
Foliar Potassium Sulfate Application Improved Photosynthetic Characteristics, Water Relations and Seedling Growth of Drought-Stressed Maize

 , ,
, ,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Estimation of Relative Water Content (%)
- FW = Fresh weight of leaves
- DW = Dry weight of leaves
- TW = Turgid weight of leaves.
2.2. Estimation of Gas Exchange Parameters
2.3. Estimation of Chlorophyll Content
2.4. Estimation of Proline (mg g−1)
2.5. Morphological Traits
2.6. Statistical Analysis
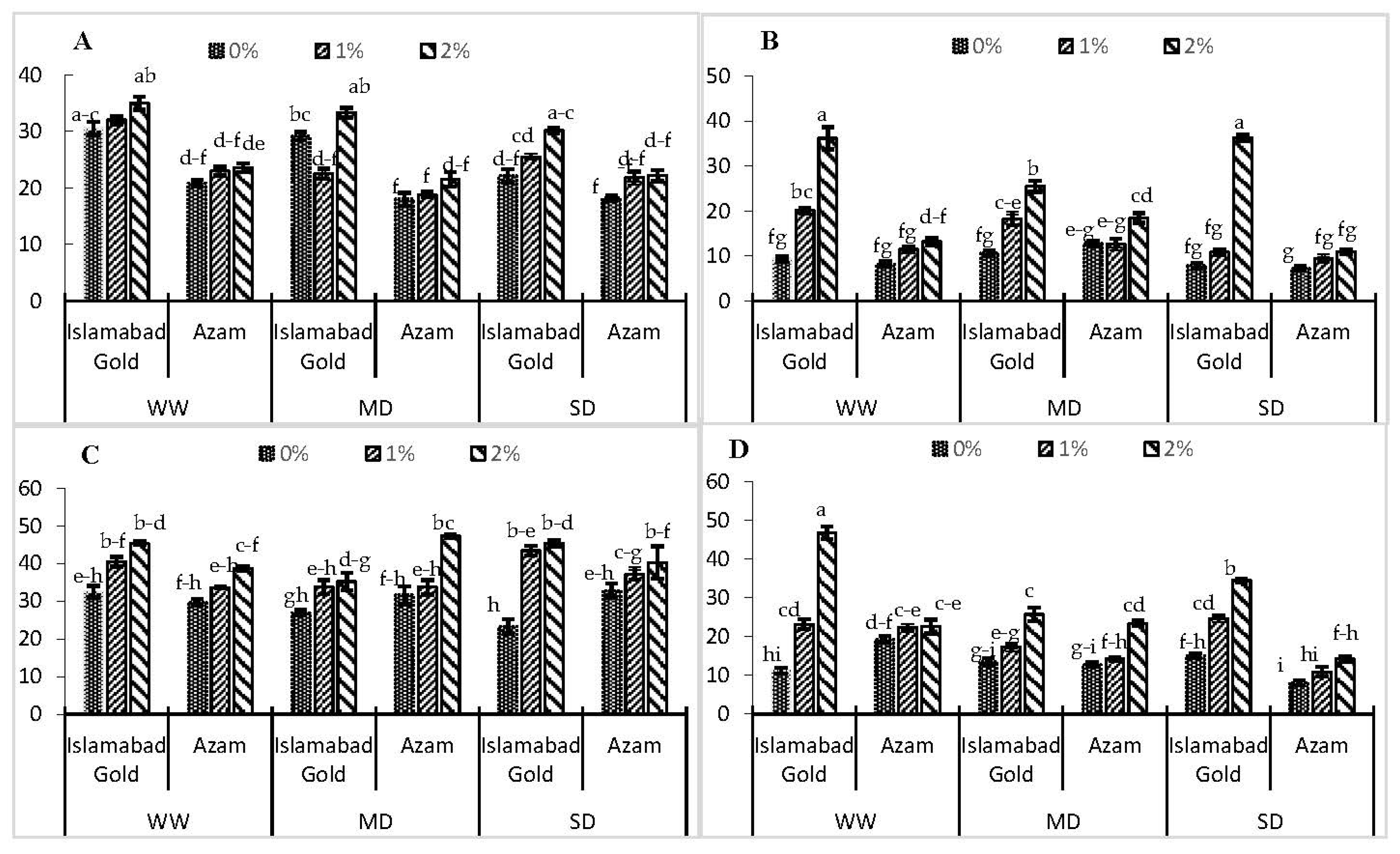
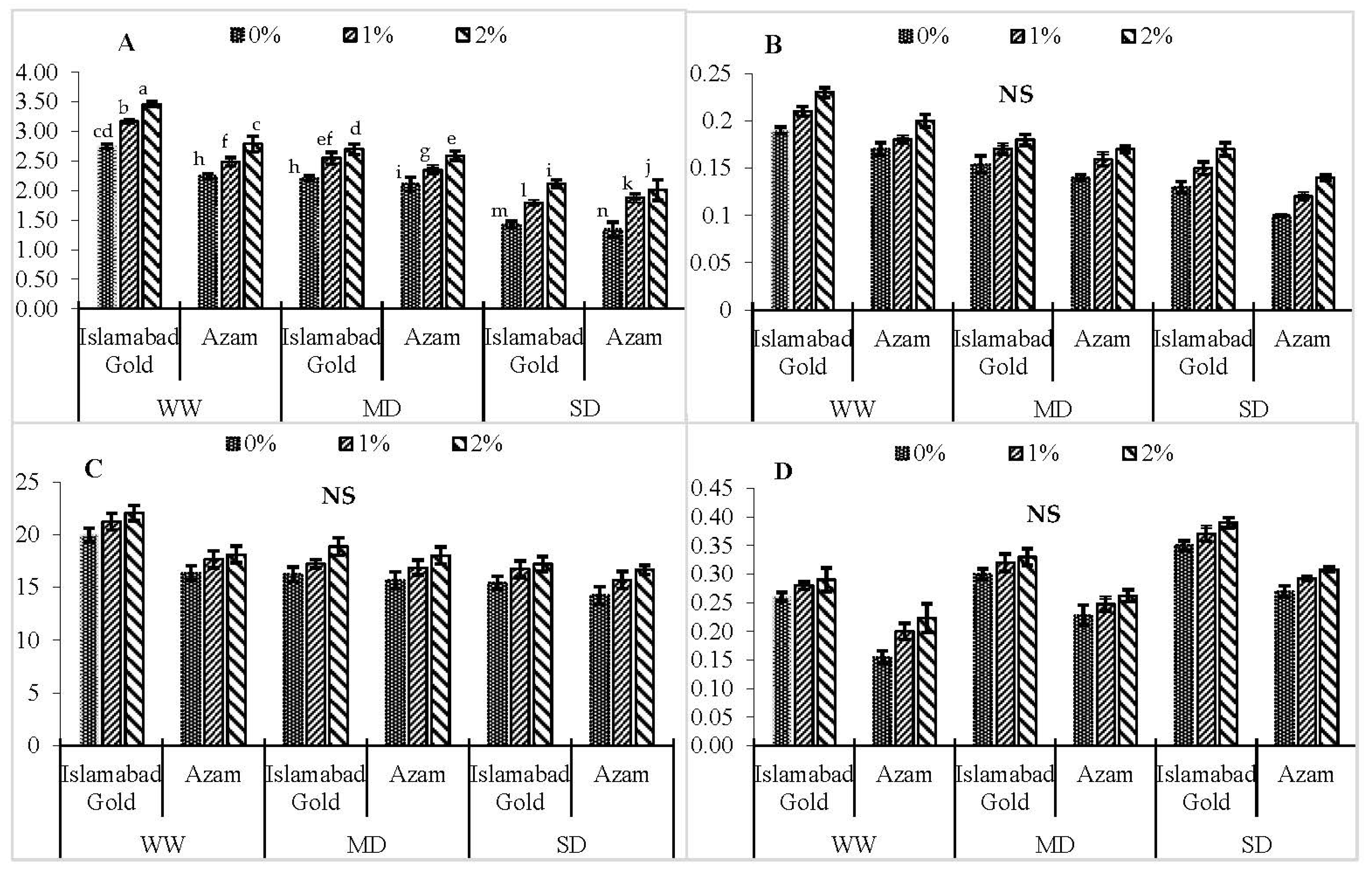
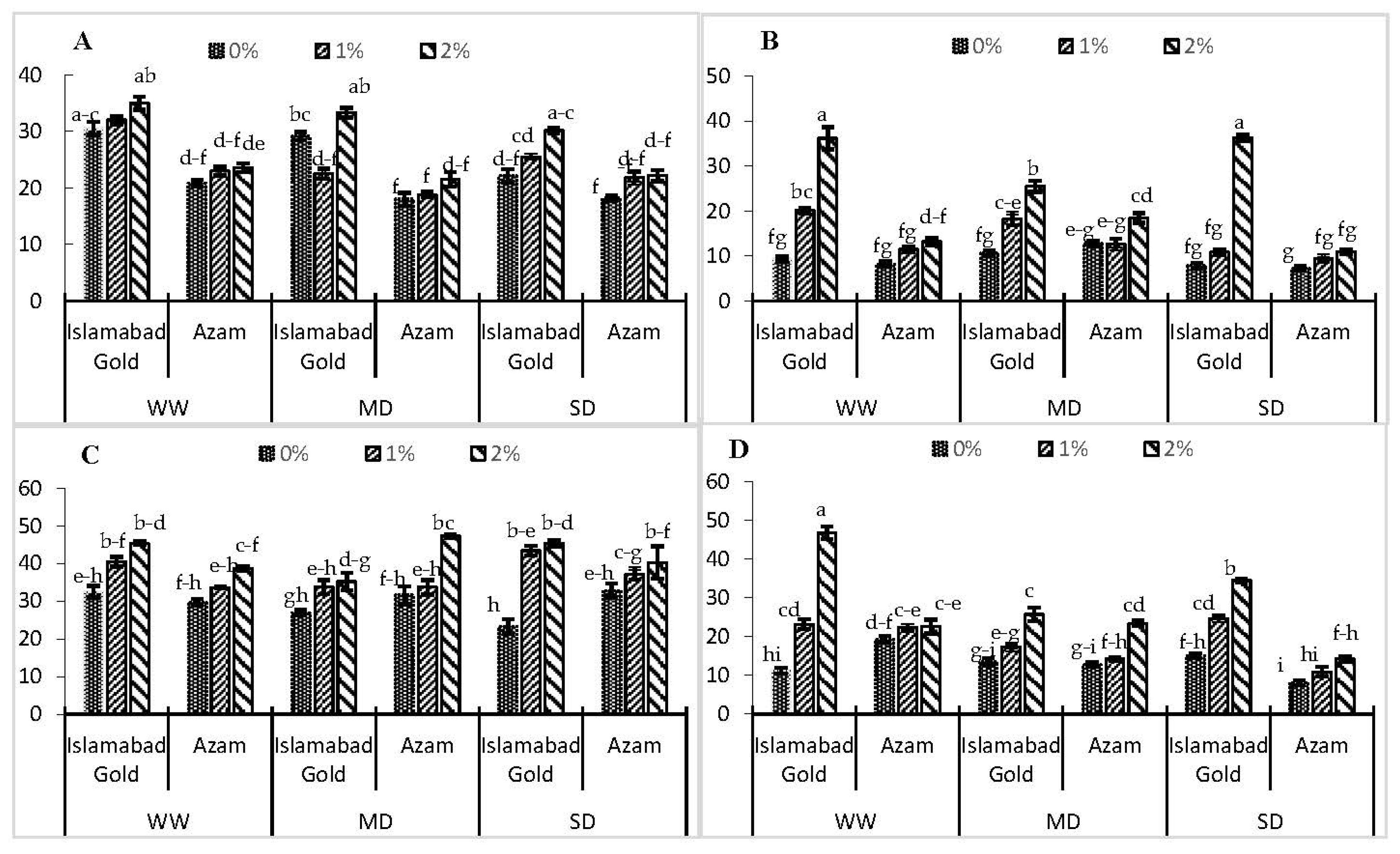
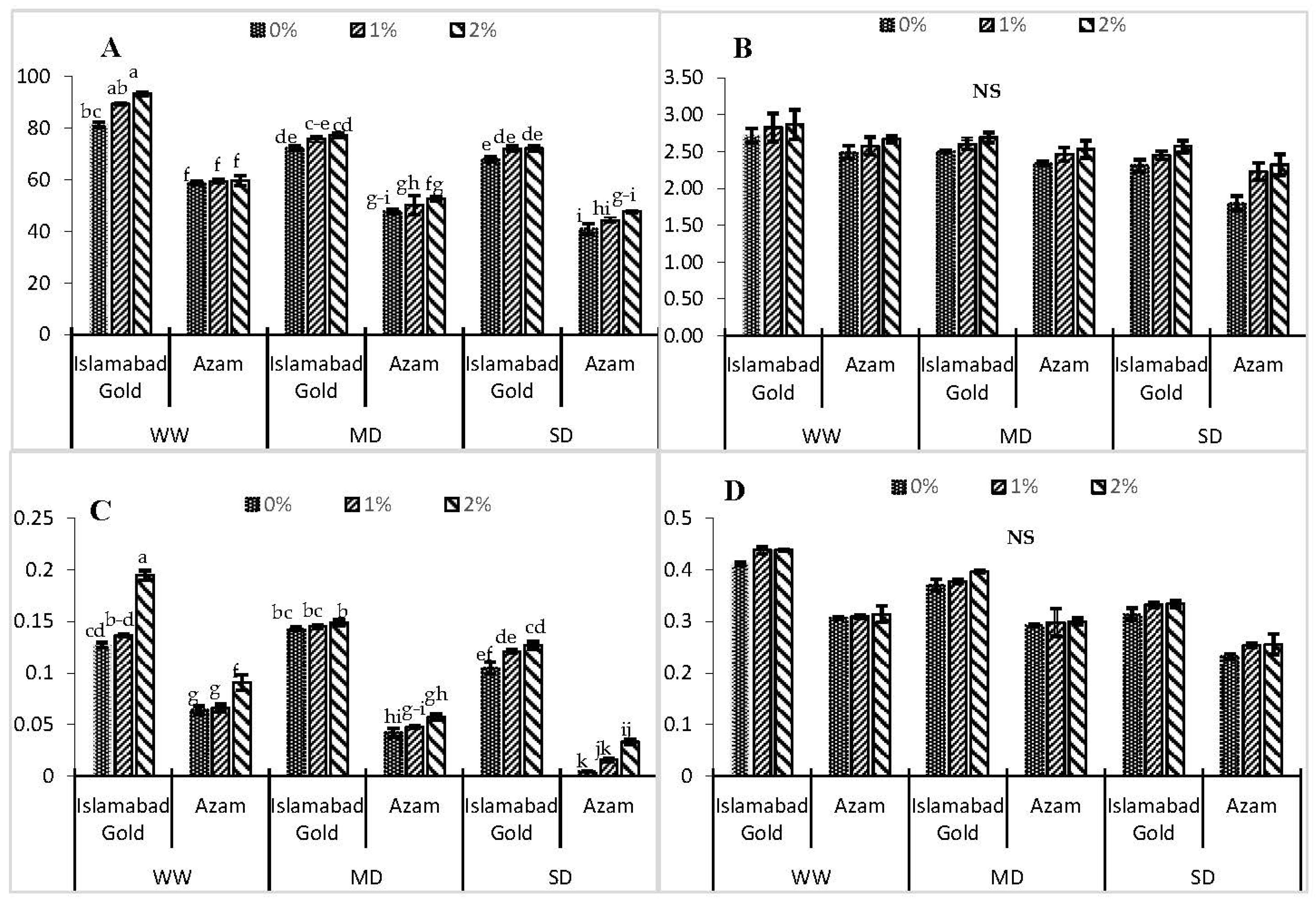
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ghosh, D.; Brahmachari, K.; Skalicky, M.; Hossain, A.; Sarkar, S.; Dinda, N.K.; Das, A.; Pramanick, B.; Moulick, D.; Brestic, M.; et al. Nutrients Supplementation through Organic Manures Influence the Growth of Weeds and Maize Productivity. Molecules 2020, 25, 4924. [Google Scholar] [CrossRef]
- Maqsood, Q.; Rana, N.A.; Iqbal, M.A.; Serap, K.A.; Asif, I.; Ayman, S. Overviewing of Weed Management Practices to Reduce Weed Seed Bank and to Increase Maize Yield. Planta Daninha 2020, 38, e020199716. [Google Scholar] [CrossRef]
- Khaliq, A.; Iqbal, M.A.; Zafar, M.; Anis, G. Appraising Economic Dimension of Maize Production under Coherent Fertilization in Azad Kashmir, Pakistan. Cust. Agroneg. 2019, 15, 243–253. [Google Scholar]
- Sariyev, A.; Barutcular, C.; Acar, M.; Hossain, A.; EL-Sabagh, A. Sub-surface Drip Irrigation in Associated with H2O2 Improved the Productivity of Maize under Clay-rich Soil of Adana, Turkey. Phyton Int. J. Exp. Bot. 2020, 89, 519–528. [Google Scholar] [CrossRef]
- De Jonge, K.C.; Taghvaeian, S.; Trout, T.J.; Comas, L.H. Comparison of Canopy Temperature-based Water Stress Indices for Maize. Agric. Water Manag. 2015, 156, 51–62. [Google Scholar] [CrossRef]
- Zalud, Z.; Hlavinka, P.; Prokes, K.; Semeradova, D.; Jan, B.; Trnka, M. Impacts of Water Availability and Drought on Maize Yield–A Comparison of 16 Indicators. Agric. Water. Manag. 2017, 188, 126–135. [Google Scholar] [CrossRef]
- ELSabagh, A.; Hossain, A.; Iqbal, M.A.; Barutçular, C.; Islam, M.S.; Çiğ, F.; Erman, M.; Sytar, O.; Brestic, M.; Wasaya, A.; et al. Maize Adaptability to Heat Stress under Changing Climate. In Plant Stress Physiology; IntechOpen: London, UK, 2020. [Google Scholar]
- Iqbal, A.; Iqbal, M.A.; Amir, I.; Zubair, A.; Muhammad, M.; Zahoor, A.; Muhammad, U.F.; Ghulam, A.; Faisal, M. Boosting Forage Yield and Quality of Maize (Zea mays L.) with Multi-species Bacterial Inoculation in Pakistan. Phyton Int. J. Exp. Bot. 2017, 86, 84–88. [Google Scholar]
- Daryanto, S.; Wang, L.; Jacinthe, P.A. Global Synthesis of Drought Effects on Maize and Wheat Production. PLoS ONE 2016, 11, e0156362. [Google Scholar] [CrossRef] [PubMed]
- Molla, S.H.; Nakasathien, S.; Ali, A.; Khan, A.; Alam, R.; Hossain, A.; Farooq, M.; El Sabagh, A. Influence of Nitrogen Application on Dry Biomass Allocation and Translocation in Two Maize Varieties under Short Pre-anthesis and Prolonged Bracketing Flowering Periods of Drought. Arch. Agron. Soil Sci. 2019, 65, 928–944. [Google Scholar] [CrossRef]
- Wasaya, A.; Zhang, X.; Fang, Q.; Zongzheng, Y. Root Phenotyping for Drought Tolerance: A Review. Agronomy 2018, 8, 241. [Google Scholar] [CrossRef] [Green Version]
- Maqsood, M.; Shehzad, M.A.; Ahmad, S.; Mushtaq, S. Performance of Wheat (Triticum aestivum L.) Genotypes Associated with Agronomical Traits under Water Stress Conditions. Asian J. Pharm. Biol. Res. 2012, 2, 45–50. [Google Scholar]
- Selvakumar, G.; Panneerselvam, P.; Ganeshamurthy, A.N. Bacterial Mediated Alleviation of Abiotic Stress in Crops. In Bacteria in Agrobiology: Stress Management; Maheshwari, D.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 205–224. [Google Scholar]
- Flexas, J.; Bota, J.; Loreto, F.; Cornic, G.; Sharkey, T.D. Diffusive and Metabolic Limitations to Photosynthesis under Drought and Salinity in C3 Plants. Plant Biol. 2004, 6, 269–279. [Google Scholar] [CrossRef]
- Samarah, N.H.; Alqudah, A.M.; Amayreh, J.A.; McAndrews, G.M. The Effect of Late-terminal Drought Stress on Yield Components of Four Barley Cultivars. J. Agron. Crop Sci. 2009, 195, 427–441. [Google Scholar] [CrossRef]
- Majid, M.A.; Islam, M.S.; EL Sabagh, A.; Hasan, M.K.; Barutcular, C.; Ratnasekera, D.; Islam, M.S. Evaluation of Growth and Yield Traits in Corn under Irrigation Regimes in Sub-tropical Climate. J. Exp. Biol. Agric. Sci. 2017, 5, 143–150. [Google Scholar]
- Ge, T.; Sui, F.; Bai, L.; Tong, C.; Sun, N. Effects of Water Stress on Growth, Biomass Partitioning, and Water-use Efficiency in Summer Maize (Zea mays L.) throughout the Growth Cycle. Acta Physiol. Plant. 2012, 34, 1043–1053. [Google Scholar] [CrossRef]
- Wasaya, A.; Abbas, T.; Yasir, T.A.; Sarwar, N.; Aziz, A.; Javaid, M.M.; Akram, S. Mitigating Drought Stress in Sunflower (Helianthus annuus L.) Through Exogenous Application of β-Aminobutyric Acid. J. Soil Sci. Plant Nutr. 2021. [Google Scholar] [CrossRef]
- Sehgal, A.; Sita, K.; Siddique, K.H.; Kumar, R.; Bhogireddy, S.; Varshney, R.K.; HanumanthaRao, B.; Nair, R.M.; Prasad, P.V.; Nayyar, H. Drought or/and heat-stress effects on seed filling in food crops: Impacts on functional biochemistry, seed yields, and nutritional quality. Front. Plant Sci. 2018, 9, 1705. [Google Scholar] [CrossRef] [Green Version]
- Fahad, S.; Ullah, A.; Ali, U.; Ali, E.; Saud, S.; Hakeem, K.; Alharby, H.; Sabagh, A.; Barutcular, C.; Kamran, M.; et al. Drought Tolerance in Plants Role of Phytohormones and Scavenging System of ROS. In Plant Tolerance to Environmental Stress Role of Phytoprotectants; Hasanuzzaman, M., Fujita, M., Oku, H., Tofazzal Islam, M., Eds.; CRC Press: Boca Raton, FL, USA, 2019; pp. 1–12. [Google Scholar]
- Kimaro, A.A.; Mpanda, M.; Rioux, J.; Aynekulu, E.; Shaba, E.; Thiongo, S.; Mutuo, M.; Abwanda, P.; Shepherd, S.K.; Neuf, H.; et al. Is Conservation Agriculture ‘Climate-smart for Maize Farmers in the Highlands of Tanzania? Nutr. Cycle. Agroecosystem 2016, 105, 217–228. [Google Scholar] [CrossRef]
- Habben, J.E.; Schussler, J.R. Effects of Drought Stress on Maize Kernel Set. In Maize Kernel Development; Centre for Agriculture and Bioscience International: Oxfordshire, UK, 2017. [Google Scholar]
- Waraich, E.A.; Ahmad, R.; Ashraf, M.Y. Role of Mineral Nutrition in Alleviation of Drought Stress in Plants. Aust. J. Crop Sci. 2011, 5, 764. [Google Scholar]
- Aslam, M.; Zamir, M.S.I.; Afzal, I.; Yaseen, M.; Mubeen, M.; Shoaib, A. Drought Stress, Its Effect on Maize Production and Development of Drought Tolerance through Potassium Application. Cercet. Agron. 2013, 46, 99–114. [Google Scholar]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The Critical Role of Potassium in Plant Stress Response. Int. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.; Zahan, F.; Sikdar, M.S.; El Sabagh, A.; Barutçular, C.; Islam, M.S.; Ratnasekera, D. Evaluation of Salt Tolerance Mung Bean Genotypes and Mitigation of Salt Stress through Potassium Nitrate Fertilization. Fresen. Environ. Bull. 2017, 26, 7218–7226. [Google Scholar]
- Abbasi, H.; Jamil, M.; Haq, A.; Ali, S.; Ahmad, R.; Malik, Z. Salt Stress Manifestation on Plants, Mechanism of Salt Tolerance and Potassium Role in Alleviating It: A Review. Zemdirb. Agric. 2016, 103, 229–238. [Google Scholar] [CrossRef] [Green Version]
- DaCosta, M.; Huang, B.R. Osmotic Adjustment Associated with Variation in Bentgrass Tolerance to Drought Stress. J. Amer. Soc. Hortic. Sci. 2006, 131, 338–344. [Google Scholar] [CrossRef]
- Jin, S.H.; Huang, J.Q.; Li, X.Q.; Zheng, B.S.; Wu, J.S.; Wang, Z.J.; Liu, G.H.; Chen, M. Effects of Potassium Supply on Limitations of Photosynthesis by Mesophyll Diffusion Conductance in Carya cathayensis. Tree Physiol. 2011, 31, 1142–1151. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.A.; Oosterhuis, D.M. Efficacy of Foliar Application to Cotton of Potassium Compounds at Different pH Levels. In Proceedings of the Beltwide Cotton Conferences, San Antonio, TX, USA, 4–7 January 1995. [Google Scholar]
- Mallarino, A.P.; Haq, M.U.; Wittry, D.; Bermudez, M. Variation in Soybean Response to Early Season Foliar Fertilization among and within Fields. Agron. J. 2001, 93, 1220–1226. [Google Scholar] [CrossRef]
- Gewin, V. Food an Underground Revolution. Nature 2010, 466, 552–553. [Google Scholar] [CrossRef] [PubMed]
- Martineau, E.; Domec, J.C.; Bosc, A.; Denoroy, P.; Fandino, V.A.; Lavres, J.J.; Jordan-Meille, L. The Effects of Potassium Nutrition on Water Use in Field-grown Maize (Zea mays L.). Environ. Exp. Bot. 2017, 134, 62–71. [Google Scholar] [CrossRef]
- Etienne, P.; Diquelou, S.; Prudent, M.; Salon, C.; Maillard, A.; Ourry, A. Macro and Micronutrient Storage in Plants and Their Remobilization When Facing Scarcity: The Case of Drought. Agriculture 2018, 8, 14. [Google Scholar] [CrossRef] [Green Version]
- Subbarao, G.V.; Wheeler, R.M.; Stutte, G.W.; Levine, L.H. Low Potassium Enhances Sodium Uptake in Red-beet under Moderate Saline Conditions. J. Plant Nutr. 2000, 23, 1449–1470. [Google Scholar] [CrossRef] [Green Version]
- Long, S.P.; Bernacchi, C.J. Gas Exchange Measurements, What Can They Tell Us about the Understanding Limitations of Photosynthesis? Procedures and Sources of Error. J. Exp. Bot. 2003, 54, 2393–2401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnon, D.T. Copper Enzymes in Isolated Chloroplasts Polyphenol-oxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bates, L.S.; Waldron, R.P.; Teaxe, I.W. Rapid Determination of Free Proline for Water Stress Studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Steel, R.G.D.; Torrie, J.H.; Dicky, D.A. Principles and Procedures of Statistics. A Biometrical Approach, 3rd ed.; McGraw Hill Book Int. Co.: New York, NY, USA, 1997; pp. 172–177. [Google Scholar]
- Pettigrew, W.T. Potassium Influences on Yield and Quality Production for Maize, Wheat, Soybean and Cotton. Physiol. Plant. 2008, 1339, 670–681. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.G.; Zhao, X.H.; Jiang, C.J.; Li, C.H.; Cong, S.; Wu, D.; Chen, Y.Q.; Yu, H.Q.; Wang, C.Y. Effects of Potassium Deficiency on Photosynthesis and Photoprotection Mechanisms in Soybean (Glycine max (L.) Merr.). J Integ. Agric. 2015, 14, 856–863. [Google Scholar] [CrossRef]
- Burdon, J.J.; Rebetzke, G.; Morell, M. Targets and Traits in the Pre-breeding Development Pipeline at Csiro 2012 WA Agribusiness Crop Updates; Western Australian Agriculture Authority: Perth, Australia, 2012. [Google Scholar]
- Henry, A. IRRI’s Drought Stress Research in Rice with Emphasis on Roots: Accomplishments Over the Last 50 Years. Plant Root. 2013, 7, 5–19. [Google Scholar] [CrossRef] [Green Version]
- Ayalew, H.; Ma, X.; Yan, G. Screening Wheat (Triticum spp.) Genotypes for Root Length under Contrasting Water Regimes: Potential Sources of Variability for Drought Resistance Breeding. J. Agron. Crop Sci. 2015, 201, 189–194. [Google Scholar] [CrossRef]
- Bahrami-Rad, S.; Hajiboland, R. Effect of Potassium Application in Drought-stressed Tobacco (Nicotiana rustica L.) Plants: Comparison of Root with Foliar Application. Ann. Agric. Sci. 2017, 62, 121–130. [Google Scholar] [CrossRef]
- Römheld, V.; Kirkby, E.A. Research on Potassium in Agriculture: Needs and Prospects. Plant Soil 2010, 335, 155–180. [Google Scholar] [CrossRef]
- Fazeli, F.; Ghorbanli, M.; Niknam, V. Effect of Drought on Biomass, Protein Content, Lipid Peroxidation and Antioxidant Enzymes in Two Sesame Cultivars. Biol. Plant. 2007, 51, 98–103. [Google Scholar] [CrossRef]
- Degenkolbe, T.; Do, P.T.; Zuther, E.; Repsilber, D.; Walther, D.; Hincha, D.K.; Köhl, K.I. Expression Profiling of Rice Cultivars Differing in Their Tolerance to Long-term Drought Stress. Plant Mol. Biol. 2009, 69, 133–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hell, R.; Mendel, R.R. Cell Biology of Metals and Nutrients; Springer: New York, NY, USA, 2010; pp. 209–246. [Google Scholar]
- Singh, P.; Blanke, M.M. Deficiency of Potassium but Not Phosphorus Enhances Root Respiration. Plant Growth Regul. 2000, 32, 77–81. [Google Scholar] [CrossRef]
- Sritontip, C.; Khaosumain, Y.; Changjeraja, S.; Changjeraja, R. Effects of Light Intensity and Potassium Chlorate on Photosynthesis and Flowering in ‘Do’ Longan. Acta Hortic. 2008, 787, 285–288. [Google Scholar] [CrossRef]
- Xu, Y.W.; Zou, Y.T.; Husaini, A.M.; Zeng, J.W.; Guan, L.L.; Liu, Q.; Wu, W. Optimization of Potassium for Proper Growth and Physiological Response of Houttuynia Cordata Thunb. Environ. Exp. Bot. 2011, 71, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Kanai, S.; Moghaieb, R.E.; El-Shemy, H.A.; Panigrahi, R.; Mohapatra, P.K.; Ito, J.; Nguyen, N.T.; Saneoka, H.; Fujita, K. Potassium Deficiency Affects Water Status and Photosynthetic Rate of the Vegetative Sink in Green House Tomato Prior to Its Effects on Source Activity. Plant Sci. 2011, 180, 368–374. [Google Scholar] [CrossRef]
- Tartar, O. Influence of Water Stress on Leaf Relative Water Content of Wheat. Asian J. Plant Sci. 2008, 7, 409–412. [Google Scholar] [CrossRef] [Green Version]
- Cochrane, T.T.; Cochrane, T.A. The Vital Role of Potassium in the Osmotic Mechanism of Stomata Aperture Modulation and Its Link with Potassium Deficiency. Plant Signal. Behav. 2009, 4, 240–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Z.; Lu, J.; Pan, Y.; Lu, P.; Li, X.; Cong, R.; Ren, T. Anatomical Variation of Mesophyll Conductance under Potassium Deficiency Has a Vital Role in Determining Leaf Photosynthesis. Plant Cell Environ. 2016, 39, 2428–2439. [Google Scholar] [CrossRef] [PubMed]
- Bednarz, C.W.; Oosterhuis, D.M.; Evans, R.D. Leaf Photosynthesis and Carbon Isotope Discrimination of Cotton in Response to Potassium Deficiency. Environ. Exp. Bot. 1998, 39, 131–139. [Google Scholar] [CrossRef]
- Chakhchar, A.; Lamaoui, M.; Wahbi, S.; Ferradous, A.; El Mousadik, A.; Ibnsouda-Koraichi, S.; Filali-Maltouf, A.; Modafar, C.E. Leaf Water Status, Osmoregulation and Secondary Metabolism as a Model for Depicting Drought Tolerance in Argania spinosa. Acta Physiol. Planta 2015, 37, 80–96. [Google Scholar] [CrossRef]
- Shemi, R.; Wang, R.; Gheith, E.S.M.; Hussain, H.A.; Hussain, S.; Irfan, M.; Cholidah, L.; Zhang, K.; Zhang, S.; Wang, L. Effects of Salicylic Acid, Zinc and Glycine Betaine on Morpho-physiological Growth and Yield of Maize under Drought Stress. Sci. Rep. 2021, 11, 3195. [Google Scholar] [CrossRef] [PubMed]
- Anjum, S.A.; Wang, L.C.; Farooq, M.; Khan, I.; Xue, L.L. Methyl Jasmonate-induced Alteration in Lipid Peroxidation, Antioxidative Defense System and Yield in Soybean under Drought. J. Agron. Crop. Sci. 2011, 197, 296–301. [Google Scholar] [CrossRef]
- Karimpour, M. Effect of Drought Stress on RWC and Chlorophyll Content on Wheat (Triticum durum L.) Genotypes. World Ess. J. 2019, 7, 52–56. [Google Scholar]
- Rong-Hua, L.; Pei-Guo, G.; Baum, M.; Grando, S.; Ceccarelli, S. Evaluation of Chlorophyll Content and Fluorescence Parameters as Indicators of Drought Tolerance in Barley. Agric. Sci. China 2006, 5, 751–757. [Google Scholar]
- Asgharipour, M.R.; Heidari, M. Effect of Potassium Supply on Drought Resistance in Sorghum: Plant Growth and Macronutrient Content. Pak. J. Agri. Sci. 2011, 48, 197–204. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
| SOV | DF | RL | RDW | SL | SDW | E | A | gs |
|---|---|---|---|---|---|---|---|---|
| Variety (V) | 1 | 141.5 ** | 3.9 ns | 44.1* | 589.2 ** | 1.7 ** | 2112.5 ** | 0.021 ** |
| Field Capacity (FC) | 2 | 91.6 ** | 41.2 ** | 226.4 ** | 241.5 ** | 10.2 ** | 311.0 ** | 0.007 ** |
| Potassium (K) | 2 | 234.6 ** | 932.2 ** | 61.6 ** | 968.2 ** | 34.4 ** | 34.0 ** | 0.001 * |
| V × FC | 2 | 212.0 ** | 479.4 ** | 178.4 ** | 196.8 ** | 72.3 ** | 0.007 ns | 0.004 ** |
| V × K | 2 | 55.0 ** | 22.2 ** | 108.8 ** | 228.3 ** | 11.9 ** | 0.010 ns | 0.000 ns |
| FC × K | 4 | 30.3 ** | 29.2 ** | 472.7 ** | 31.8 ** | 10.4 ** | 0.001 ns | 0.001 ns |
| V × FC × K | 4 | 8.9 ** | 246.2 ** | 355.9 ** | 132.9 ** | 18.3 ** | 0.001 ns | 0.000 ns |
| Error | 36 | 2.3 | 2.8 | 9.1 | 3.0 | 0.2057 | 1.85074 | 0.00024 |
| SOV | DF | Proline | RWC | Chl a | Chl b | T Chl |
|---|---|---|---|---|---|---|
| Variety (V) | 1 | 0.085 ** | 9657.7 ** | 0.001 ns | 0.010 ** | 0.120 ** |
| Field Capacity (FC) | 2 | 0.015 ** | 1224.2 ** | 0.002 ns | 0.008 ** | 0.031 ** |
| Potassium (K) | 2 | 0.004 ** | 145.7 ** | 0.002 ns | 0.008 ** | 0.002 ** |
| V × FC | 2 | 0.005 ** | 16.1 * | 0.005 ** | 0.000 ** | 0.002 ** |
| V × K | 2 | 0.002 * | 13.7 ns | 0.010 ** | 0.008 ** | 0.000 ns |
| FC × K | 4 | 0.000 ns | 1.01 ns | 0.002 ns | 0.011 ** | 0.000 ns |
| V × FC × K | 4 | 0.000 ns | 19.4 ** | 0.002 ns | 0.010 ** | 0.000 ns |
| Error | 36 | 0.085 | 5.03 | 0.00071 | 0.00004 | 0.00031 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wasaya, A.; Affan, M.; Ahmad Yasir, T.; Atique-ur-Rehman; Mubeen, K.; Rehman, H.u.; Ali, M.; Nawaz, F.; Galal, A.; Iqbal, M.A.; et al. Foliar Potassium Sulfate Application Improved Photosynthetic Characteristics, Water Relations and Seedling Growth of Drought-Stressed Maize. Atmosphere 2021, 12, 663. https://doi.org/10.3390/atmos12060663
Wasaya A, Affan M, Ahmad Yasir T, Atique-ur-Rehman, Mubeen K, Rehman Hu, Ali M, Nawaz F, Galal A, Iqbal MA, et al. Foliar Potassium Sulfate Application Improved Photosynthetic Characteristics, Water Relations and Seedling Growth of Drought-Stressed Maize. Atmosphere. 2021; 12(6):663. https://doi.org/10.3390/atmos12060663
Chicago/Turabian StyleWasaya, Allah, Muhammad Affan, Tauqeer Ahmad Yasir, Atique-ur-Rehman, Khuram Mubeen, Haseeb ur Rehman, Muqarrab Ali, Farukh Nawaz, Ahmed Galal, Muhammad Aamir Iqbal, and et al. 2021. "Foliar Potassium Sulfate Application Improved Photosynthetic Characteristics, Water Relations and Seedling Growth of Drought-Stressed Maize" Atmosphere 12, no. 6: 663. https://doi.org/10.3390/atmos12060663
APA StyleWasaya, A., Affan, M., Ahmad Yasir, T., Atique-ur-Rehman, Mubeen, K., Rehman, H. u., Ali, M., Nawaz, F., Galal, A., Iqbal, M. A., Islam, M. S., El-Sharnouby, M., Rahman, M. H. u., & EL Sabagh, A. (2021). Foliar Potassium Sulfate Application Improved Photosynthetic Characteristics, Water Relations and Seedling Growth of Drought-Stressed Maize. Atmosphere, 12(6), 663. https://doi.org/10.3390/atmos12060663