
Meteorological Influences on Short-Term Carbon-Water Relationships in Two Forests in Subtropical China


Abstract
1. Introduction
2. Data and Methods
2.1. Study Sites
2.2. Data and Data Preprocessing
2.3. Statistical Methods
3. Results and Discussion
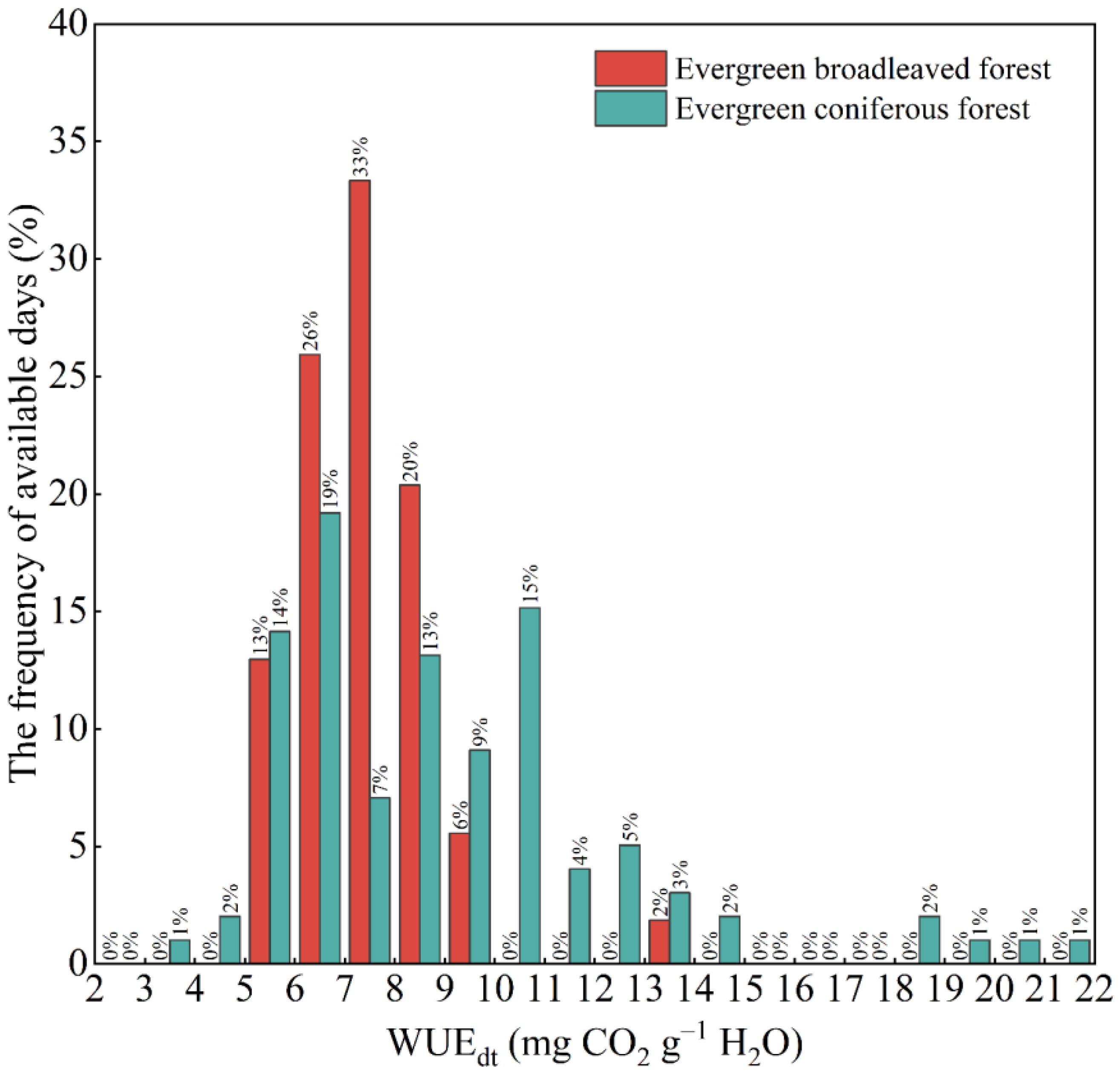
3.1. Daily Water Use Efficiency
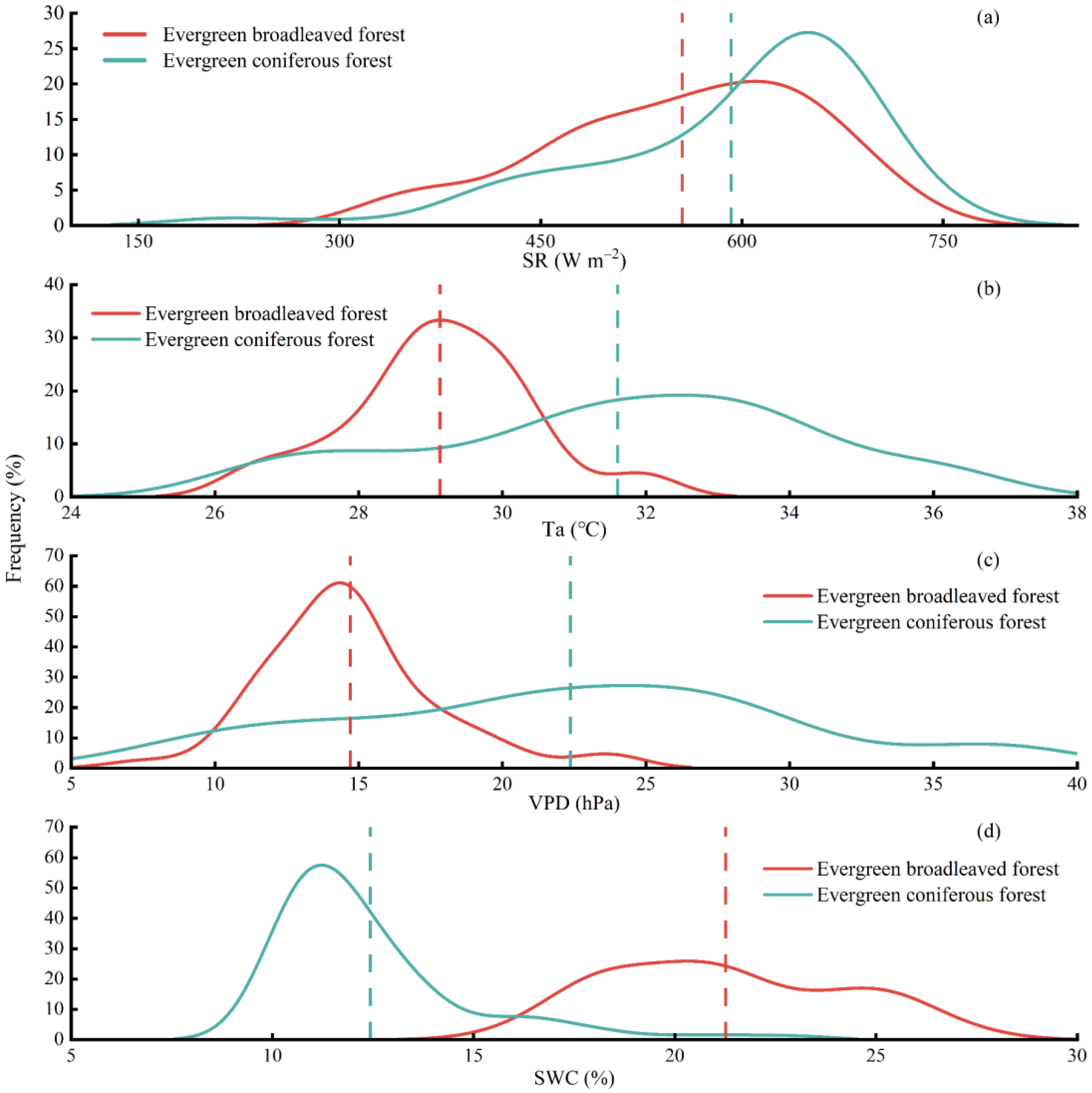
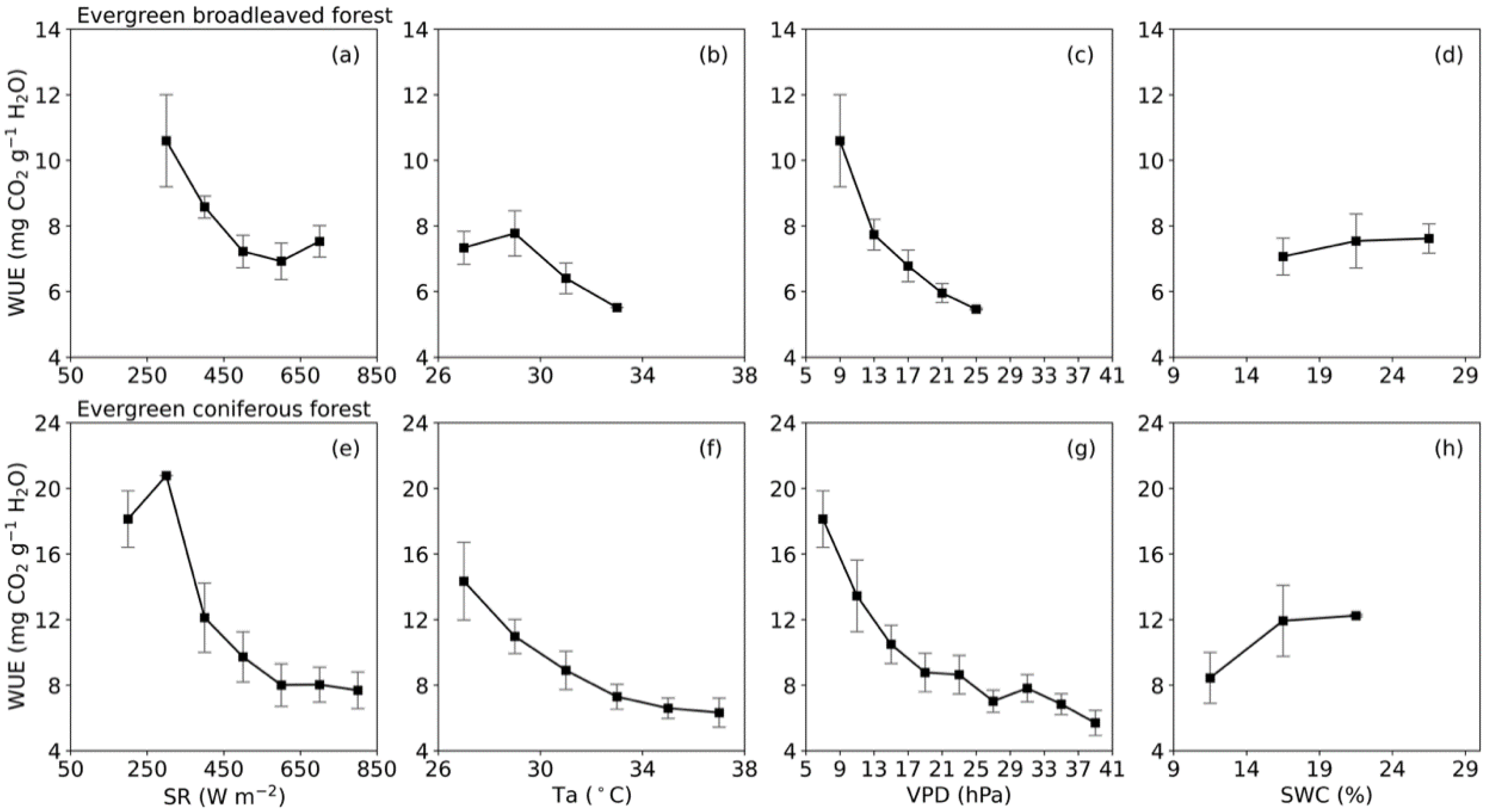
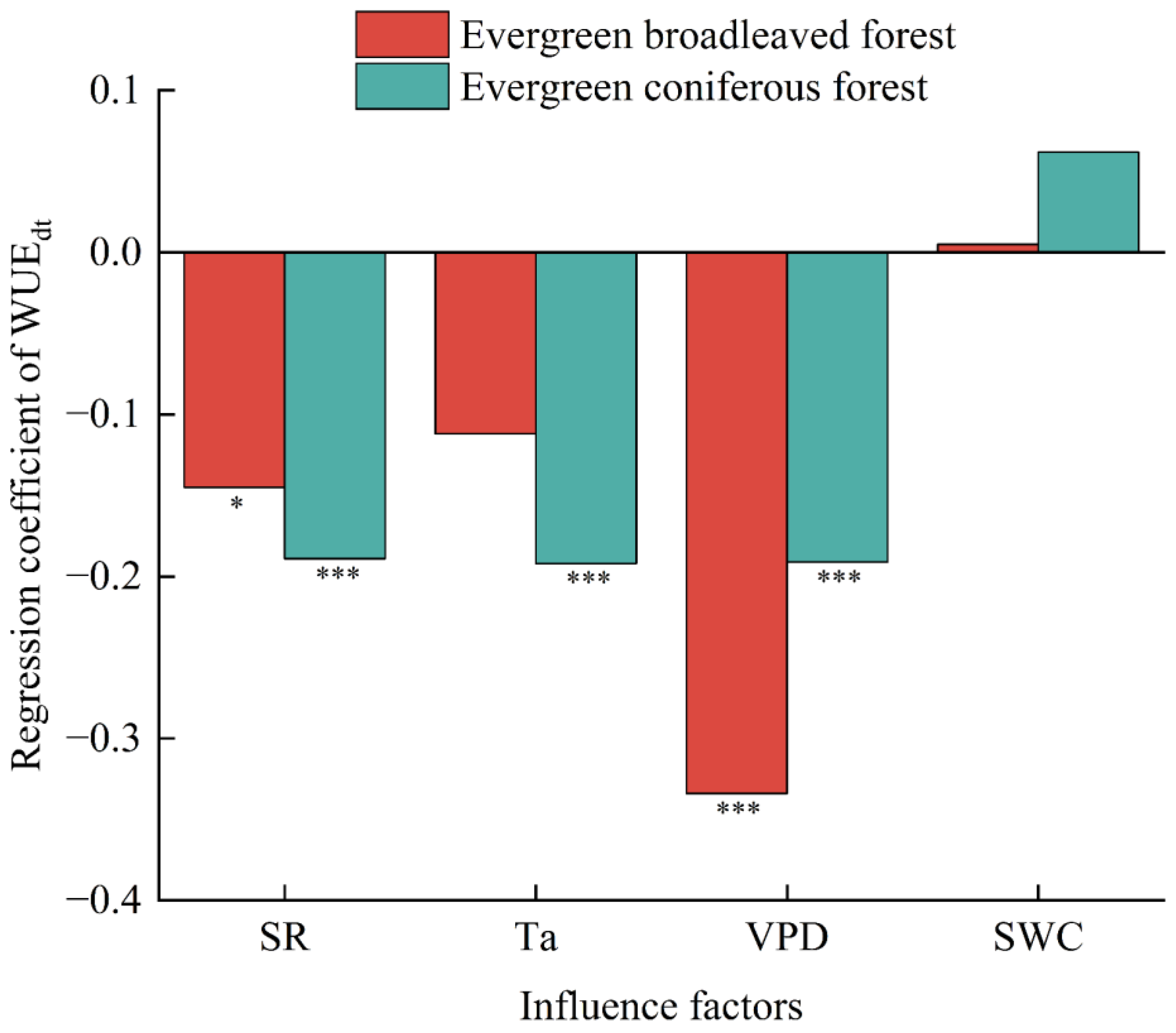
3.2. Meteorological Influences on Daily Water Use Efficiency
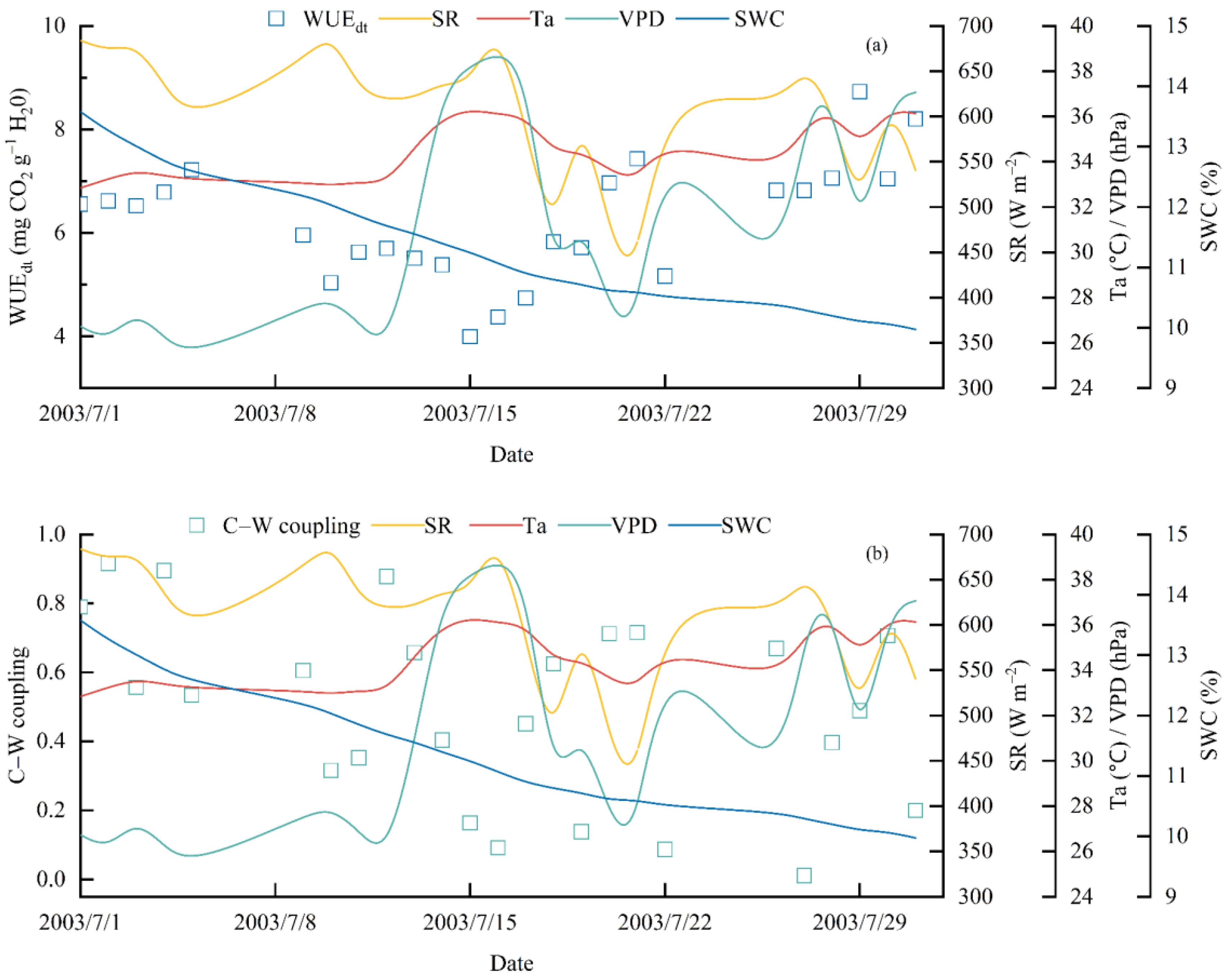
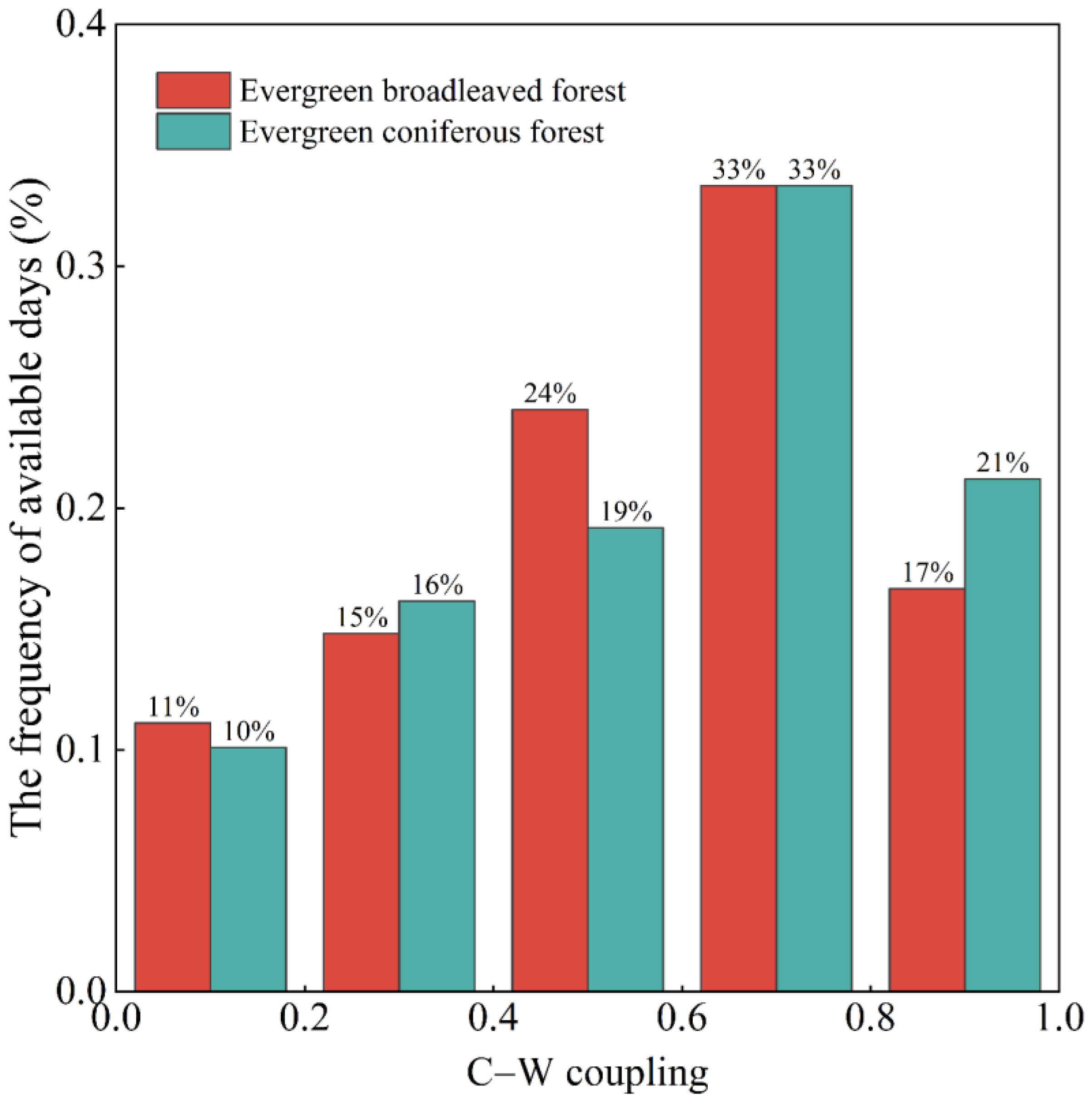
3.3. GPP–ET Coupling
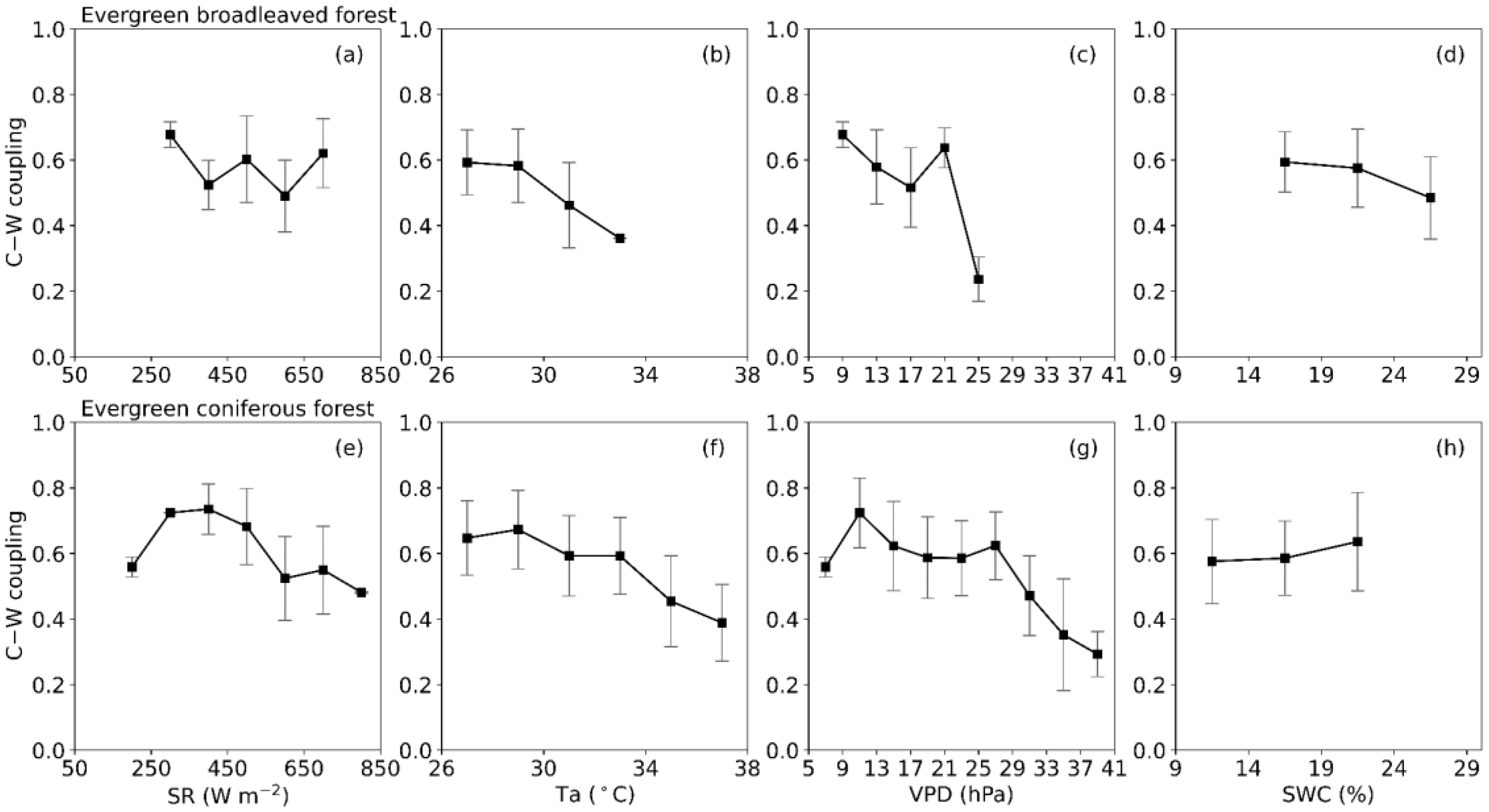
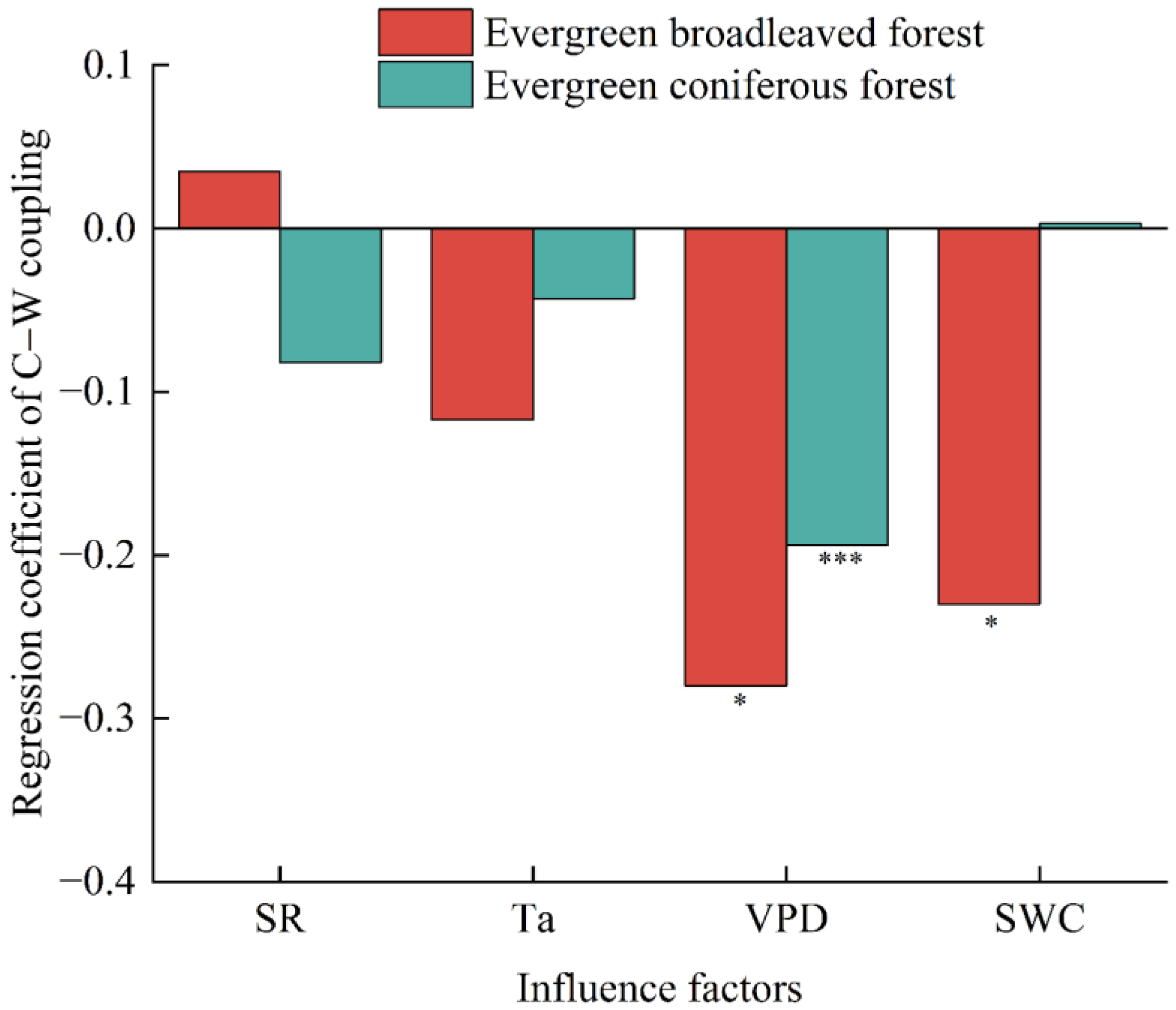
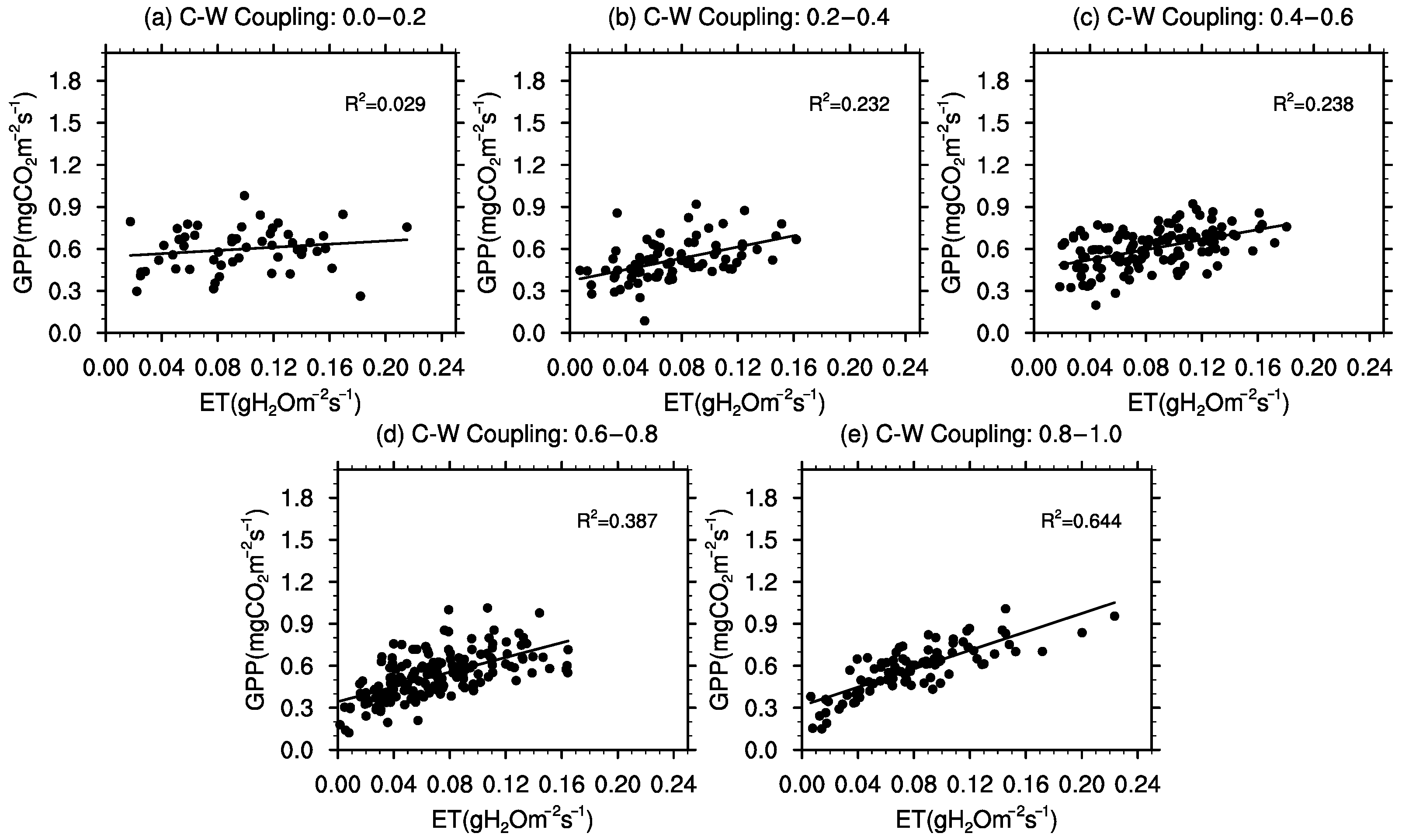
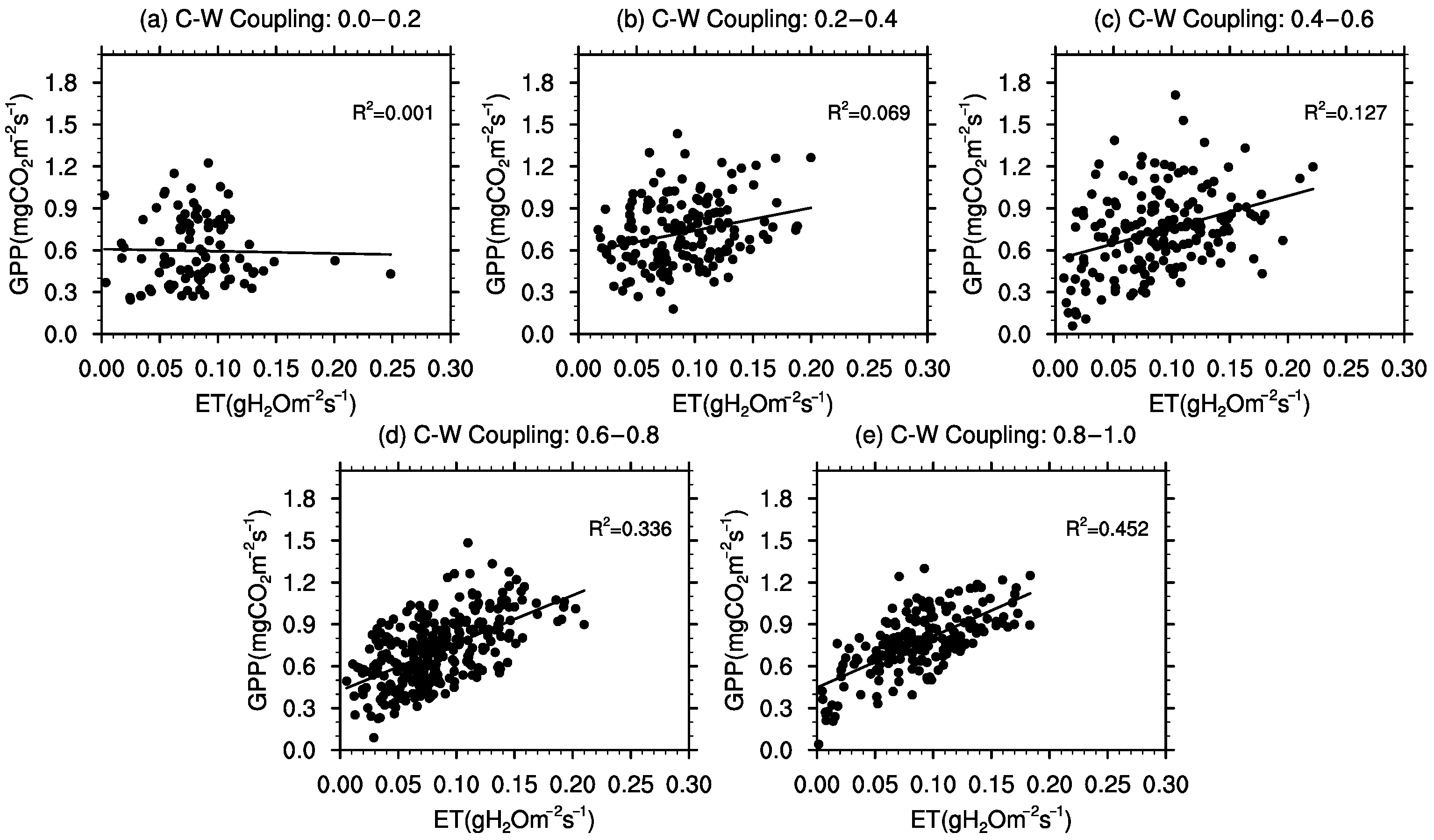
3.4. Meteorological Influences on C–W Coupling
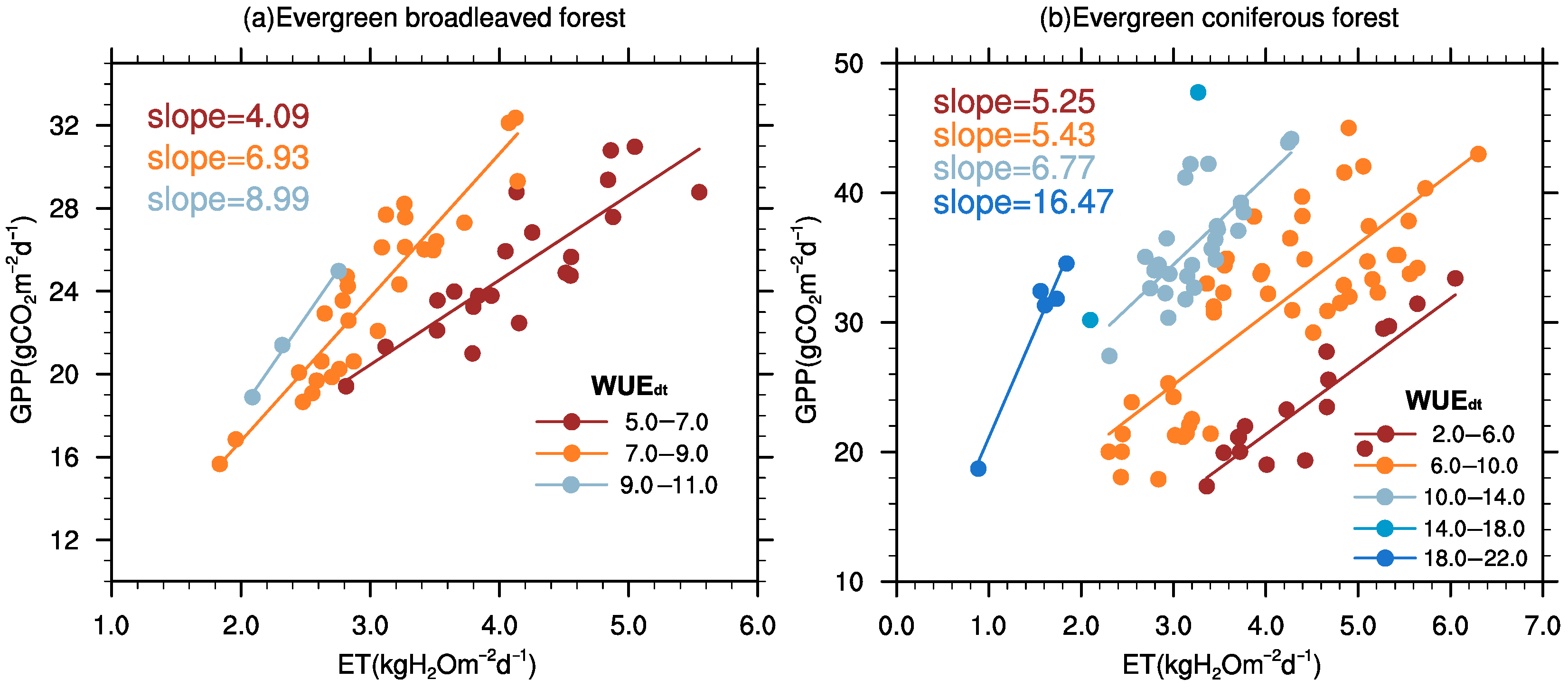
3.5. GPP–ET Relationship Based on WUEdt and Coupling Strength r
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nelson, J.A.; Carvalhais, N.; Migliavacca, M.; Reichstein, M.; Jung, M. Water-stress-induced breakdown of carbon–water relations: Indicators from diurnal FLUXNET patterns. Biogeosciences 2018, 15, 2433–2447. [Google Scholar] [CrossRef]
- Yu, G.; Song, X.; Wang, Q.; Liu, Y.; Guan, D.; Yan, J.; Sun, X.; Zhang, L.; Wen, X. Water-use efficiency of forest ecosystems in eastern China and its relations to climatic variables. New Phytol. 2008, 177, 927–937. [Google Scholar] [CrossRef]
- Gao, L.; Kang, S.; Bai, X.; Li, S.; Niu, J.; Ding, R. Water-carbon relationships and variations from the canopy to ecosystem scale in a sparse vineyard in the northwest China. J. Hydrol. 2021, 600, 126469. [Google Scholar] [CrossRef]
- Hou, Q.; Pei, T.; Yu, X.; Chen, Y.; Ji, Z.; Xie, B. The seasonal response of vegetation water use efficiency to temperature and precipitation in the Loess Plateau, China. Glob. Ecol. Conserv. 2022, 33, e01984. [Google Scholar] [CrossRef]
- Liu, D.; Mishra, A.K.; Ray, D.K. Sensitivity of global major crop yields to climate variables: A non-parametric elasticity analysis. Sci. Total Environ. 2020, 748, 141431. [Google Scholar] [CrossRef] [PubMed]
- Kirschbaum, M.U. Direct and indirect climate change effects on photosynthesis and transpiration. Plant Biol. 2004, 6, 242–253. [Google Scholar] [CrossRef]
- Kramer, R.D.; Ishii, H.R.; Carter, K.R.; Miyazaki, Y.; Cavaleri, M.A.; Araki, M.G.; Azuma, W.A.; Inoue, Y.; Hara, C. Predicting effects of climate change on productivity and persistence of forest trees. Ecol. Res. 2020, 35, 562–574. [Google Scholar] [CrossRef]
- Price, D.T.; Alfaro, R.I.; Brown, K.J.; Flannigan, M.D.; Fleming, R.A.; Hogg, E.H.; Girardin, M.P.; Lakusta, T.; Johnston, M.; McKenney, D.W. Anticipating the consequences of climate change for Canada’s boreal forest ecosystems. Environ. Rev. 2013, 21, 322–365. [Google Scholar] [CrossRef]
- Wu, J.; Serbin, S.P.; Ely, K.S.; Wolfe, B.T.; Dickman, L.T.; Grossiord, C.; Michaletz, S.T.; Collins, A.D.; Detto, M.; McDowell, N.G. The response of stomatal conductance to seasonal drought in tropical forests. Glob. Chang. Biol. 2020, 26, 823–839. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, P.G. The interpretation of the variations in leaf water potential and stomatal conductance found in canopies in the field. Philos. Trans. R. Soc. London B Biol. Sci. 1976, 273, 593–610. [Google Scholar]
- Ball, J.T.; Woodrow, I.E.; Berry, J.A. A model predicting stomatal conductance and its contribution to the control of photosynthesis under different environmental conditions. In Progress in Photosynthesis Research; Biggins, J., Ed.; Springer: Dordrecht, The Netherlands, 1987; pp. 221–224. [Google Scholar]
- Novick, K.A.; Ficklin, D.L.; Stoy, P.C.; Williams, C.A.; Bohrer, G.; Oishi, A.C.; Papuga, S.A.; Blanken, P.D.; Noormets, A.; Sulman, B.N. The increasing importance of atmospheric demand for ecosystem water and carbon fluxes. Nat. Clim. Chang. 2016, 6, 1023–1027. [Google Scholar] [CrossRef]
- Baldocchi, D.; Falge, E.; Gu, L.; Olson, R.; Hollinger, D.; Running, S.; Anthoni, P.; Bernhofer, C.; Davis, K.; Evans, R. FLUXNET: A new tool to study the temporal and spatial variability of ecosystem-scale carbon dioxide, water vapor, and energy flux densities. Bull. Am. Meteorol. Soc. 2001, 82, 2415–2434. [Google Scholar] [CrossRef]
- Beer, C.; Ciais, P.; Reichstein, M.; Baldocchi, D.; Law, B.E.; Papale, D.; Soussana, J.-F.; Ammann, C.; Buchmann, N.; Frank, D. Temporal and among-site variability of inherent water use efficiency at the ecosystem level. Glob. Biogeochem. Cycles 2009, 23, GB2018. [Google Scholar] [CrossRef]
- Boese, S.; Jung, M.; Carvalhais, N.; Reichstein, M. The importance of radiation for semiempirical water-use efficiency models. Biogeosciences 2017, 14, 3015–3026. [Google Scholar] [CrossRef]
- Law, B.E.; Falge, E.; Gu, L.V.; Baldocchi, D.D.; Bakwin, P.; Berbigier, P.; Davis, K.; Dolman, A.J.; Falk, M.; Fuentes, J.D. Environmental controls over carbon dioxide and water vapor exchange of terrestrial vegetation. Agric. For. Meteorol. 2002, 113, 97–120. [Google Scholar] [CrossRef]
- Niu, S.; Wu, M.; Han, Y.; Xia, J.; Li, L.; Wan, S. Water-mediated responses of ecosystem carbon fluxes to climatic change in a temperate steppe. New Phytol. 2008, 177, 209–219. [Google Scholar] [CrossRef]
- Li, H.; Wang, C.; Zhang, F.; He, Y.; Shi, P.; Guo, X.; Wang, J.; Zhang, L.; Li, Y.; Cao, G. Atmospheric water vapor and soil moisture jointly determine the spatiotemporal variations of CO2 fluxes and evapotranspiration across the Qinghai-Tibetan Plateau grasslands. Sci. Total Environ. 2021, 791, 148379. [Google Scholar] [CrossRef] [PubMed]
- Pang, J.; Li, H.; Yu, F.; Geng, J.; Zhang, W. Environmental controls on water use efficiency in a hilly tea plantation in southeast China. Agric. Water Manag. 2022, 269, 107678. [Google Scholar] [CrossRef]
- Zhang, M.; Huang, X.; Chuai, X.; Yang, H.; Lai, L.; Tan, J. Impact of land use type conversion on carbon storage in terrestrial ecosystems of China: A spatial-temporal perspective. Sci. Rep. 2015, 5, 10233. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Zhong, Q.; Yang, H.; Xu, C.; Hua, W.; Li, B. Patterns and driving factors of leaf C, N, and P stoichiometry in two forest types with different stand ages in a mid-subtropical zone. For. Ecosyst. 2022, 9, 100005. [Google Scholar] [CrossRef]
- Liu, X.; Trogisch, S.; He, J.-S.; Niklaus, P.A.; Bruelheide, H.; Tang, Z.; Erfmeier, A.; Scherer-Lorenzen, M.; Pietsch, K.A.; Yang, B. Tree species richness increases ecosystem carbon storage in subtropical forests. Proc. R. Soc. B 2018, 285, 20181240. [Google Scholar] [CrossRef]
- Xiang, W.; Zhou, J.; Ouyang, S.; Zhang, S.; Lei, P.; Li, J.; Deng, X.; Fang, X.; Forrester, D.I. Species-specific and general allometric equations for estimating tree biomass components of subtropical forests in southern China. Eur. J. For. Res. 2016, 135, 963–979. [Google Scholar] [CrossRef]
- Pastorello, G.; Trotta, C.; Canfora, E.; Chu, H.; Christianson, D.; Cheah, Y.-W.; Poindexter, C.; Chen, J.; Elbashandy, A.; Humphrey, M. The FLUXNET2015 dataset and the ONEFlux processing pipeline for eddy covariance data. Sci. Data 2020, 7, 225. [Google Scholar] [CrossRef]
- Lin, C.; Gentine, P.; Frankenberg, C.; Zhou, S.; Kennedy, D.; Li, X. Evaluation and mechanism exploration of the diurnal hysteresis of ecosystem fluxes. Agric. For. Meteorol. 2019, 278, 107642. [Google Scholar] [CrossRef]
- Liu, W.; Mo, X.; Liu, S.; Lin, Z.; Lv, C. Attributing the changes of grass growth, water consumed and water use efficiency over the Tibetan Plateau. J. Hydrol. 2021, 598, 126464. [Google Scholar] [CrossRef]
- Meloun, M.; Militký, J.; Hill, M.; Brereton, R.G. Crucial problems in regression modelling and their solutions. Analyst 2002, 127, 433–450. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Patel, N.R.; Bhattacharya, B.K.; Soni, P.; Parida, B.R.; Parihar, J.S. Analyzing the dynamics and inter-linkages of carbon and water fluxes in subtropical pine (Pinus roxburghii) ecosystem. Agric. For. Meteorol. 2014, 197, 206–218. [Google Scholar] [CrossRef]
- Wu, X.; Li, X.; Chen, Y.; Bai, Y.; Tong, Y.; Wang, P.; Liu, H.; Wang, M.; Shi, F.; Zhang, C. Atmospheric water demand dominates daily variations in water use efficiency in alpine meadows, northeastern Tibetan Plateau. J. Geophys. Res. Biogeosci. 2019, 124, 2174–2185. [Google Scholar] [CrossRef]
- Sulman, B.N.; Roman, D.T.; Yi, K.; Wang, L.; Phillips, R.P.; Novick, K.A. High atmospheric demand for water can limit forest carbon uptake and transpiration as severely as dry soil. Geophys. Res. Lett. 2016, 43, 9686–9695. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Sharkey, T.D. Stomatal conductance and photosynthesis. Annu. Rev. Plant Physiol. 1982, 33, 317–345. [Google Scholar] [CrossRef]
- Farquhar, G.D.; von Caemmerer, S.; Berry, J.A. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef]
- Zhang, Y.; Kang, S.; Ward, E.J.; Ding, R.; Zhang, X.; Zheng, R. Evapotranspiration components determined by sap flow and microlysimetry techniques of a vineyard in northwest China: Dynamics and influential factors. Agric. Water Manag. 2011, 98, 1207–1214. [Google Scholar] [CrossRef]
- Sakata, T.; Nakano, T.; Kachi, N. Effects of internal conductance and Rubisco on the optimum temperature for leaf photosynthesis in Fallopia japonica growing at different altitudes. Ecol. Res. 2015, 30, 163–171. [Google Scholar] [CrossRef]
- Xue, L.; Fu, F.; Chen, X.; Liu, Y.; Han, Q.; Liao, S.; Wei, Q. Analysis on water use efficiency of Populus euphratica forest ecosystem in arid area. Theor. Appl. Climatol. 2021, 145, 717–730. [Google Scholar] [CrossRef]
- Zhang, F.; Ju, W.; Shen, S.; Wang, S.; Yu, G.; Han, S. How recent climate change influences water use efficiency in East Asia. Theor. Appl. Climatol. 2014, 116, 359–370. [Google Scholar] [CrossRef]
- Zhou, S.; Duursma, R.A.; Medlyn, B.E.; Kelly, J.W.G.; Prentice, I.C. How should we model plant responses to drought? An analysis of stomatal and non-stomatal responses to water stress. Agric. For. Meteorol. 2013, 182, 204–214. [Google Scholar] [CrossRef]
- Blanken, P.D.; Rouse, W.R. Evidence of water conservation mechanisms in several subarctic wetland species. J. Appl. Ecol. 1996, 33, 842–850. [Google Scholar] [CrossRef]
- Scott, R.L.; Edwards, E.A.; Shuttleworth, W.J.; Huxman, T.E.; Watts, C.; Goodrich, D.C. Interannual and seasonal variation in fluxes of water and carbon dioxide from a riparian woodland ecosystem. Agric. For. Meteorol. 2004, 122, 65–84. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sites | Dinghushan (CN-Din) | Qianyanzhou (CN-Qia) |
|---|---|---|
| Vegetation type | Evergreen broadleaved forest | Evergreen coniferous forest |
| Latitude (°N) | 23.17 | 26.74 |
| Longitude (°W) | 112.53 | 115.06 |
| Elevation (m) | 300 | 102 |
| Mean annual temperature (°C) | 19.6 | 19.0 |
| Mean annual precipitation (mm) | 1618.1 | 1466.8 |
| Leaf area index (LAI, m2/m2) | 4.0 | 3.5 |
| Mean daytime solar radiation (W m−2) in summer | 555.6 | 592.0 |
| Mean daytime air temperature (°C) in summer | 29.1 | 31.6 |
| Mean daytime vapor pressure deficit (hPar) in summer | 14.7 | 22.4 |
| Mean daytime soil water content (%) in summer | 21.26 | 12.43 |
| Dominant species | Pinus massoniana Lam, Pinus elliottii Engelm, Cunninghamia lanceolate et al. | Schima superb, Castanea henry, Pinus massoniana Lamb et al. |
| Frequency | Half-hour | Half-hour |
| Time series | 2003–2005 | 2003–2005 |
| Broadleaved Forest | Coniferous Forest | |
|---|---|---|
| WUE (mg CO2 g−1 H2O) | 7.387 ± 1.385 | 9.014 ± 3.466 |
| C–W coupling (r) | 0.551 ± 0.236 | 0.585 ± 0.254 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, J.; Liu, J.; Yang, M.; Wang, R. Meteorological Influences on Short-Term Carbon-Water Relationships in Two Forests in Subtropical China. Atmosphere 2023, 14, 457. https://doi.org/10.3390/atmos14030457
Pan J, Liu J, Yang M, Wang R. Meteorological Influences on Short-Term Carbon-Water Relationships in Two Forests in Subtropical China. Atmosphere. 2023; 14(3):457. https://doi.org/10.3390/atmos14030457
Chicago/Turabian StylePan, Jihua, Jane Liu, Mengmiao Yang, and Rong Wang. 2023. "Meteorological Influences on Short-Term Carbon-Water Relationships in Two Forests in Subtropical China" Atmosphere 14, no. 3: 457. https://doi.org/10.3390/atmos14030457
APA StylePan, J., Liu, J., Yang, M., & Wang, R. (2023). Meteorological Influences on Short-Term Carbon-Water Relationships in Two Forests in Subtropical China. Atmosphere, 14(3), 457. https://doi.org/10.3390/atmos14030457