Abstract
Background/Objectives: Cold stress is the main environmental factor that affects the growth and development of rice, leading to a decrease in its yield and quality. However, the molecular mechanism of rice’s low-temperature resistance remains incompletely understood. Methods: In this study, we conducted a joint analysis of miRNA and mRNA expression profiles in the cold-resistant material Yongning red rice and the cold-sensitive material B3 using high-throughput sequencing. Results: 194 differentially expressed miRNAs (DEMIs) and 14,671 differentially expressed mRNAs (DEMs) were identified. Among them, 19 DEMIs, including miR1437, miR1156, miR166, miR1861, and miR396_2 family members, showed opposite expression during the early or late stages of low-temperature treatment in two varieties, while 13 DEMIs were specifically expressed in Yongning red rice, indicating that these miRNAs are involved in rice’s resistance to low temperature. In the transcriptome analysis, 218 DEMs exhibited opposite expressions during the early or late stages of low-temperature treatment in two varieties. GO enrichment analysis indicated that these DEMs were enriched in biological processes such as a defense response to fungi, a defense response to bacteria, a plant-type cell wall modification, single-organism cellular processes, a response to chitin, and the regulation of a plant-type hypersensitive response, as well as in cellular components such as the apoplast, nucleus, vacuole, plasma membrane, and plasmodesma. Twenty-one genes were further selected as potential candidates for low-temperature resistance. The joint analysis of miRNA and mRNA expression profiles showed that 38 miRNAs corresponding to 39 target genes were candidate miRNA–mRNA pairs for low-temperature resistance. Conclusions: This study provides valuable resources for determining the changes in miRNA and mRNA expression profiles induced by low temperatures and enables the provision of valuable information for further investigating the molecular mechanisms of plant resistance to low temperatures and for the genetic improvement of cold-resistant varieties.
1. Introduction
Rice (Oryza sativa L.) is both a model plant and one that provides carbohydrate needs for over half of the world’s population [1]. As a tropical and subtropical crop, rice is susceptible to low-temperature stress. Under low-temperature conditions, rice may experience yellowing of leaves, wilting, tillering reduction, delayed heading, and spikelet sterility. In severe cases, it may even lead to plant death, ultimately resulting in a decrease in yield [2,3,4,5]. With the increase in rice cultivation area, rice planting is expanding from tropical and subtropical regions to high-altitude areas and cool summer areas, and the possibility of cold damage is increasing. Therefore, enhancing the cold tolerance of rice varieties is very important, indicating the need to explore the molecular mechanisms of rice cold tolerance.
MicroRNAs (miRNAs) are a class of noncoding endogenous small RNA in eukaryotes, typically ranging in length from 19 to 24 nucleotides (nt) [6,7]. It mainly regulates the expression level of its target gene mRNA through shear degradation, inhibition of translation, and chromatin remodeling (methylation). Numerous studies have shown that plant miRNAs play a crucial role in plant growth and development, epigenetics, and the response to biotic and abiotic stresses [7,8,9,10,11]. miR1432 negatively regulates abiotic stress tolerance by targeting OsACAs and affects plant growth and development [8]; meanwhile, another study found that it can also regulate drought tolerance by directly targeting the CALMODULIN-LIKE2 gene in rice [12]. The overexpression of miR535 in rice can lead to low-temperature-induced cell death, excessive accumulation of ROS, and impaired osmotic regulation [13]. In maize, miR169s negatively regulates resistance to bipolaris maydis by affecting the SA-dependent signaling pathway and ROS accumulation [14]. miR397b participates in the regulation of root hair growth by targeting genes encoding reduced residual arabinose (RRA1/RRA2) in Arabidopsis (Arabidopsis thaliana) [15]. miR160, miR165, and miR166 are both involved in regulating the number, size, and weight of Arabidopsis seeds in a ROS-dependent manner [16].
The rapid development of sequencing technology and bioinformatics has led to the emergence of high-throughput sequencing as a prominent approach in the study of plant low-temperature defense mechanisms [17,18,19,20,21]. For example, through comparative transcriptome analysis, 795 DEMs related to low-temperature response were identified in the indica rice (O. sativa L. ssp. indica) TGMS 33S variety (cold-resistant material). Functional and enrichment analysis of these DEMs showed that the expression of genes related to nutritional reserve activity was significantly up-regulated in 33S, indicating that these genes play an important role in low-temperature response [17]. In Camellia japonica (Naidong), 4544 significant DEMs were identified through transcriptomics, and analysis revealed that transcription factors, lipid metabolism, and carbon metabolism play important roles in Camellia japonica (Naidong) resistance to low-temperature stress [18]. In Dongxiang wild rice, 16 significant DEMIs under low-temperature stress were identified through miRNA sequencing [19]. Among these miRNAs, miR408-5p was found to be significantly upregulated under low-temperature stress. The overexpression of miR408 has been shown to improve cold tolerance in rice [19,20]. Similarly, through miRNA sequencing, under low-temperature stress, 145 known and 876 novel miRNAs were identified; among them, miR164, MiR156, miR167, miR169, miR394, miR396, and miR398 were involved in response to low-temperature stress in maize [21].
Although there have been many independent reports on the low-temperature response transcriptome profiles of miRNAs and mRNAs, there are few reports on the joint analysis of miRNAs and mRNAs transcriptome profiles to reveal the mechanisms of low-temperature resistance. In order to further elucidate the molecular mechanism of rice response to low temperatures, this study applied high-throughput sequencing for the first time to combine analysis of the expression profiles of miRNAs and mRNAs in japonica rice (O. sativa L. ssp. japonica). The research results provide valuable resources for studying the specific expression of miRNA and mRNA in rice under low-temperature stress and help deepen our understanding of the miRNA–mRNA regulatory mechanism in rice under low-temperature stress.
2. Materials and Methods
2.1. Plant Materials and Stress Treatment
The japonica rice (O. sativa L. ssp. japonica) landrace Yongning red rice was provided by the Rice Research Institute of Yunnan Agricultural University. It is a high-altitude japonica rice distributed across Ninglang County, Lijiang City, Yunnan Province, China, which is the highest rice-producing area in the world at an altitude of 1800–2750 m. It has the characteristic of extremely low-temperature tolerance. Japonica rice (O. sativa L. ssp. japonica) strain B3 was provided by the Panxi Crops Research and Utilization Key Laboratory of Sichuan Province from Xichang University. It is sensitive to low temperatures. To investigate the low-temperature tolerance mechanism of Yongning red rice, Yongning red rice and B3 were used for a combined miRNA and transcriptome analysis. For analysis, three biological replicates were employed for each treatment, with 100 seedlings constituting the sample size for each replicate. Rice seeds were grown to the 3–4 leaf stage under a 12 h light/12 h dark cycle at 28 °C. The 3–4 leaf seedlings were then treated at 5 °C for a 12 h light/12 h dark cycle. Two rice lines were sampled at low temperatures for 0, 24, and 48 h. Five seedlings were randomly selected from each replicate to collect their root, stem, and leaf tissues. After washing away the soil, the tissues were immediately frozen in liquid nitrogen and stored at −80 °C until RNA extraction. The samples taken from the Yongning red rice line at 0, 24, and 48 h were labeled R0, Re, and Rl, respectively. The samples taken from the B3 line at 0, 24, and 48 h were labeled S0, Se, and Sl, respectively.
2.2. RNA Extraction
The total RNA was extracted from a mixture of rice roots, stems, and leaves using a Trizol reagent (Life Technologies, Carlsbad, CA, USA), according to the manufacturer’s instructions. The concentration and purity of the RNA were determined using a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Wilmington, DE, USA). RNA integrity was assessed using the RNA Nano 6000 Assay Kit on the Agilent Bioanalyzer 2100 system (Agilent Technologies, Palo Alto, CA, USA).
2.3. miRNA Library Construction and Sequencing
Small RNA libraries were constructed using the VAHTS™ Small RNA Library Prep Kit for Illumina (Vazyme, Nanjing, China) according to the manufacturer’s instructions. The small RNAs were first ligated with the 3′NEXTflex Adenylated Adapter, followed by the 5′ SR Adapter. After first-strand synthesis and PCR amplification, the final bands were purified and sequenced on Illumina’s NovaSeq 6000 platform. After sequencing, the reads were processed to remove any low-quality reads, reads containing greater than or equal to 10% unknown bases, adaptor sequences, and reads of <18 nt and >30 nt.
2.4. mRNA Library Construction and Sequencing
A total of 1 μg of RNA per sample was used to prepare the RNA libraries. Sequencing libraries were prepared using the Hieff NGS Ultima Dual-mode mRNA Library Prep Kit for Illumina (Yeasen, Shanghai, China), according to the manufacturer’s recommendations. The library quality was assessed on the Agilent Bioanalyzer 2100 system. Libraries were then sequenced on an Illumina NovaSeq platform to generate 150 bp paired-end reads according to the manufacturer’s instructions. Clean data were obtained by removing adapter-containing reads, ploy-N-containing reads, and low-quality reads from the raw data, and all downstream analyses were based on clean, high-quality data.
The clean reads were mapped to the rice reference genome (Oryza_sativa.MSU_v7.0) using Hisat2 tools [22]. Significant DEMs were identified by DESeq2 [23]. We used |log2(FC)| ≥ 1 and p ≤ 0.05 as thresholds to determine cold-induced DEMs.
2.5. Bioinformatics Analysis of Small RNA Sequencing Data
Clean reads were sequentially aligned with the Silva database, GtRNAdb, Repbase, and Rfam databases using Bowtie software. Unannotated reads containing miRNA sequences were obtained by filtering out noncoding RNAs (ncRNAs), such as snoRNA, tRNA, snRNA, rRNA, and repeat sequences. These unannotated reads were then compared with the rice reference genome (Oryza_sativa.MSU_v7.0) using Bowtie software to analyze their expression and distribution. To identify known miRNAs, alignment was performed with mature miRNA sequences from the reference genome and the miRBase (v22) database, including their downstream 5 nt and upstream 2 nt ranges. A mismatch of no more than one base was allowed for a sequence to be considered a known miRNA. For predicting novel miRNAs, miRDeep2 software was used to align the obtained reads to genomic positions to identify possible precursor sequences. Based on the distribution information of the reads on the precursor sequences (based on miRNA production characteristics, mathematics, star, loop) and precursor structure energy information (RNAfold randfold), Bayesian models were used for scoring, which facilitated the prediction of new miRNAs.
DEMIs were identified by DESeq2 with FC ≥ 1.5 (|Log2(FC)| ≥ 0.58) and p ≤ 0.05 as thresholds to determine cold-induced DEMIs. The target genes of the miRNAs were predicted using the TargetFinder program [24], and the sequences of the target genes were compared with the Swiss-Prot, NR, COG, GO, KOG, KEGG, and Pfam databases using BLAST tools.
2.6. qRT-PCR Analysis
Total RNA was extracted from mixed tissues of rice roots, stems, and leaves using a Trizol reagent (Life Technologies, CA, USA). Reverse transcription of miRNA and mRNA was performed using an M5 miRNA cDNA Synthesis Kit (Mei5bio, Beijing, China) and an M5 Super plus qPCR RT Kit with gDNA remover (Mei5bio, Beijing, China), respectively. Then, the M5 miRNA qPCR Assay Kit (Mei5bio, Beijing, China) and 2X M5 HiPer SYBR Premix EsTaq (Mei5bio, Beijing, China) were used for qRT-PCR analysis of miRNA and mRNA expression, using the CFX96 real-time PCR system (Bio-Rad, Berkeley, CA, USA). Three biological replicates were performed for each experiment. The expression of miRNAs and genes was calculated using the 2−ΔΔC (t) method [25]. Ubiquitin and U6 were used as internal reference genes for qRT-PCR analysis of mRNA and miRNA expression, respectively. Table S1 lists the primers used in qRT-PCR.
3. Results
3.1. Evaluation of Cold Tolerance in Rice Seedling Stage
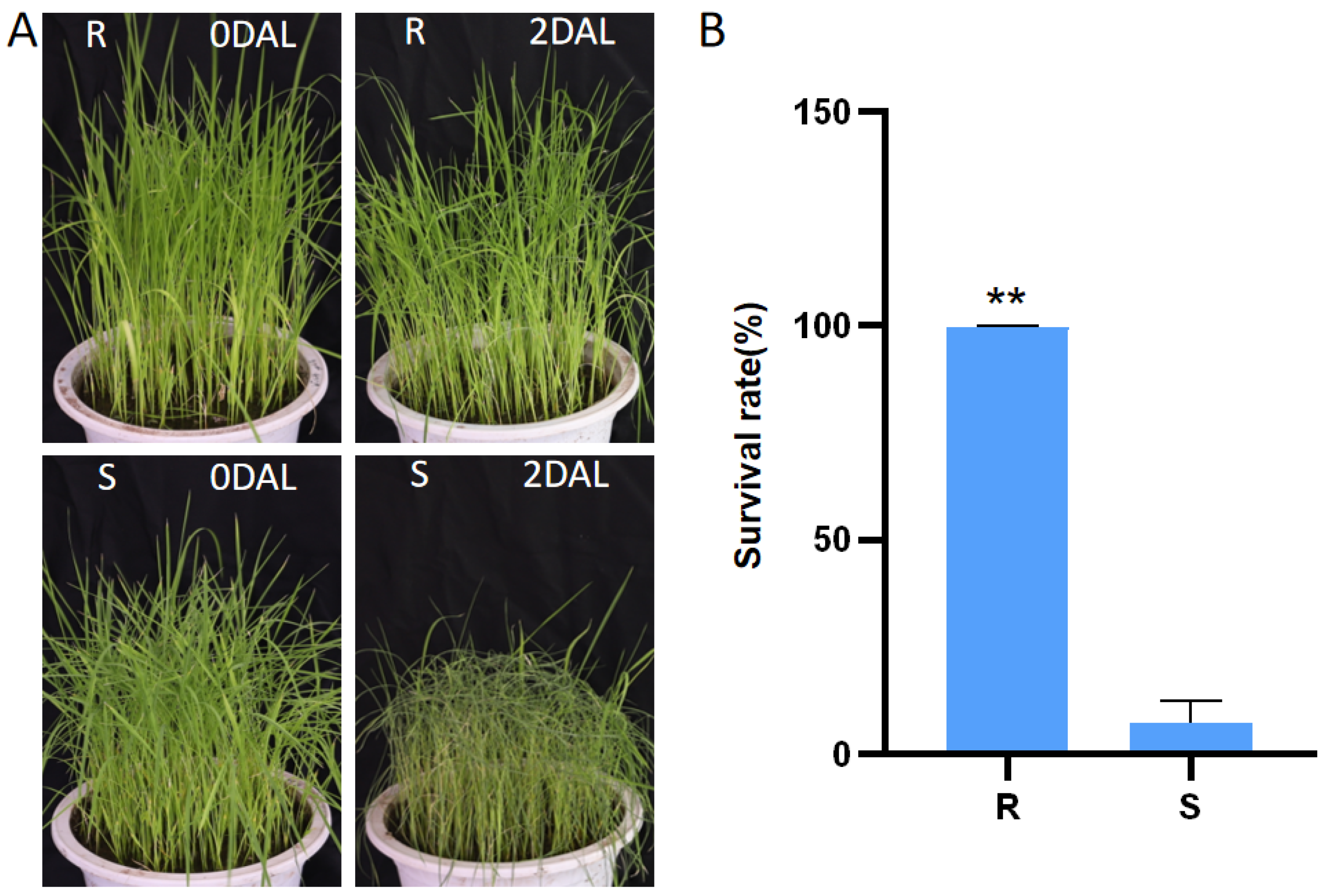
Yongning red rice exhibited a cold tolerance phenotype and had a higher survival rate compared to B3 (Figure 1). Therefore, Yongning red rice and B3 were selected for miRNA and mRNA sequencing before and after 24 and 48 h of low-temperature treatment. By comparing the miRNA and mRNA transcriptome profiles of Yongning red rice and B3, the mechanism of low-temperature tolerance in Yongning red rice was explored.
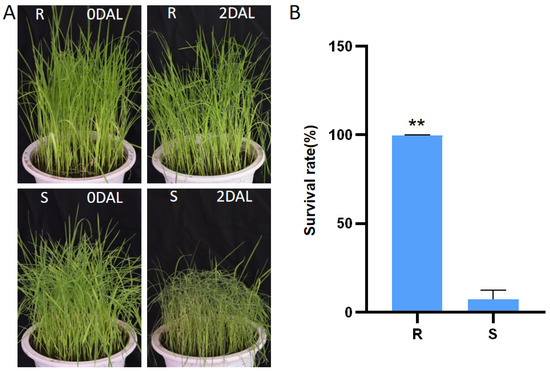
Figure 1.
Evaluation of cold tolerance in the rice seedling stage of the Yongning red rice and B3. (A) Image of cold tolerance identification of Yongning red rice and B3 seedlings. (B) Survival rates of Yongning red rice and B3 plants after 2 days of cold treatment at 5 °C and a 5-day recovery period. R stands for Yongning red rice; S stands for B3; DAL, days after low-temperature treatment; ** for p < 0.01.
3.2. Small RNA Library Construction and Sequencing
Total raw reads of 18,762,842 to 56,372,907 were obtained in R0, Re, Rl, S0, Se and Sl libraries. After removing all low-quality reads, reads with unknown bases greater than or equal to 10%, adaptor sequences, and reads of <18 nt and >30 nt, a total of 11,722,741 to 21,890,454 clean reads remained in the 18 libraries, respectively (Table S2). A total of 831 miRNAs were obtained, of which 244 were novel miRNAs, accounting for 29.36%.
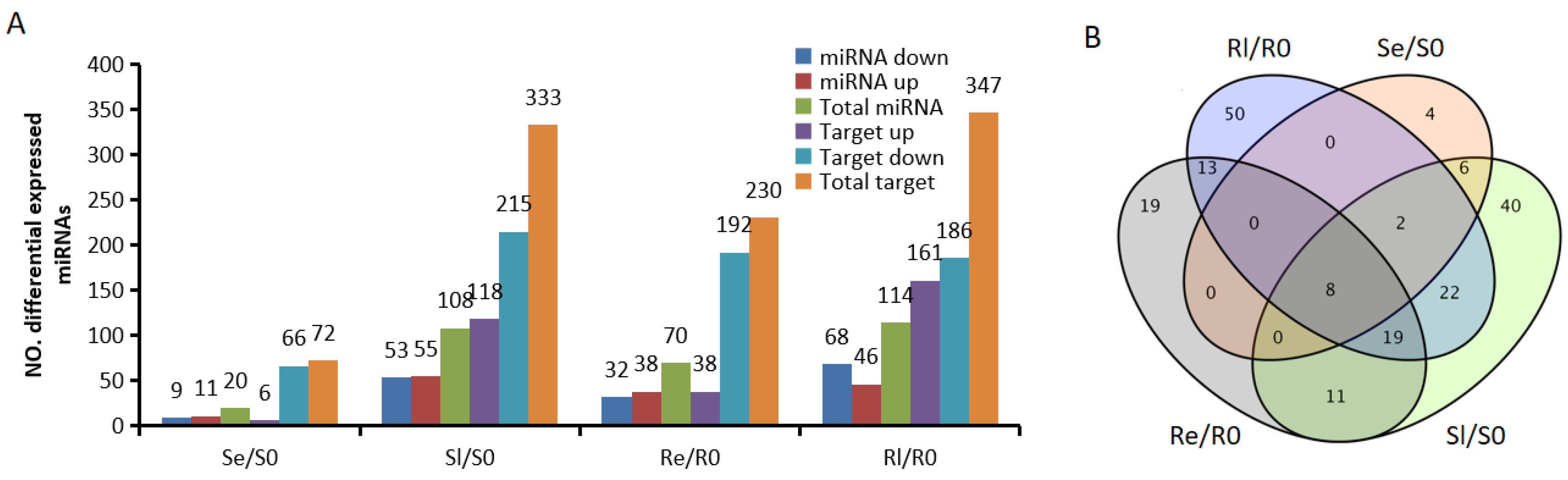
3.3. DEMIs Across Different Materials Before and After Low-Temperature Treatment
After normalizing raw sequence reads, average normalized reads from three independent biological replicates were selected for further analysis. The expression levels of miRNAs were compared across the different groups. A total of 194 DEMIs were identified among the various cryogenic treatment stages with fold changes ≥ 1.5, p < 0.05 based on the average of three replicates (Figure 2B). In the early stages of low-temperature treatment, more DEMIs were observed in Yongning red rice (70) compared to B3 (20); however, in the late stages of low-temperature treatment, the number of DEMIs in Yongning red rice (114) and B3 (108) was not significantly different (Figure 2A), suggesting that many miRNAs involved in low-temperature regulation were not immediately expressed in B3.
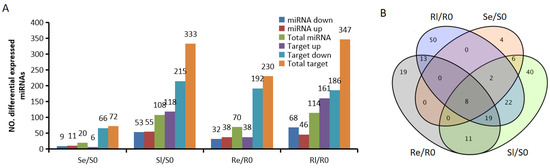
Figure 2.
DEMIs in the comparisons. (A) Number of up- and downregulated miRNAs and target genes in comparing different low-temperature treatment periods (fold change > 1.5, p < 0.05). (B) Venn diagrams of the unique and common DEMIs.
3.4. Identification of miRNAs Related to Cold Tolerance
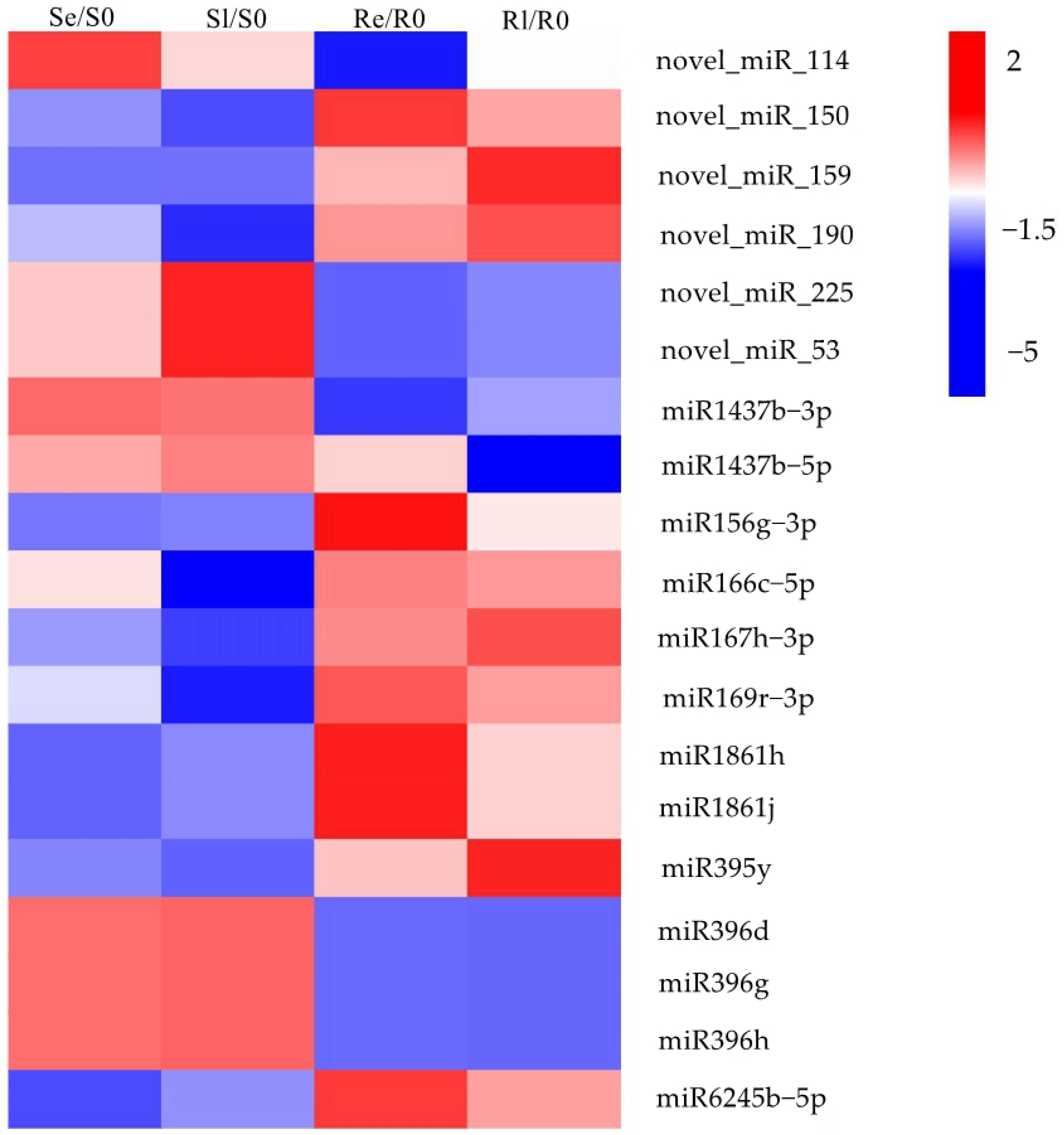
To identify miRNAs associated with plant chilling stress, Venn diagrams were used to visualize the DEMIs in early and late stages of low-temperature treatment for the two varieties (Se/S0, Sl/S0, Re/R0, and Rl/R0) (Figure 2B). In total, 69 DEMIs were expressed in Re/R0 or Rl/R0, and 13 were expressed specifically in Re/R0 and Rl/R0 (Figure 2B). Additionally, 19 DEMIs showed opposing expression patterns between the two varieties after low-temperature treatment (Figure 3). Of these, 6 were novel miRNAs, and 13 were known miRNAs, many of which belonged to known miRNA families, including miR1437, miRNA156, miR166, miR167_1, miR169_1, miR1861, miR395, miR396_2, and miR9783. Members of these miRNA families have been reported to play roles in various biological processes such as growth and development, flowering regulation, nutrient absorption, agricultural trait regulation, and responses to both biotic and abiotic stresses [26,27,28,29,30,31,32,33,34,35,36,37,38]. These DEMIs, which respond to chilling stress, may be involved in the regulation of low-temperature responses in rice.
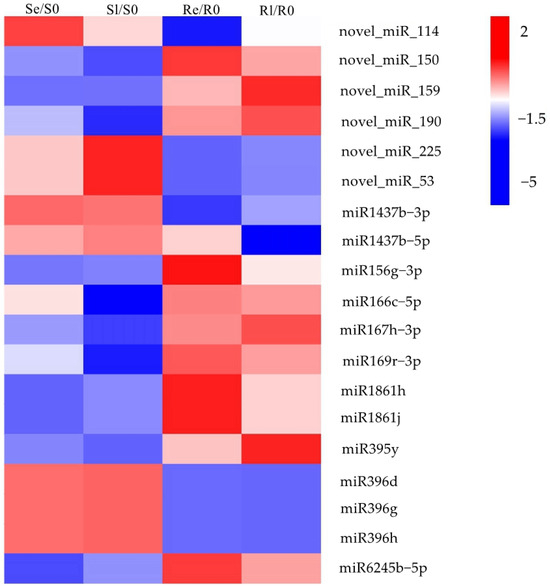
Figure 3.
Heatmap of DEMIs were opposite expressed in two varieties after low-temperature treatment. The heatmap is constructed based on log2fold change values.
3.5. General mRNA Expression Profiles
To analyze gene expression and profile all the targets of the DEMIs in response to cold tolerance, mRNA libraries were constructed. A total of 39,848,202 to 59,658,622 clean reads were sequenced from 18 mRNA libraries. After filtering out low-quality reads from the samples of Yongning red rice, 90.78–95.96% of the reads were mapped to 24,766–27,477 rice genes (Table S3). In the B3 replicates, 91.00–95.22% of reads were mapped to 26,335–28,109 rice genes (Table S3).
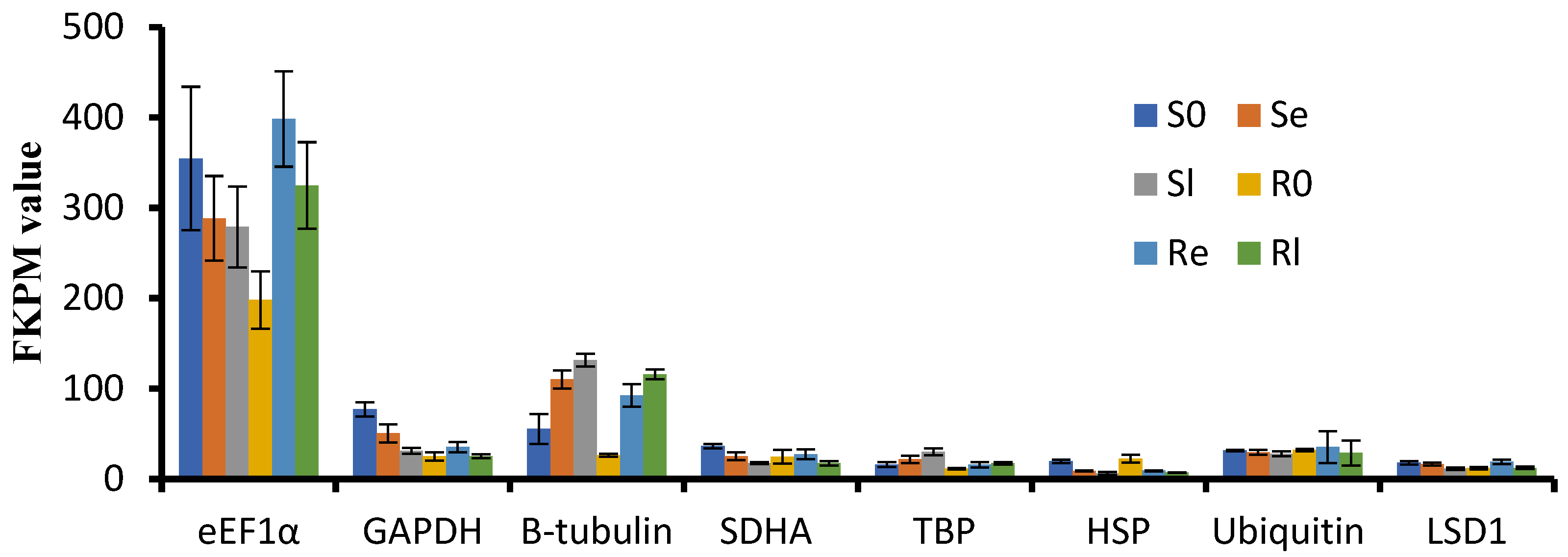
To select stable reference genes for qRT-PCR analysis during low-temperature treatment, we carefully selected eight commonly used reference genes: SDHA (Os07g0424), LSD1 (Os12g41700), TBP (Os03g45410), GAPDH (Os02g38920), β-tubulin (Os03g56810), HSP (Os03g31300), eEF1α (Os03g08020) and Ubiquitin (Os03g03920). We evaluated the respective FPKM values of these genes from our RNA-seq data (Figure 4). Expression levels of eEF1α, β-tubulin, GAPDH, and HSP showed significant changes before and after low-temperature treatment, while LSD1 and TBP were stable but relatively low in expression. Based on these findings, Ubiquitin was selected as the reference gene for qRT-PCR analysis.
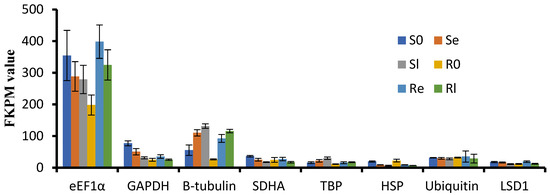
Figure 4.
FKPM values of GAPDH, SDHA, TBP, eEF1α, Ubiquitin, LSD1, β-tubulin, and HSP from mRNA-sequencing data.
3.6. DEMs in Response to Low-Temperature Treatment
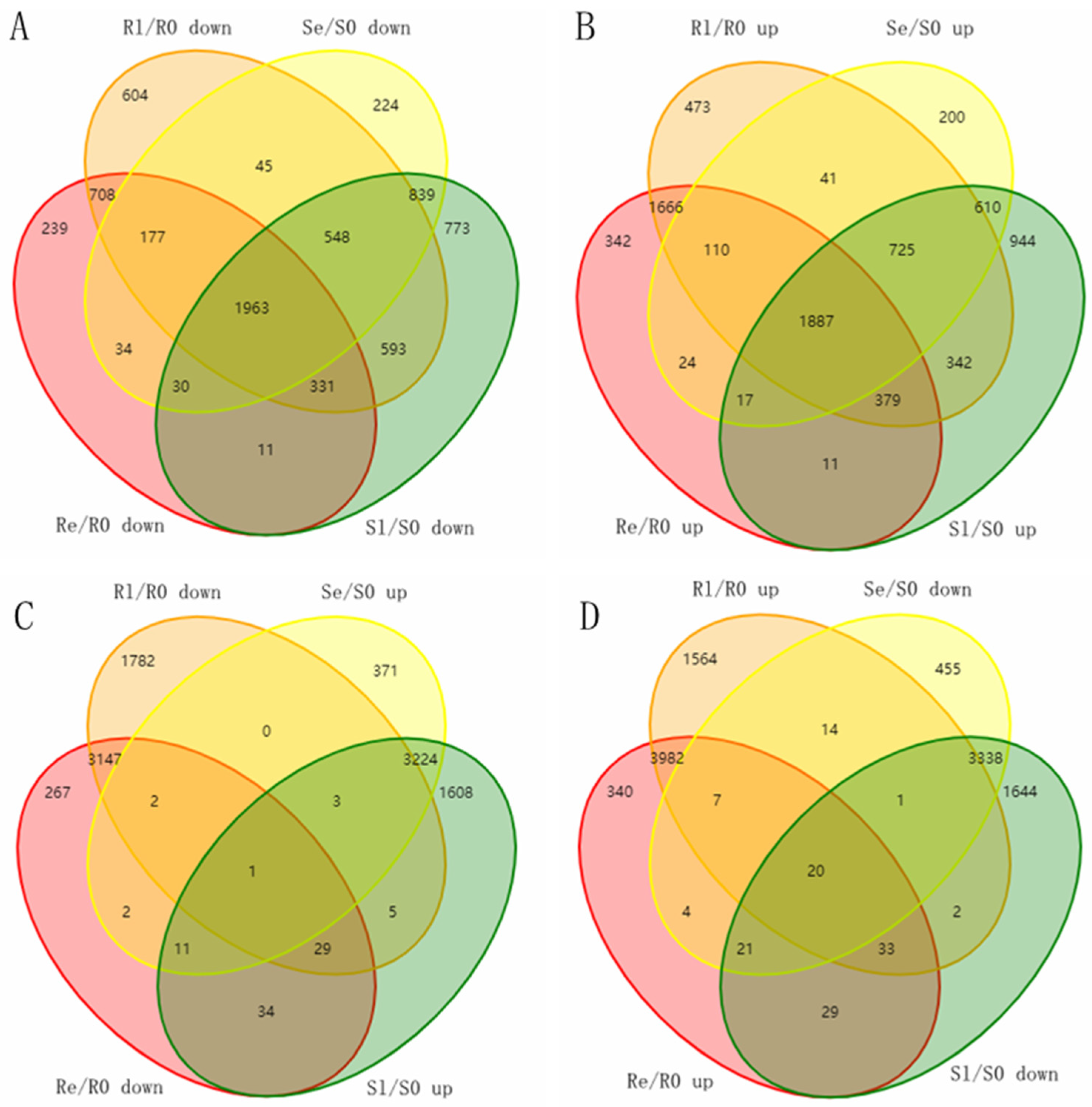
A total of 14,671 DEMs (log 2 FC ≥ 2, p < 0.05) were detected between different low-temperature treatment stages (Table 1 and Figure 5). The DEMs in the Yongning red rice and B3 plants at various low-temperature treatment stages were hierarchically clustered. Across the four comparisons, the expression patterns of the DEMs were consistent, with the similar up- or downregulation observed (Figure S1).

Table 1.
DEMs between the Yongning red rice and B3 plants in response to low-temperature treatment.
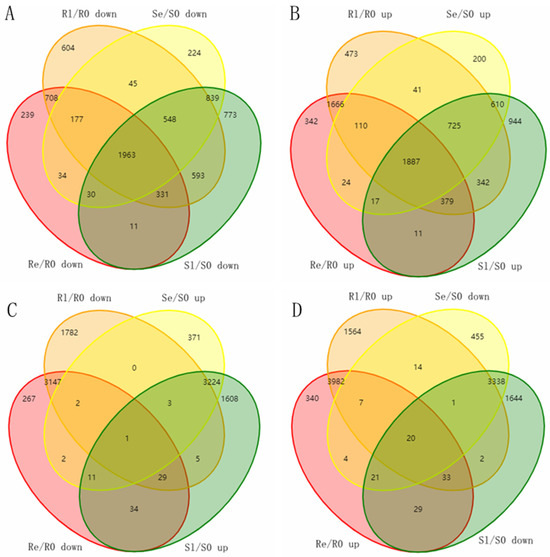
Figure 5.
Venn diagram of the unique and shared DEMs. (A) Venn diagram of downregulated genes in Se/S0, Sl/S0, Re/R0, and Rl/R0. (B) Venn diagram of upregulated genes in Se/S0, Sl/S0, Re/R0, and Rl/R0. (C) Venn diagram of upregulated genes in Se/S0, Sl/S0, and downregulated genes in Re/R0 and Rl/R0. (D) Venn diagram of downregulated genes in Se/S0, Sl/S0, and upregulated genes in Re/R0 and Rl/R0. DEMs were screened using a threshold of fold change ≥ 2, p < 0.05.
During low-temperature treatment, the number of DEMs in Yongning red rice plants was consistently higher than that in B3. Notably, we found that the number of downregulated DEMs in Yongning red rice plants was consistently lower than that in B3, while more DEMs were upregulated in Yongning red rice plants compared to B3 plants (Table 1). These differential gene expression patterns between B3 and Yongning red rice likely contribute to the stronger cold tolerance observed in Yongning red rice.
3.7. Identification of Genes Associated with Cold Tolerance
To investigate the mechanism of rice’s resistance to low temperatures, sequencing data from Yongning red rice and B3 before and after low-temperature treatment were compared. Venn diagrams were used to analyze the potential genes related to low-temperature resistance in the DEMs of two rice genotypes, with a fold change (FC) ≥ 2 and p < 0.05. In Yongning red rice, 1551 unique DEMs were downregulated, and 2481 unique DEMs were upregulated after low-temperature treatment. In contrast, in the B3 plants, 1836 unique DEMs were downregulated, and 1754 unique DEMs were upregulated after treatment (Figure 5A,B). To better understand the function of these DEMs, GO enrichment analyses were carried out. In terms of biological processes, the downregulated genes in Yongning red rice and the upregulated genes in B3 plants were both enriched in processes such as pollen tube growth, defense response to fungus, defense response to bacteria, single-organism cellular process, response to chitin, regulation of plant-type hypersensitive response, response to cadmium ion, positive regulation of transcription, DNA-templated transcription, and protein targeting to membrane (Figure S2A,D). The upregulated genes in Yongning red rice and the downregulated genes in B3 were both enriched in the single-organism cellular process and the response to cadmium ion (Figure S2B,C). Among the molecular function GO terms, the most contrasting terms between the two rice varieties included quercetin 4′-O-glucosyltransferase activity, protein binding, peptide receptor activity, and ubiquitin protein ligase binding (Figure S2). Additionally, nine cellular components—apoplast, plasma membrane, Golgi apparatus, vacuole, chloroplast, nucleus, intracellular organelle part, plasmodesma, and plasma membrane—were enriched, indicating that these cellular components play a role in the low-temperature response (Figure S2).
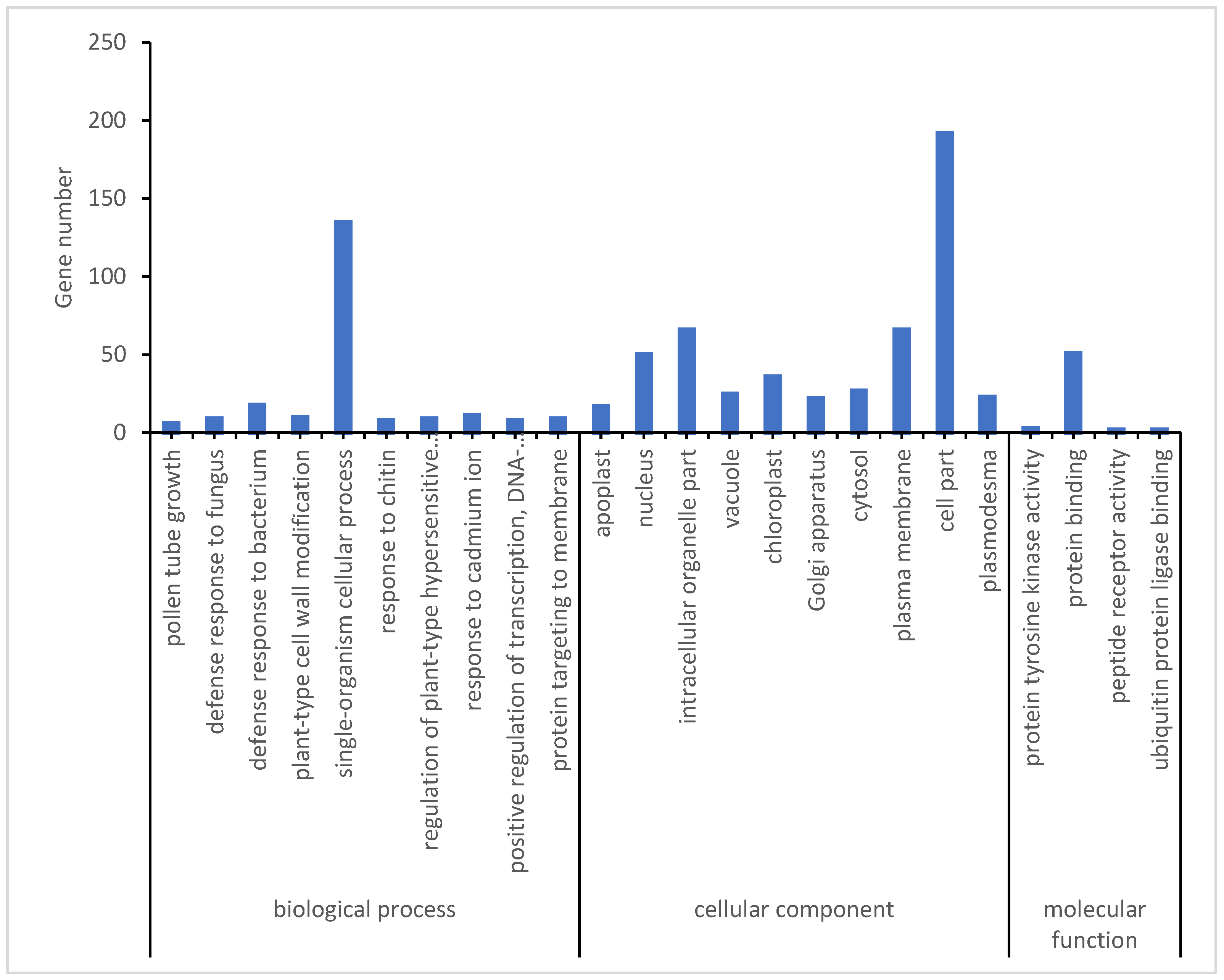
To further narrow down potential genes related to the low-temperature response, the opposite expression DEMs during the early and late stages of low-temperature treatment in two rice varieties was assessed. A total of 218 DEMs were identified in both Yongning red rice and B3 plants after low-temperature treatment, with 87 DEMs being downregulated in Yongning red rice plants and upregulated in B3, while 131 DEMs were upregulated in Yongning red rice and downregulated in B3 plants (Figure 5). GO enrichment analysis showed that these resistance-related genes were enriched in biological processes such as defense response to fungus, defense response to bacteria, plant-type cell wall modification, a single-organism cellular process, the response to chitin, and the regulation of a plant-type hypersensitive response. Cellular component enrichment included apoplast, nucleus, vacuole, plasma membrane, and plasmodesma (Figure 6). Finally, 21 genes were differentially expressed in both the Yongning red rice and the B3 after low-temperature treatment and were considered potential genes related to low-temperature resistance (Table 2). Among these, 20 genes were significantly upregulated in Yongning red rice and downregulated in B3 after treatment, while a single gene was downregulated in Yongning red rice and upregulated in B3. These DEMs were predominantly enriched in processes such as cellular processes, single-organism cellular processes, plant-type cell wall modification, stress response, metabolic processes, biological regulation, and cell wall organization. They were also enriched in cellular components such as the cell part, intracellular organelle part, nucleus, plasma membrane, and plasmodesma (Table 2).
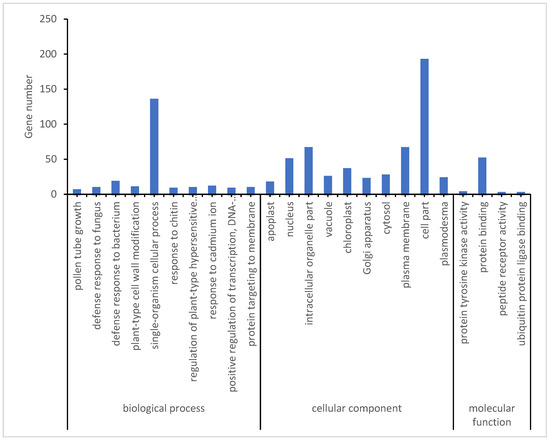
Figure 6.
GO (Gene Ontology) analysis of DEMs with opposite expression in two materials during early or late stages of low-temperature treatment.
Table 2.
Candidate genes related to low-temperature resistance exhibiting an opposite expression in the early and late stages of low-temperature treatment in two rice materials.
3.8. Integrated Analysis of miRNA and mRNA Expression Profiles
In most instances, miRNAs negatively regulate target mRNAs by repressing translation or degrading mRNAs [39]. For correlating the identified miRNAs with their target genes, the psRNA target tool was used with the parameters fold change ≥ 1.5, p < 0.05 [40]. A total of 20, 105, 70, and 114 DEMIs showed significant negative correlations with 72, 333, 230, and 347 target mRNAs in Re/R0, Rl/R0, Se/S0, and Sl/S0, respectively (Figure 2A).
To identify potential miRNA–mRNA pairs associated with low-temperature response, 82 DEMIs from Re/R0 or Rl/R0 (Figure 2B) and 19 DEMIs from different low-temperature treatment stages (Figure 3) were selected and negatively correlated with 416 target mRNAs (Table S4). These miRNAs target different mRNAs at each stage of low-temperature treatment. For example, miR167h-3p was downregulated in Se/S0 and upregulated in Rl/R0. This miRNA negatively correlated with eight upregulated target genes in Se/S0 and two downregulated target genes in Rl/R0, respectively. However, most of these targets showed similar expression trends in both Yongning red rice and B3 after low-temperature treatment (Table S4). When these miRNAs and their corresponding targets were excluded, there were 38 miRNAs corresponding to 39 target genes that were differentially expressed in Re/R0 or Rl/R0, or that exhibited an opposite expression in Yongning red rice and B3 after low-temperature treatment. These were selected as potential miRNA–mRNA candidates related to low-temperature resistance (Table 3).
Table 3.
The miRNA–mRNA interactions related to plant low-temperature response.
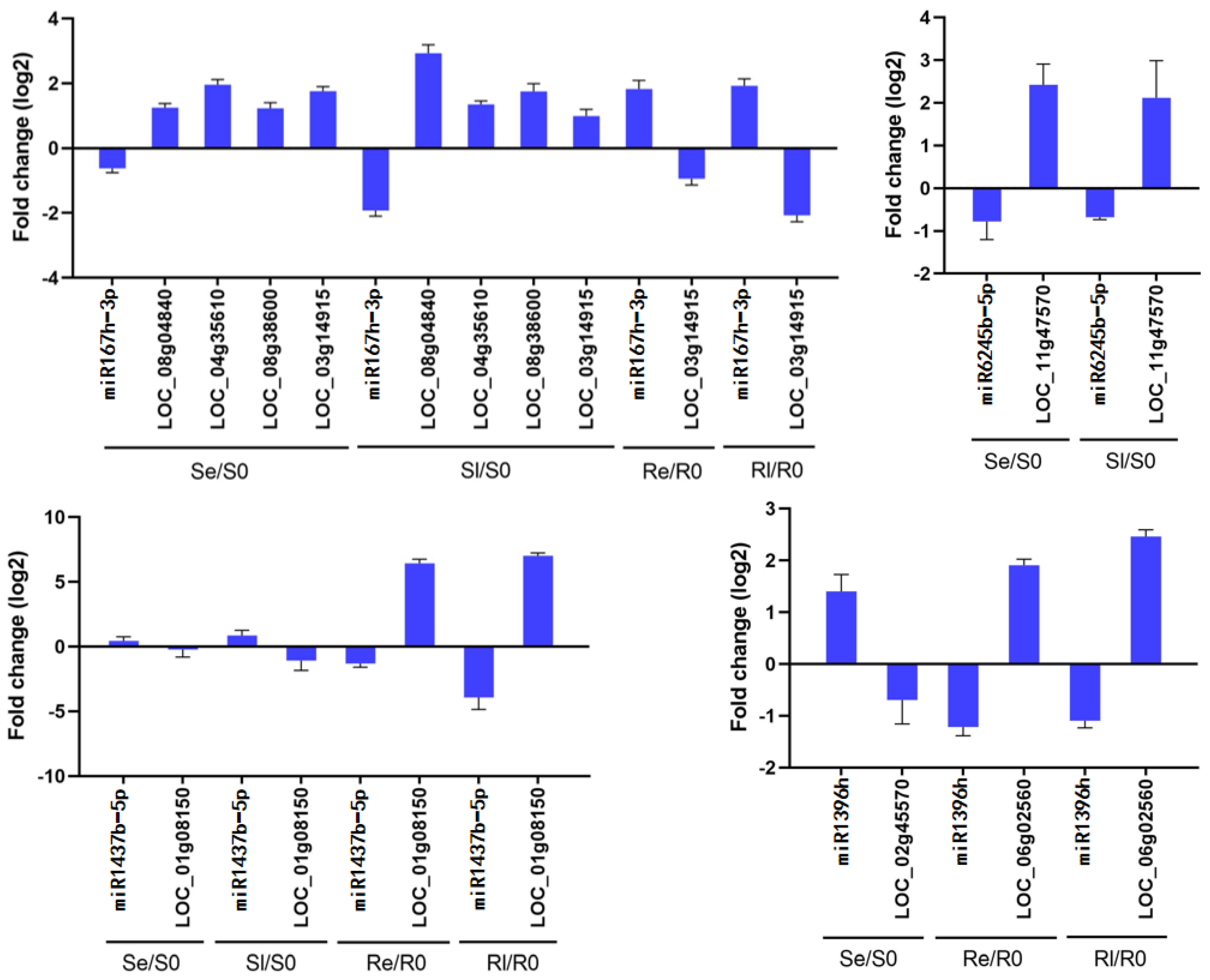
Four miRNAs and their target genes were chosen for qRT-PCR analysis to determine whether these miRNAs negatively regulate target gene expression. The results showed that miR1437b-5p negatively regulated LOC_Os01g08150 in both Yongning red rice and B3, miR167h-3p negatively regulated LOC_Os08g04840, LOC_Os04g35610, LOC_Os08g38600, and LOC_Os03g14915 in Se/S0 and Sl/S0, while it only negatively regulated LOC_Os03g14915 in Re/R0 and Rl/R0. miR6245b-5p negatively regulated LOC_Os11g47570 in both Se/S0 and Sl/S0, and miR396h negatively regulated LOC_Os02g45570 in Se/S0, while it negatively regulated LOC_Os06g02560 in Re/R0 and Rl/R0 (Figure 7). These results indicate that the qRT-PCR data are consistent with the sequencing results (Figure 7, Table 3).
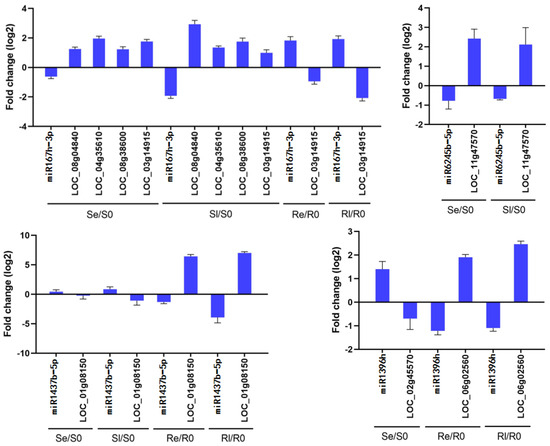
Figure 7.
Comparison of the expression patterns of miRNAs and their target genes. Data are means ± SD of three independent biological experiments.
4. Discussion
Few research reports have utilized a combined expression profile of miRNA and mRNA to analyze abiotic stress responses in plants, except for the study on the low-temperature regulation mechanism of the CTS-12 locus in common wild rice (Oryza rufipogon Griff.) [41]. This study, for the first time, reports the integrated analysis of mRNA and miRNA expression profiles in japonica rice under chilling stress, thereby improving our understanding of the regulatory mechanisms involving miRNA–mRNA interactions under chilling stress in rice.
By comparing the miRNA expression of Yongning red rice and B3 before and after low-temperature treatment, 194 DEMIs were identified (Figure 2A,B). To identify miRNAs associated with low-temperature responses, the DEMIs among four comparisons: Se/S0, Sl/S0, Re/R0, and Rl/R0 were analyzed using Venn diagrams (Figure 2B). A total of 40 DEMIs appeared in two comparisons (Re/R0 and Rl/R0), while 16 DEMIs appeared in both Se/S0 and Sl/S0 (Figure 2B). This suggests that more miRNAs are involved in low-temperature responses in Yongning red rice. Nineteen miRNAs exhibited opposite expression patterns in Yongning red rice and B3 plants after low-temperature treatment (Figure 3). Among them, miR1437b-5P, miR166c-5P, miR1861h, miR395y, miR396d, and miR396h displayed expression trends similar to those of Dongxiang common wild rice under low-temperature treatment [19], indicating that these miRNAs are likely to be involved in the defense response of rice to cold stress.
miR156, miR166, miR167_1, and miR395 have been reported to play roles in plant defense against abiotic stress and diseases. For example, miR156 participates in the regulation of drought stress by targeting StSPL9 in potato (Solanum tuberosum L.) [42]. miR166 promotes thermotolerance in Arabidopsis by regulating the expression of Heat stress transcription factors (HSFA1) [43]. miR395 is involved in the regulation of immunity in rice and the response to sulfate stress in Brassica juncea [36,44]. miR167_1 induces salicylic acid expression and stomatal closure by regulating target genes AUXIN RESPONSE FACTOR 6 (ARF6) and ARF8, positively modulating Arabidopsis defense against Pseudomonas syringae [33]. Members of the miR156, miR166, miR167_1, and miR395 families were upregulated in Yongning red rice and downregulated in the B3 plants (Figure 3), suggesting their involvement in rice’s low-temperature response. There are also research reports that miR166 can affect nutrient absorption, photosynthesis, and grain quality in rice [31]. miR167_1 negatively regulates the expression of auxin (IAA) under high osmotic stress, influencing Arabidopsis root architecture [45]. These reports suggest miRNAs might indirectly participate in the stress response to low temperatures by altering metabolic processes and hormone regulation.
To identify low-temperature resistance-related genes, RNA sequencing was carried out in Yongning red rice and B3 under low-temperature treatment. Transcriptome analysis revealed significant differences in the response of Yongning red rice and B3 to low-temperature treatment. Compared to B3 plants, Yongning red rice exhibited a stronger defense response to low temperature at both early and late stages, with more upregulated DEMs (fold change ≥ 2) detected in Yongning red rice. Furthermore, upregulated DEMs were significantly more abundant than downregulated ones in Yongning red rice, indicating a stronger activation of cold-responsive genes.
To explore the defense mechanism of Yongning red rice against cold stress, DEMs during the early and late stages of low-temperature treatment in both varieties were analyzed. A total of 218 DEMs were oppositely expressed, with most being upregulated in Yongning red rice, suggesting that these DEMs positively regulate rice’s resistance to low temperatures. Ultimately, 21 genes were selected as potential candidates for participation in low-temperature defense (Table 2). Most of these genes were upregulated in Yongning red rice and downregulated in B3 plants after low-temperature treatment, with the exception of LOC-Os07g45410, highlighting their crucial role in the rice’s cold stress response. The gene GRAIN INCOMPLETE FILLING 1 (GIF1), involved in sugar homeostasis mediated by cell wall invertase, plays an essential role in constitutive and induced defense responses [46]. Non-specific lipid transfer proteins (nsLTPs) contribute to various biological processes, including disease resistance [47,48,49,50,51], stress resistance [52,53,54,55,56,57], the regulation of germination and grain weight [58], and the formation of epidermal cutin and wax [59,60]. The EXPA gene family is also known to be vital in abiotic stresses, growth, and development [61,62,63,64].
Integrated expression analyses of miRNA and mRNA help identify functional miRNA–mRNA pairs related to rice resistance to cold stress. In this study, 82 specific DEMIs in Yongning red rice (Figure 2B) and 19 oppositely expressed miRNAs (Figure 3) corresponding to 416 target genes were identified under low-temperature treatment (Table S3). However, only 38 miRNAs corresponding to 39 target genes were potentially associated with rice’s low-temperature response (Table 3). For instance, miR169, known to negatively regulate immunity against Magnaporthe oryzae (rice blast fungus) [39], exhibited opposite expression patterns in Yongning red rice and B3 plants after low-temperature treatment (Figure 3). However, its target genes showed a similar expression pattern in both plant types, suggesting miR169 and its targets are not directly involved in cold stress response. This discrepancy could be attributed to several factors: (1) most target genes exhibited similar expression trends in both Yongning red rice and B3 plants after cold treatment, (2) the plant defense response to low temperature involves both systemic and local reactions, and many targets may not be expressed under these conditions, and (3) the criteria used for identifying DEMIs and DEMs may have overlooked key interactions.
5. Conclusions
In this study, we constructed 18 libraries of Yongning red rice and B3 plants before and after low-temperature treatment. These libraries were sequenced to identify mRNAs and miRNAs associated with cold resistance. Key miRNAs from families such as miR1437, miR156, miR166, miR1861, miR395, and miR396_2 were identified as playing significant roles in the rice’s resistance to low temperatures. DEMs involved in cold stress resistance were found to be related to plant cell wall modification, cellular processes, chitin response, and regulation of plant hypersensitive responses. Additionally, 38 miRNAs corresponding to 39 target genes were identified as potential miRNA–mRNA pairs linked to cold stress resistance. The integrated analysis of miRNAs and cold-responsive genes in rice provides a foundation for further research on the functions of miRNAs and their targets in low-temperature stress response, establishing a molecular basis for future studies on how plants adapt to cold environments.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/genes16010038/s1, Figure S1: Heatmap of DEMIs in Yongning red rice and B3 after low-temperature treatment; Figure S2: GO (Gene Ontology) analysis of DEMs in Yongning red rice and B3 after low-temperature treatment; Table S1: Primers used in qRT-PCR; Table S2: Summary of small RNA sequences data; Table S3: Summary of mRNA expression libraries; Table S4: The miRNA-mRNA pairs related to the resistance of rice to low temperatures.
Author Contributions
F.L., D.L. and L.C. planned and designed the research. F.L., M.Y., J.Z., X.Z., C.W., W.Z., D.L. and L.C. performed the experiments and conducted the fieldwork. F.L., M.Y., J.Z., D.L. and L.C. analyzed the data. F.L., X.Z., D.L. and L.C. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (31860108), Natural Science Foundation of Sichuan Province (2024NSFSC0317), the Open Project Program of Panxi Crops Research and Utilization Key Laboratory of Sichuan Province (XNFZ21015), The High-end Foreign Expert Program of the ‘Yunnan Talent Support Plan’ (No), the Major Science and Technology Projects of Yunnan (202202AE09002102), the Central Leading Local Science and Technology Development Project (202207AA110010).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the first author Fan Luo. The data are not publicly available due to privacy restrictions.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Shahzad, N.; Nabi, H.G.; Qiao, L.; Li, W. The molecular mechanism of cold-stress tolerance: Cold responsive genes and their mechanisms in rice (Oryza sativa L.). Biology 2024, 13, 442. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Tian, C.; Song, S.; Li, R. Insights on the enhancement of chilling tolerance in Rice through over-expression and knock-out studies of OsRBCS3. Plant Signal. Behav. 2024, 19, 2318514. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Fu, M.; Du, J.; Wang, M.; Zhang, S.; Li, S.; Chen, J.; Zha, W.; Li, C.; Liu, K.; et al. Identification of the cold-related genes COLD11 and OsCTS11 via BSA-seq and fine mapping at the rice seedling stage. Rice 2024, 17, 72. [Google Scholar] [CrossRef]
- Zuo, J.; Wei, C.; Liu, X.; Jiang, L.; Gao, J. Multifunctional transcription factor YABBY6 regulates morphogenesis, drought and cold stress responses in rice. Rice 2024, 17, 69. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, P.; Das, A.; Datta, S.; Banerjee, I.; Tripathy, S.; Chaudhuri, S. Understanding the early cold response mechanism in IR64 indica rice variety through comparative transcriptome analysis. BMC Genom. 2020, 21, 425. [Google Scholar] [CrossRef]
- Xia, H.; Yu, S.; Kong, D.; Xiong, J.; Ma, X.; Chen, L.; Luo, L. Temporal responses of conserved miRNAs to drought and their associations with drought tolerance and productivity in rice. BMC Genom. 2020, 21, 232. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O. Origin, biogenesis, and activity of plant microRNAs. Cell. 2009, 136, 669–687. [Google Scholar] [CrossRef]
- Dai, Y.; Feng, X.; Liu, Z.; Wang, M.; Zhou, Y.; Cui, L.; Wei, X.; Zhu, Z. miR1432 negatively regulates cold tolerance by targeting OsACAs. Plant Cell Environ. 2024, 47, 5443–5456. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Zhang, X.; Li, W.; Pan, D.; Ma, B.; Duan, X.; Dong, C.; Wang, L.; Zhao, M.; Zhao, S.; et al. A resource for functional investigation of miRNAs in rice responses to viral infection. Plant Biotechnol. J. 2024, 22, 3380–3382. [Google Scholar] [CrossRef]
- Sunkar, R.; Zhu, J.K. Novel and stress-regulated microRNAs and other small RNAs from Arabidopsis. Plant Cell 2004, 16, 2001–2019. [Google Scholar] [CrossRef] [PubMed]
- Ucar, S.; Yaprak, E.; Yigider, E.; Kasapoglu, A.G.; Oner, B.M.; Ilhan, E.; Ciltas, A.; Yildirim, E.; Aydin, M. Genome-wide analysis of miR172-mediated response to heavy metal stress in chickpea (Cicer arietinum L.): Physiological, biochemical, and molecular insights. BMC Plant Biol. 2024, 24, 1063. [Google Scholar] [CrossRef]
- Luo, G.; Li, L.; Yang, X.; Yu, Y.; Gao, L.; Mo, B.; Chen, X.; Liu, L. MicroRNA1432 regulates rice drought stress tolerance by targeting the CALMODULIN-LIKE2 gene. Plant Physiol. 2024, 195, 1954–1968. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Shen, Y.; Yang, J.; Cai, X.; Li, H.; Zhu, Y.; Jia, B.; Sun, X. miR535 negatively regulates cold tolerance in rice. Mol. Breed. 2020, 40, 14. [Google Scholar] [CrossRef]
- Xie, S.; Li, H.; Lu, J.; Li, J.; Song, Z.; Jiang, H. A novel member of miR169 family negatively regulates maize resistance against Bipolaris maydis. Plant Dis. 2024, 108, 3518–3526. [Google Scholar] [CrossRef] [PubMed]
- Gaddam, S.R.; Sharma, A.; Bhatia, C.; Trivedi, P.K. A network comprising ELONGATED HYPOCOTYL 5, microRNA397b, and auxin-associated factors regulates root hair growth in Arabidopsis. Plant Physiol. 2024, 196, 1460–1474. [Google Scholar] [CrossRef]
- Pawłasek, N.; Sokołowska, A.; Koter, M.; Oracz, K. The interaction between miR165/166 and miR160 regulates Arabidopsis thaliana seed size, weight, and number in a ROS-dependent manner. Planta. 2024, 260, 72. [Google Scholar] [CrossRef]
- Xie, H.; Zhu, M.; Yu, Y.; Zeng, X.; Tang, G.; Duan, Y.; Wang, J.; Yu, Y. Comparative transcriptome analysis of the cold resistance of the sterile rice line 33S. PLoS ONE 2022, 17, e0261822. [Google Scholar] [CrossRef] [PubMed]
- Fan, M.; Yang, K.; Zhou, R.; Liu, Q.; Guo, X.; Sun, Y. Temporal transcriptome profiling reveals candidate genes involved in cold acclimation of Camellia japonica (Naidong). Plant Physiol. Biochem. 2021, 167, 795–805. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Shi, W.; Ma, X.; Zhao, J.; Wang, S.; Tan, L.; Sun, C.; Liu, F. Identification of microRNAs responding to cold stress in Dongxiang common wild rice. Genome. 2019, 62, 635–642. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Yang, J.; Cai, X.; Shen, Y.; Cui, N.; Zhu, Y.; Jia, B.; Sun, X. The opposite roles of OsmiR408 in cold and drought stress responses in Oryza sativa. Mol. Breed. 2018, 38, 120. [Google Scholar] [CrossRef]
- Božić, M.; Micić, D.I.; Delić, N.; Nikolić, A. Maize miRNAs and their putative target genes involved in chilling stress response in 5-day old seedlings. BMC Genom. 2024, 25, 479. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Allen, E.; Xie, Z.; Gustafson, A.M.; Carrington, J.C. MicroRNA-directed phasing during trans-acting siRNA biogenesis in plants. Cell 2005, 121, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Hua, L.; Zhang, Z.; Yang, B.; Li, W. CRISPR-induced miRNA156-recognition element mutations in TaSPL13 improve multiple agronomic traits in wheat. Plant Biotechnol. J. 2023, 21, 536–548. [Google Scholar] [CrossRef] [PubMed]
- Ferigolo, L.F.; Vicente, M.H.; Correa, J.P.O.; Barrera-Rojas, C.H.; Silva, E.M.; Silva, G.F.F.; Carvalho, A.; Peres, L.E.P.; Ambrosano, G.B.; Margarido, G.R.A.; et al. Gibberellin and the miRNA156-targeted SlSBPs synergistically regulate tomato floral meristem determinacy and ovary patterning. Development 2023, 150, dev201961. [Google Scholar] [CrossRef]
- Yan, B.; Li, F.; Ma, Q.; Shen, T.; Jiang, J.; Li, H. The miR156-SPL4/SPL9 module regulates leaf and lateral branch development in Betula platyphylla. Plant Sci. 2023, 338, 111869. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Song, Z.; Xie, Y.; Cheng, H.; Yan, H.; Sun, F.; Liu, H.; Shen, J.; Li, L.; He, X.; et al. High temperature inhibits vascular development via the PIF4-miR166-HB15 module in Arabidopsis. Curr. Biol. 2023, 33, 3203–3214. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, S.; Zhang, D.; Li, X.; Gao, Z.; Jiang, Z. The miR166-mRNA network regulates vascular tissue differentiation in Moso bamboo. Front. Genet. 2022, 13, 893956. [Google Scholar] [CrossRef]
- Iwamoto, M. In-frame editing of transcription factor gene RDD1 to suppress miR166 recognition influences nutrient uptake, photosynthesis, and grain quality in rice. Sci. Rep. 2022, 12, 10795. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Huang, S.; Xie, H. Advances in the regulation of plant development and stress response by miR167. Front. Biosci. (Landmark Ed.) 2021, 26, 655–665. [Google Scholar] [CrossRef] [PubMed]
- Caruana, J.C.; Dhar, N.; Raina, R. Overexpression of Arabidopsis microRNA167 induces salicylic acid-dependent defense against Pseudomonas syringae through the regulation of its targets ARF6 and ARF8. Plant Direct. 2020, 4, e00270. [Google Scholar] [CrossRef]
- Rao, S.; Gupta, A.; Bansal, C.; Sorin, C.; Crespi, M.; Mathur, S. A conserved HSF:miR169:NF-YA loop involved in tomato and Arabidopsis heat stress tolerance. Plant J. 2022, 112, 7–26. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhao, S.L.; Li, J.L.; Hu, X.H.; Wang, H.; Cao, X.L.; Xu, Y.J.; Zhao, Z.X.; Xiao, Z.Y.; Yang, N.; et al. Osa-miR169 negatively regulates rice immunity against the blast fungus magnaporthe oryzae. Front. Plant Sci. 2017, 8, 2. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Hui, S.; Lv, Y.; Zhang, M.; Chen, D.; Tian, J.; Zhang, H.; Liu, H.; Cao, J.; Xie, W.; et al. miR395-regulated sulfate metabolism exploits pathogen sensitivity to sulfate to boost immunity in rice. Mol. Plant 2022, 15, 671–688. [Google Scholar] [CrossRef]
- Wang, L.; Hou, J.; Xu, H.; Zhang, Y.; Huang, R.; Wang, D.; He, X.Q. The PtoTCP20-miR396d-PtoGRF15 module regulates secondary vascular development in Populus. Plant Commun. 2023, 4, 100494. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Shen, J.; Zhang, C.; Guo, Q.; Liang, H.; Hou, X. Characterization and bioinformatics analysis of ptc-miR396g-5p in response to drought stress of Paeonia ostii. Noncoding RNA Res. 2022, 7, 150–158. [Google Scholar] [CrossRef]
- Ma, Z.; Wang, J.; Li, C. Research progress on miRNAs and artificial miRNAs in insect and disease resistance and breeding in plants. Genes 2024, 15, 1200. [Google Scholar] [CrossRef]
- Dai, X.; Zhuang, Z.; Zhao, P.X. psRNATarget: A plant small RNA target analysis server (2017 release). Nucleic Acids Res. 2018, 46, W49–W54. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Xiao, W.; Sun, J.; Chen, M.; Ma, M.; Cao, Y.; Cen, W.; Li, R.; Luo, J. An integration of microRNA and transcriptome sequencing analysis reveal regulatory roles of miRNAs in response to chilling stress in wild rice. Plants 2022, 11, 977. [Google Scholar] [CrossRef] [PubMed]
- Pescador-Dionisio, S.; Robles-Fort, A.; Parisi, B.; García-Robles, I.; Bassolino, L.; Mandolino, G.; Real, M.D.; Rausell, C. Contribution of the regulatory miR156-SPL9 module to the drought stress response in pigmented potato (Solanum tuberosum L.). Plant Physiol. Biochem. 2024, 217, 109195. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Cao, Y.; Zhang, J.; Zhu, C.; Tang, G.; Yan, J. The miR165/166-PHABULOSA module promotes thermotolerance by transcriptionally and posttranslationally regulating HSFA1. Plant Cell. 2023, 35, 2952–2971. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, E.; Pokhriyal, E.; Jain, A.; Lal, M.; Khari, M.; Jalan, K.; Das, S. The non-canonically organized members of MIR395 gene family in Brassica juncea are associated with developmentally regulated, sulfate-stress responsive bidirectional promoters that exhibit orientation-dependent differential transcriptional activity. Plant Sci. 2024, 348, 112214. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, N.; Wang, H.; Kasahara, H.; Liu, J.; Macpherson, C.; Machida, Y.; Kamiya, Y.; Hannah, M.A.; Chua, N.H. IAA-Ala Resistant3, an evolutionarily conserved target of miR167, mediates Arabidopsis root architecture changes during high osmotic stress. Plant Cell 2012, 24, 3590–3602. [Google Scholar] [CrossRef]
- Sun, L.; Yang, D.L.; Kong, Y.; Chen, Y.; Li, X.Z.; Zeng, L.J.; Li, Q.; Wang, E.T.; He, Z.H. Sugar homeostasis mediated by cell wall invertase GRAIN INCOMPLETE FILLING 1 (GIF1) plays a role in pre-existing and induced defence in rice. Mol. Plant Pathol. 2014, 15, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Dou, W.; Song, H.; Deng, H.; Tian, Z.; Chen, R.; Liu, Z.; Jiao, Z.; Akhberdi, O. Insights into the functional mechanism of the non-specific lipid transfer protein nsLTP in Kalanchoe fedtschenkoi (Lavender scallops). Protein Expr. Purif. 2025, 226, 106607. [Google Scholar] [CrossRef] [PubMed]
- Shang, K.; Wang, C.; Wang, X.; Wang, Y.; Xu, K.; Zhou, S.; Liu, H.; Zhu, X.; Zhu, C. Non-specific lipid transfer protein StLTP6 promotes virus infection by inhibiting jasmonic acid signalling pathway in response to PVS TGB1. Plant Cell Environ. 2024, 4, 100494. [Google Scholar] [CrossRef]
- Li, H.; Liu, Y.; Gao, W.; Zhu, J.; Zhang, H.; Wang, Z.; Liu, C.; Li, X. Genome-wide characterization of small secreted peptides in Nicotiana tabacum and functional assessment of NtLTP25 in plant immunity. Physiol. Plant. 2024, 176, e14436. [Google Scholar] [CrossRef]
- Zhu, F.; Cao, M.Y.; Zhu, P.X.; Zhang, Q.P.; Lam, H.M. Non-specific LIPID TRANSFER PROTEIN 1 enhances immunity against tobacco mosaic virus in Nicotiana benthamiana. J. Exp. Bot. 2023, 74, 5236–5254. [Google Scholar] [CrossRef]
- Bvindi, C.; Howe, K.; Wang, Y.; Mullen, R.T.; Rogan, C.J.; Anderson, J.C.; Goyer, A. Potato non-specific lipid transfer protein StnsLTPI.33 is associated with the production of reactive oxygen species, plant growth, and susceptibility to Alternaria solani. Plants 2023, 12, 3129. [Google Scholar] [CrossRef] [PubMed]
- Dhar, N.; Caruana, J.; Erdem, I.; Raina, R. An Arabidopsis DISEASE RELATED NONSPECIFIC LIPID TRANSFER PROTEIN 1 is required for resistance against various phytopathogens and tolerance to salt stress. Gene 2020, 753, 144802. [Google Scholar] [CrossRef]
- Peng, C.N.; Xing, Y.; Wang, Q.B.; Wang, C.C.; Zhang, X.Y.; Chen, D.Y.; Song, Y.Z.; Zhu, C.X. Expression of tobacco Lipid Transfer Protein NtLTP4 enhances tolerance to abiotic and biotic stresses in transgenic potato lines. Potato Res. 2022, 65, 631–647. [Google Scholar] [CrossRef]
- Xiao, Y.; Xiao, C.; He, X.; Yang, X.; Tong, Z.; Wang, Z.; Sun, Z.; Qiu, W. A novel non-specific lipid transfer protein gene, CmnsLTP6.9, enhanced osmotic and drought tolerance by regulating ROS scavenging and remodeling lipid profiles in chinese chestnut (Castanea mollissima Blume). Plants 2023, 12, 3916. [Google Scholar] [CrossRef]
- Yang, Y.; Song, H.; Yao, P.; Zhang, S.; Jia, H.; Ye, X. NtLTPI.38, a plasma membrane-localized protein, mediates lipid metabolism and salt tolerance in Nicotiana tabacum. Int. J. Biol. Macromol. 2023, 242 Pt 2, 125007. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Yao, P.; Zhang, S.; Jia, H.; Yang, Y.; Liu, L. A non-specific lipid transfer protein, NtLTPI.38, positively mediates heat tolerance by regulating photosynthetic ability and antioxidant capacity in tobacco. Plant Physiol. Biochem. 2023, 200, 107791. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Wang, S.; Qin, J.; Sun, C.; Liu, F. The lipid transfer protein OsLTPL159 is involved in cold tolerance at the early seedling stage in rice. Plant Biotechnol. J. 2020, 18, 756–769. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Guo, L.; Cui, Y.; Yan, X.; Ouyang, J.; Li, S. Lipid transfer protein, OsLTPL18, is essential for grain weight and seed germination in rice. Gene 2023, 883, 147671. [Google Scholar] [CrossRef]
- DeBono, A.; Yeats, T.H.; Rose, J.K.; Bird, D.; Jetter, R.; Kunst, L.; Samuels, L. Arabidopsis LTPG is a glycosylphosphatidylinositol-anchored lipid transfer protein required for export of lipids to the plant surface. Plant Cell 2009, 21, 1230–1238. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.B.; Go, Y.S.; Bae, H.J.; Park, J.H.; Cho, S.H.; Cho, H.J.; Lee, D.S.; Park, O.K.; Hwang, I.; Suh, M.C. Disruption of glycosylphosphatidylinositol-anchored lipid transfer protein gene altered cuticular lipid composition, increased plastoglobules, and enhanced susceptibility to infection by the fungal pathogen Alternaria brassicicola. Plant Physiol. 2009, 150, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ma, J.; He, F.; Wang, L.; Zhang, T.; Liu, D.; Xu, Y.; Li, F.; Feng, X. A study on the functional identification of overexpressing winter wheat expansin gene TaEXPA7-B in rice under salt stress. Int. J. Mol. Sci. 2024, 25, 7707. [Google Scholar] [CrossRef] [PubMed]
- Ribas, A.F.; Silva, N.V.E.; Santos, T.B.D.; Abrantes, F.L.; Custódio, C.C.; Machado-Neto, N.B.; Vieira, L.G.E. Regulation of α-expansins genes in Arabidopsis thaliana seeds during post-osmopriming germination. Physiol. Mol. Biol. Plants 2019, 25, 511–522. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Luo, Y.; Wang, G.; Feng, C.; Li, H. Genome-wide identification of expansin genes in Brachypodium distachyon and functional characterization of BdEXPA27. Plant Sci. 2020, 296, 110490. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Zhou, F.; Chen, Y.; Wu, H.; Yin, T. Genome-wide analysis of the expansin gene family in Populus and characterization of expression changes in response to phytohormone (abscisic acid) and abiotic (low-temperature) stresses. Int. J. Mol. Sci. 2023, 24, 7759. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).