
Adaptability of Prunus cerasifera Ehrh. to Climate Changes in Multifunctional Landscape


 , and
, and
Abstract
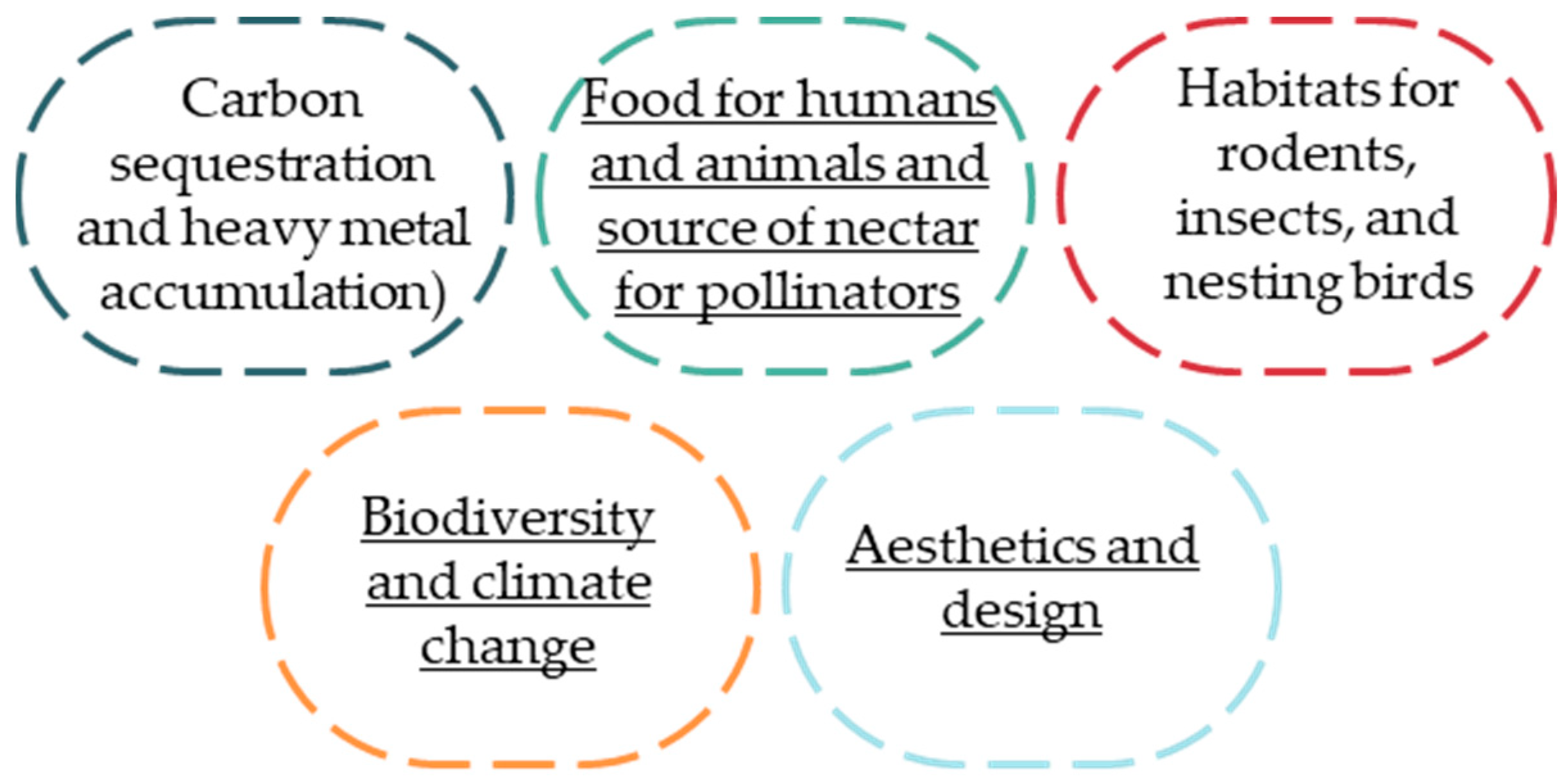
1. Introduction
2. Materials and Methods
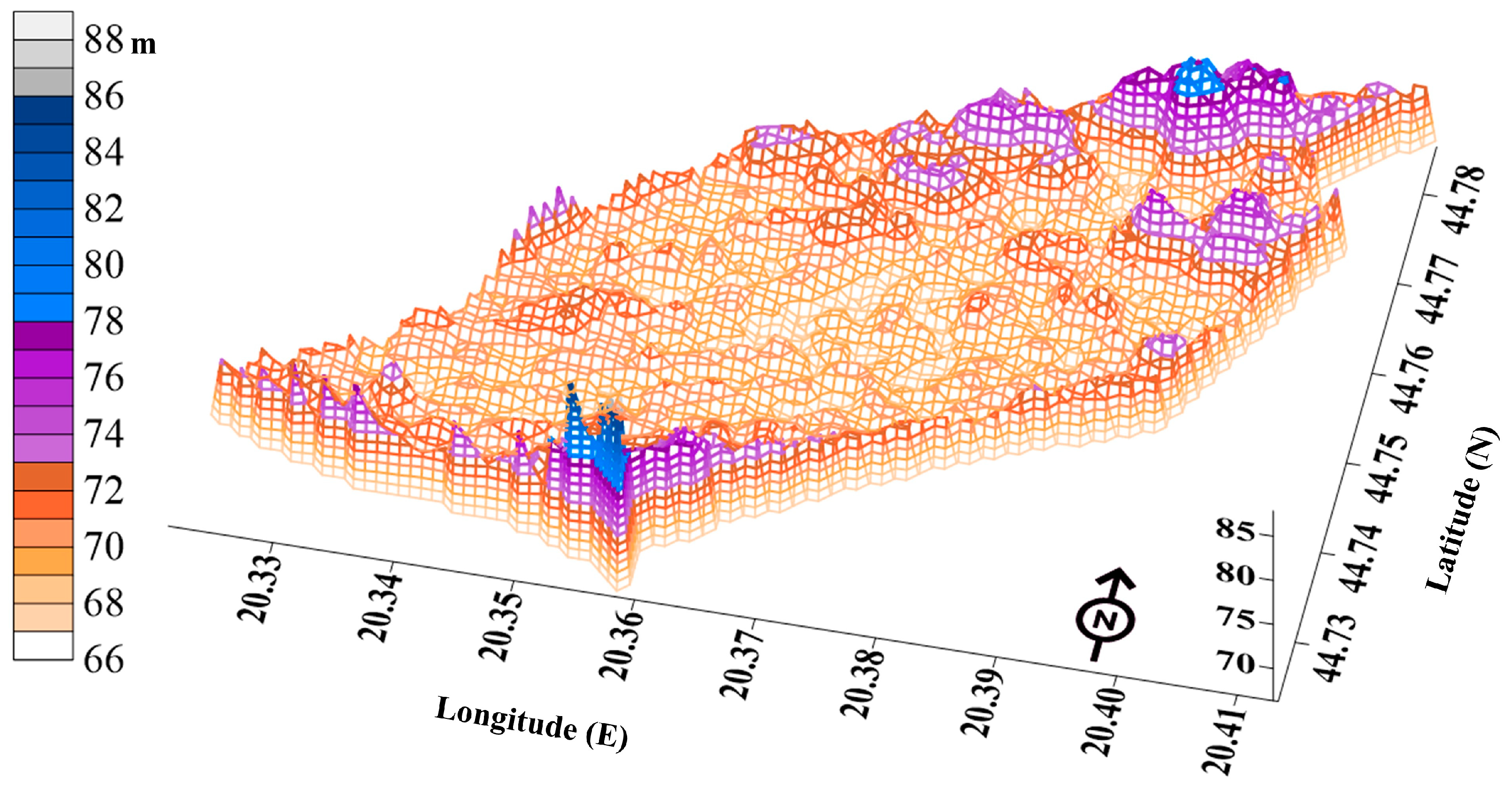
2.1. Habitat Characteristics
2.2. Phenology
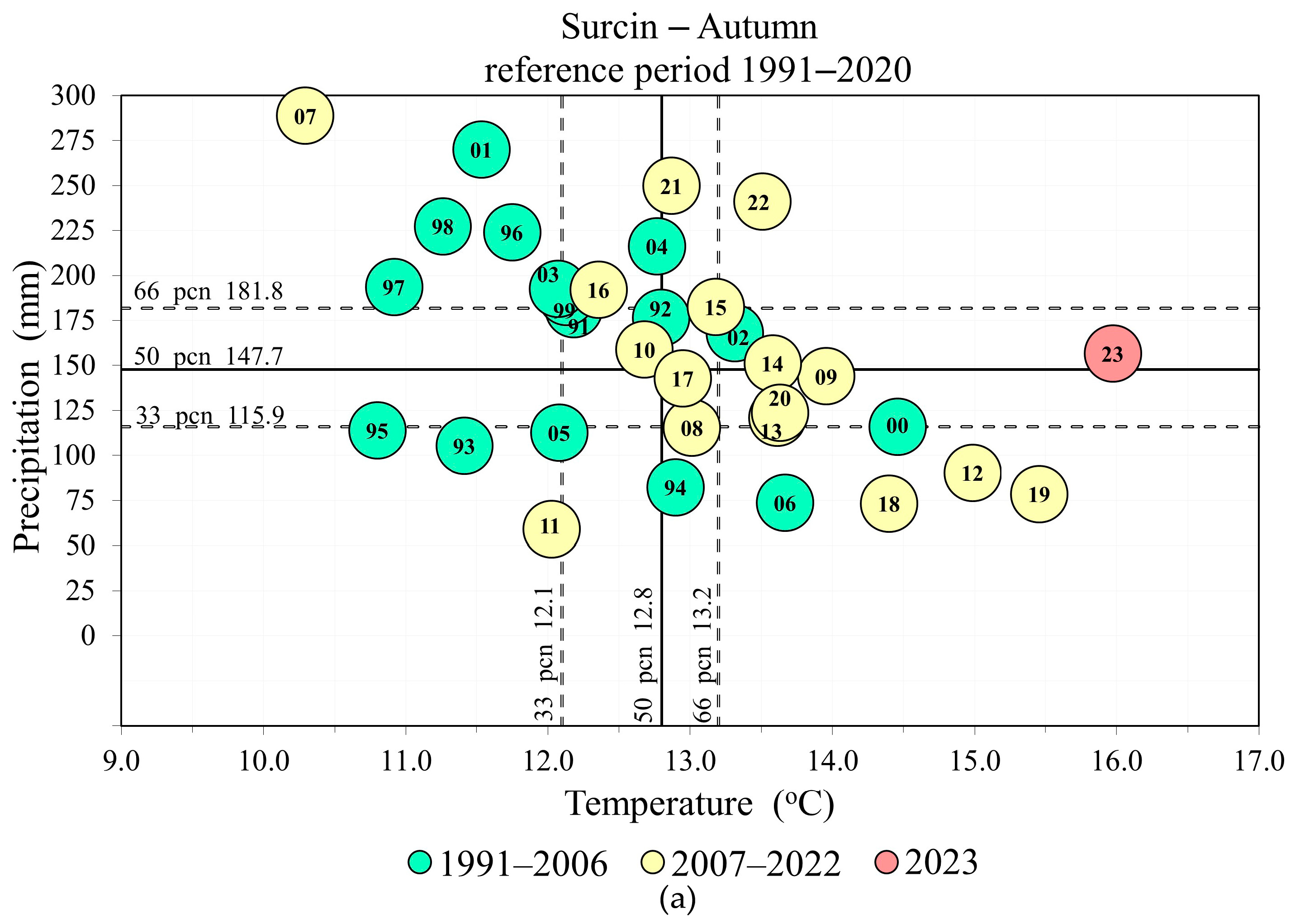
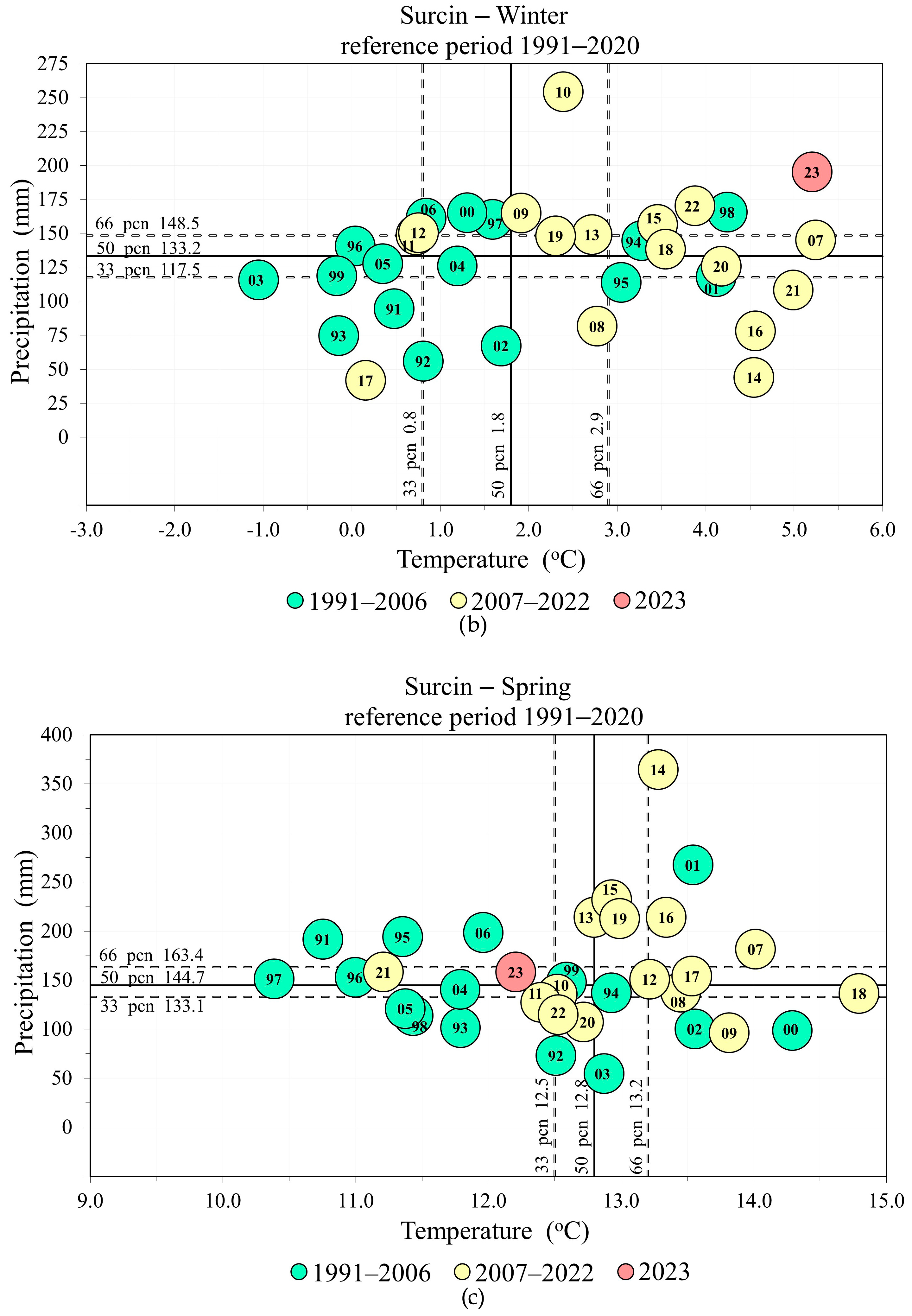
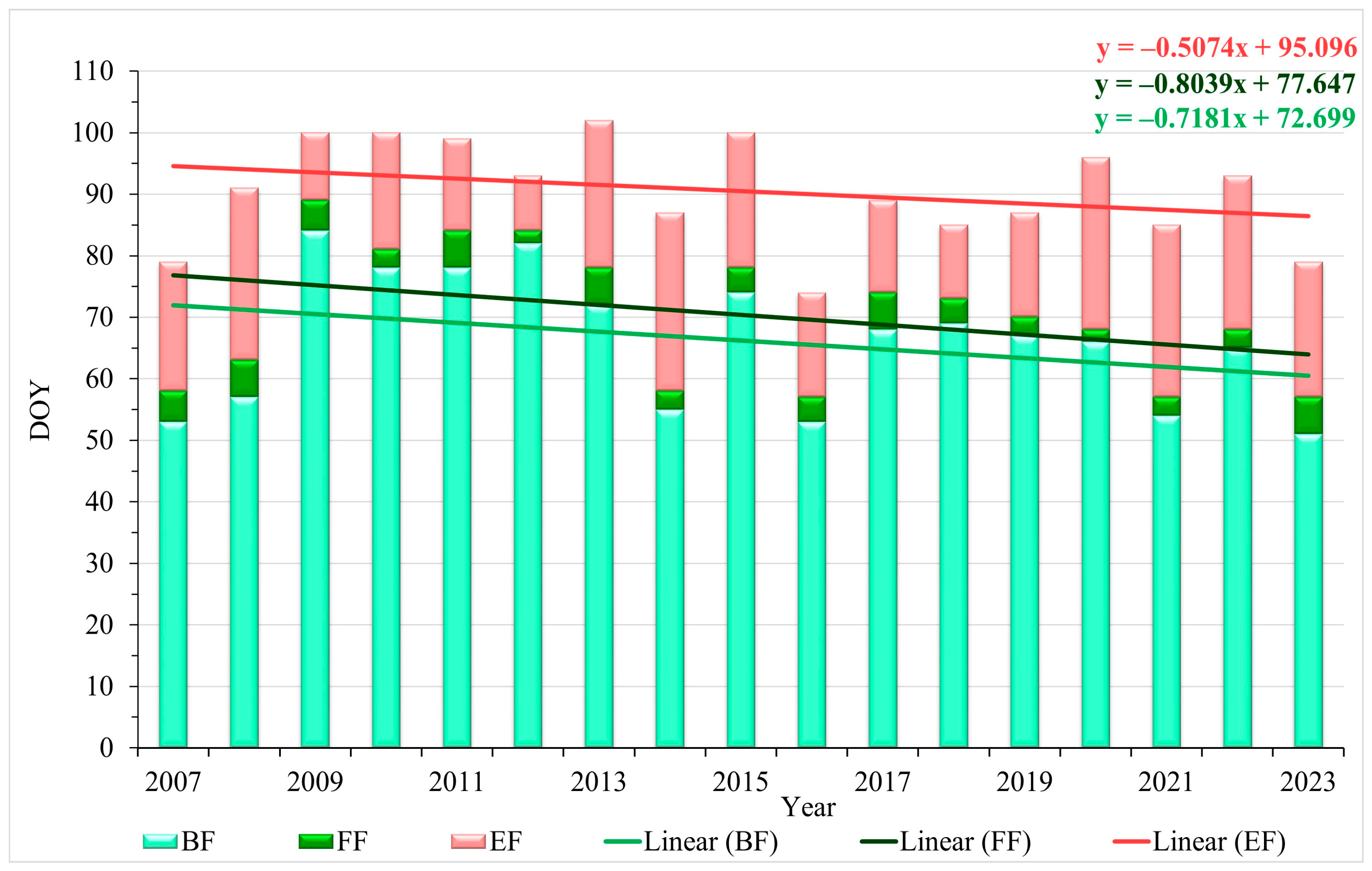
3. Results
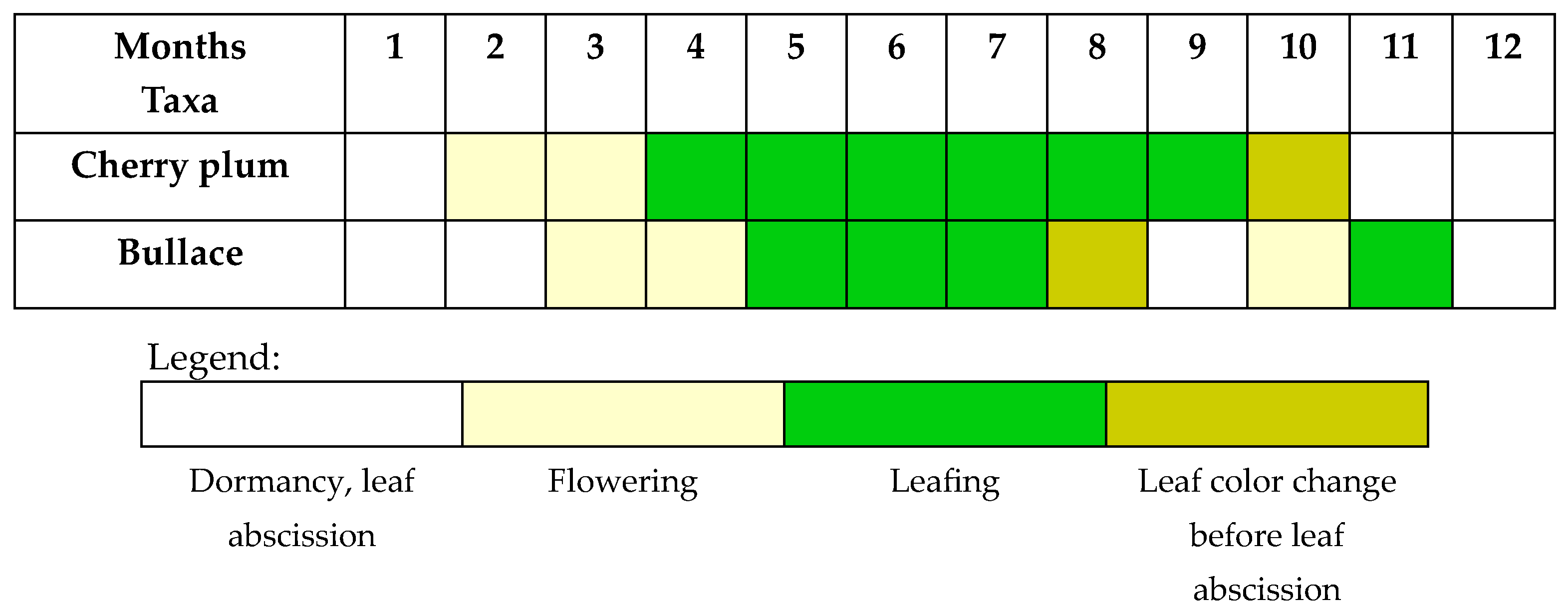
A MultiFunctional Landscape: Phenology of Cherry Plum and Bullace during 2023
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- IPCC. Summary for Policymakers: Synthesis Report—Climate Change 2023—Synthesis Report; Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2023; pp. 1–34. [Google Scholar]
- Randall, T.; Cousins, A.L.; Neilson, L.; Price, M.; Hardman, C.A.; Wilkinson, L.L. Sustainable food consumption across Western and Non-Western cultures: A scoping review considering the theory of planned behavior. Food Qual. Prefer. 2024, 114, 105086. [Google Scholar] [CrossRef]
- Shahid, M.; Dumat, C.; Khalida, S.; Schreck, E.; Xiong, T.; Nabeel, N.K. Foliar heavy metal uptake, toxicity and detoxification in plants: A comparison of foliar and root metal uptake. J. Hazard. Mater. 2017, 325, 36–58. [Google Scholar] [CrossRef]
- Cetin, M.; Onac, A.K.; Sevik, H.; Sen, B. Temporal and regional change of some air pollution parameters in Bursa. Air Qual. Atmos. Health 2019, 12, 311–316. [Google Scholar] [CrossRef]
- Qarri, F.; Lazo, P.; Stafilov, T.; Bekteshi, L.; Baceva, K.; Marka, J. Survey of atmospheric deposition of Al, Cr, Fe, Ni, V, and Zn in Albania by using moss biomonitoring and ICP-AES. Air Qual. Atmos. Health 2014, 7, 297–307. [Google Scholar] [CrossRef]
- Zhang, S.; Song, H.; Li, X.; Luo, S. Urban Parks Quality Assessment Using Multi-Dimension Indicators in Chengdu, China. Land 2024, 13, 86. [Google Scholar] [CrossRef]
- Song, P.; Gao, L. Landscape design of colleges based on the concept of ecological protection-taking Jilin Communications Polytechnic as an example. Int. J. Phytoremediat. 2024, 26, 314–323. [Google Scholar] [CrossRef]
- Smithwick, E.A.H.; Baka, J.; Bird, D.; Blaszscak-Boxe, C.; Cole, C.A.; Fuentes, J.D.; Gergel, S.E.; Glenna, L.L.; Grady, C.; Hunt, C.A.; et al. Regenerative landscape design: An integrative framework to enhance sustainability planning. Ecol. Soc. 2023, 28, 5. [Google Scholar] [CrossRef]
- Wang, H.; Tassinary, L.G.; Newman, G.D. Developing the health effect assessment of landscape (HEAL) Tool: Assessing the health effects of community greenspace morphology design on non-communicable diseases. Landsc. Urban Plann. 2024, 244, 104990. [Google Scholar] [CrossRef]
- Roman, L.A.; McPherson, E.G.; Scharenbroch, B.C.; Bartens, J. Identifying common practices and challenges for local urban tree monitoring programs across the United States. Arboric. Urban For. 2013, 39, 292–299. [Google Scholar] [CrossRef]
- Martin, M.P.; Simmons, C.; Ashton, M.S. Survival is not enough: The effects of microclimate on the growth and health of three common urban tree species in San Francisco, California. Urban For. Urban Green. 2016, 19, 1–6. [Google Scholar] [CrossRef]
- Ognjanov, V.; Cerović, S. Selection and utilization of minor fruit tree species. Acta Hortic. 2004, 663, 569–574. [Google Scholar] [CrossRef]
- Cosmulescu, S.N.; Ionica, M.E.; Mutu, N. Evaluation on the genetic diversity of phenotypic traits in myrobalan plum (Prunus cerasifera Ehrh.). South-West J. Hortic. Biol. Environ. 2018, 9, 25–34. [Google Scholar]
- Nikolić, D.; Rakonjac, V. Divergence of myrobalan (Prunus cerasifera Ehrh.) types on the territory of Serbia. Genetika 2007, 39, 333–342. [Google Scholar] [CrossRef]
- Mišić, P.D. Gene banks and preselection in fruit plants. Genetika 1983, 15, 361–368. [Google Scholar]
- Baraldi, R.; Chieco, C.; Neri, L.; Facini, O.; Rapparini, F.; Morrone, L.; Rotondi, A.; Carriero, G. An integrated study on air mitigation potential of urban vegetation: From a multi-trait approach to modeling. Urban For. Urban Green. 2019, 41, 127–138. [Google Scholar] [CrossRef]
- Šebek, G. The phenological and pomological traits of autochthonous plum cultivars in the area of North Montenegro. Acta Sci. Pol. Hortorum Cultus 2016, 15, 45–63. [Google Scholar]
- Milutinovic, M.; Nikolic, D. Genetical variability of myrobalan (Prunus cerasifera) seedlings. Acta Hortic. 1994, 359, 217–224. [Google Scholar] [CrossRef]
- Ogašanović, D.; Korać, M.; Miletić, R.; Milutinović, M.; Mišić, P.D.; Miranović, K.; Nikolić, M.; Ognjanov, V.; Paunović, S.A.; Tešović, Ž.; et al. Genetic resources of the most important fruit tree species of FR Yugoslavia. [Genetički resursi važnijih vrsta voćaka SR Jugoslavije]. Savremena Poljoprivreda 1997, 46, 161–173. (In Serbian) [Google Scholar]
- Božović, Đ.; Jaćimović, V. Resistance to low temperatures, causing disease and pests of isolated genotypes of sedge (Prunus cerasifera Ehrh.) in the area of upper Polimlje. [Otpornost na niske temperature, prouzrokovače bolesti ištetotičine izdvojenih genotipova džanarike (Prunus cerasifera Ehrh.) u području gornjeg Polimlja]. Zbornik Naučnih Radova sa XVII Savetovanja Agronoma, Veterinara i Tehnologa 2003, 9, 185–191. (In Serbian) [Google Scholar]
- Li, Q.; Zhu, Y.; Zhu, Z. Calculation and Optimization of the Carbon Sink Benefits of Green Space Plants in Residential Areas: A Case Study of Suojin Village in Nanjing. Sustainability 2023, 15, 607. [Google Scholar] [CrossRef]
- Zhang, B.; Feng, Q.; Xue, Y.; Liu, D.; Zhao, C.; Yan, S.; Li, J. Accumulation characteristics, seasonal changes, and source analysis of heavy metals in major greening tree species on common highways in Fuzhou, China. Chin. J. Appl. Environ. Biol. 2022, 28, 1199–1208. [Google Scholar] [CrossRef]
- Turkyilmaz, A.; Sevik, H.; Cetin, M.; Ahmaida Saleh, E. Changes in Heavy Metal Accumulation Depending on Traffic Density in Some Landscape Plants. Pol. J. Environ. Stud. 2018, 27, 2277–2284. [Google Scholar] [CrossRef]
- Ganji Moghaddam, E.; Hossein Ava, S.; Akhavan, S.; Hosseini, S. Phenological and pomological characteristics of some plum (Prunus spp.) cultivars grown in Mashhad, Iran. Crop Breed. J. 2011, 1, 105–108. [Google Scholar] [CrossRef]
- Milošević, T.; Milošević, N. Plum (Prunus spp.) Breeding. In Advances in Plant Breeding Strategies: Fruits; Al-Khayri, J., Jain, S.M., Johnson, D.V., Eds.; Springer: Cham, Switzerland, 2018; Volume 3, pp. 165–215. ISBN 978-3-319-91943-0. [Google Scholar]
- Butac, M. Plum Breeding. In Prunus; Küden, A., Küden, A., Eds.; IntechOpen: London, UK, 2020; ISBN 978-1-83962-608-1. [Google Scholar] [CrossRef]
- Halpern, C.B.; Spies, T.A. Plant Species Diversity in Natural and Managed Forests of the Pacific Northwest. Ecol. Appl. 1995, 5, 913–934. [Google Scholar] [CrossRef]
- Carey, A.B. Biocomplexity and restoration of biodiversity in temperate coniferous forest: Inducing spatial heterogeneity with variable-density thinning. Forestry 2003, 76, 127–136. [Google Scholar] [CrossRef]
- Czortek, P.; Adamowski, W.; Kamionka-Kanclerska, K.; Karpińska, O.; Zalewski, A.; Dyderski, M. Patterns of Prunus cerasifera early invasion stages into a temperate primeval forest. Biol. Invasions 2023, 26, 633–647. [Google Scholar] [CrossRef]
- Zuo, J.; Zhao, Z.Y. Green building research–current status and future agenda: A review. Renew. Sustain. Energy Rev. 2014, 30, 271–281. [Google Scholar] [CrossRef]
- Radutoiu, D.; Stefanescu, D.M. Aesthetics of ruderal vegetation in the urban and peri-urban areas of Oltenia (Romania). Ann. Univ. Craiova Agric. Mont. Cadastre Ser. 2017, XLVII, 223–228. [Google Scholar]
- Lo Piccolo, E.; Landi, M. Red-leafed species for urban “greening” in the age of global climate change. J. For. Res. 2021, 32, 151–159. [Google Scholar] [CrossRef]
- Cosmulescu, S.; Buican Stanciu, A.; Ionescu, M. The influence of temperature on phenology of ornamental woody species in urban environment. scientific papers. Sci. Pap. Ser. B Hortic. 2020, 64, 61–67. [Google Scholar]
- Richardson, A.D.; Anderson, R.S.; Arain, M.A.; Barr, A.G.; Bohrer, G.; Chen, G.; Chen, J.M.; Ciais, P.; Davis, K.J.; Desai, A.R.; et al. Terrestrial biosphere models need better representation of vegetation phenology: Results from the North American Carbon Program Site Synthesis. Glob. Chang. Biol. 2012, 18, 566–584. [Google Scholar] [CrossRef]
- Burkle, L.A.; Marlin, J.C.; Knight, T.M. Plant-pollinator interactions over 120 years: Loss of species, co-occurrence, and function. Science 2013, 339, 1611–1615. [Google Scholar] [CrossRef]
- Xia, J.; Niu, S.; Ciais, P.; Janssens, I.A.; Chen, J.; Ammann, C.; Arain, A.; Blanken, P.D.; Cescatti, A.; Bonal, D.; et al. Joint control of terrestrial gross primary productivity by plant phenology and physiology. Proc. Natl. Acad. Sci. USA 2015, 112, 2788–2793. [Google Scholar] [CrossRef]
- Fitter, A.; Fitter, R.; Harris, I.; Williamson, M. Relationships between first flowering date and temperature in the flora of a locality in central England. Funct. Ecol. 1995, 9, 55–60. [Google Scholar] [CrossRef]
- Menzel, A. Trends in phenological phases in Europe between 1951 and 1996. Int. J. Biometeorol. 2000, 44, 76–81. [Google Scholar] [CrossRef]
- Rötzer, T.; Chmielewski, F.M. Phenological maps of Europe. Clim. Res. 2001, 18, 249–257. [Google Scholar] [CrossRef]
- Sparks, T.H.; Menzel, A. Observed changes in seasons: An overview. Int. J. Climatol. 2002, 22, 1715–1726. [Google Scholar] [CrossRef]
- Vitasse, Y.; Porté, A.J.; Kremer, A.; Michalet, R.; Delzon, S. Responses of canopy duration to temperature changes in four temperate tree species: Relative contributions of spring and autumn leaf phenology. Oecologia 2009, 161, 187–198. [Google Scholar] [CrossRef]
- Beaubien, E.; Hamann, A. Spring Flowering Response to Climate Change between 1936 and 2006 in Alberta, Canada. BioScience 2011, 61, 514–524. [Google Scholar] [CrossRef]
- Stojicic, Đ. The Impact of Environmental Factors on Flowering of Woody Species from Magnoliophyta group in Belgrade. Ph.D. Thesis, Univerzitet u Beogradu—Šumarski Fakultet, Belgrade, Serbia, 2014; p. 243. (In Serbian). [Google Scholar]
- Wang, H.; Zhong, S.; Tao, Z.; Dai, J.; Ge, Q. Changes in flowering phenology of woody plants frm 1963 to 2014 in North China. Int. J. Biometeorol. 2019, 63, 579–590. [Google Scholar] [CrossRef]
- Ocokoljić, M.; Petrov, Đ.; Galečcić, N.; Skočajić, D.; Košanin, O.; Simović, I. Phenological Flowering Patterns of Woody Plants in the Function of Landscape Design: Case Study Belgrade. Land 2023, 12, 706. [Google Scholar] [CrossRef]
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef]
- Hamann, A.; Wang, T. Potential effects of climate change on ecosystem and tree species distribution in British Columbia. Ecology 2006, 87, 2773–2786. [Google Scholar] [CrossRef]
- Qiu, J.; Carpenter, S.R.; Booth, E.G.; Motew, M.; Zipper, S.C.; Kucharik, C.J.; Loheide, S.P., II; Turner, M.G. Understanding relationships among ecosystem services across spatial scales and over time. Environ. Res. Lett. 2018, 13, 054020. [Google Scholar] [CrossRef]
- Preve´y, J.S.; Parker, L.E.; Harrington, C.A. Projected impacts of climate change on the range and phenology of three culturally-important shrub species. PLoS ONE 2020, 15, e0232537. [Google Scholar] [CrossRef]
- Richardson, A.D. Climate change, phenology, and phenological control of vegetation feedbacks to the climate system. Agric. For. Meteorol. 2013, 169, 156173. [Google Scholar] [CrossRef]
- Bagne, K.; Friggens, M.; Finch, D. A System for Assessing Vulnerability of Species (SAVS) to Climate Change; General Technical Report RMRS-GTR-257; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2011; p. 28.
- Morellato, L.P.C.; Alberton, B.; Alvarado, S.T.; Borges, B.; Buisson, E.; Camargo, M.G.G.; Cancian, L.F.; Carstensen, D.W.; Escobar, D.F.E.; Leite, P.T.P.; et al. Linking plant phenology to conservation biology. Biol. Conserv. 2016, 195, 60–72. [Google Scholar] [CrossRef]
- Dalhaus, T.; Musshoff, O.; Finger, R. Phenology Information Contributes to Reduce Temporal Basis Risk in Agricultural Weather Index Insurance. Sci. Rep. 2018, 8, 46. [Google Scholar] [CrossRef]
- Grigoriev, A.; Lukicheva, L.; Korzin, V.; Gorina, V. Biotic and abiotic factors limiting the productivity of Prunus cerasifera subsp. macrocarpa (Ehrh.) Erem. et Garcov. Acta Hortic. 2021, 1322, 193–200. [Google Scholar] [CrossRef]
- Duchovskis, P.; Stanys, V.; Sasnauskas, A.; Bobinas, C. Cold resistance of Prunus domestica L. and Prunus cerasifera Ehrh. in Lithuania. Acta Hortic. 2007, 734, 299–303. [Google Scholar] [CrossRef]
- Augspurger, C.K. Reconstructing patterns of temperature, phenology, and frost damage over 124 years: Spring damage risk is increasing. Ecology 2013, 94, 41–50. [Google Scholar] [CrossRef]
- Ru, X.; Jiang, Y.; Luo, Q.; Wang, R.; Feng, X.; Wang, J.; Wang, Z.; Li, M.; Qu, Z.; Su, B.; et al. Evaluating late spring frost risks of apple in the Loess Plateau of China under future climate change with henological modeling approach. Sci. Hortic. 2023, 308, 111604. [Google Scholar] [CrossRef]
- Landi, M.; Lo Piccolo, E.; Pellegrini, E.; Agati, G.; Giordano, C.; Giordani, T.; Lorenzini, G.; Malorgio, F.; Massai, R.; Nali, C.; et al. The use of red species for urban “greening” in the age of climate change. Agrochimica 2019, 157–161. [Google Scholar]
- Simović, I.; Šikoparija, B.; Panić, M.; Radulović, M.; Lugonja, P. Remote Sensing of Poplar Phenophase and Leaf Miner Attack in Urban Forests. Remote Sens. 2022, 14, 6331. [Google Scholar] [CrossRef]
- Available online: http://www.sllistbeograd.rs/pdf/2020/153-2020.pdf (accessed on 18 September 2023).
- Available online: https://www.srbijavode.rs/images/dokumenti/vodoprivredna_osnova_republike_srbije.pdf (accessed on 18 September 2023).
- Anđelković, M. Geology of the Wider Area of Belgrade 4—Paleogeography [Geologija Šire Okoline Beograda 4—Paleogeografija]; Zavod za Regionalnu Geologiju i Paleontologiju: Belgrade, Serbia, 1989. (In Serbian) [Google Scholar]
- Direkcija za Građevinsko Zamljište i Izgradnju Beograda. General Urban Plan of Belgrade, “Official Gazette of the City of Belgrade”, Number 11/16; Direkcija za Građevinsko Zamljište i Izgradnju Beograda: Belgrade, Serbia, 2016. [Google Scholar]
- Teofilović, A.; Cvejić, J.; Jovanović, S.; Lakušić, D.; Tutundžić, A.; Mitrović, M.; Čavić, K.; Jeftić, N.; Glamočić, B.; Vasiljević, N.; et al. Mapping and Evaluation of Belgrade Habitats. Results of the Project “Belgrade Green Regulations—Phase III”; Electronic Edition; Secretariat for Environmental Protection of Belgrade: Belgrade, Serbia, 2008; Volume 1, pp. 1–112. [Google Scholar]
- Meier, U. BBCH-Monograph. Growth Stages of Plants [Entwicklungsstadien von Pflanzen. Estadios de las Plantas. Stades De Développement des Plantes]; Blackwell Wissenschafts-Verla: Berlin, Germany; Vienna, Austria, 1997; p. 622. [Google Scholar]
- Stilinović, S. Semenarstvo Šumskog i Ukrasnog Drveća i Žbunja; OOUR Institut za Šumarstvo Šumarskog Fakulteta u Beogradu: Belgrade, Serbia, 1985; p. 399. [Google Scholar]
- Vučetić, V. Secular trend analysis of growing degree-days in Croatia. Idojaras 2009, 113, 39–46. [Google Scholar]
- WMO. State of the Global Climate 2021: WMO Provisional Report; WMO No. 1290; World Meteorological Organization (WMO), World Climate Data and Monitoring Programme: Geneva, Switzerland, 2021; p. 57. Available online: https://library.wmo.int/doc_num.php?explnum_id=10859 (accessed on 18 September 2023).
- Lalić, B.; Ejcinger, J.; Dalamarta, A.; Orlandini, S.; Firanj Sremac, A.; Paher, B. Meteorology and Climatology for Agronomists. [Meteorologija i Klimatologija za Agronome]; Univerzitet u Novom Sadu—Poljoprivredni Fakultet: Novi Sad, Serbia, 2021; p. 219. (In Serbian) [Google Scholar]
- Available online: https://www.hidmet.gov.rs/ciril/meteorologija/klimatologija_godisnjaci.php (accessed on 1 November 2023).
- Glowacka, A.; Sitarek, M.; Rozparaa, E.; Podwyszynska, M. Pomological characteristics and ploidy levels of Japanese plum (Prunus salicina Lindl.) cultivars preserved in Poland. Plants 2021, 10, 884. [Google Scholar] [CrossRef]
- Silva, I.A.; Da Silva, D.M.; De Carvalho, G.H.; Batalha, M.A. Reproductive phenology of Brazilian savannas and riparian forests: Environmental and phylogenetic issues. Ann. For. Sci. 2011, 68, 1207–1215. [Google Scholar] [CrossRef]
- CaraDonna, P.J.; Inouye, D.W. Phenological responses to climate change do not exhibit phylogenetic signal in a subalpine plant community. Ecology 2015, 96, 355–361. [Google Scholar] [CrossRef]
- Sevik, H.; Cetin, M.; Ozel, H.B.; Pinar, B. Determining toxic metal concentration changes in landscaping plants based on some factors. Air Qual. Atmos. Health 2019, 12, 983–991. [Google Scholar] [CrossRef]
- Ding, H.; Xu, L.; Elmore, A.J.; Shi, Y. Vegetation Phenology Influenced by Rapid Urbanization of The Yangtze Delta Region. Remote Sens. 2020, 12, 1783. [Google Scholar] [CrossRef]
- Ocokoljić, M.; Petrov, Đ. Decorative Dendrology. [Dekorativna Dendrologija]; Univerzitet u Beogradu—Šumarski Fakultet: Belgrade, Serbia, 2022; p. 409. (In Serbian) [Google Scholar]
- POWO Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. Published on the Internet. 2023. Available online: https://www.plantsoftheworldonline.org (accessed on 28 November 2023).
- Hartmann, W.; Neumüller, M. Plum Breeding. In Breeding Plantation Tree Crops: Temperate Species; Jain, S.M., Priyadarshan, P.M., Eds.; Springer: New York, NY, USA, 2009; Volume 6, pp. 161–231. [Google Scholar]
- Wang, Y.X.; Chen, X.; Zhang, Y.; Chen, X. Antioxidant activities and major anthocyanins of myrobalan plum (Prunus cerasifera Ehrh.). Food Sci. 2012, 77, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Fanning, K.J.; Topp, B.; Russell, D.; Stanleyd, R.; Netzel, M. Japanese plums (Prunus salicina Lindl.) and phytochemicals—Breeding, horticultural practice, postharvest storage, processing and bioactivity. J. Sci. Food Agric. 2014, 94, 2137–2147. [Google Scholar] [CrossRef]
- Slepneva, T.; Makarenko, S. Economic and biological evaluation hybrids of Prunus cerasifera Ehrem. var. pissardii Bail. in conditions of the Southern Urals. BIO Web Conf. 2020, 24, 00082. [Google Scholar] [CrossRef]
- Dunaevskaya, E.V.; Gorina, V.M.; Melkozerova, E.A.; Khetagurov, K.M.; Kozyrev, A.K. Prunus cerasifera Ehrh. fruits are a source of biologically active substances and high-quality feedstock for Food for Specific Health Use. E3S Web Conf. 2021, 282, 04008. [Google Scholar] [CrossRef]
- Shamsolshoara, Y.; Seied, M.; Gharesheikhbayat, R.; Pirkhezri, M.; Davoodi, D. Phenological, morphological, and pomological characterizations of three promising plum and apricot natural hybrids. Taiwania 2021, 66, 466–478. [Google Scholar] [CrossRef]
- Lhotka, O.; Brönnimann, S. Possible Increase of Vegetation Exposure to Spring Frost under Climate Change in Switzerland. Atmosphere 2020, 11, 391. [Google Scholar] [CrossRef]
- Chitu, E.; Sumedrea, D.; Paltineanu, C. Phenolocial and climatic simulation of the late frost damage in plum orchard under the climate conditions of Romania. Acta Hortic. 2008, 803, 139–146. [Google Scholar] [CrossRef]
- Yaacoubi, A.E.; Oukabli, A.; Hafidi, M.; Farrera, I.; Ainane, T.; Cherkaoui, S.I.; Legave, J.M. Validated model for apple flowering prediction in the Mediterranean area in response to temperature variation. Sci. Hortic. 2019, 249, 59–64. [Google Scholar] [CrossRef]
- Vujadinović Mandić, M.; Vuković Vimić, A.; Ranković-Vasić, Z.; Đurović, D.; Ćosić, M.; Sotonica, D.; Nikolić, D.; Đurđević, V. Observed Changes in Climate Conditions and Weather-Related Risks in Fruit and Grape Production in Serbia. Atmosphere 2022, 13, 948. [Google Scholar] [CrossRef]
- Jochner, S.; Menzel, A. Urban phenological studies—Past, present, future, Environmental Pollution. Environ. Pollut. 2015, 203, 250–261. [Google Scholar] [CrossRef]
- Deng, L.; Shen, W.; Lin, Y.; Gao, W.; Lin, J. Surveillance Camera-Based Monitoring of Plant Flowering Phenology. In Geo-Spatial Knowledge and Intelligence, Proceedings of the 4th International Nonference on Feo-Informatics in Resource Management and Sustainable Ecosystem, GRMSE 2016, Hong Kong, China, 18–20 November 2016; Yuan, H., Geng, J., Bian, F., Eds.; Part of the Communications in Computer and Information Science; Springer: Singapore, 2017; 698p, p. 698. [Google Scholar] [CrossRef]
- Deng, L.; Lin, Y.; Yan, L.; Tesfamichael, S.; Billen, R.; Yao, Y.; Yao, W.; Chen, X.; Fang, X.; Wang, C.; et al. Urban plant phenology monitoring: Expanding the functions of widespread surveillance cameras to nature rhythm understanding. RSASE Remote Sens. Appl. Soc. Environ. 2019, 15, 100232. [Google Scholar] [CrossRef]
- Didukh, Y.P.; Kucher, O.O.; Zavialova, L.V. Phenological reaction of vascular plants to the extreme thermal regime in autumn–winter of 2019 in Ukraine. Ukr. Bot. J. 2020, 77, 143–155. [Google Scholar] [CrossRef]
- Surányi, D. Phenological and flower morphological studies on different plum and prune cultivars. Int. J. Hortic. Sci. 2013, 19, 43–52. [Google Scholar] [CrossRef]
- Zverko, J.; Vido, J.; Škvareninová, J.; Škvarenina, J. Early Onset of Spring Phenological Phases in the Period 2007–2012 Compared to the Period 1931–1960 as a Potential Bioindicator of Nvironmental Changes in the National Nature Reserve Boky (Slovakia); Rožnovský, J., Litschmann, T., Eds.; Mendel a Bioklimatologie: Brno, Czech Republic, 2014; Volume 9, pp. 3–5. ISBN 978-80-210-6983-1. [Google Scholar]
- Buican Stanciu, A.; Ionescu, M.; Cosmulescu, S.N. The influence of urban conditions on the phenology of some ornamental species. Biharean Biol. 2021, 15, 75–79. [Google Scholar]
- Onal, K. Collecting and evaluation of myrobalan plum (Prunus cerasifera Ehrh.) from southeast Anatolia. Plant Genet. Resour. Newsl. 1998, 115, 49–50. [Google Scholar]
- Buican Stanciu, A.; Ionescu, M.; Cosmulescu, S.N. Spring phenology of some ornamental species, as an indicator of temperature increase in the urban climate area. Not. Sci. Biol. 2021, 13, 11007. [Google Scholar] [CrossRef]
- Robeson, S.M. Increasing growing-season length in Illinois during the 20th century. Clim. Chang. 2002, 52, 219–238. [Google Scholar] [CrossRef]
- Wang, H.; Zexing, T.; Wang, H.; Ge, Q. Varying temperature sensitivity of bud-burst date at different temperature conditions. Int. J. Biometeorol. 2021, 65, 357–367. [Google Scholar] [CrossRef]
- Egea, J.; Ortega, E.; Martínez-Gómez, P.; Dicenta, F. Chilling and heat requirements of almond cultivars for flowering. Environ. Exp. Bot. 2003, 50, 79–85. [Google Scholar] [CrossRef]
- Ruiz, D.; Campoy, J.A.; Egea, J. Chilling and heat requirements of apricot cultivars for flowering. Environ. Exp. Bot. 2007, 61, 254–263. [Google Scholar] [CrossRef]
- Nečas, T.; Rampáčková, E.; Göttingerová, M.; Kiss, T.; Ondrášek, I. Evaluation of non-traditional plum cultivars for growing in the Czech conditions. Acta Hortic. 2021, 1322, 113–124. [Google Scholar] [CrossRef]
- Nguyen, A. A Benefit–Cost Analysis of Ten Urban Landscaping Trees in Berkley; CA Drubin/Barnes Lab: Berkeley, CA, USA, 2005; pp. 1–26. [Google Scholar]
- Karlović, K.; Jeran, N.; Dujmović Purgar, D.; Židovec, V.; Bolarić, S.; Vokurka, A. Sustainable landscaping with Prunus. Acta Hortic. 2020, 1288, 75–84. [Google Scholar] [CrossRef]
- Keenan, T.; Gray, J.; Friedl, M.; Toomey, M.; Bohrer, G.; Hollinger, D.; Munger, J.W.; O’Keefe, J.; Schmid, H.P.; Wing, I.S.; et al. Net carbon uptake has increased through warming-induced changes in temperate forest phenology. Nat. Clim. Chang. 2014, 4, 598–604. [Google Scholar] [CrossRef]
- Churkina, G.; Schimel, D.; Braswell, B.H.; Xiao, X. Spatial analysis of growing season length control over net ecosystem exchange. Glob. Chang. Biol. 2005, 11, 1777–1787. [Google Scholar] [CrossRef]
- Dragoni, D.; Schmid, H.P.; Wayson, C.A.; Potter, H.; Grimmond, C.S.B.; Randolph, J.C. Evidence of increased net ecosystem productivity associated with a longer vegetated season in a deciduous forest in south-central Indiana, USA. Glob. Chang. Biol. 2011, 17, 886–897. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | BF DOY | FF DOY | EF DOY | BF GDD | FF GDD | EF GDD |
|---|---|---|---|---|---|---|
| Minimum | 51.000 | 57.000 | 74.000 | 85.529 | 103.173 | 152.667 |
| Maximum | 84.000 | 89.000 | 102.000 | 153.082 | 162.887 | 309.247 |
| Range | 33.000 | 32.000 | 28.000 | 67.553 | 59.715 | 156.581 |
| 1st Quartile | 55.000 | 58.000 | 85.000 | 103.115 | 125.548 | 210.594 |
| Median | 67.000 | 70.000 | 91.000 | 114.788 | 134.595 | 221.087 |
| 3rd Quartile | 74.000 | 78.000 | 99.000 | 131.219 | 141.786 | 256.229 |
| Mean | 66.235 | 70.412 | 90.529 | 116.062 | 134.837 | 230.662 |
| Standard deviation (n) | 10.547 | 10.505 | 8.197 | 19.367 | 15.704 | 35.722 |
| Variation coefficient (n) | 0.159 | 0.149 | 0.091 | 0.167 | 0.116 | 0.155 |
| Kurtosis (Pearson) | −1.263 | −1.287 | −0.919 | −0.996 | −0.508 | 0.110 |
| Standard error (Kurtosis (Fisher) | 1.063 | 1.063 | 1.063 | 1.063 | 1.063 | 1.063 |
| Mean Air Temperature (°C) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Months Period | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | |
| Tmean 1991/2020 | 1.0 | 3.0 | 7.5 | 12.9 | 17.6 | 21.4 | 23.3 | 23.2 | 18.0 | 12.8 | 7.4 | 2.2 | 12.5 |
| Tmean 2007/2022 | 1.8 | 4.2 | 8.0 | 13.5 | 17.8 | 22.2 | 24.1 | 24.0 | 18.6 | 13.0 | 8.2 | 3.3 | 13.2 |
| Tmean 2023 | 5.1 | 4.1 | 9.3 | 10.6 | 16.8 | 21.2 | 25.3 | 23.8 | 21.9 | 17.02 | 9.0 | 5.9 | 14.2 |
| Deviation Tmean 2023 from the reference period | 4.2 | 1.6 | 2.0 | −3.0 | −1.2 | −0.9 | 2.0 | 0.7 | 4.2 | 4.25 | 1.6 | 3.8 | 1.7 |
| Deviation Tmean 2023 from period 2007–2022 | 3.4 | 0.4 | 1.7 | −3.6 | −1.2 | −1.5 | 1.2 | −0.1 | 3.8 | 4.05 | 0.8 | 2.7 | 1.7 |
| Mean maximum air temperature (°C) | |||||||||||||
| Tmax 1991/2020 | 4.5 | 7.4 | 12.9 | 18.4 | 23.2 | 26.9 | 29.0 | 29.3 | 24.1 | 18.5 | 11.9 | 5.5 | 17.6 |
| Tmax 2007/2022 | 5.3 | 8.6 | 13.2 | 19.1 | 23.2 | 27.5 | 29.8 | 30.1 | 24.5 | 18.5 | 12.7 | 6.6 | 18.3 |
| Tmax 2023 | 8.7 | 9.0 | 14.8 | 15.5 | 22.0 | 26.0 | 31.0 | 30.0 | 28.3 | 23.4 | 13.4 | 10.4 | 19.4 |
| Deviation Tmax 2023 from the reference period | 4.2 | 1.6 | 2.0 | −2.9 | −1.2 | −0.9 | 2.0 | 0.7 | 4.2 | 4.9 | 1.5 | 4.9 | 1.7 |
| Deviation Tmax 2023 from period 2007–2022 | 3.4 | −0.4 | 1.7 | −3.6 | −1.2 | −1.5 | 1.2 | −0.1 | 3.8 | 4.9 | 0.7 | 3.8 | 1.1 |
| Mean minimum air temperature (°C) | |||||||||||||
| Tmin 1991/2020 | −2.3 | −1.0 | 2.6 | 7.1 | 11.8 | 15.4 | 16.8 | 16.9 | 12.6 | 7.9 | 3.6 | −0.9 | 7.5 |
| Tmin 2007/2022 | −1.6 | 0.1 | 3.1 | 7.4 | 12.1 | 16.2 | 17.6 | 17.5 | 13.1 | 8.2 | 4.4 | 0.2 | 8.2 |
| Tmin 2023 | 2.5 | 0.0 | 4.0 | 5.8 | 11.9 | 16.1 | 18.7 | 18.0 | 15.8 | 11.4 | 4.9 | 1.9 | 9.2 |
| Deviation Tmin 2023 from the reference period | 4.2 | 1.6 | 2.0 | −3.0 | −1.2 | −0.9 | 2.0 | 0.7 | 4.2 | 3.4 | 1.3 | 2.8 | 1.7 |
| Deviation Tmin 2023 from period 2007–2022 | 3.4 | 0.4 | 1.7 | −3.6 | −1.2 | −1.5 | 1.2 | −0.1 | 3.8 | 3.2 | 0.5 | 1.7 | 1.1 |
| Total and mean amounts of precipitation (mm) | |||||||||||||
| Period | I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | ∑ |
| Prec 1991/2020 | 42.4 | 34.0 | 41.7 | 47.4 | 68.1 | 80.1 | 58.2 | 54.0 | 56.0 | 50.7 | 45.5 | 48.3 | 626.4 |
| Prec 2007/2022 | 44.9 | 38.2 | 48.4 | 40.4 | 82.5 | 74.2 | 51.0 | 45.3 | 52.3 | 50.0 | 48.5 | 50.0 | 625.8 |
| Prec 2023 | 71.6 | 54.4 | 29.9 | 58.6 | 70.1 | 102.0 | 43.4 | 60.6 | 46.9 | 16.8 | 93.0 | 31.6 | 678.9 |
| Deviation Prec 2023 from the reference period | 69 | 60 | −28 | 24 | 3 | 27 | −25 | 12 | −16 | −67 | 104 | −35 | 8 |
| Deviation Prec 2023 from period 2007–2022 | 59 | 43 | −38 | 45 | −15 | 37 | −15 | 34 | −10 | −66 | 92 | −37 | 8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrov, D.; Ocokoljić, M.; Galečić, N.; Skočajić, D.; Simović, I. Adaptability of Prunus cerasifera Ehrh. to Climate Changes in Multifunctional Landscape. Atmosphere 2024, 15, 335. https://doi.org/10.3390/atmos15030335
Petrov D, Ocokoljić M, Galečić N, Skočajić D, Simović I. Adaptability of Prunus cerasifera Ehrh. to Climate Changes in Multifunctional Landscape. Atmosphere. 2024; 15(3):335. https://doi.org/10.3390/atmos15030335
Chicago/Turabian StylePetrov, Djurdja, Mirjana Ocokoljić, Nevenka Galečić, Dejan Skočajić, and Isidora Simović. 2024. "Adaptability of Prunus cerasifera Ehrh. to Climate Changes in Multifunctional Landscape" Atmosphere 15, no. 3: 335. https://doi.org/10.3390/atmos15030335
APA StylePetrov, D., Ocokoljić, M., Galečić, N., Skočajić, D., & Simović, I. (2024). Adaptability of Prunus cerasifera Ehrh. to Climate Changes in Multifunctional Landscape. Atmosphere, 15(3), 335. https://doi.org/10.3390/atmos15030335