
N/P Addition Is More Likely Than N Addition Alone to Promote a Transition from Moss-Dominated to Graminoid-Dominated Tundra in the High-Arctic

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
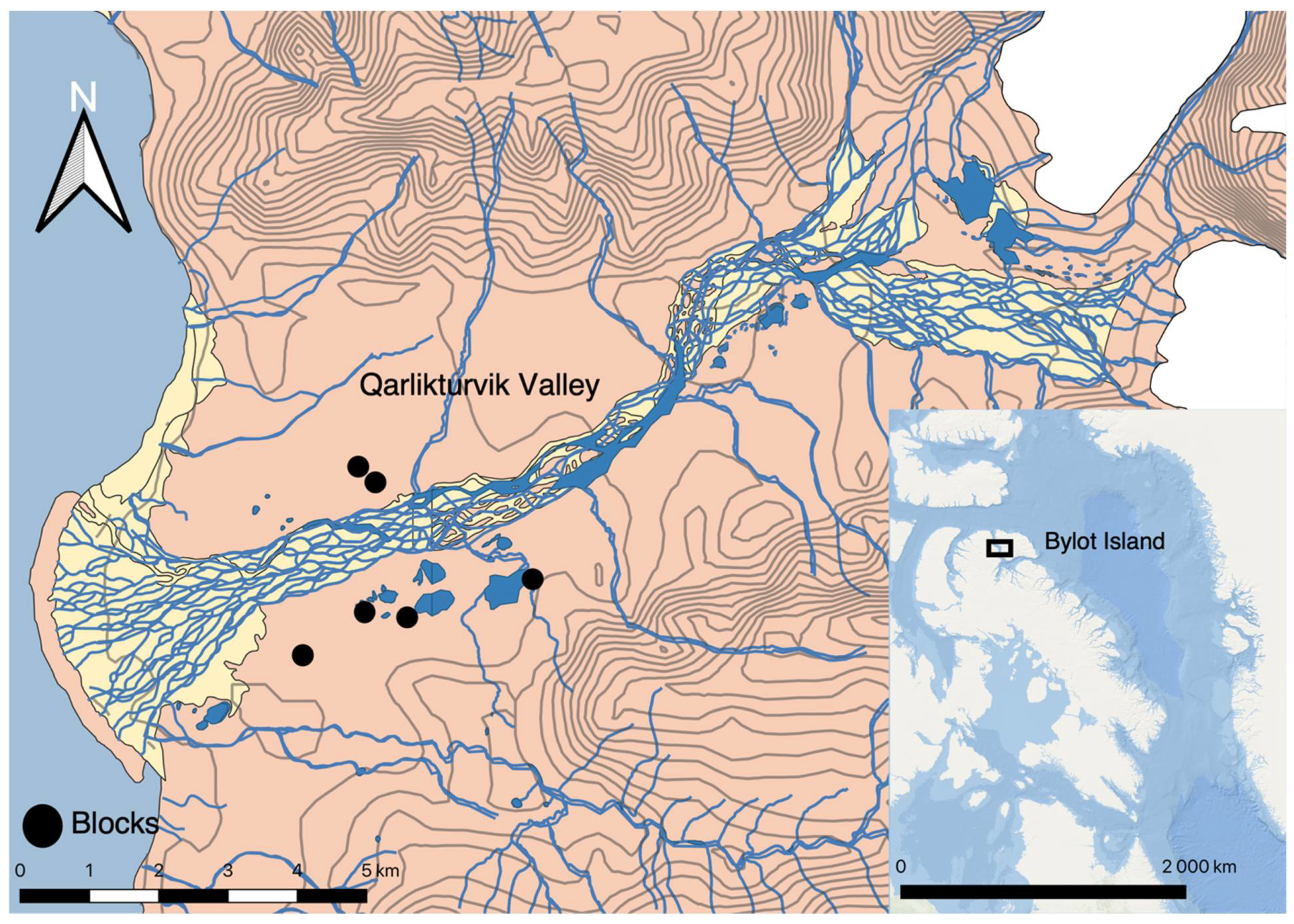
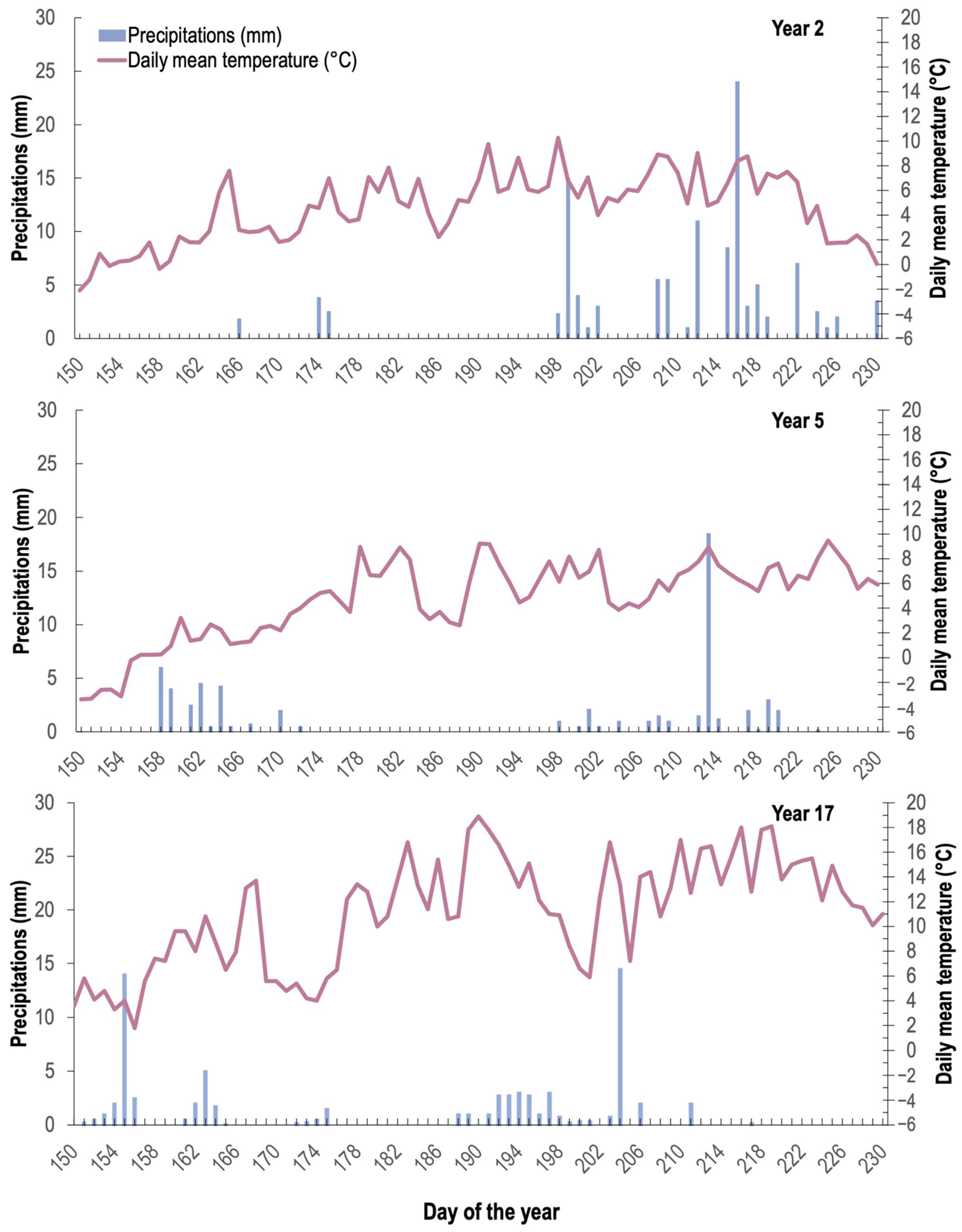
2.1. Study Area
2.2. Experimental Design and Fertilization
2.3. Aboveground Net Primary Productivity (ANPP)
2.4. Vegetation Composition
2.5. Chemical Analyses
2.6. Statistical Analysis
3. Results
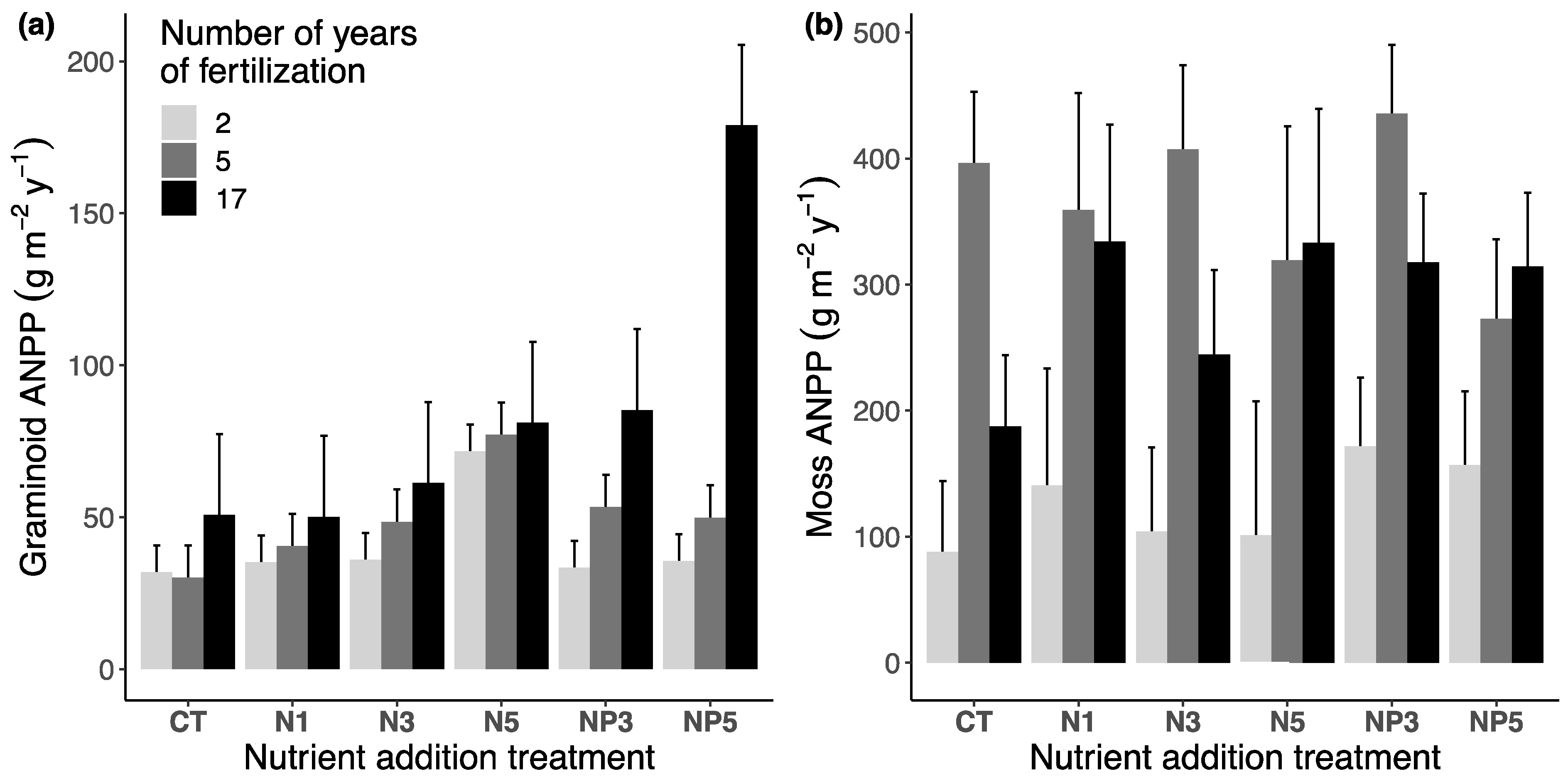
3.1. Aboveground Net Primary Productivity (ANPP)
3.1.1. Fertilization Effect after 17 Years
3.1.2. Temporal Effect of Fertilization
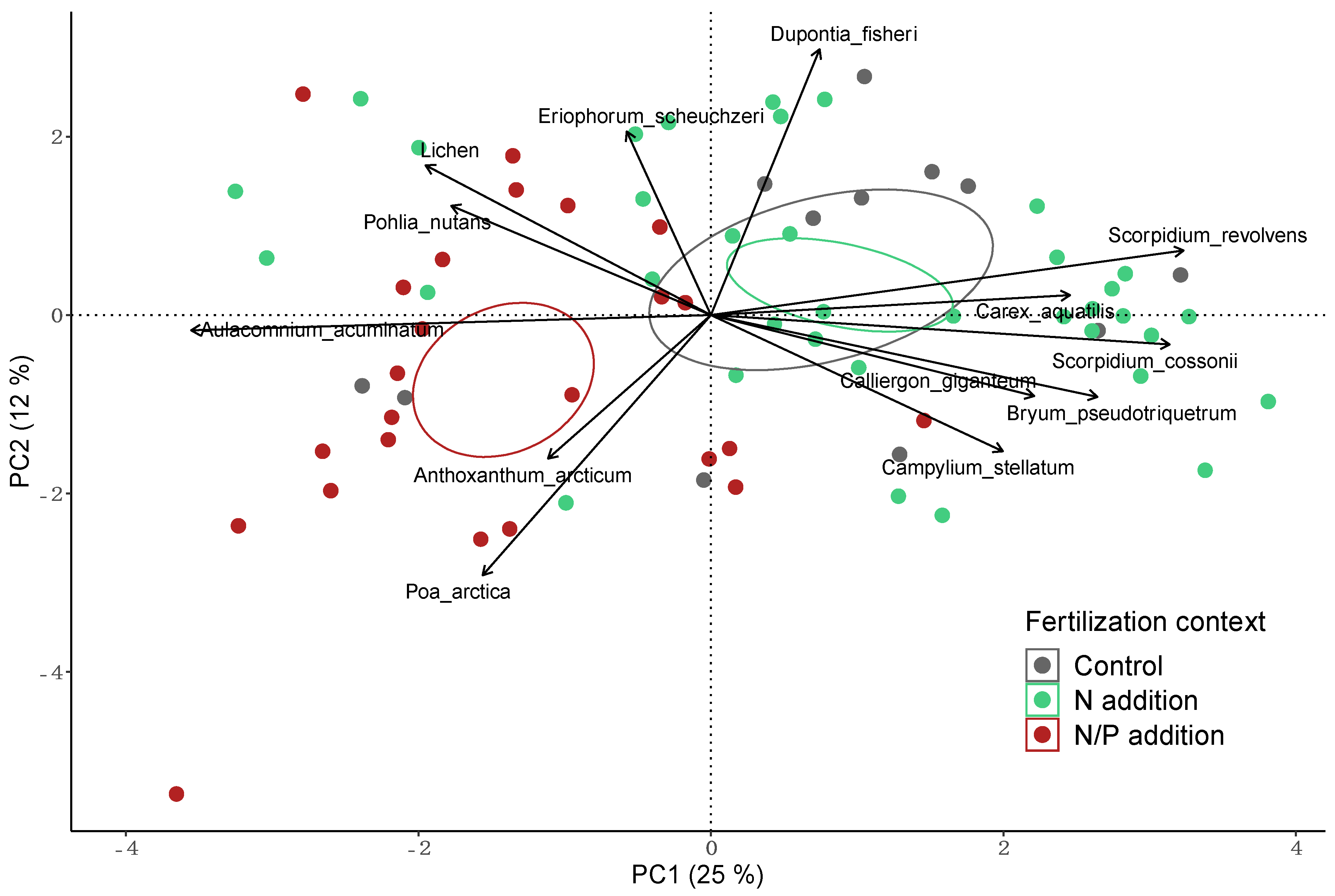
3.2. Vegetation Composition
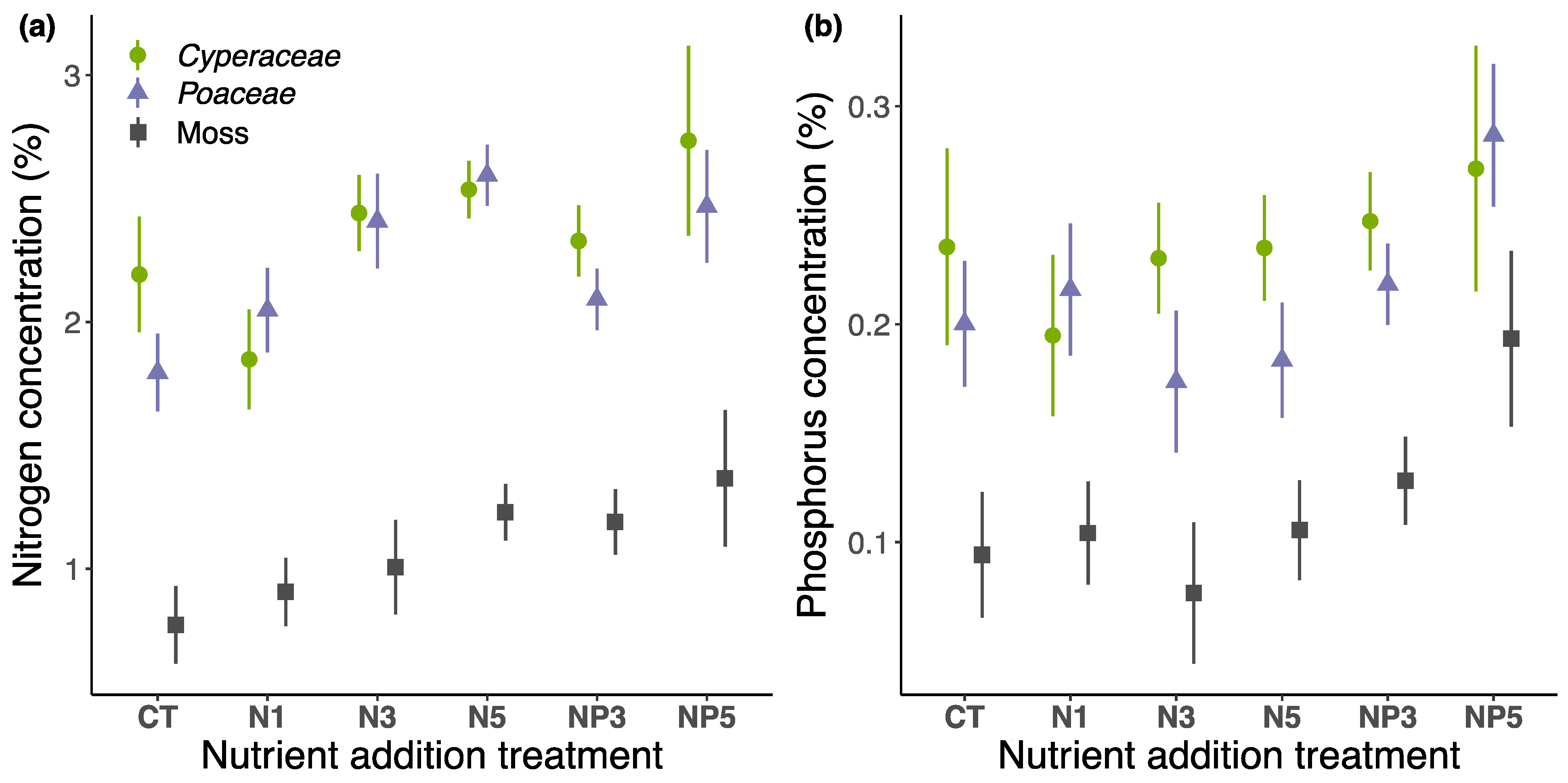
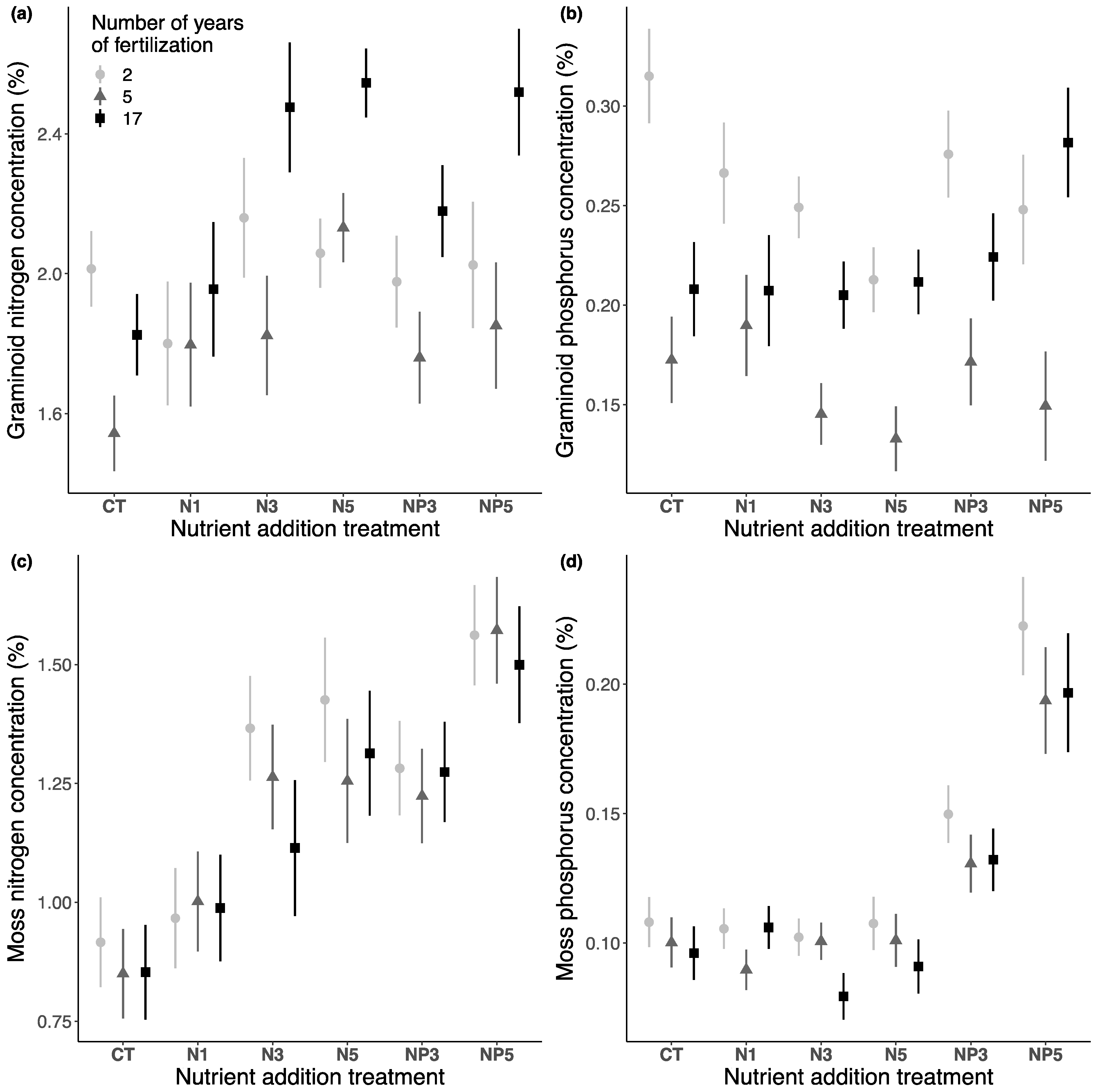
3.3. Chemical Concentration
3.3.1. Fertilization Effect after 17 Years
3.3.2. Temporal Effect of Fertilization
4. Discussion
4.1. Nitrogen Deposition vs. Global Warming
4.2. Lag in the Response Time
4.3. Limitation of the Study
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strata | Family | Species | Authority |
|---|---|---|---|
| Shrub | Salicaceae | Salix arctica | Pallas |
| Salix reticulata | Linnaeus | ||
| Graminoid | Cyperaceae | Carex aquatilis | Wahlenberg |
| Eriophorum angustifolium | Honckeny | ||
| Eriophorum scheuchzerii | Hoppe | ||
| Juncaceae | Luzula confusa | Lindeberg | |
| Luzula nivalis | (Laestadius) Sprengel | ||
| Poaceae | Alopecurus magellanicus | Lamarck | |
| Arctagrostis latifolia | (R. Brown) Grisebach | ||
| Dupontia fisheri | R. Brown | ||
| Festuca brachyphylla | Schultes & Schultes f | ||
| Anthoxanthum arcticum | Veldkamp | ||
| Pleuropogon sabinei | R. Brown | ||
| Poa arctica | R. Brown | ||
| Forb | Asteraceae | Tephroseris palustris | L. Reichenbach |
| Brassicaceae | Cardamine pratensis | Linnaeus | |
| Draba sp. | Linnaeus | ||
| Eutrema edwardsii | R. Brown | ||
| Caryophyllaceae | Cerastium arcticum | Lange | |
| Silene uralensis subsp. uralensis | Ruprecht Bocquet | ||
| Stellaria longipes | Goldie | ||
| Orobanchaceae | Pedicularis langsdorffii subsp. arctica | (R. Brown) Pennell ex Hultén | |
| Pedicularis sudetica | Willdenow | ||
| Saxifragaceae | Chrysosplenium tetrandrum | Th. Fries | |
| Saxifraga cernua | Linnaeus | ||
| Saxifraga hirculus | Linnaeus | ||
| Saxifraga oppositifolia | Linnaeus | ||
| Bryophytes | Amblystegiaceae | Campylium stellatum | (Hedwig) C. Jensen |
| Tomenthypnum nitens | (Hedwig) Loeske | ||
| Aneuraceae | Aneura pinguis | (Linnaeus) Dumort | |
| Aulacomniaceae | Aulacomnium acuminatum | (Lindberg & Arnell) Kindberg | |
| Aulacomnium palustre | (Hedwig) Schwagrichen | ||
| Aulacomnium turgidum | (Wahlenberg) Schwagrichen | ||
| Brachytheciaceae | Brachythecium turgidum | (Hartman) Kindberg | |
| Bryaceae | Bryum cryophilum | Mårtensson | |
| Bryum pseudotriquetrum | (Hedwig) Gaertner, Meyer & Scherbius | ||
| Calliergonaceae | Calliergon giganteum | (Schimper) Kindberg | |
| Scorpidium cossonii | (Schimper) Hedenas | ||
| Scorpidium revolvens | (Swartz) Rubers | ||
| Dicranaceae | Oncophorus wahlenbergii | Bridel | |
| Meesiaceae | Meesia triquetra | (Linnaeus ex Jolyclerc) Ångström | |
| Mielichhoferiaceae | Pohlia nutans | (Hedwig) Lindberg | |
| Mniaceae | Cinclidium arcticum | (Bruch & Schimper) Schimper, Kongl | |
| Pseudobryum cinclidioides | (Huebener) T.J. Kop. | ||
| Polytrichaceae | Polytrichum commune | Hedwig | |
| Polytrichum swartzii | Hartman | ||
| Sphagnaceae | Sphagnum sp. | Linnaeus |
| Plot | Fertilization Context | PC1 | PC2 | Plot | Fertilization Context | PC1 | PC2 |
|---|---|---|---|---|---|---|---|
| 1 | CT | 3.46 | 0.48 | 37 | N | 2.78 | −0.01 |
| 2 | CT | 2.94 | −0.17 | 38 | N | 0.19 | 0.02 |
| 3 | N | 3.09 | 0.37 | 39 | N | −2.18 | 0.40 |
| 4 | N | 3.27 | −0.20 | 40 | N | −0.46 | 1.38 |
| 5 | N | 3.16 | −0.85 | 41 | CT | −0.39 | −1.40 |
| 6 | N | 3.10 | −0.17 | 42 | CT | 1.36 | −1.36 |
| 7 | N | 3.55 | −0.07 | 43 | NP | −2.80 | −1.39 |
| 8 | N | 3.05 | 0.24 | 44 | NP | −2.31 | −2.03 |
| 9 | NP | 1.42 | −1.24 | 45 | NP | −2.50 | −0.67 |
| 10 | NP | −0.25 | 0.13 | 46 | NP | −2.18 | −0.21 |
| 11 | NP | −1.32 | −2.54 | 47 | N | 0.95 | 0.02 |
| 12 | NP | 0.34 | −0.02 | 48 | N | 0.39 | −0.41 |
| 13 | N | 2.67 | 0.54 | 49 | N | −0.66 | 0.64 |
| 14 | N | 2.53 | 1.07 | 50 | N | −0.22 | 1.97 |
| 15 | N | 3.52 | −1.95 | 51 | N | −0.28 | 0.27 |
| 16 | N | 3.90 | −1.03 | 52 | N | −2.01 | 1.66 |
| 17 | CT | 2.15 | 1.43 | 53 | CT | 0.87 | 2.89 |
| 18 | CT | 1.90 | 1.58 | 54 | CT | 0.82 | 1.42 |
| 19 | NP | −2.85 | −2.46 | 55 | NP | −1.12 | −0.97 |
| 20 | NP | −3.59 | −5.69 | 56 | NP | −0.84 | 1.43 |
| 21 | NP | 0.43 | −1.52 | 57 | NP | −2.91 | 2.61 |
| 22 | NP | −0.09 | −1.62 | 58 | NP | −1.75 | 1.70 |
| 23 | N | 2.60 | −0.10 | 59 | N | −0.06 | 2.52 |
| 24 | N | 2.65 | −0.02 | 60 | N | −2.42 | 2.49 |
| 25 | N | 0.53 | 0.88 | 61 | CT | −2.51 | −0.92 |
| 26 | N | 0.81 | 2.29 | 62 | CT | −2.81 | −0.72 |
| 27 | N | 0.34 | 2.39 | 63 | N | 0.86 | −0.49 |
| 28 | N | −0.80 | 2.03 | 64 | N | 1.01 | −1.99 |
| 29 | CT | 0.69 | 1.45 | 65 | N | 0.18 | −0.13 |
| 30 | CT | 0.97 | 1.12 | 66 | N | 1.35 | −0.12 |
| 31 | NP | −1.32 | 0.47 | 67 | N | 1.97 | −2.43 |
| 32 | NP | −1.55 | 0.15 | 68 | N | −0.99 | −2.09 |
| 33 | NP | −0.33 | 1.11 | 69 | NP | −0.86 | −1.58 |
| 34 | NP | −0.81 | 1.36 | 70 | NP | −2.18 | −1.97 |
| 35 | N | −3.08 | 0.53 | 71 | NP | −2.45 | −1.01 |
| 36 | N | −2.84 | 1.49 | 72 | NP | −2.58 | −1.27 |
| Species | PC1 | PC2 |
|---|---|---|
| Aulacomnium acuminatum * | 0.54 * | 0.33 * |
| Aneura pinguis | −0.01 | 0.01 |
| Brachythecium turgidum | 0.04 | 0.07 |
| Bryum pseudotriquetrum | −0.21 | −0.10 |
| Calliergon giganteum | −0.11 | −0.08 |
| Campylium stellatum | −0.16 | −0.27 * |
| Cinclidium arcticum | 0.03 | −0.09 |
| Meesia triquetra | −0.11 | −0.11 |
| Oncophorus wahlenbergii | 0.00 | 0.00 |
| Pohlia nutans | 0.09 | 0.20 |
| Polytrichum swartzii | 0.08 | 0.03 |
| Scorpidium cossonii * | −0.40 * | −0.22 * |
| Scorpidium revolvens * | −0.42 * | −0.01 |
| Sphagnum sp. | −0.01 | 0.05 |
| Tomenthypnum nitens | 0.07 | −0.14 |
| Lichen * | 0.13 | 0.24 * |
| Eriophorum scheuchzeri * | 0.02 | 0.30 * |
| Arctagrostis latifolia | −0.20 | 0.09 |
| Carex aquatilis * | −0.25 * | 0.07 |
| Chrysosplenium tetrandrum | 0.01 | −0.03 |
| Dupontia fisheri * | −0.07 | 0.34 * |
| Anthoxanthum arcticum | 0.13 | −0.05 |
| Pedicularis langsdorffii subsp. arctica | −0.11 | 0.10 |
| Poa arctica * | 0.30 * | −0.72 * |
| Draba sp. | 0.03 | 0.03 |
| Salix arctica | 0.12 | 0.11 |

References
- Remmert, H. Arctic Animal Ecology; Springer: Berlin, Germany, 1980; 252p. [Google Scholar]
- Tedrow, J.C.F. Soils of the Polar Landscapes; Rutgers University Press: New Brunswick, NJ, USA, 1977. [Google Scholar]
- Heal, O.; Flanagan, P.W.; French, D.D.; MacLean, J.S.F. Decomposition and accumulation of organic matter in tundra. In Tundra Ecosystems: A Comparative Analysis; Cambridge University Press: Cambridge, UK, 1981; pp. 587–633. [Google Scholar]
- Mikan, C.J.; Schimel, J.P.; Doyle, A.P. Temperature controls of microbial respiration in arctic tundra soils above and below freezing. Soil Biol. Biochem. 2002, 34, 1785–1795. [Google Scholar] [CrossRef]
- Chapin, F.S.; Rincon, E.; Huante, P. Environmental responses of plants and ecosystems as predictors of the impact of global change. J. Biosci. 1993, 18, 515–524. [Google Scholar] [CrossRef]
- Nadelhoffer, K.J.; Giblin, A.E.; Shaver, G.R.; Linkins, A.E. Microbial processes and plant nutrient availability in arctic soils. In Arctic Ecosystems in a Changing Climate; Academic Press, Inc.: San Diego, CA, USA, 1992. [Google Scholar]
- Shaver, G.R.; Giblin, A.E.; Nadelhoffer, K.J.; Thieler, K.K.; Downs, M.R.; Laundre, J.A.; Rastetter, E.B. Carbon turnover in Alaskan tundra soils: Effects of organic matter quality, temperature, moisture and fertilizer. J. Ecol. 2006, 94, 740–753. [Google Scholar] [CrossRef] [Green Version]
- McLaren, J.R.; Buckeridge, K.M. Enhanced plant leaf P and unchanged soil P stocks after a quarter century of warming in the arctic tundra. Ecosphere 2021, 12, e03838. [Google Scholar] [CrossRef]
- Marty, C.; Houle, D.; Gagnon, C.; Courchesne, F. The relationships of soil total nitrogen concentrations, pools and C:N ratios with climate, vegetation types and nitrate deposition in temperate and boreal forests of eastern Canada. Catena 2017, 152, 163–172. [Google Scholar] [CrossRef]
- Smith, C.K.; Coyea, M.R.; Munson, A.D. Soil carbon, nitrogen, and phosphorus stocks and dynamics under disturbed black spruce forests. Ecol. Appl. 2000, 10, 775–788. [Google Scholar] [CrossRef]
- Jiang, J.; Wang, Y.P.; Yang, Y.H.; Yu, M.X.; Wang, C.; Yan, J.H. Interactive effects of nitrogen and phosphorus additions on plant growth vary with ecosystem type. Plant Soil 2019, 440, 523–537. [Google Scholar] [CrossRef]
- Klupar, I.; Rocha, A.V.; Rastetter, E.B. Alleviation of nutrient co-limitation induces regime shifts in post-fire community composition and productivity in Arctic tundra. Glob. Chang. Biol. 2021, 27, 3324–3335. [Google Scholar] [CrossRef]
- Post, E.; Alley, R.B.; Christensen, T.R.; Macias-Fauria, M.; Forbes, B.C.; Gooseff, M.N.; Iler, A.; Kerby, J.T.; Laidre, K.L.; Mann, M.E.; et al. The polar regions in a 2 degrees C warmer world. Sci. Adv. 2019, 5, 12. [Google Scholar] [CrossRef] [Green Version]
- Sistla, S.A.; Moore, J.C.; Simpson, R.T.; Gough, L.; Shaver, G.R.; Schimel, J.P. Long-term warming restructures Arctic tundra without changing net soil carbon storage. Nature 2013, 497, 615–618. [Google Scholar] [CrossRef]
- Nadelhoffer, K.J.; Giblin, A.E.; Shaver, G.R.; Laundre, J.A. Effects of temperature and substrate quality on element mineralization in 6 arctic soils. Ecology 1991, 72, 242–253. [Google Scholar] [CrossRef]
- Schmidt, I.; Jonasson, S.; Shaver, G.; Michelsen, A.; Nordin, A. Mineralization and distribution of nutrients in plants and microbes in four arctic ecosystems: Responses to warming. Plant Soil 2002, 242, 93–106. [Google Scholar] [CrossRef]
- Koyama, A.; Wallenstein, M.D.; Simpson, R.T.; Moore, J.C. Soil bacterial community composition altered by increased nutrient availability in Arctic tundra soils. Front. Microbiol. 2014, 5, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mack, M.C.; Schuur, E.A.G.; Bret-Harte, M.S.; Shaver, G.R.; Chapin, F.S. Ecosystem carbon storage in arctic tundra reduced by long-term nutrient fertilization. Nature 2004, 431, 440–443. [Google Scholar] [CrossRef] [PubMed]
- Nowinski, N.S.; Trumbore, S.E.; Schuur, E.A.G.; Mack, M.C.; Shaver, G.R. Nutrient addition prompts rapid destabilization of organic matter in an arctic tundra ecosystem. Ecosystems 2008, 11, 16–25. [Google Scholar] [CrossRef] [Green Version]
- Chapin, F.S. Integrated responses of plants to stress. Bioscience 1991, 41, 29–36. [Google Scholar] [CrossRef]
- Jiang, Y.Y.; Rocha, A.V.; Rastetter, E.B.; Shaver, G.R.; Mishra, U.; Zhuang, Q.L.; Kwiatkowski, B.L. C-N-P interactions control climate driven changes in regional patterns of C storage on the North Slope of Alaska. Landsc. Ecol. 2016, 31, 195–213. [Google Scholar] [CrossRef]
- Shaver, G.R.; Chapin, F.; Gartner, B.L. Factors limiting seasonal growth and peak biomass accumulation in eriophorum vaginatum in alaskan tussock tundra. J. Ecol. 1986, 74, 257–278. [Google Scholar] [CrossRef]
- Marion, G.M.; Hastings, S.J.; Oberbauer, S.F.; Oechel, W.C. Soil-plant element relationships in a tundra ecosystem. Holarct. Ecol. 1989, 12, 296–303. [Google Scholar] [CrossRef]
- DeMarco, J.; Mack, M.C.; Bret-Harte, M.S.; Burton, M.; Shaver, G.R. Long-term experimental warming and nutrient additions increase productivity in tall deciduous shrub tundra. Ecosphere 2014, 5, 22. [Google Scholar] [CrossRef]
- Stewart, E.J.; Dawson, J.; Draper, D. Cruise Tourism and Residents in Arctic Canada: Development of a Resident Attitude Typology. J. Hosp. Tour. Manag. 2011, 18, 95–106. [Google Scholar] [CrossRef]
- Harsem, O.; Eide, A.; Heen, K. Factors influencing future oil and gas prospects in the Arctic. Energy Policy 2011, 39, 8037–8045. [Google Scholar] [CrossRef] [Green Version]
- Winther, M.; Christensen, J.H.; Plejdrup, M.S.; Ravn, E.S.; Eriksson, O.F.; Kristensen, H.O. Emission inventories for ships in the arctic based on satellite sampled AIS data. Atmos. Environ. 2014, 91, 1–14. [Google Scholar] [CrossRef]
- Held, I.M.; Soden, B.J. Robust responses of the hydrological cycle to global warming. J. Clim. 2006, 19, 5686–5699. [Google Scholar] [CrossRef]
- Choudhary, S.; Blaud, A.; Osborn, A.M.; Press, M.C.; Phoenix, G.K. Nitrogen accumulation and partitioning in a High Arctic tundra ecosystem from extreme atmospheric N deposition events. Sci. Total Environ. 2016, 554, 303–310. [Google Scholar] [CrossRef]
- Serreze, M.C.; Francis, J.A. The arctic amplification debate. Clim. Chang. 2006, 76, 241–264. [Google Scholar] [CrossRef] [Green Version]
- De Schrijver, A.; De Frenne, P.; Ampoorter, E.; Van Nevel, L.; Demey, A.; Wuyts, K.; Verheyen, K. Cumulative nitrogen input drives species loss in terrestrial ecosystems. Glob. Ecol. Biogeogr. 2011, 20, 803–816. [Google Scholar] [CrossRef]
- van der Wal, R.; Pearce, I.S.K.; Brooker, R.W. Mosses and the struggle for light in a nitrogen-polluted world. Oecologia 2005, 142, 159–168. [Google Scholar] [CrossRef]
- Virtanen, R.; Johnston, A.E.; Crawley, M.J.; Edwards, G.R. Bryophyte biomass and species richness on the Park Grass Experiment, Rothamsted, UK. Plant Ecol. 2000, 151, 129–141. [Google Scholar] [CrossRef]
- Alatalo, J.M.; Little, C.J.; Jagerbrand, A.K.; Molau, U. Vascular plant abundance and diversity in an alpine heath under observed and simulated global change. Sci. Rep. 2015, 5, 11. [Google Scholar] [CrossRef] [Green Version]
- Chagnon, C.; Boudreau, S. Shrub canopy induces a decline in lichen abundance and diversity in Nunavik (Quebec, Canada). Arct. Antarct. Alp. Res. 2019, 51, 521–532. [Google Scholar] [CrossRef]
- Lang, S.I.; Cornelissen, J.H.C.; Shaver, G.R.; Ahrens, M.; Callaghan, T.V.; Molau, U.; Ter Braak, C.J.F.; Holzer, A.; Aerts, R. Arctic warming on two continents has consistent negative effects on lichen diversity and mixed effects on bryophyte diversity. Glob. Chang. Biol. 2012, 18, 1096–1107. [Google Scholar] [CrossRef]
- Vuorinen, K.E.M.; Oksanen, L.; Oksanen, T.; Pyykonen, A.; Olofsson, J.; Virtanen, R. Open tundra persist, but arctic features decline-Vegetation changes in the warming Fennoscandian tundra. Glob. Chang. Biol. 2017, 23, 3794–3807. [Google Scholar] [CrossRef] [PubMed]
- Walker, M.D.; Wahren, C.H.; Hollister, R.D.; Henry, G.H.R.; Ahlquist, L.E.; Alatalo, J.M.; Bret-Harte, M.S.; Calef, M.P.; Callaghan, T.V.; Carroll, A.B.; et al. Plant community responses to experimental warming across the tundra biome. Proc. Natl. Acad. Sci. USA 2006, 103, 1342–1346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bobbink, R.; Hicks, K.; Galloway, J.; Spranger, T.; Alkemade, R.; Ashmore, M.; Bustamante, M.; Cinderby, S.; Davidson, E.; Dentener, F.; et al. Global assessment of nitrogen deposition effects on terrestrial plant diversity: A synthesis. Ecol. Appl. 2010, 20, 30–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hobbie, S.E.; Gough, L. Foliar and soil nutrients in tundra on glacial landscapes of contrasting ages in northern Alaska. Oecologia 2002, 131, 453–462. [Google Scholar] [CrossRef]
- Chapin, F.S.; Shaver, G.R. Physiological and growth responses of arctic plants to a field experiment simulating climatic change. Ecology 1996, 77, 822–840. [Google Scholar] [CrossRef]
- Kotanen, P.M. Fates of added nitrogen in freshwater arctic wetlands grazed by Snow Geese: The role of mosses. Arct. Antarct. Alp. Res. 2002, 34, 219–225. [Google Scholar] [CrossRef]
- Pouliot, R.; Rochefort, L.; Gauthier, G. Moss carpets constrain the fertilizing effects of herbivores on graminoid plants in arctic polygon fens. Botany 2009, 87, 1209–1222. [Google Scholar] [CrossRef]
- Gauthier, G.; Bêty, J.; Cadieux, M.C.; Legagneux, P.; Doiron, M.; Chevallier, C.; Lai, S.; Tarroux, A.; Berteaux, D. Long-term monitoring at multiple trophic levels suggests heterogeneity in responses to climate change in the Canadian Arctic tundra. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2013, 368, 20120482. [Google Scholar] [CrossRef] [Green Version]
- Ellis, C.J.; Rochefort, L. Century-scale development of polygon-patterned tundra wetland, Bylot Island (73° N, 80° W). Ecology 2004, 85, 963–978. [Google Scholar] [CrossRef]
- Massé, H.; Rochefort, L.; Gauthier, G. Carrying capacity of wetland habitats used by breeding greater snow geese. J. Wildl. Manag. 2001, 65, 271–281. [Google Scholar] [CrossRef]
- Gamon, J.A.; Kershaw, G.P.; Williamson, S.; Hik, D.S. Microtopographic patterns in an arctic baydjarakh field: Do fine-grain patterns enforce landscape stability? Environ. Res. Lett. 2012, 7, 6. [Google Scholar] [CrossRef]
- Zona, D.; Lipson, D.A.; Zulueta, R.C.; Oberbauer, S.F.; Oechel, W.C. Microtopographic controls on ecosystem functioning in the Arctic Coastal Plain. J. Geophys. Res.-Biogeosci. 2011, 116, 12. [Google Scholar] [CrossRef] [Green Version]
- Marchand-Roy, M. L’effet Fertilisant de la Grande Oie des Neiges: Cinq ans de Suivi de L’azote et du Phosphore dans les Polygones de tourbe de l’Île Bylot au Nunavut; Université Laval: Québec, QC, Canada, 2009. [Google Scholar]
- Gauthier, G.; Berteaux, D.; Bêty, J.; Tarroux, A.; Therrien, J.F.; McKinnon, L.; Legagneux, P.; Cadieux, M.C. The tundra food web of Bylot Island in a changing climate and the role of exchanges between ecosystems. Ecoscience 2011, 18, 223–235. [Google Scholar] [CrossRef]
- Godin, É. Arctic Change 2020 Abstracts (continued). Arct. Sci. 2020, 1, 240–368. [Google Scholar] [CrossRef]
- CEN. Climate Station Data from Bylot Island in Nunavut, Canada, v. 1.11 (1992–2019); CEN: Québec, QC, Canada, 2020. [Google Scholar] [CrossRef]
- Vivanco, M.G.; Theobald, M.R.; Garcia-Gomez, H.; Garrido, J.L.; Prank, M.; Aas, W.; Adani, M.; Alyuz, U.; Andersson, C.; Bellasio, R.; et al. Modeled deposition of nitrogen and sulfur in Europe estimated by 14 air quality model systems: Evaluation, effects of changes in emissions and implications for habitat protection. Atmos. Chem. Phys. 2018, 18, 10199–10218. [Google Scholar] [CrossRef] [Green Version]
- Ackerman, D.; Millet, D.B.; Chen, X. Global Estimates of Inorganic Nitrogen Deposition Across Four Decades. Glob. Biogeochem. Cycles 2019, 33, 100–107. [Google Scholar] [CrossRef] [Green Version]
- Hik, D.S.; Jefferies, R.L. Increase in the net aboveground primary production of a salt-marsh forage grass: A test of the predictions of the herbivore-optimization. J. Ecol. 1990, 78, 180–195. [Google Scholar] [CrossRef] [Green Version]
- Kielland, K. Amino-acid absorption by arctic plants—Implication for plant nutrition and nitrogen cycling. Ecology 1994, 75, 2373–2383. [Google Scholar] [CrossRef]
- Morrissette-Boileau, C.; Boudreau, S.; Tremblay, J.P.; Cote, S.D. Simulated caribou browsing limits the effect of nutrient addition on the growth of Betula glandulosa, an expanding shrub species in Eastern Canada. J. Ecol. 2018, 106, 1256–1265. [Google Scholar] [CrossRef]
- Prager, C.M.; Naeem, S.; Boelman, N.T.; Eitel, J.U.H.; Greaves, H.E.; Heskel, M.A.; Magney, T.S.; Menge, D.N.L.; Vierling, L.A.; Griffin, K.L. A gradient of nutrient enrichment reveals nonlinear impacts of fertilization on Arctic plant diversity and ecosystem function. Ecol. Evol. 2017, 7, 2449–2460. [Google Scholar] [CrossRef] [PubMed]
- Saucier, V.; Champagne, E.; Cote, S.D.; Tremblay, J.P. Combined effects of simulated browsing, warming and nutrient addition on forage availability for migratory caribou in Nunavik, Canada. Polar Biol. 2019, 42, 1561–1570. [Google Scholar] [CrossRef]
- Shaver, G.R.; Chapin, F.S. Long-term responses to factorial, NPK fertilizer treatment by alaskan wet and moist tundra sedge species. Ecography 1995, 18, 259–275. [Google Scholar] [CrossRef]
- Hicks, L.C.; Yuan, M.Y.; Brangari, A.; Rousk, K.; Rousk, J. Increased Above- and Belowground Plant Input Can Both Trigger Microbial Nitrogen Mining in Subarctic Tundra Soils. Ecosystems 2022, 25, 105–121. [Google Scholar] [CrossRef]
- Limpens, J.; Granath, G.; Gunnarsson, U.; Aerts, R.; Bayley, S.; Bragazza, L.; Bubier, J.; Buttler, A.; van den Berg, L.J.L.; Francez, A.J.; et al. Climatic modifiers of the response to nitrogen deposition in peat-forming Sphagnum mosses: A meta-analysis. New Phytol. 2011, 191, 496–507. [Google Scholar] [CrossRef] [Green Version]
- Aerts, R.; van Bodegom, P.M.; Cornelissen, J.H.C. Litter stoichiometric traits of plant species of high-latitude ecosystems show high responsiveness to global change without causing strong variation in litter decomposition. New Phytol. 2012, 196, 181–188. [Google Scholar] [CrossRef]
- Shaw, A.N.; Cleveland, C.C. The effects of temperature on soil phosphorus availability and phosphatase enzyme activities: A cross-ecosystem study from the tropics to the Arctic. Biogeochemistry 2020, 151, 113–125. [Google Scholar] [CrossRef]
- Haag, R.W. Nutrient limitations to plant production in 2 tundra communities. Can. J. Bot.-Rev. Can. Bot. 1974, 52, 103–116. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate. IPCC: Climate Change 2021: The Physical Science Basis; Intergovernmental Panel on Climate: Geneva, Switzerland, 2021. [Google Scholar]
- Gauthier, G.; Hughes, J.; Reed, A.; Beaulieu, J.; Rochefort, L. Effect of grazing by Greater Snow Geese on the production of graminoids at an arctic site (Bylot Island, NWT, Canada). J. Ecol. 1995, 83, 653–664. [Google Scholar] [CrossRef]
- Pouliot, R.; Marchand-Roy, M.; Rochefort, L.; Gauthier, G. Estimating moss growth in arctic conditions: A comparison of three methods. Bryologist 2010, 113, 322–332. [Google Scholar] [CrossRef]
- Clymo, R.S. Growth of sphagnum—Methods of measurement. J. Ecol. 1970, 58, 13. [Google Scholar] [CrossRef]
- Isaac, R.A.; Johnson, W.C. Determination of total nitrogen in plant-tissue, using a block digestor. J. Assoc. Off. Anal. Chem. 1976, 59, 98–100. [Google Scholar] [CrossRef]
- Nkonge, C.; Ballance, G.M. A sensitive colorimetric procedure for nitrogen determination in micro-kjeldahl digests. J. Agric. Food Chem. 1982, 30, 416–420. [Google Scholar] [CrossRef]
- Tandon, H.L.S.; Cescas, M.P.; Tyner, E.H. An acid-free vanadate-molybdate reagent for determination of total phosphorus in soils. Soil Sci. Soc. Am. Proc. 1968, 32, 48–51. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression; Sage Publications: Los Angeles, CA, USA, 2018. [Google Scholar]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef]
- Stuble, K.L.; Bewick, S.; Fisher, M.; Forister, M.L.; Harrison, S.P.; Shapir, A.M.; Latimer, A.M.; Fox, L.R. The promise and the perils of resurveying to understand global change impacts. Ecol. Monogr. 2021, 91, 14. [Google Scholar] [CrossRef]
- Holm, S. A simple sequentially rejective multiple test procedure. Scand. J. Stat. 1979, 6, 65–70. [Google Scholar]
- Team., R.C. R: A language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: Vienna, Austria; New York, NY, USA, 2009. [Google Scholar]
- Bates, D.; Machler, M.; Bolker, B.M.; Walker, S.C. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Okasen, F.; Blanchet, G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Wagner, H. Vegan: Community Ecology Package; CRAN.R-Project: Hesinki, Finalnd, 2015. [Google Scholar]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Gignac, L.D.; Vitt, D.H.; Zoltai, S.C.; Bayley, S.E. Bryophyte response surfaces along climatic, chemical, and physical gradients in peatlands of Western Canada. Nova Hedwig. 1991, 53, 27–71. [Google Scholar]
- Gordon, C.; Wynn, J.M.; Woodin, S.J. Impacts of increased nitrogen supply on high Arctic heath: The importance of bryophytes and phosphorus availability. New Phytol. 2001, 149, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Rustad, L.E.; Campbell, J.L.; Marion, G.M.; Norby, R.J.; Mitchell, M.J.; Hartley, A.E.; Cornelissen, J.H.C.; Gurevitch, J.; Gcte, N. A meta-analysis of the response of soil respiration, net nitrogen mineralization, and aboveground plant growth to experimental ecosystem warming. Oecologia 2001, 126, 543–562. [Google Scholar] [CrossRef] [PubMed]
- Oulehle, F.; Rowe, E.C.; Myska, O.; Chuman, T.; Evans, C.D. Plant functional type affects nitrogen use efficiency in high-Arctic tundra. Soil Biol. Biochem. 2016, 94, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Michelsen, A.; Graglia, E.; Schmidt, I.K.; Jonasson, S.; Sleep, D.; Quarmby, C. Differential responses of grass and a dwarf shrub to long-term changes in soil microbial biomass C, N and P following factorial addition of NPK fertilizer, fungicide and labile carbon to a heath. New Phytol. 1999, 143, 523–538. [Google Scholar] [CrossRef]
- Epstein, H.E.; Beringer, J.; Gould, W.A.; Lloyd, A.H.; Thompson, C.D.; Chapin, F.S.; Michaelson, G.J.; Ping, C.L.; Rupp, T.S.; Walker, D.A. The nature of spatial transitions in the Arctic. J. Biogeogr. 2004, 31, 1917–1933. [Google Scholar] [CrossRef]
- Nordin, A.; Schmidt, I.K.; Shaver, G.R. Nitrogen uptake by arctic soil microbes and plants in relation to soil nitrogen supply. Ecology 2004, 85, 955–962. [Google Scholar] [CrossRef]
- Tye, A.M.; Young, S.D.; Crout, N.M.J.; West, H.M.; Stapleton, L.M.; Poulton, P.R.; Laybourn-Parry, J. The fate of N-15 added to high Arctic tundra to mimic increased inputs of atmospheric nitrogen released from a melting snowpack. Glob. Chang. Biol. 2005, 11, 1640–1654. [Google Scholar] [CrossRef]
- Wust-Galley, C.; Volk, M.; Bassin, S. Interaction of climate change and nitrogen deposition on subalpine pastures. J. Veg. Sci. 2020, 12, e12946. [Google Scholar] [CrossRef]
- Bassin, S.; Schalajda, J.; Vogel, A.; Suter, M. Different types of sub-alpine grassland respond similarly to elevated nitrogen deposition in terms of productivity and sedge abundance. J. Veg. Sci. 2012, 23, 1024–1034. [Google Scholar] [CrossRef]
- Shane, M.W.; Cawthray, G.R.; Cramer, M.D.; Kuo, J.; Lambers, H. Specialized ‘dauciform’ roots of Cyperaceae are structurally distinct, but functionally analogous with ‘cluster’ roots. Plant Cell Environ. 2006, 29, 1989–1999. [Google Scholar] [CrossRef] [PubMed]
- Cusell, C.; Kooijman, A.; Lamers, L.P.M. Nitrogen or phosphorus limitation in rich fens?—Edaphic differences explain contrasting results in vegetation development after fertilization. Plant Soil 2014, 384, 153–168. [Google Scholar] [CrossRef]
- Verhoeven, J.T.A.; Schmitz, M.B. Control of plant-growth by nitrogen and phosphorus in mesotrophic fens. Biogeochemistry 1991, 12, 135–148. [Google Scholar] [CrossRef] [Green Version]
- Heijmans, M.; Berendse, F.; Arp, W.J.; Masselink, A.K.; Klees, H.; de Visser, W.; van Breemen, N. Effects of elevated carbon dioxide and increased nitrogen deposition on bog vegetation in the Netherlands. J. Ecol. 2001, 89, 268–279. [Google Scholar] [CrossRef]
- Limpens, J.; Berendse, F.; Klees, H. How phosphorus availability affects the impact of nitrogen deposition on Sphagnum and vascular plants in bogs. Ecosystems 2004, 7, 793–804. [Google Scholar] [CrossRef]
- Elmendorf, S.C.; Henry, G.H.R.; Hollister, R.D.; Bjork, R.G.; Bjorkman, A.D.; Callaghan, T.V.; Collier, L.S.; Cooper, E.J.; Cornelissen, J.H.C.; Day, T.A.; et al. Global assessment of experimental climate warming on tundra vegetation: Heterogeneity over space and time. Ecol. Lett. 2012, 15, 164–175. [Google Scholar] [CrossRef]
- Iversen, C.M.; Sloan, V.L.; Sullivan, P.F.; Euskirchen, E.S.; McGuire, A.D.; Norby, R.J.; Walker, A.P.; Warren, J.M.; Wullschleger, S.D. The unseen iceberg: Plant roots in arctic tundra. New Phytol. 2015, 205, 34–58. [Google Scholar] [CrossRef]
- Liu, X.; Wang, Z.; Li, X.M.; Bao, W.K. Nitrogen and phosphorus translocation of forest floor mosses as affected by a pulse of these nutrients. J. Plant Ecol. 2020, 13, 633–640. [Google Scholar] [CrossRef]
- Clymo, R.S. Ion exchange in Sphagnum and its relation to bog ecology. Ann. Bot. 1963, 27, 309–324. [Google Scholar] [CrossRef]
- Clarke, G.C.S.; Greene, S.W.; Greene, D.M. Productivity of bryophytes in polar regions. Ann. Bot. 1971, 35, 99–108. [Google Scholar] [CrossRef]
- Zechmeister, H.G. Growth rates of five pleurocarpous moss species under various climatic conditions. J. Bryol. 1995, 18, 455–468. [Google Scholar] [CrossRef]
- Asada, T.; Warner, B.G. Growth of mosses in relation to climate factors in a hypermaritime coastal peatland in British Columbia, Canada. Bryologist 2003, 106, 516–527. [Google Scholar] [CrossRef]
- Callaghan, T.V.; Carlsson, B.A.; Sonesson, M.; Temesvary, A. Between-year variation in climate-related growth of circumarctic populations of the moss Hylocomium splendens. Funct. Ecol. 1997, 11, 157–165. [Google Scholar] [CrossRef]
- Valéry, L.; Cadieux, M.C.; Gauthier, G. Spatial heterogeneity of primary production as both cause and consequence of foraging patterns of an expanding Greater Snow Goose colony. Ecoscience 2010, 17, 9–19. [Google Scholar] [CrossRef]
- Gauthier, G.; Bêty, J.; Giroux, J.-F.; Rochefort, L. Trophic interactions in a High Arctic Snow Goose Colony. Integr. Comp. Biol. 2004, 44, 119–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishizawa, K.; Deschamps, L.; Maire, V.; Bety, J.; Levesque, E.; Kitagawa, R.; Masumoto, S.; Gosselin, I.; Morneault, A.; Rochefort, L.; et al. Long-term consequences of goose exclusion on nutrient cycles and plant communities in the High-Arctic. Polar Sci. 2021, 27, 13. [Google Scholar] [CrossRef]
- Kuhnel, R.; Roberts, T.J.; Bjorkman, M.P.; Isaksson, E.; Aas, W.; Holmen, K.; Strom, J. 20-Year Climatology of NO3- and NH4+ Wet Deposition at Ny-Alesund, Svalbard. Adv. Meteorol. 2011, 2011, 406508. [Google Scholar] [CrossRef] [Green Version]
- Bilbrough, C.J.; Welker, J.M.; Bowman, W.D. Early spring nitrogen uptake by snow-covered plants: A comparison of arctic and alpine plant function under the snowpack. Arct. Antarct. Alp. Res. 2000, 32, 404–411. [Google Scholar] [CrossRef]
- Rasmussen, L.H.; Zhang, W.X.; Ambus, P.; Michelsen, A.; Jansson, P.E.; Kitzler, B.; Elberling, B. Nitrogen transport in a tundra landscape: The effects of early and late growing season lateral N inputs on arctic soil and plant N pools and N2O fluxes. Biogeochemistry 2022, 157, 69–84. [Google Scholar] [CrossRef]
- Myers-Smith, I.H.; Grabowski, M.M.; Thomas, H.J.D.; Angers-Blondin, S.; Daskalova, G.N.; Bjorkman, A.D.; Cunliffe, A.M.; Assmann, J.J.; Boyle, J.S.; McLeod, E.; et al. Eighteen years of ecological monitoring reveals multiple lines of evidence for tundra vegetation change. Ecol. Monogr. 2019, 89, 21. [Google Scholar] [CrossRef] [Green Version]
- Brouillet, L.; Coursol, F.; Meades, S.J.; Favreau, M.; Anions, M.; Bélisle, P.; Desmet, P. VASCAN, the Database of Vascular Plants of Canada; Canadensys: Ottawa, ON, Canada, 2010. [Google Scholar]
- Faubert, J.; Leclerc, S.; Anions, M.; D’Aoûst, M.; Favreau, M.; Higgins, K.; Lamond, M.; Lavoie, A.; Lapointe, M.; Nadeau, S.; et al. Base de Données des Bryophytes du Québec-Labrador (BRYOQUEL); Société Québecoise de Bryologie: Québec, QC, Canada, 2014. [Google Scholar]






| Fertilization Context | Nutrient Addition Treatment | N (g m−2 Year−1) | P (g m−2 Year−1) |
|---|---|---|---|
| Control | CT | 0 | 0 |
| N1 | 1 | 0 | |
| N addition | N3 | 3 | 0 |
| N5 | 5 | 0 | |
| N/P addition | NP3 | 3 | 0.5 |
| NP5 | 5 | 1 |
| Graminoid | Moss | |||||
|---|---|---|---|---|---|---|
| ANPP | ANPP | |||||
| F | P | df | F | P | df | |
| A. Fertilization effect after 17 years | ||||||
| Fertilization (F) | 3.9 | 0.0046 | 5, 54 | 0.8 | 0.0901 | 5, 25 |
| Family (Fa) | 4.8 | 0.0314 | 1, 54 | |||
| F × Fa | 3.7 | 0.0062 | 5, 54 | |||
| B. Temporal effect of fertilization | ||||||
| Fertilization (F) | 9.5 | <0.0001 | 5, 85 | 1.0 | 0.4075 | 5, 85 |
| Time (T) | 9.7 | 0.0002 | 2, 85 | 26.0 | <0.0001 | 2, 85 |
| F × T | 2.1 | 0.0356 | 10, 85 | 0.8 | 0.6008 | 10, 85 |
| C. Contrasts | t | P | df | t | P | df |
| Nitrogen linear | 2.4 | 0.0318 | 15 | 0.1 | 0.9874 | 15 |
| Control vs. N addition 1 | 0.5 | 0.6224 | 54 | 0.9 | 0.3630 | 25 |
| Control vs. N/P addition 2 | 2.5 | 0.0171 | 54 | 2.7 | 0.0136 | 25 |
| N addition vs. N/P addition | 2.7 | 0.0102 | 54 | 2.4 | 0.0246 | 25 |
| Fertilization Context | Treatment | Strata | Species | Variation (%) | t | P |
|---|---|---|---|---|---|---|
| Control | CT | Graminoid | Eriophorum scheuchzeri | +18.9 | 6.4 | <0.0001 |
| N addition | N1 | Moss | Scorpidium spp. | −50.5 | −7.5 | <0.0001 |
| Bryum pseudotriquetrum | −2.1 | −3.7 | 0.004 | |||
| Graminoid | Eriophorum scheuchzeri | +8.3 | 4.4 | 0.001 | ||
| N3 | Graminoid | Carex aquatilis | +15.1 | 5.1 | 0.0004 | |
| N5 | Moss | Scorpidium spp. | −43.1 | −8.4 | <0.0001 | |
| Graminoid | Carex aquatilis | +15.9 | 3.8 | 0.003 | ||
| Eriophorum scheuchzeri | +26.1 | 4.0 | 0.002 | |||
| N/P addition | NP3 | Moss | Aulacomnium acuminatum | +24.5 | 4.9 | 0.0005 |
| NP5 | Moss | Scorpidium spp. | −58.6 | −5.5 | 0.0002 |
| Fertilization Context | Treatment | Strata | Comparison | |||||
|---|---|---|---|---|---|---|---|---|
| Years 2–5 | Years 5–17 | Years 2–17 | ||||||
| Variation (%) | P | Variation (%) | P | Variation (%) | P | |||
| N addition | N5 | Moss | −7.4 | 0.1885 | −32.6 | 0.0016 * | −37.6 | 0.0003 * |
| N5 | Graminoid | −4.9 | 0.5589 | 43.1 | 0.2033 | 36.0 | 0.2259 | |
| N/P addition | NP5 | Moss | −3.4 | 0.3693 | −46.9 | <0.0001 * | −49.0 | <0.0001 * |
| NP5 | Graminoid | −20.9 | 0.0413 | 222.3 | <0.0001 * | 155.0 | <0.0001 * | |
| Nitrogen Concentration | Phosphorus Concentration | |||||
|---|---|---|---|---|---|---|
| F | P | df | F | P | df | |
| A. Fertilization effect after 17 years | ||||||
| Fertilization (F) | 24.4 | <0.0001 | 5, 32 | 2.2 | 0.3332 | 5, 32 |
| Family (Fa) | 1.1 | 0.3054 | 1, 32 | 5.1 | 0.0474 | 1, 32 |
| F × Fa | 1.5 | 0.2181 | 5, 32 | 0.05 | 0.6622 | 5, 32 |
| B. Temporal effect of fertilization | ||||||
| Fertilization (F) | 9.1 | <0.0001 | 5, 82 | 2.8 | 0.0209 | 5, 81 |
| Time (T) | 19.8 | <0.0001 | 2, 82 | 39.9 | <0.0001 | 2, 81 |
| F × T | 2.1 | 0.0328 * | 10, 82 | 1.4 | 0.2157 | 10, 81 |
| C. Contrasts | t | P | df | t | P | df |
| Nitrogen linear | 4.2 | 0.0003 | 22 | 0.2 | 0.9 | 22 |
| Control vs. N addition 1 | 1.7 | 0.0925 | 32 | 0.4 | 0.6782 | 32 |
| Control vs. N/P addition 2 | 1.8 | 0.0742 | 32 | 1.0 | 0.3261 | 32 |
| N addition vs. N/P addition | 0.7 | 0.4739 | 32 | 2.0 | 0.0489 | 32 |
| Nitrogen Concentration | Phosphorus Concentration | |||||
|---|---|---|---|---|---|---|
| F | P | df | F | P | df | |
| A. Fertilization effect after 17 years | ||||||
| Fertilization (F) | 9.0 | <0.0001 | 5, 19 | 16.5 | <0.0001 | 5, 19 |
| B. Temporal effect of fertilization | ||||||
| Fertilization (F) | 26.2 | <0.0001 | 5, 76 | 23.0 | <0.0001 | 5, 76 |
| Time (T) | 1.3 | 0.2631 | 2, 76 | 3.9 | 0.0245 | 2, 76 |
| F × T | 0.3 | 0.9665 | 10, 76 | 1.1 | 0.3894 | 10, 76 |
| C. Contrasts | t | P | df | t | P | df |
| Nitrogen linear | 3.2 | 0.0099 | 12 | 0.1 | 0.8284 | 12 |
| Control vs. N addition 1 | 1.7 | 0.0972 | 19 | 0.005 | 0.9849 | 19 |
| Control vs. N/P addition 2 | 3.1 | 0.0063 | 19 | 3.4 | 0.0028 | 19 |
| N addition vs. N/P addition | 3.4 | 0.0028 | 19 | 4.4 | 0.0003 | 19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gignac, C.; Rochefort, L.; Gauthier, G.; Lévesque, E.; Maire, V.; Deschamps, L.; Pouliot, R.; Marchand-Roy, M. N/P Addition Is More Likely Than N Addition Alone to Promote a Transition from Moss-Dominated to Graminoid-Dominated Tundra in the High-Arctic. Atmosphere 2022, 13, 676. https://doi.org/10.3390/atmos13050676
Gignac C, Rochefort L, Gauthier G, Lévesque E, Maire V, Deschamps L, Pouliot R, Marchand-Roy M. N/P Addition Is More Likely Than N Addition Alone to Promote a Transition from Moss-Dominated to Graminoid-Dominated Tundra in the High-Arctic. Atmosphere. 2022; 13(5):676. https://doi.org/10.3390/atmos13050676
Chicago/Turabian StyleGignac, Charles, Line Rochefort, Gilles Gauthier, Esther Lévesque, Vincent Maire, Lucas Deschamps, Rémy Pouliot, and Mylène Marchand-Roy. 2022. "N/P Addition Is More Likely Than N Addition Alone to Promote a Transition from Moss-Dominated to Graminoid-Dominated Tundra in the High-Arctic" Atmosphere 13, no. 5: 676. https://doi.org/10.3390/atmos13050676