
Rapid Sampling Protocol of Isoprene Emission Rate of Palm (Arecaceae) Species Using Excised Leaves

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
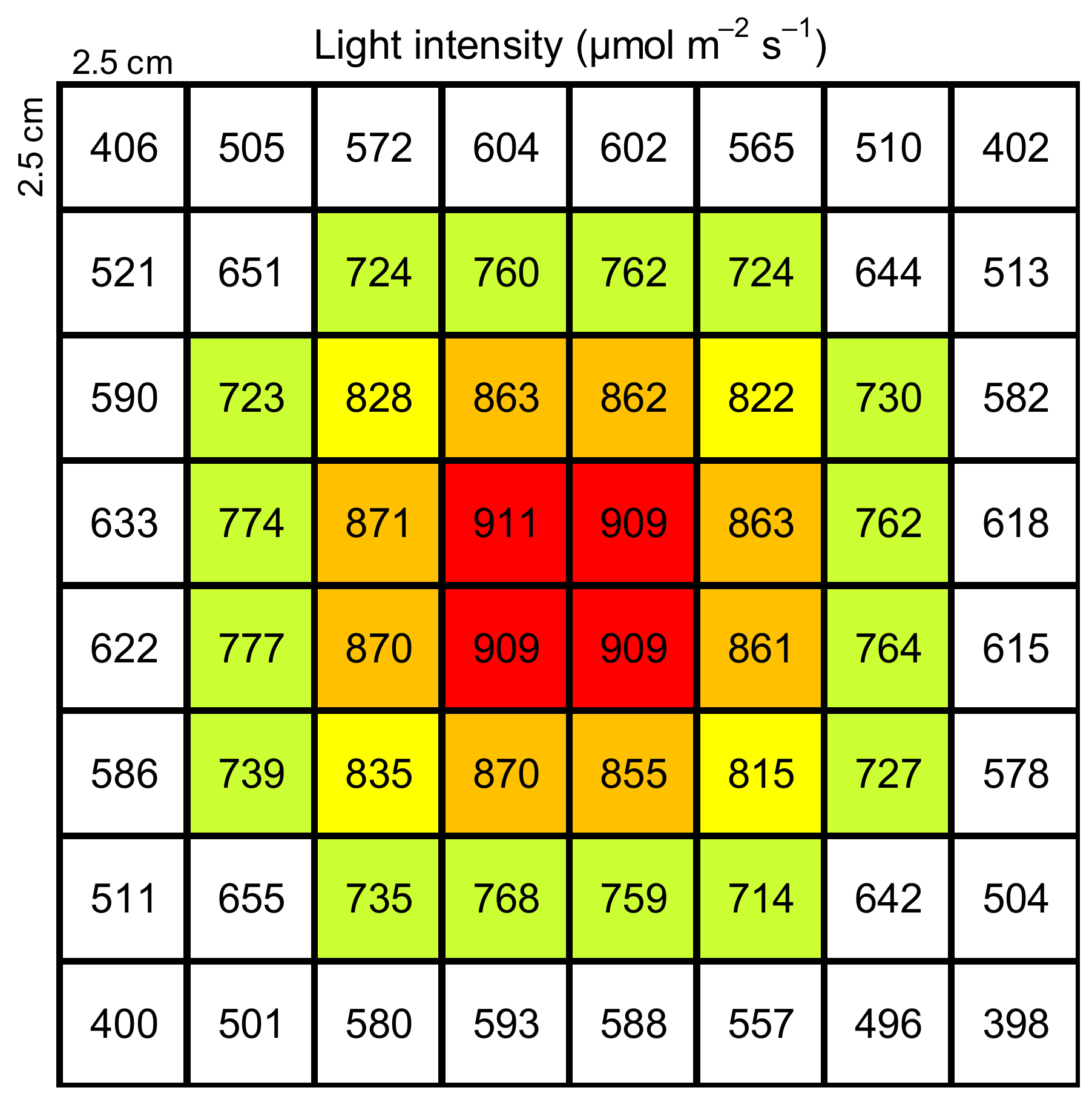
2.2. Measurement of Temperature Response of Isoprene Emission Rate Using Enclosure Method
2.3. Isoprene Emission Rate for Different Incubation Periods
2.4. Comparison of Measurement Using Excised Leaves and Measurement Based on Enclosure Method
2.5. Gas Chromatography Analysis
2.6. Calculation of Gas Exchange and Carbon Ratio
3. Results
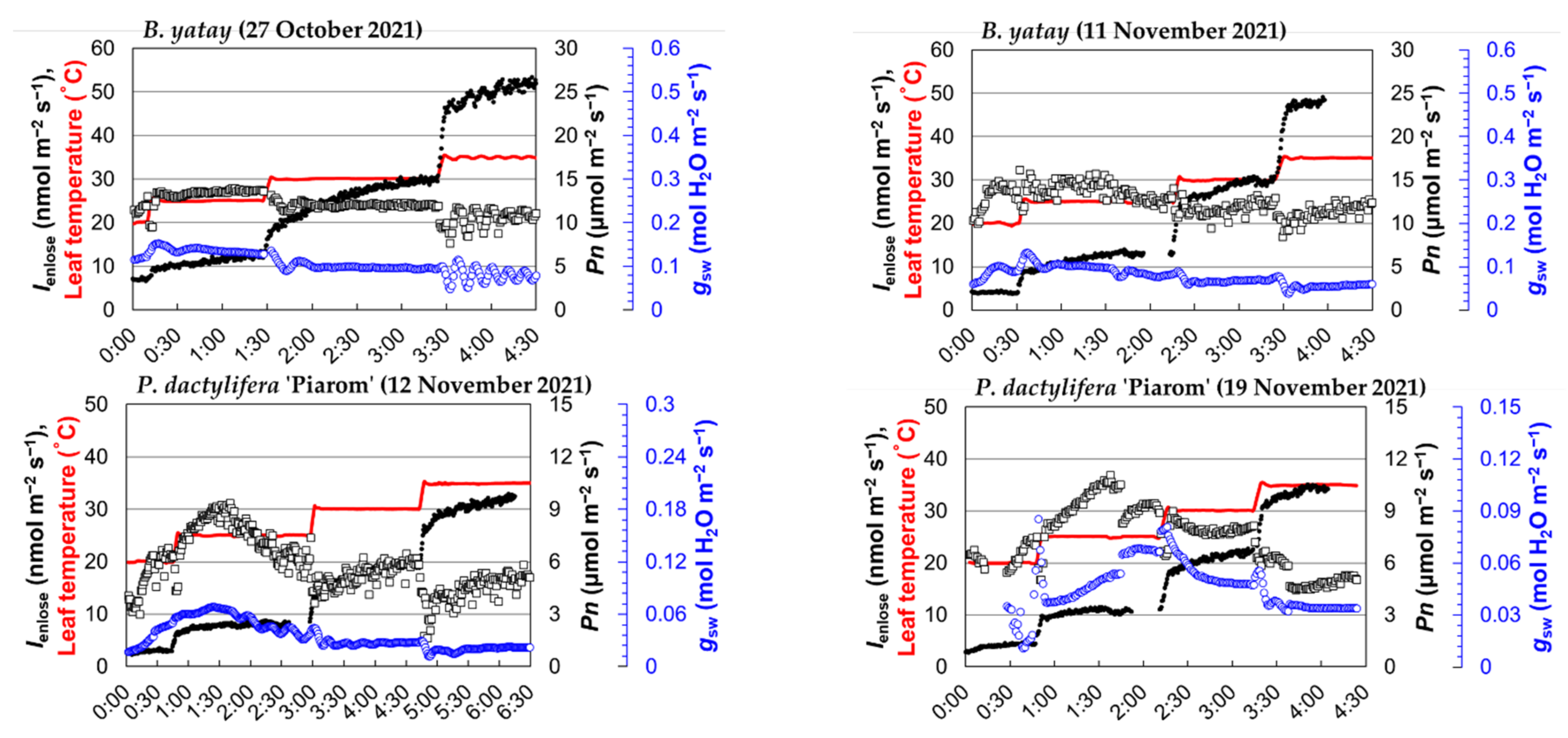
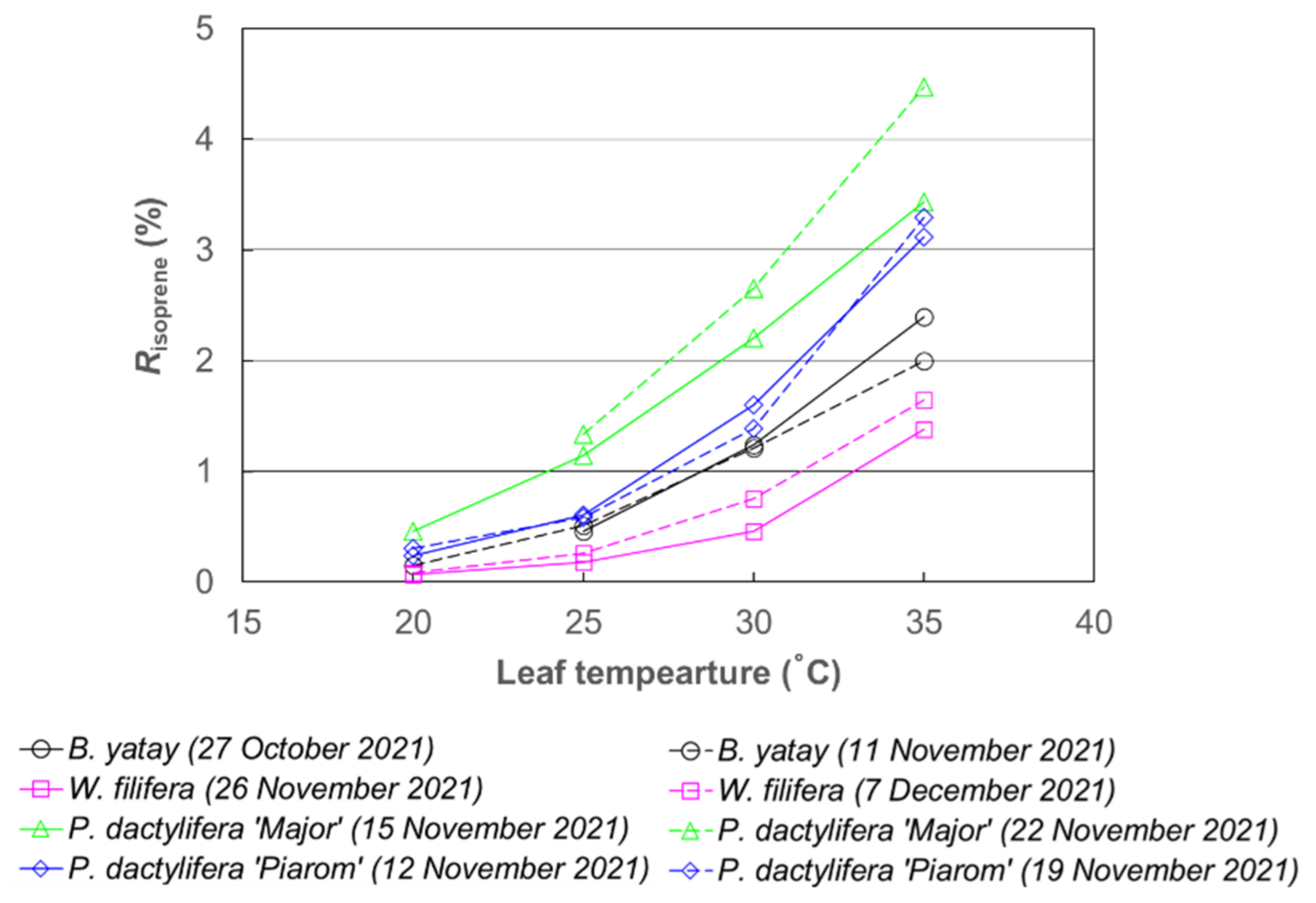
3.1. Response of Isoprene Emission Rate to Leaf Temperature
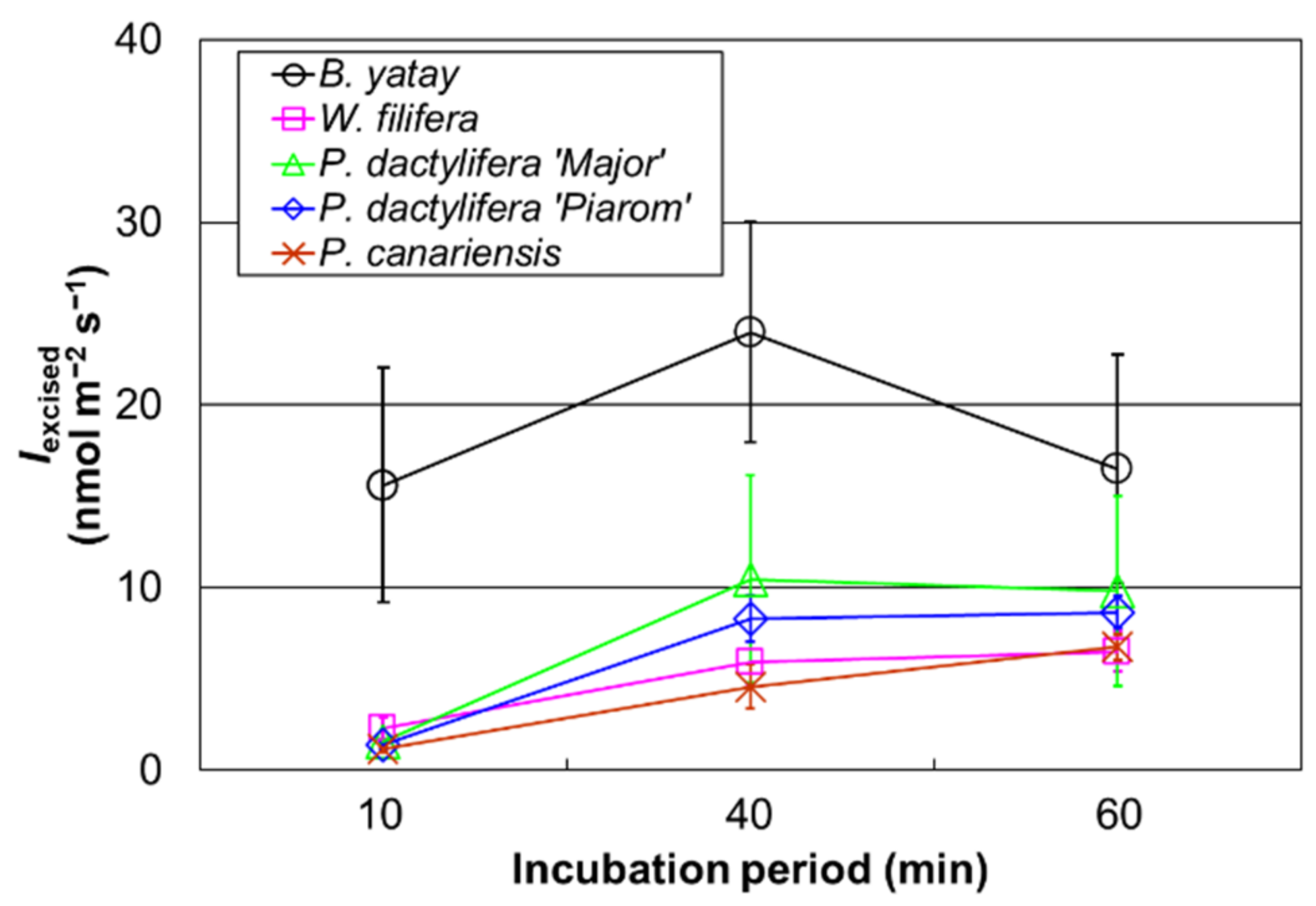
3.2. Isoprene Emission Rate for Different Incubation Periods
3.3. Comparison of Isoprene Emission Rates Obtained Using Excised Leaves and Enclosure Method
4. Discussion
4.1. Temperature Response of Isoprene Emission Rate of Palm Species
4.2. Determination of Appropriate Incubation Period for Isoprene Measurement Using Excised Palm Leaves
4.3. Suitability of Measurement Using Excised Leaves to Evaluate Isoprene Emission from Palm Leaves
4.4. Development of Method Using Excised Leaves to Measure Isoprene Emission Rate
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guenther, A.B.; Jiang, X.; Heald, C.L.; Sakulyanontvittaya, T.; Duhl, T.; Emmons, L.K.; Wang, X. The Model of Emissions of Gases and Aerosols from Nature Version 2.1 (MEGAN2.1): An Extended and Updated Framework for Modeling Biogenic Emissions. Geosci. Model Dev. 2012, 15, 1471–1492. [Google Scholar] [CrossRef] [Green Version]
- Sindelarova, K.; Granier, C.; Bouarar, I.; Guenther, A.; Tilmes, S.; Stavrakou, T.; Müller, J.-F.; Kuhn, U.; Stefani, P.; Knorr, W. Global data set of biogenic VOC emissions calculated by the MEGAN model over the last 30 years. Atmos. Chem. Phys. 2014, 14, 9317–9341. [Google Scholar] [CrossRef] [Green Version]
- Saunois, M.; Stavert, A.R.; Poulter, B.; Bousquet, P.; Canadell, J.G.; Jackson, R.B.; Raymond, P.A.; Dlugokencky, E.J.; Houweling, S.; Patra, P.K.; et al. The Global Methane Budget 2000–2017. Earth Syst. Sci. Data 2020, 12, 1561–1623. [Google Scholar] [CrossRef]
- Huang, G.; Brook, R.; Crippa, M.; Janssens-Maenhout, G.; Schieberle, C.; Dore, C.; Guizzardi, D.; Muntean, M.; Schaaf, E.; Friedrich, R. Speciation of anthropogenic emissions of non-methane volatile organic compounds: A global gridded data set for 1970–2012. Atmos. Chem. Phys. 2017, 17, 7683–7701. [Google Scholar] [CrossRef] [Green Version]
- Teng, A.P.; Crounse, J.D.; Wennberg, P.O. Isoprene Peroxy Radical Dynamics. JACS 2017, 139, 5367–5377. [Google Scholar] [CrossRef] [Green Version]
- Tani, A.; Mochizuki, T. Review: Exchanges of volatile organic compounds between terrestrial ecosystems and the atmosphere. J. Agric. Meteorol. 2021, 77, 66–80. [Google Scholar] [CrossRef]
- Dunker, A.M.; Koo, B.; Yarwood, G. Ozone sensitivity to isoprene chemistry and emissions and anthropogenic emissions in central California. Atmos. Environ. 2016, 145, 326–337. [Google Scholar] [CrossRef]
- Hewitt, C.N.; MacKenzie, A.R.; Di Carlo, P.; Di Marco, C.F.; Dorsey, J.R.; Evans, M.; Fowler, D.; Gallagher, M.W.; Hopkins, J.R.; Jones, C.E.; et al. Nitrogen management is essential to prevent tropical oil palm plantations from causing ground-level ozone pollution. Proc. Natl. Acad. Sci. USA 2009, 106, 18447–18451. [Google Scholar] [CrossRef] [Green Version]
- Duane, M.; Poma, B.; Rembges, D.; Astorga, C.; Larsen, B.R. Isoprene and its degradation products as strong ozone precursors in Insubria, Northern Italy. Atmos. Environ. 2002, 36, 3867–3879. [Google Scholar] [CrossRef]
- Pang, X.; Mu, Y.; Zhang, Y.; Lee, X.; Yuan, J. Contribution of isoprene to formaldehyde and ozone formation based on its oxidation products measurement in Beijing, China. Atmos. Environ. 2009, 43, 2142–2147. [Google Scholar] [CrossRef]
- Kaser, L.; Peron, A.; Graus, M.; Striednig, M.; Wohlfahrt, G.; Juráň, S.; Karl, T. Interannual Variability of BVOC Emissions in an Alpine City. Atmos. Chem. Phys. Discuss. 2021, 22, 5603–5618. [Google Scholar] [CrossRef]
- Fowler, D.; Amann, M.; Anderson, R.; Ashmore, M.; Cox, P.; Depledge, M.; Derwent, D.; Grennfelt, P.; Hewitt, N.; Hov, O.; et al. Ground-Level Ozone in the 21st Century: Future Trends, Impacts and Policy Implications, RS1276 ed.; The Royal Society: London, UK, 2008; Volume 15/08, pp. 57–66. [Google Scholar]
- Agyei, T.; Juráň, S.; Edwards-Jonášová, M.; Fischer, M.; Švik, M.; Komínková, K.; Ofori-Amanfo, K.K.; Marek, M.V.; Grace, J.; Urban, O. The Influence of Ozone on Net Ecosystem Production of a Ryegrass-Clover Mixture under Field Conditions. Atmosphere 2021, 12, 1629. [Google Scholar] [CrossRef]
- Kroll, J.H.; Ng, N.L.; Murphy, S.M.; Flagan, R.C.; Seinfeld, J.H. Secondary organic aerosol formation from isoprene photooxidation under high-NOx conditions. Geophys. Res. Lett. 2005, 32, L18808. [Google Scholar] [CrossRef] [Green Version]
- Kroll, J.H.; Ng, N.L.; Murphy, S.M.; Flagan, R.C.; Seinfeld, J.H. Secondary organic aerosol formation from isoprenephotooxidation. Environ. Sci. Technol. 2006, 40, 1869–1877. [Google Scholar] [CrossRef] [Green Version]
- Collins, W.J.; Derwent, R.G.; Johnson, C.E.; Stevenson, D.S. The Oxidation of Organic Compounds in the Troposphere and their Global Warming Potentials. Clim. Change 2002, 52, 453–479. [Google Scholar] [CrossRef]
- Pike, R.C.; Young, P.J. How plants can influence tropospheric chemistry: The role of isoprene emissions from the biosphere. Weather 2009, 64, 332–336. [Google Scholar] [CrossRef] [Green Version]
- Archibald, A.T.; Levine, J.G.; Abraham, N.L.; Cooke, M.C.; Edwards, P.M.; Heard, D.E.; Jenkin, M.E.; Karunaharan, A.; Pike, R.C.; Monks, P.S.; et al. Impacts of HOx regeneration and recycling in the oxidation of isoprene: Consequences for the composition of past, present and future atmospheres. Geophys. Res. Lett. 2011, 38, L05804. [Google Scholar] [CrossRef] [Green Version]
- Guenther, A.B.; Zimmerman, P.R.; Harley, P.C.; Monson, R.K.; Fall, R. Isoprene and monoterpene emission rate variability: Model evaluations and sensitivity analyses. J. Geophys. Res. 1993, 98, 12609–12617. [Google Scholar] [CrossRef] [Green Version]
- Guenther, A.; Zimmerman, P.R.; Klinger, L.; Greenberg, J.P.; Ennis, C.; Davis, K.; Pollock, W.; Westberg, H.; Allwine, G.; Geron, C.D. Estimates of regional natural volatile organic compound fluxes from enclosure and ambient measurements. J. Geophys. Res. 1996, 101, 1345–1359. [Google Scholar] [CrossRef] [Green Version]
- Tani, A.; Fushimi, K. Effects of temperature and light intensity on isoprene emission of Edgeworthia chrysantha. J. Agric. Meteorol. 2005, 61, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Tani, A.; Kawawata, Y. Isoprene emission from the major native Quercus spp. in Japan. Atmos. Environ. 2008, 42, 4540–4550. [Google Scholar] [CrossRef]
- Okumura, M.; Kosugi, Y.; Tani, A. Biogenic volatile organic compound emissions from bamboo species in Japan. J. Agric. Meteorol. 2018, 74, 40–44. [Google Scholar] [CrossRef] [Green Version]
- Chang, T.; Kume, T.; Okumura, M.; Kosugi, Y. Characteristics of isoprene emission from moso bamboo leaves in a forest in central Taiwan. Atmos. Environ. 2019, 211, 288–295. [Google Scholar] [CrossRef]
- Chang, T.W.; Kosugi, Y.; Okumura, M.; Jiao, L.; Chen, S.; Xu, D.; Liu, Z.; Shibata, S.; Chang, K.H. Isoprene emission characteristics of tall and dwarf bamboos. Atmos. Environ. X 2021, 12, 100136. [Google Scholar] [CrossRef]
- Day, M.E. Influence of temperature and leaf-to-air vapor pressure deficit on net photosynthesis and stomatal conductance in red spruce (Picea rubens). Tree Physiol. 2000, 20, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Pimentel, C.; Rafael, V.R.; Santos, M.G.; Oliveira, R.F.; Machado, E.C. Effects of changes in the photosynthetic photon flux density on net gas exchange of Citrus limon and Nicotiana tabacum. Braz. J. Plant Physiol. 2004, 16, 77–82. [Google Scholar] [CrossRef]
- Tanaka, Y.; Shiraiwa, T.; Nakajima, A.; Sato, J.; Nakazaki, T. Leaf Gas Exchange Activity in Soybean as Related to Leaf Traits and Stem Growth Habit. Crop Sci. 2008, 48, 1925–1932. [Google Scholar] [CrossRef]
- Tanaka, Y. Measurements and its Applications of Leaf Gas Exchange. Jpn. J. Crop Sci. 2016, 85, 339–346. [Google Scholar] [CrossRef] [Green Version]
- Baker, W.J.; Dransfield, J. Beyond Genera Palmarum: Progress and prospects in palm systematics. Bot. J. Linn. 2016, 182, 207–233. [Google Scholar] [CrossRef] [Green Version]
- Eiserhardt, W.L.; Svenning, J.C.; Kissling, W.D.; Balslev, H. Geographical ecology of the palms (Arecaceae): Determinants of diversity and distributions across spatial scales. Ann. Bot. 2011, 108, 1391–1416. [Google Scholar] [CrossRef] [Green Version]
- Tomlinson, P.B. The uniqueness of palms. Bot. J. Linn. 2006, 151, 5–14. [Google Scholar] [CrossRef]
- Harley, P.C.; Monson, R.K.; Lerdau, M.T. Ecological and evolutionary aspects of isoprene emission from plants. Oecologia 1999, 118, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Dani, K.G.S.; Jamie, I.M.; Prentice, I.C.; Atwell, B.J. Evolution of isoprene emission capacity in plants. Trends Plant Sci. 2014, 19, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, T.B.; Baker, C.R.; Walker, A.P.; McDowell, N.; Alistair Rogers, A.; Higuchi, N.; Chambers, J.Q.; Jardine, K.J. Stimulation of isoprene emissions and electron transport rates as key mechanisms of thermal tolerance in the tropical species Vismia guianensis. Glob. Change. Biol. 2020, 26, 5928–5941. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, M.T.; Sudol, M.; Bloch, L.; Winer, A.M. Low-emitting urban forests: A taxonomic methodology for assigning isoprene and monoterpenes emission rates. Atmos. Environ. 1996, 30, 1437–1452. [Google Scholar] [CrossRef]
- Geron, C.; Owen, S.; Guenther, A.; Greenberg, J.; Rasmussen, R.; Bai, J.H.; Li, Q.J.; Baker, B. Volatile organic compounds from vegetation in southern Yunnan Province, China: Emission rates and some potential regional implications. Atmos. Environ. 2006, 40, 1759–1773. [Google Scholar] [CrossRef] [Green Version]
- Jardine, K.J.; Zorzanelli, R.F.; Gimenez, B.O.; Oliveira Piva, L.R.D.; Teixeira, A.; Fontes, C.G.; Robles, E.; Higuchi, N.; Chambers, J.Q.; Martin, S.T. Leaf isoprene and monoterpene emission distribution across hyperdominant tree genera in the Amazon basin. Phytochemistry 2020, 175, 112366. [Google Scholar] [CrossRef]
- Barfod, A.S.; Balhara, M.; Dransfield, J.; Balslev, H. SE Asian Palms for Agroforestry and Home Gardens. Forests 2015, 6, 4607–4616. [Google Scholar] [CrossRef] [Green Version]
- Food and Agriculture Organization of the United Nations (FAO) FAOSTAT. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 2 April 2022).
- Litvak, M.E.; Loreto, F.; Harley, P.C.; Sharkey, T.D.; Monson, R.K. The response of isoprene emission rate and photosynthetic rate to photon flux and nitrogen supply in aspen and white oak trees. Plant Cell Environ. 1996, 19, 549–559. [Google Scholar] [CrossRef]
- Geron, C.; Nie, D.; Arnts, R.R.; Sharkey, T.D.; Singsaas, E.L.; Vanderveer, P.J.; Guenther, A.; Sickles, J.E.; Kleindienst, T.E. Biogenic isoprene emission: Model evaluation in a southeastern United States bottomland deciduous forest. J. Geophys. Res. 1997, 102, 18889–18901. [Google Scholar] [CrossRef] [Green Version]
- Geron, C.; Harley, P.; Guenther, A. Isoprene emission capacity for US tree species. Atmos. Environ. 2001, 35, 3341–3352. [Google Scholar] [CrossRef] [Green Version]
- Arab, L.; Kreuzwieser, J.; Kruse, J.; Zimmer, I.; Ache, P.; Alfarraj, S.; Al-Rasheid, K.A.S.; Schnitzler, J.; Hedrich, R.; Rennenberg, H. Acclimation to heat and drought—Lessons to learn from the date palm (Phoenix dactylifera). Environ. Exp. Bot. 2016, 125, 20–30. [Google Scholar] [CrossRef] [Green Version]
- Harley, P.; Guenther, A.; Zimmerman, P. Environmental controls over isoprene emission in deciduous oak canopies. Tree Physiol. 1997, 17, 705–714. [Google Scholar] [CrossRef] [Green Version]
- Hanson, D.T.; Sharkey, T.D. Effect of growth conditions on isoprene emission and other thermotolerance-enhancing compounds. Plant Cell Environ. 2001, 24, 929–936. [Google Scholar] [CrossRef]
- Brilli, F.; Dani, K.G.S.; Pasqualini, S.; Costarelli, A.; Cannavò, S.; Paolocci, F.; Zittelli, G.C.; Mugnai, G.; Baraldi, R.; Loreto, F. Exposure to different light intensities affects emission of volatiles and accumulations of both pigments and phenolics in Azolla filiculoides. Physiol. Plant. 2022, 174, e13619. [Google Scholar] [CrossRef]
- Tingey, D.T.; Manning, M.; Grothaus, L.C.; Burns, W.F. The influence of light and temperature on isoprene emission rates from live oak. Physiol. Plant. 1979, 47, 112–118. [Google Scholar] [CrossRef]
- Guenther, A.B.; Monson, R.K.; Fall, R. Isoprene and monoterpene emission rate variability: Observations with eucalyptus and emission rate algorithm development. J. Geophys. Res. 1991, 96, 10799–10808. [Google Scholar] [CrossRef]
- Sharkey, T.D.; Loreto, F. Water stress, temperature, and light effects on the capacity for isoprene emission and photosynthesis of kudzu leaves. Oecologia 1993, 95, 328–333. [Google Scholar] [CrossRef]
- Rasulov, B.; Bichele, I.; Hüve, K.; Vislap, V.; Niinemets, Ü. Acclimation of isoprene emission and photosynthesis to growth temperature in hybrid aspen: Resolving structural and physiological controls. Plant Cell Environ. 2015, 38, 751–766. [Google Scholar] [CrossRef] [Green Version]
- Monson, R.K.; Fall, R. Isoprene Emission from Aspen Leaves: Influence of Environment and Relation to Photosynthesis and Photorespiration. Plant Physiol. 1989, 90, 267–274. [Google Scholar] [CrossRef] [Green Version]
- Fall, R.; Monson, R.K. Isoprene emission rate and intercellular isoprene concentration as influenced by stomatal distribution and conductance. Plant Physiol. 1992, 100, 987–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasulov, B.; Hüve, K.; Bichele, I.; Laisk, A.; Niinemets, Ü. Temperature Response of Isoprene Emission in Vivo Reflects a Combined Effect of Substrate Limitations and Isoprene Synthase Activity: A Kinetic Analysis. Plant Physiol. 2010, 154, 1558–1570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tezara, W.; Mitchell, V.J.; Driscoll, S.D.; Lawlor, D.W. Water stress inhibits plant photosynthesis by decreasing coupling factor and ATP. Nature 1999, 401, 914–917. [Google Scholar] [CrossRef]
- Wildermuth, M.C.; Fall, R. Light-Dependent Isoprene Emission (Characterization of a Thylakoid-Bound Isoprene Synthase in Salix discolor Chloroplasts). Plant Physiol. 1996, 112, 171–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pétron, G.; Harley, P.; Greenberg, J.; Guenther, A. Seasonal temperature variations influence isoprene emission. Geophys. Res. Lett. 2001, 28, 1707–1710. [Google Scholar] [CrossRef]
- Rasulov, B.; Hüve, K.; Valbe, M.; Laisk, A.; Niinemets, U. Evidence That Light, Carbon Dioxide, and Oxygen Dependencies of Leaf Isoprene Emission Are Driven by Energy Status in Hybrid Aspen. Plant Physiol. 2009, 151, 448–460. [Google Scholar] [CrossRef] [Green Version]
- Müller, P.; Li, X.P.; Niyogi, K.K. Non-Photochemical Quenching. A Response to Excess Light Energy. Plant Physiol. 2001, 125, 1558–1566. [Google Scholar] [CrossRef] [Green Version]
- Kuzma, J.; Fall, R. Leaf Isoprene Emission Rate Is Dependent on Leaf Development and the Level of Isoprene Synthase. Plant Physiol. 1993, 101, 435–440. [Google Scholar] [CrossRef] [Green Version]
- Logan, B.A.; Monson, R.K. Thermotolerance of Leaf Discs from Four Isoprene-Emitting Species Is Not Enhanced by Exposure to Exogenous Isoprene. Plant Physiol. 1999, 120, 821–826. [Google Scholar] [CrossRef] [Green Version]
- Loreto, F.; Mannozzi, M.; Maris, C.; Nascetti, P.; Ferranti, F.; Pasqualini, S. Ozone Quenching Properties of Isoprene and Its Antioxidant Role in Leaves. Plant Physiol. 2001, 126, 993–1000. [Google Scholar] [CrossRef] [Green Version]
- Loreto, F.; Sharkey, T.D. Isoprene emission by plants is affected by transmissible wound signals. Plant Cell Environ. 1993, 16, 563–570. [Google Scholar] [CrossRef]
- Loreto, F.; Barta, C.; Brilli, F.; Nogues, I. On the induction of volatile organic compound emissions by plants as consequence of wounding or fluctuations of light and temperature. Plant Cell Environ. 2006, 29, 1820–1828. [Google Scholar] [CrossRef]
- Monson, R.K.; Harley, P.C.; Litvak, M.E.; Wildermuth, M.; Guenther, A.B.; Zimmerman, P.R.; Fall, R. Environmental and developmental controls over the seasonal pattern of isoprene emission from aspen leaves. Oecologia 1994, 99, 260–270. [Google Scholar] [CrossRef]
- Rasulov, B.; Copolovici, L.; Laisk, A.; Niinemets, Ü. Postillumination Isoprene Emission: In Vivo Measurements of Dimethylallyldiphosphate Pool Size and Isoprene Synthase Kinetics in Aspen Leaves. Plant Physiol. 2009, 149, 1609–1618. [Google Scholar] [CrossRef] [Green Version]
- Van Gardingen, P.R.; Grace, J. Vapour Pressure Deficit Response of Cuticular Conductance in Intact Leaves of Fagus sylvatica L. J. Exp. Bot. 1992, 43, 1293–1299. [Google Scholar] [CrossRef]
- Hoad, S.P.; Grace, J.; Jeffree, C.E. A leaf disc method for measuring cuticular conductance. J. Exp. Bot. 1996, 47, 431–437. [Google Scholar] [CrossRef]
- Schuster, A.; Burghardt, M.; Alfarhan, A.; Bueno, A.; Hedrich, R.; Leide, J.; Thomas, J.; Riederer, M. Effectiveness of cuticular transpiration barriers in a desert plant at controlling water loss at high temperatures. AoB Plants 2016, 8, plw027. [Google Scholar] [CrossRef]
- Bueno, A.; Alfarhan, A.; Arand, K.; Burghardt, M.; Deininger, A.; Hedrich, R.; Leide, J.; Seufert, P.; Staiger, S.; Riederer, M. Effects of temperature on the cuticular transpiration barrier of two desert plants with water-spender and water-saver strategies. J. Exp. Bot. 2019, 70, 1613–1625. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, T.-W.; Okamoto, H.; Tani, A. Rapid Sampling Protocol of Isoprene Emission Rate of Palm (Arecaceae) Species Using Excised Leaves. Atmosphere 2022, 13, 778. https://doi.org/10.3390/atmos13050778
Chang T-W, Okamoto H, Tani A. Rapid Sampling Protocol of Isoprene Emission Rate of Palm (Arecaceae) Species Using Excised Leaves. Atmosphere. 2022; 13(5):778. https://doi.org/10.3390/atmos13050778
Chicago/Turabian StyleChang, Ting-Wei, Hiroshi Okamoto, and Akira Tani. 2022. "Rapid Sampling Protocol of Isoprene Emission Rate of Palm (Arecaceae) Species Using Excised Leaves" Atmosphere 13, no. 5: 778. https://doi.org/10.3390/atmos13050778