid Catabolism during Seedling Establishment. Mol. Plant 2020, 13, 984–1000. [Google Scholar] [CrossRef] [PubMed]
- Weitbrecht, K.; Müller, K.; Leubner-Metzger, G. First off the Mark: Early Seed Germination. J. Exp. Bot. 2011, 62, 3289–3309. [Google Scholar] [CrossRef]
- Zhu, L.; Zhao, X.; Xu, Y.; Wang, Q.; Wang, H.; Wu, D.; Jiang, L. Effect of Germination Potential on Storage Lipids and Transcriptome Changes in Premature Developing Seeds of Oilseed Rape (Brassica napus L.). Theor. Appl. Genet. 2020, 133, 2839–2852. [Google Scholar] [CrossRef]
- Feussner, I.; Kühn, H.; Wasternack, C. Lipoxygenase-Dependent Degradation of Storage Lipids. Trends Plant Sci. 2001, 6, 268–273. [Google Scholar] [CrossRef]
- Duan, X.; Jiang, W.; Wu, K.; Chen, J.; Li, Y.; Tao, Z. Integrating Transcriptomics and Hormones Dynamics Reveal Seed Germination and Emergence Process in Polygonatum cyrtonema Hua. Int. J. Mol. Sci. 2023, 24, 3792. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhao, G.; Li, Y.; Wei, S.; Dong, Y.; Jiao, R. Integrative Analysis of the Transcriptome and Metabolome Reveals the Mechanism of Chinese Fir Seed Germination. Forests 2023, 14, 676. [Google Scholar] [CrossRef]
- Li, J.; Li, X.; Bai, Y.; Xie, Y.; Li, L.; Mu, S.; Gao, J. Transcriptome Analysis of Energy Supply Process During Seed Germination in Phyllostachys edulis. Plant Mol. Biol. Rep. 2023, 41, 489–511. [Google Scholar] [CrossRef]
- Singh, P.; Arif, Y.; Miszczuk, E.; Bajguz, A.; Hayat, S. Specific Roles of Lipoxygenases in Development and Responses to Stress in Plants. Plants 2022, 11, 979. [Google Scholar] [CrossRef] [PubMed]
- Vogt, T. Phenylpropanoid biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar] [CrossRef] [PubMed]
- Vanholme, R.; Demedts, B.; Morreel, K.; Ralph, J.; Boerjan, W. Lignin Biosynthesis and Structure. Plant Physiol. 2010, 153, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Zhong, R.; Cui, D.; Ye, Z. Secondary Cell Wall Biosynthesis. New Phytol. 2019, 221, 1703–1723. [Google Scholar] [CrossRef] [PubMed]
- Campilho, A.; Nieminen, K.; Ragni, L. The Development of the Periderm: The Final Frontier between a Plant and Its Environment. Curr. Opin. Plant Biol. 2020, 53, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Ganapathy, G.; Keerthi, D.; Nair, R.A.; Pillai, P. Correlation of Phenylalanine Ammonia Lyase (PAL) and Tyrosine Ammonia Lyase (TAL) Activities to Phenolics and Curcuminoid Content in Ginger and Its Wild Congener, Zingiber zerumbet Following Pythium myriotylum Infection. Eur. J. Plant Pathol. 2016, 145, 777–785. [Google Scholar] [CrossRef]
- Wu, Z.; Wang, N.; Hisano, H.; Cao, Y.; Wu, F.; Liu, W.; Bao, Y.; Wang, Z.; Fu, C. Simultaneous regulation of F5H in COMT-RNAi transgenic switchgrass alters effects of COMT suppression on syringyl lignin biosynthesis. Plant Biotechnol. J. 2019, 17, 836–845. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, F.; Wei, T.; Yang, C.; Yang, Y.; Luo, Z.; Jiang, Y. Combined Analysis of Untargeted Metabolomics and Transcriptomics Revealed Seed Germination and Seedling Establishment in Zelkova schneideriana. Genes 2024, 15, 488. https://doi.org/10.3390/genes15040488
Yan F, Wei T, Yang C, Yang Y, Luo Z, Jiang Y. Combined Analysis of Untargeted Metabolomics and Transcriptomics Revealed Seed Germination and Seedling Establishment in Zelkova schneideriana. Genes. 2024; 15(4):488. https://doi.org/10.3390/genes15040488
Chicago/Turabian StyleYan, Fengxia, Tangmei Wei, Chao Yang, Yanbing Yang, Zaiqi Luo, and Yunli Jiang. 2024. "Combined Analysis of Untargeted Metabolomics and Transcriptomics Revealed Seed Germination and Seedling Establishment in Zelkova schneideriana" Genes 15, no. 4: 488. https://doi.org/10.3390/genes15040488
APA StyleYan, F., Wei, T., Yang, C., Yang, Y., Luo, Z., & Jiang, Y. (2024). Combined Analysis of Untargeted Metabolomics and Transcriptomics Revealed Seed Germination and Seedling Establishment in Zelkova schneideriana. Genes, 15(4), 488. https://doi.org/10.3390/genes15040488