Abstract
Atmospheric sounding using the Tu-134 Optik aircraft-laboratory was conducted in September 2020 over the seas of the Russian sector of the Arctic Ocean, namely the Barents, Kara, Laptev, East Siberian, Chukchi and Bering seas. Unique samples of atmospheric aerosols at altitudes from 200 and up to 10,000 m were taken, including samples for the identification of cultivated microorganisms and their genetic analysis. Data on the concentration and diversity of bacteria and fungi isolated from 24 samples of atmospheric aerosols are presented; the main phenotypic and genomic characteristics were obtained for 152 bacterial cultures; and taxonomic belonging was determined. The concentration of cultured microorganisms detected in aerosols of different locations was similar, averaging 5.5 × 103 CFU/m3. No dependence of the number of isolated microorganisms on the height and location of aerosol sampling was observed. The presence of pathogenic and condto shitionally pathogenic bacteria, including those referred to in the genera Staphylococcus, Kocuria, Rothia, Comamonas, Brevundimonas, Acinetobacter, and others, as well as fungi represented by the widely spread genera Aureobasidium, Aspergillus, Alternaria, Penicillium, capable of causing infectious and allergic diseases were present in most analyzed samples. Obtained data reveal the necessity of systematic studies of atmospheric microbiota composition to combat emerging population diseases.
1. Introduction
The polar regions of the Earth, characterized by low average annual temperatures, have long attracted the attention of scientists from the point of view of studying the microbiota present there. The first microbiological studies in the Russian Arctic were carried out in areas adjacent to the mainland and islands—in the shelf part of the Arctic seas or on the continental slope—and were published back in 1914 [1]. Studies of microorganisms began to cover not only terrestrial, but also aquatic samples [2], as well as samples of Arctic atmospheric air obtained using aircrafts [3,4,5] in the mid-20th century.
Recently, progressive environmental changes have been observed in the Arctic: the melting of glaciers and permafrost and the reduction in the ice area of the Arctic Ocean, which also affects the composition of the biota [6,7]. The Arctic is affected by global warming, and the temperature increase observed here is twice that of similar changes in other parts of the globe [8,9]. When studying the organic and biological components of the atmosphere, one of the main types of research activity in the polar regions remains marine expeditions using ice-class vessels and polar regions monitoring groups using Arctic stations [10,11,12,13,14,15,16]. The dominant species were the fungi Aureobasidium pullulans, Chaetomium globosum, Cladosporium cladosporioides, Penicillium spp. and a number of others, with numbers up to 2.54 × 102 CFU/m3 of air, which is consistent with the results obtained in cold regions such as Antarctica and Canada [17,18,19,20,21,22]. A significant part of the identified fungi were opportunistic human pathogens. The key routes of microbial dispersal are anthropogenic transfer (invasion) and the spread of microorganisms through the air. A noticeable deterioration in the condition of people, especially those with weakened immune systems, is possible if there are significant microorganism concentrations in the air. In this regard, there is a need to study aerosols of organic and inorganic composition containing biogenic components entering the region.
As existing reviews indicate [23,24], there is no unified monitoring network in the Arctic region and, therefore, aircraft sounding is used to study the composition and concentration of aerosols [3,4,5,14,20,25,26]. The aircraft method for studying the composition of atmospheric air is the most accurate and widely used in the Arctic, however, information on the number and species composition of the aeromicrobiota in the Arctic Russian territories is scarce in the literature, mainly relating to the concentration and composition of fungi in the near surface atmosphere [27,28]. Information on bacteria in atmospheric aerosols of the Arctic is practically absent.
The purpose of this study was to obtain data on the vertical distribution of bacteria and fungi in atmospheric aerosols during aircraft sounding of the atmosphere over the seas of the Arctic Ocean of the Russian sector of the Arctic: Barents, Kara, Laptev, East Siberian and Chukotka, as well as over the Bering Sea at altitudes of 200 and up to 10,000 m.
2. Materials and Methods
2.1. Flight Route
Airborne sounding of the atmosphere was carried out using a modified Tu-134 Optik laboratory aircraft [26,29]. The route diagram of the entire experiment is shown in Figure 1. Profiles of measured values were obtained from the minimum permissible heights (red and other warm tones of trajectories on the schematic map) to the upper troposphere (purple tones of trajectories on the schematic map).
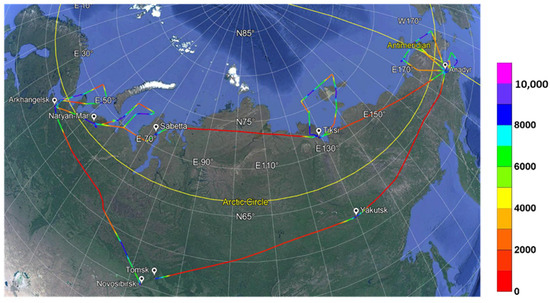
Figure 1.
Schematic map of the routes of the Optik Tu-134 aircraft laboratory in the regions of the Russian Arctic. Figure 1 is constructed based on data from the work presented in [26].
2.2. Aerosol Sampling
Atmospheric aerosols were collected into impingers manufactured by JSC Experimental Design Bureau of Fine Biological Engineering, Kirishi, Russia. The device is made of stainless steel with a critical nozzle that ensures constant flow rate at a pressure difference of more than 4 × 104 Pa of air through the device. The required pressure drop is ensured by the difference in the pressure of the flow entering the air intake when the aircraft is moving and the pressure is outside. The particles are inertially deposited into the liquid swirled by the incoming flow (with a flow rate of 50 ± 5 L/min) along the walls of the device. In total, 50 mL of Hanks solution (ICN Biomedicals, Costa Mesa, CA, USA) is used as a sorbing liquid. The retention efficiency of this device for aerosols larger than 0.3 μm (the minimum size of known bacteria) exceeds 80%, amounting to an almost constant value of 90 ± 15% for particles with a diameter of more than 2 μm. To eliminate the loss of aerosol particles in the connecting tubes, impingers are located near the isokinetic sample inlet. Geographical sampling conditions for microbiological analyzes are presented in Table 1. This table also presents the detected concentrations of culturable microorganisms in the air included in the samples.

Table 1.
Sampling conditions of bioaerosols samples and the concentration of culturable microorganisms in the air samples taken in 2020 (expressed in number of Colony Forming Units (CFU) per cubic meter). Time is shown in Greenwich Mean Time.
2.3. The Concentration of Culturable Microorganisms
The concentration of culturable microorganisms was determined by sowing aerosol samples onto liquid and agar nutrient media: LB (Difco, Franklin Lakes, NJ, USA)—to identify saprophytic bacteria; depleted LB medium (1:10 dilution)—for isolating microorganisms inhibited by excess organic matter; starch-ammonia medium [30]—to identify actinomycetes; soil agar—for soil microorganisms; Sabouraud’s medium [31]—for identifying fungi. Petri dishes with inoculations were incubated at a temperature of 28–30 °C and—to identify psychrophiles—at 6–10 °C for 3–14 days. The calculation of the number of culturable microorganisms in samples, expressed in decimal logarithms of the number of colony-forming units (CFU), was carried out according to standard methods [32]. Considering the size of the volumes of the atmospheric air samples taken for analysis, the minimum threshold for detecting the concentration of viable fungi in the atmosphere was 40 CFU/m3, and for bacteria was 100 CFU/m3. The error in determining the concentration of microorganisms for the cultivation conditions described above was ±0.2 log10.
2.4. The Phenotypic Characteristics of the Isolated Microorganisms
The phenotypic characteristics of the isolated microorganisms were determined in accordance with the guidelines [30]. The morphological features of the isolates were studied by observing cells of vital- and fixed-stained preparations using an Axioskop 40 microscope “Carl Zeiss” (Gottingen, Germany).
2.5. The Enzymatic Activity of the Isolated Microorganisms
The enzymatic activity of the isolated microorganisms was determined by testing the proteolytic, amylolytic, lecithinase, nuclease, and lipase activities of the isolates [29]. Pathogenic properties were determined by the presence of alkaline phosphatase, hemolytic, phospholipase (lecithinase), plasmacoagulase, fibrinolytic, and gelatinase activities [33].
2.6. Destruction of Oil
Microbial cultures isolated from Arctic aerosols were tested for their ability to degrade oil by sowing on agarized and liquid 8E mediums [34]. Up to 1–5% of high-viscosity oil from the Usinsk field (Komi Republic, Russian Federation) was added to the medium as the only carbon source. The average density of oil was 0.920–0.986 g/cm3.
2.7. Microorganisms’ Antibiotic Properties
To determine the resistance of coccus strains to antibiotics using the disc diffusion method, the following drugs were used (µg/disc): gentamicin (10), ciprofloxacin (30), kanamycin (30), vancomycin (30), levofloxacin (5), oxacillin (10), benzylpenicillin (10 units), erythromycin (15), clindomycin (2). For spore-forming bacteria, a slightly different recommended set was used: gentamicin (10), ciprofloxacin (5), vancomycin (30), levofloxacin (5), norfloxacin (10), imipenem (10), linozolid (30). The discs with the antibiotics used were produced by NICF LLC (Russia). The Mueller–Hinton medium was used in the experiment according to [35].
The antibiotic activity of spore-forming bacteria was studied using the method of delayed antagonism [36] against opportunistic test strains of bacteria and yeast: Escherichia coli 6645 ATCC 25922, Candida albicans 620 Y-583, Staphylococcus aureus ATCC 6538, Klebsiella pneumoniae B-378 B-4894, and Bacillus cereus ATCC 1070 2 from the collection of bacteria, bacteriophages, and fungi at the Federal Budgetary Research Institution «State Research Center for Virology and Biotechnology “Vector”» of Rospotrebnadzor.
2.8. Microorganisms’ Taxonomy
The taxonomy of microorganism strains was determined based on the total results of phenotypic and genomic analysis. Molecular biological identification of bacteria was carried out by determining the nucleotide sequence of a fragment (about 700 bp) of the 16S ribosomal RNA gene [37]. Total nucleic acids were isolated from the prepared material using a commercial reagent kit “RIBO-prep” (InterLabService, Moscow, Russia) in accordance with the manufacturer’s instructions. Next, PCR was carried out using the PCR mixture “5X ScreenMix” (Evrogen, Moscow, Russia) and specific primers for amplification of the 16S rRNA gene fragment (oligodeoxyribonucleotide primers ACTCCTACGGGAGGCAGCAG and CGACRRCCATGCANCACCT; with a 5′→3′ structure). Detection of PCR products was carried out using gel electrophoresis [38]. Determination of the nucleotide sequences of PCR products purified using the AMPure XP Reagent kit was carried out using the BigDye™ Terminator v.3.1 Cycle Sequencing Kit on an ABI PRISM 3500 Genetic Analyzer automatic sequencer (Applied Biosystems, Foster City, CA, USA) [39]. Analysis of the obtained nucleotide sequences was carried out using the Sequencher program. Identification of microorganisms based on analysis of the primary nucleotide sequence was carried out in the GenBank/EMBL/DDBJ database (http://www.ncbi.nlm.nih.gov, accessed on 12 September 2023) using multiple alignment in the BLASTn program (http://www.ncbi.nlm.nih.gov, accessed on 12 September 2023).
Fungi were mainly identified by morpho-physiological characteristics, molecular genetic methods were used selectively for a small number of strains.
ITS (intergenic ribosomal spacer sequence) was used for the identification of fungal strains using molecular genetic methods. The amplification reaction of ITS-fragment of genomic DNA was carried out using oligonucleotides ITS1 and ITS4 for the first round of PCR (obtained fragment~700 bp), ITS3 and ITS4 for the second round of PCR (300 bp). Sequencing reaction of the obtained PCR fragments was performed using the same oligonucleotides and BigDye v.3.1 reagent (Applied Biosystems, USA) under standard conditions. The obtained sequences were analyzed using ABI Sequence Scanner and FinchTV 4.1 software. Sequences were compared with those available in the NCBI GenBank database (http://www.ncbi.nlm.nih.gov, accessed on 12 September 2023) using the BLASTN algorithm, and phylogenetic analysis was performed using the MEGA 7.0.21 program.
The studied strains are stored for low-temperature freezing in the collection of bacteria, bacteriophages, and fungi at the Federal Budgetary Research Institution «State Research Center for Virology and Biotechnology “Vector”» of Rospotrebnadzor.
3. Results
3.1. Microorganisms Concentrations and Diversity
Flights were carried out over the Barents Sea, White Sea, Kara Sea, Laptev Sea, East Siberian Sea, Chukchi Sea and over the Anadyr Gulf of the Bering Sea. For microbiological analysis, 24 samples of atmospheric aerosols were taken into impingers at altitudes from 200 to 10,000 m. Analysis of the reverse trajectories of air masses from which samples were taken using the HYSPLIT program [40,41] shows that their trajectories mainly passed through the northern territories and water areas that are not rich in powerful sources of bioaerosols. Only some backward trajectories for samples with the highest concentrations of culturable microorganisms came from more southern regions (Supplementary Materials, Figures S1–S6). Consequently, there was no reason to expect high concentrations of culturable microorganisms in all air samples over the Arctic seas.
The concentration of culturable microorganisms found in all samples averaged 5.5 × 103 CFU/m3, which is close to the concentrations of atmospheric bioaerosols previously discovered for the more southern region of Russia [42,43]. There was no dependence on the number of isolated microorganisms on the height of aerosol sampling (Table 1). From the studied aerosol samples, 252 culturable isolates of bacteria and fungi were isolated. The obligate psychrophiles expected in samples of Antarctic aerosols, cultivated under experimental conditions, were not detected.
Fungi, including representatives of the widespread genera Penicillium, Aspergillus, and Aureobasidium, were identified only in seven of 24 studied samples, occurring both at high altitudes up to 9000 m and at the level 200–300 m above the water surface. Their highest concentration was isolated from a sample taken over the Barents Sea at an altitude of 500-200 m, accounting for 30% of the number of culturable microorganisms isolated from this sample. In the remaining samples, fungi were either not detected or were present in insignificant quantities (from 0.14 to 4.41%, Figure 2, Table 1 and Table 2). The isolated fungi can cause both infectious diseases and allergic reactions. According to the results of studies conducted in pulmonology clinics in different countries, the frequency of sensitization to Aspergillus spp. in patients with bronchial asthma in China was 5.5%; in New Zealand 18.4%; in Saudi Arabia 22.6%; in India 38.5%; in Russia 36%; the number of patients with this disease in the world is approximately 6 million, in Russia this number is 175 thousand [44,45,46].
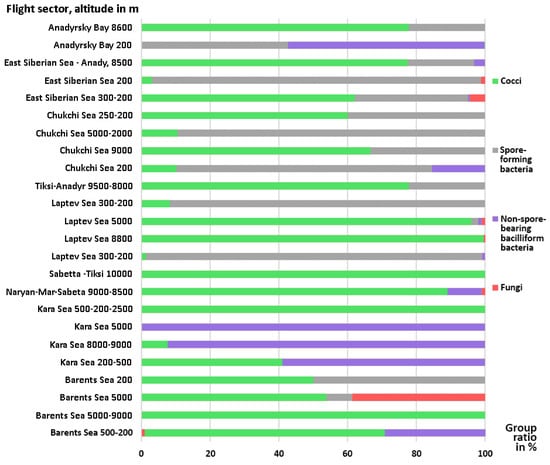
Figure 2.
The ratio of groups of detected culturable microorganisms in samples of Arctic aerosols selected during aircraft sounding of the atmosphere in different regions at different altitudes.
Table 2.
Diversity of culturable microorganisms in samples of atmospheric bioaerosols of the Arctic, determined from the results of analysis of phenotypic and genomic characteristics.
Gram-positive endospore-forming bacteria of the genera Lysinibacillus and Bacillus were most abundant in aerosols at altitudes of 200–300 m, accounting for up to 97.8% of the total number of microbial isolates isolated from the sample (Figure 2). Two samples collected at high altitudes of 9000 and 2000–5000 m above the Chukchi Sea were exceptions and also contained noticeable amounts of spore-forming bacteria—33.3 and 89.4%, respectively, of the total isolated from the samples. In other samples, bacilli were observed in much smaller quantities or were absent.
Non-spore-forming bacteria were isolated from 12 samples found at different altitudes: 15, 30, 57.6, and 58.8% of the total number of isolated non-spore-forming bacteria were in samples taken at altitudes of 500-200 m (Chukchi, Barents, Kara Sea, Anadyr Sea). bay, respectively). The largest number of these were isolated from three samples over the Kara Sea (up to 99.78%) and in a sample taken at an altitude of 200 m above the Gulf of Anadyr—57.06% of those isolated from the sample (Figure 2). It is possible that these samples were taken in locations where the influence of land-based sources was small. The composition and concentration of isolated microorganisms corresponds to the information that oceanic air masses contain mainly Gram-negative bacteria of marine origin caught in the aerosol from the surface of the water, since it is Gram-negative bacteria that are up to 80-95% of the total number of bacteria in seawater [47].
Coccal forms of bacteria isolated from most of the samples were represented by both saprotrophic bacteria of the genus Micrococcus and a group of opportunistic bacteria of the genera Staphylococcus, Acinetobacter, Rothia and a number of others that can cause infectious diseases (Table 2).
Among the isolates of this group, bacteria were found that have signs of pathogenicity: phosphatase, lecithinase, and hemolytic activities. Pathogenic microorganisms use a number of genetic strategies to invade the host—the secretion of aggression enzymes is one of them. Testing for signs of pathogenicity of 145 bacterial isolates from different samples revealed 30 spore-forming bacteria, 15 cocci, and 20 non-spore-forming bacteria that secrete hemolysins which destroy red blood cells. In total, 45 bacterial cultures possessed lipolytic enzymes of varying activity and specificity involved in the destruction of cell membranes. Bacteria belonging to the genera Bacillus and Staphylococcus were discovered, producing highly active phospholipases that determine the invasiveness of microorganisms.
Such a sign of pathogenicity as the production of alkaline phosphatase was found in most of the 145 tested strains, among them the most active were 20 cultures of the genus Bacillus, 17 cocci of the genus Staphylococcus and Jeotgaliococcus, and 15 non-spore-bearing bacteria of the genera Acinetobacter, Brevundiomonas, and Microbacterium.
3.2. Antibiotic Activities of Isolates
Important are the ongoing studies to determine the antibiotic activity of microbial isolates from the Arctic and Antarctic in order to identify new promising producers of antibiotics that make it possible to overcome the increasing resistance to antimicrobial drugs of clinical strains of pathogens [48,49,50,51].
The antibiotic activity of 34 spore-forming bacteria was studied using the method of delayed antagonism against opportunistic test cultures E. coli, C. albicans, S. aureus, K. pneumoniae, and B. cereus. Almost all strains of bacilli effectively inhibited the growth of the Candida strain, different effects indicate different sets of compounds secreted by the bacilli (zones of lysis and growth inhibition were up to 45 mm, Table S1), 16 out of 34 tested spore-forming bacteria inhibited the growth of S. aureus, 9 strains of B. cereus. E. coli zones were negligible. The K. pneumoniae strain was resistant to secreted metabolites of all bacilli studied.
The presence and concentration of staphylococci in the air determines the degree of its contamination by pathogenic microorganisms. Control of the resistance of such atmospheric contaminants to antibiotics is an important factor in determining their danger to public health. Of the 36 tested strains of the staphylococcal group, 15 showed resistance to benzylpenicillin, expressed to varying degrees, 7 strains were resistant to kanamycin, 3 to vancomycin, and one each to gentamicin and oxacillin (Tables S2 and S3). In general, one can say that the staphylococcal strains isolated from northern aerosols showed high sensitivity to the antibiotics used in the experiment in contrast to the control strains S. aureus MRSA B-1352 and S. epidermidis MRSE B-1350, represented by clinical isolates. The test strain S. aureus 209-p B-1266, recommended by the Russian Pharmacopoeia for monitoring drug resistance of strains, showed sensitivity to all drugs used (Tables S2 and S3).
Spore-forming bacteria also showed high sensitivity to antibiotic drugs with a few exceptions (Tables S4 and S5). Three strains Sp-32, Sp-69 and Sp-127, resistant to 3–5 antibiotics, can be classified as multi-resistant. Linozolid turned out to be the least effective antibiotic, while 16 out of 40 tested strains showed resistance to it.
3.3. Bacterial Growth at Different Temperatures
Another sign that indirectly indicates the ability of a microorganism to cause a pathogenic process is its active growth at a temperature of 37 °C. The studied strains were tested for their ability to grow in the temperature range of 6–37 °C when sown on the agar medium of the Russian Federation and incubated for 48 h, the results obtained are presented in Table 3. Eighteen strains out of 145 tested were distinguished by active growth at a temperature of 6–9 °C are classified as psychrotolerant bacteria. The rest were mesophilic microorganisms, and more than half of them grew very well at 37 °C (Table 3).
Table 3.
Ability of the studied strains to grow at different temperatures.
3.4. Biotechnological Properties of Isolates
A significant aspect of the study of microorganisms in polar habitats is their ability to secrete various lytic compounds at low temperatures, which can be used in biotechnological processes and in reclamation processes carried out in similar temperature conditions [52,53,54,55,56]. The results of testing isolated bacteria and yeasts as oil destructors are provided in the work presented in [34]. The ability to grow at different temperature conditions was previously demonstrated in Table 3. Psychrotolerant microorganisms capable of active destruction of oil at low temperatures include bacterial strains Sp-1, Sp-16, Sp-18, Sp-22, Sp-22-1, and Sp-116, as well as yeast Sp-71, Sp-91 (Figure 3B,C). The most active oil destructor turned out to be the psychrotolerant strain Rhodococcus sp Sp-116 [34], deposited in the collection of bacteria, bacteriophages, and fungi under No. B-1388. During 10 days of incubation of this strain at room temperature, oil added up to 2% to the cultivation medium was almost completely utilized (Figure 3A).

Figure 3.
Oil utilization by Rhodococcus sp. Sp-116 (A) and emulsification of petroleum products (B,C) by yeast strains Sp-71 and Sp-91 isolated from aerosols.
4. Discussion
The formation of the Arctic aeromycobiota occurs both due to the arrival of microorganisms from natural landscapes and due to the appearance of bacteria and fungi as a result of anthropogenic transfer (invasion) that has increased in the Arctic territory in recent years. The dominant agents of biological transport between land, water, and the atmosphere are air flows that facilitate the transfer of particles with immobilized microorganisms and other biological objects over large horizontal distances and heights. An increase in the concentration of fungi of the genera Penicillium, Aspergillus, and others in Arctic aerosols growing at 37 °C is associated with their possible pathogenicity for humans [26], which requires control of the concentration and composition of atmospheric mycota to prevent possible fungal infections. The work presented in [57] showed that the majority of the mycobiota in the studied air samples consisted of biodegraders of various materials and conditional human pathogens. The dominant species and their uneven distribution in the air of residential and work premises and in adjacent areas were identified. There is a dominance of dark-colored microorganisms producing melanins, which are well-adapted to overcome unfavorable environmental conditions. In our studies, these are fungi of the genera Aspergillus and Aureobasidium (Table 2).
Knowledge of the sources and transport pathways of bioaerosols is necessary for a comprehensive understanding of the role of microorganisms in the atmosphere and control of the spread of associated epidemic diseases. The coronavirus disease 2019 (COVID-19) pandemic has raised widespread concerns about bioaerosols, which may have significant public health impacts. The Arctic aerial microbiota is formed due to the supply of microorganisms from natural local landscapes, large-scale propagation events associated with the supply of biomaterials from distant sources, and also as a result of increased anthropogenic load, leading to risks of biological pollution, which contributes to the spread of cosmopolitan species in the Arctic, such as pathogenic fungi Aspergillus, Penicillium, and others. Transoceanic and transcontinental dust events play a role in the transfer of pathogens, expanding the biogeographic range of organisms through long-distance dispersal [58]. Satellite observations have shown atmospheric transport of dust from China more than 10,000 km over the Pacific Ocean, and Eurasia has been identified as the main source of anthropogenic aerosol pollution in the polar regions of North America [59]. Pathogens moving through the air over long distances are able to spread diseases across continents and between them [60]. Seasonal meningococcal meningitis in Africa [61] and valley fever caused by Coccidioides spores in arid regions on both American continents are associated with the transfer of the pathogen with dust components [62]. The spread of microorganisms over long distances in unfavorable atmospheric conditions is ensured by various protective means, due to the production of extracellular polymeric substances and pigments, the formation of spores or other dormant structures, association with vectors, and also due to osmo- and thermo-protectors, antioxidants, or specific enzymes [63]. The long-held view of atmospheric transport as neutral to dispersal outcomes in ecology is no longer tenable.
Noteworthy is the presence in Arctic aerosols of bacteria of the genus Staphylococcus, which belong to the fourth pathogenicity group (Table 2). Species such as S. epidermidis, S. warneri, and S. hominis isolated from aerosols are detected primarily in hospital-acquired infections. Found as a harmless commensal on the skin of humans and animals, these bacteria, having increased virulence and pathogenicity as well as frequent resistance to antibiotics, are capable of causing infections in patients with weakened immune systems. The presence of staphylococci in aerosols of cold habitats was previously shown in the literature [64].
Bacteria of the genus Acinetobacter isolated from the studied aerosols are free-living saprophytes, but can cause meningitis and septicemia in humans against the background of suppressed immune system function [65]. Among the isolated microbial isolates, similar information about the possibility of causing diseases is known for microorganisms of the genus Kocuria, which cause infections in patients with weakened immune systems [66]; for bacteria of the genus Curtobacterium of the order Actinomycetales, isolated from clinical material during infectious processes of various localizations; for bacteria of the genus Rothia from the family Micrococcaceae [67]; as well as for bacteria of the genus Comamonas, isolates of which are obtained mainly from plants and soil habitats, but are also capable of causing an infectious disease [68]. Opportunistic bacteria of the genus Brevundimonas have been isolated; they are usually isolated from patients with impaired immunity and concomitant chronic diseases [69]. Saprotrophic microorganisms of the genus Pseudarthrobacter, representatives of the widespread genera Bacillus, Lysinibacillus, Nocardia, and symbiotic bacteria of the genus Rhizobium were also isolated from Arctic aerosols. Facultatively anaerobic, halotolerant bacteria of the genus Jeotgalicoccus, known for their ability to grow at NaCl concentrations from 0.1 to 16%, were isolated too [70].
Pathogenic microorganisms use a number of genetic strategies to invade the host, and the secretion of aggression enzymes is one of them. As mentioned above, testing for signs of pathogenicity of 145 bacterial isolates from different samples revealed 30 spore-forming bacteria, 15 cocci, and 20 non-spore-forming bacteria that secrete hemolysins that destroys red blood cells. Fifty-five cultures possessed lipolytic enzymes of varying activity and specificity involved in the destruction of cell membranes. Bacteria belonging to the genera Bacillus and Staphylococcus were found to produce highly active phospholipases that determine the invasiveness of microorganisms [71]. Such a sign of pathogenicity as the production of alkaline phosphatase was found in most of the tested strains. Among them, the most active were 20 cultures of the genus Bacillus, 17 cocci of the genus Staphylococcus and Jeotgaliococcus, and 15 non-spore-forming bacteria of the genera Acinetobacter, Brevundiomonas, and Microbacterium.
Global climatic changes and increasing anthropogenic load imply further changes in the Arctic biota and enrichment of the air environment with microorganisms that was not typical before. The results obtained are preliminary and need further research. For an objective assessment of the data obtained, additional flights with route repetition are required. Further studies of the Arctic microbiota are planned as part of a comprehensive expedition using the Tu-134 Optik laboratory aircraft. Unfortunately, aircraft sounding is an expensive research method and cannot be used as much as researchers would like in order to obtain the most correct data. We hope that integration of international research programs will help to obtain comprehensive monitoring results.
5. Conclusions
Analysis of the studied samples of Arctic atmospheric aerosols revealed a significant diversity of culturable bacteria and fungi with concentrations averaging 5.5 × 103 CFU/m3, regardless of altitude and sampling location at altitudes from 200 m to 10,000 m.
The presence in aerosols of opportunistic fungi and bacteria, including representatives of the genus Staphylococcus and antibiotic-resistant bacteria, makes it necessary to control the microbiological composition of aerosols to prevent infectious diseases.
The discovered highly effective psychrotolerant bacteria—oil destructors—are suitable for the creation of consortia of microorganisms and for use in the remediation of oil-contaminated areas of Siberia and the Arctic.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/atmos15030365/s1, Figure S1: Backward trajectory of air masses from which an air sample was taken at an altitude of 200 m above the Barents Sea; Figure S2: Backward trajectories of air masses from which an air sample was taken at altitudes of 200–500 m above the Kara Sea; Figure S3: Backward trajectories of air masses from which an air sample was taken at altitudes of 200–300 m above the Laptev Sea; Figure S4: Backward trajectory of air masses from which an air sample was taken at an altitude of 500 m above the East Siberian Sea; Figure S5: Backward trajectory of air masses from which an air sample was taken at an altitude of 9000 m above the Chukchi Sea; Figure S6: Backward trajectories of air masses from which an air sample was taken at altitudes 2000–5000 m above the Chukchi Sea; Table S1: Antifungal effect of bacilli strains on the pathogenic strain of Candida albicans determined using the diffusion method on an agarized nutrient medium; Table S2: Sensitivity determination to antibiotics of strains belonging to the group of staphylococci using the disk diffusion method; Table S3: Interpretation of growth retardation zone diameters in determining the sensitivity of cocci to antimicrobial agents using the disk-diffusion method; Table S4: Sensitivity determination to antibiotics using the disk diffusion method of strains of spore-forming bacteria; Table S5: Interpretation of growth retardation zone diameters in determining the sensitivity of strains of spore-forming bacteria to antimicrobial agents using the disk-diffusion method.
Author Contributions
Conceptualization, B.D.B., I.S.A. and A.S.S.; methodology, A.S.S. and I.S.A.; validation, I.S.A., G.A.B. and O.V.O.; formal analysis, I.S.A. and G.A.B.; investigation, O.V.O., I.S.A., L.I.P., M.E.R., N.A.S. and D.V.S.; resources, B.D.B. and A.S.S.; writing—original draft preparation, I.S.A., G.A.B. and A.S.S.; writing—review and editing, A.S.S.; supervision, A.S.S.; project administration, B.D.B.; funding acquisition, B.D.B. and A.S.S. All authors have read and agreed to the published version of the manuscript.
Funding
The work was funded by State orders of Rospotrebnadzor GZ 14/18 and GZ 11/21, State order of IAO SB RAS (agreement No. 121031500342-0), and agreement no. 075-15-2021-934 with Ministry of Science and Higher Education of the Russian Federation.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article and supplementary materials.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Issatchenko, B.L. Research on Bacteria in the Arctic Ocean; Tipografia V.O. Kirshbauma: Petrograd, Russia, 1914; Available online: https://elib.rgo.ru/safe-view/123456789/218205/1/UnVQUkxJQjEyMDg0Mjc5LlBERg (accessed on 16 January 2024). (In Russian)
- Kriss, A.E. Marine Microbiology: (Deep Sea); Publishing House of the USSR Academy of Sciences: Moscow, Russia, 1955. (In Russian) [Google Scholar]
- Kelly, C.D.; Layn, S. Bacteria found in the air over Canada and the American Arctic. Can. J. Microbiol. 1957, 3, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Pady, S.M.; Kapica, L. Air-borne fungi in the Arctic and other parts of Canada. Can. J. Bot. 1953, 31, 309–323. [Google Scholar] [CrossRef]
- Pady, S.M.; Kelly, C.D.; Polunin, N. Arctic aerobiology II: Preliminary report on fungi and bacteria isolated from the air in 1947. Nature 1948, 162, 379–381. [Google Scholar] [CrossRef] [PubMed]
- Overpeck, J.; Hughen, K.; Hardy, D.; Bradley, R.; Case, R. Arctic environmental change of the last four centuries. Science 1997, 278, 1251–1256. [Google Scholar] [CrossRef]
- Schmale, J.; Zieger, P.; Ekman, A.M.L. Aerosols in current and future Arctic climate. Nat. Clim. Chang. 2021, 11, 95–105. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2013: The Physical Science Basis; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- Serreze, M.C.; Barrett, A.P.; Stroeve, J.C.; Kindig, D.N.; Holland, M.M. The emergence of surface-based Arctic amplification. Cryosphere 2009, 3, 11–19. [Google Scholar] [CrossRef]
- Johansen, S.; Hafsten, U. Airborne pollen and spore registrations at Ny-Ålesund, Svalbard, summer 1986. Polar Res. 1988, 6, 11–17. [Google Scholar] [CrossRef][Green Version]
- Feltracco, M.; Barbaro, E.; Hoppe, C.J.M.; Wolf, K.K.E.; Spolaor, A.; Layton, R.; Keuschnig, C.; Barbante, C.; Gambaro, A.; Larose, C. Airborne bacteria and particulate chemistry capture Phytoplankton bloom dynamics in an Arctic fjord. Atmos. Environ. 2021, 256, 118458. [Google Scholar] [CrossRef]
- Jensen, L.Z.; Glasius, M.; Gryning, S.-E.; Massling, A.; Finster, K.; Šantl-Temkiv, T. Seasonal variation of the atmospheric bacterial community in the Greenlandic High Arctic is influenced by weather events and local and distant sources. Front. Microbiol. 2022, 13, 909980. [Google Scholar] [CrossRef]
- Kawana, K.; Taketani, F.; Matsumoto, K.; Miyakawa, T.; Tobo, Y.; Iwamoto, Y.; Ito, A.; Kanaya, Y. Roles of marine biota in the formation of atmospheric bioaerosols, cloud condensation nuclei, and ice-nucleating particles over the North Pacific Ocean, Bering Sea, and Arctic Ocean. Atmos. Chem. Phys. 2023, 24, 1777–1799. [Google Scholar] [CrossRef]
- Ferrero, L.; Sangiorgi, G.; Perrone, M.G.; Rizzi, C.; Cataldi, M.; Markuszewski, P.; Pakszys, P.; Makuch, P.; Petelski, T.; Becagli, S.; et al. Chemical composition of aerosol over the Arctic Ocean from Summer ARctic EXpedition (AREX) 2011–2012 cruises: Ions, amines, elemental carbon, organic matter, polycyclic aromatic hydrocarbons, n-alkanes, metals, and rare earth elements. Atmosphere 2019, 10, 54. [Google Scholar] [CrossRef]
- Yttri, K.E.; Bäcklund, A.; Conen, F.; Eckhardt, S.; Evangeliou, N.; Fiebig, M.; Kasper-Giebl, A.; Gold, A.; Gundersen, H.; Myhre, C.L.; et al. Composition and sources of carbonaceous aerosol in the European Arctic at Zeppelin Observatory, Svalbard. Atmos. Chem. Phys. 2023, 24, 2731–2758. [Google Scholar] [CrossRef]
- Kravchishina, M.D.; Klyuvitkin, A.A.; Novigatsky, A.N.; Glukhovets, D.I.; Shevchenko, V.P.; Belan, B.D. Cruise 89 (First Leg) of the R/V Akademik Mstislav Keldysh: Climate experiment in interaction with the Tu-134 Optik Flying Laboratory. Oceanology 2023, 63, 428–431. [Google Scholar] [CrossRef]
- Duncan, S.M.; Farrell, R.L.; Jordan, N.; Jurgens, J.A.; Blanchette, R.A. Monitoring and identification of airborne fungi at historic locations on Ross Island, Antarctic. Polar Sci. 2010, 4, 275–283. [Google Scholar] [CrossRef][Green Version]
- Li, D.; Kendrick, B. A year-round outdoor aeromycological study in Waterloo, Ontario, Canada. Grana 1995, 34, 199–207. [Google Scholar] [CrossRef]
- Malard, L.A.; Avila-Jimenez, M.-L.; Schmale, J.; Cuthbertson, L.; Cockerton, L.; Pearce, D.A. Aerobiology over the Southern Ocean—Implications for bacterial colonization of Antarctica. Environ. Int. 2022, 169, 107492. [Google Scholar] [CrossRef]
- Kobayashi, F. Direct sampling and bioanalyses of atmospheric bioaerosols using a tethered balloon over Syowa Station, Antarctica. Polar Sci. 2022, 32, 100842. [Google Scholar] [CrossRef]
- Feltracco, M.; Zangrando, R.; Barbaro, E.; Becagli, S.; Park, K.-T.; Vecchiato, M.; Caiazzo, L.; Traversi, R.; Severi, M.; Barbante, C.; et al. Characterization of free L- and D-amino acids in size-segregated background aerosols over the Ross Sea, Antarctica. Sci. Total Environ. 2023, 879, 163070. [Google Scholar] [CrossRef]
- Holmberg, S.M.; Jørgensen, N.O.G. Insights into abundance, adaptation and activity of prokaryotes in arctic and Antarctic environments. Polar Biol. 2023, 46, 381–396. [Google Scholar] [CrossRef]
- Duncan, B.N.; Ott, L.E.; Abshire, J.B.; Brucker, L.; Carroll, M.L.; Carton, J.; Comiso, J.C.; Dinnat, E.P.; Forbes, B.C.; Gonsamo, A.; et al. Space-based observations for understanding changes in the Arctic-Boreal Zone. Rev. Geophys. 2020, 58, e2019RG000652. [Google Scholar] [CrossRef]
- Cusworth, D.H.; Duren, R.M.; Yadav, V.; Thorpe, A.K.; Verhulst, K.; Sander, S.; Hopkins, F.; Rafiq, T.; Miller, C.E. Synthesis of methane observations across scales: Strategies for deploying a multitiered observing network. Geophys. Res. Lett. 2020, 47, GL087869. [Google Scholar] [CrossRef]
- Miller, C.E.; Griffith, P.C.; Goetz, S.J.; Hoy, E.E.; Pinto, N.; McCubbin, I.B.; Thorpe, A.K.; Hofton, M.; Hodkinson, D.; Hansen, C.; et al. An overview of ABoVE airborne campaign data acquisitions and science opportunities. Environ. Res. Lett. 2019, 14, 080201. [Google Scholar] [CrossRef]
- Belan, B.D.; Ancellet, G.; Andreeva, I.S.; Antokhin, P.N.; Arshinova, V.G.; Arshinov, M.Y.; Balin, Y.S.; Barsuk, V.E.; Belan, S.B.; Chernov, D.G.; et al. Integrated airborne investigation of the air composition over the Russian Sector of the Arctic. Atmos. Meas. Tech. 2022, 15, 3941–3967. [Google Scholar] [CrossRef]
- Kirtsideli, I.Y.; Vlasov, D.Y.; Krylenkov, V.A.; Sokolov, V.T. Airborne fungal in the areas of Russian arctic station near White, Barents and Kara sea. Micol. Phytopathol. 2011, 45, 228–239. (In Russian) [Google Scholar]
- Kirtsideli, I.Y.; Abakumov, E.V.; Teshebaev, S.B.; Zelenskaya, M.S.; Vlasov, D.Y.; Krylenkov, V.A.; Ryabusheva, Y.V.; Sokolov, V.T.; Barantsevich, E.P. Microbial communities in regions of Arctic settlements. Hyg. Sanit. 2016, 95, 923–929. [Google Scholar] [CrossRef]
- Anokhin, G.G.; Antokhin, P.N.; Arshinov, M.Y.; Barsuk, V.E.; Belan, B.D.; Belan, S.B.; Davydov, D.K.; Ivlev, G.A.; Kozlov, A.V.; Kozlov, V.S.; et al. OPTIK Tu-134 aicraft laboratory. Atmos. Ocean. Opt. 2011, 24, 805–816. (In Russian) [Google Scholar]
- Gerhardt, F.; Murray, R.G.E.; Wood, W.A.; Krieg, N.R. Methods of General Bacteriology, 2nd ed.; Publisher American Society for Microbiology: Washington, DC, USA, 1994. [Google Scholar]
- Saggie, J. Methods of Soil Microbiology; Kolos Publishers: Moscow, Russia, 1983. (In Russian) [Google Scholar]
- Ashmarin, I.P.; Vorobyov, A.A. Statistical Methods in Microbiological Studies; Medgiz: Leningrad, Russia, 1962. (In Russian) [Google Scholar]
- Labinskaya, A.C.; Volina, E.G. (Eds.) Guide to Medical Microbiology. General and Sanitary Microbiology. Book I; Binom: Moscow, Russia, 2020. (In Russian) [Google Scholar]
- Andreeva, I.S.; Emelyanova, E.K.; Malinkin, A.A.; Rebus, M.E.; Safatov, A.S. Screening of bacteria isolated from Arctic atmospheric aerosols for oil degradation ability. Bull. Nizhnevartovsk State Univ. 2023, 3, 4–17. (In Russian) [Google Scholar] [CrossRef]
- Federal Center for State Sanitary and Epidemiological Surveillance of the Ministry of Health of Russia. Determination of the Sensitivity of Microorganisms to Antibacterial Drugs: Guidelines; Federal Center for State Sanitary and Epidemiological Surveillance of the Ministry of Health of Russia: Moscow, Russia, 2004. (In Russian) [Google Scholar]
- Federal Center for Hygiene and Epidemiology of Rospotrebnadzor. Guidelines for Sanitary and Epidemiological Assessing the Safety and Functional Potential of Probiotic Microorganisms Used for Food Production Products: Guidelines; Federal Center for Hygiene and Epidemiology of Rospotrebnadzor: Moscow, Russia, 2011. (In Russian) [Google Scholar]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef]
- Osterman, L.A. Methods for Studying Proteins and Nucleic Acids: Electrophoresis and Ultracentrifugation; Nauka: Moscow, Russia, 1996. (In Russian) [Google Scholar]
- Watts, D.; MacBeath, J.R. Automated fluorescent DNA sequencing on the ABI PRISM 310 Genetic Analyzer. Methods Mol. Biol. 2001, 167, 153–170. [Google Scholar] [CrossRef]
- Draxler, R.R.; Hess, G.D. An overview of the HYSPLIT_4 modelling system for trajectories, dispersion and deposition. Aust. Meteorol. Mag. 1998, 47, 295–308. [Google Scholar]
- Freitag, S.; Clarke, A.D.; Howell, S.G.; Kapustin, V.N.; Campos, T.; Brekhovskikh, V.L.; Zhou, J. Combining airborne gas and aerosol measurements with HYSPLIT: A visualization tool for simultaneous evaluation of air mass history and back trajectory consistency. Atmos. Meas. Tech. 2014, 7, 107–128. [Google Scholar] [CrossRef]
- Safatov, A.S.; Buryak, G.A.; Olkin, S.E.; Reznikova, I.K.; Makarov, V.I.; Popova, S.A. Analysis of monitoring data on organic/elemental carbon and total protein in ground air layer aerosol in the South of Western Siberia. Atmos. Ocean. Opt. 2014, 27, 164–168. [Google Scholar] [CrossRef]
- Safatov, A.; Andreeva, I.; Buryak, G.; Olkin, S.; Reznikova, I.; Belan, B.; Panchenko, M.; Simonenkov, D. Long-term studies of biological components of atmospheric aerosol: Trends and variability. Atmosphere 2022, 13, 651. [Google Scholar] [CrossRef]
- Klimko, N.N.; Kozlova, Y.I.; Khostelidi, S.N.; Shadrivova, O.V.; Borzova, Y.V.; Vasilyeva, N.V. The prevalence of serous and chronic fungal diseases in Russian Federation on Life Program Model. Probl. Med. Mycol. 2014, 1, 3–9. (In Russian) [Google Scholar]
- Kozlova, Y.I.; Sobolev, A.V.; Frolova, E.V.; Aak, O.V.; Burygina, E.V.; Klimko, N.N. Allergic bronchopulmonary aspergillosis in patients with asthma. Russ. Allergol. J. 2015, 2, 37–46. (In Russian) [Google Scholar] [CrossRef]
- Maturu, V.N.; Agarwal, R. Prevalence of Aspergillus sensitization and allergic bronchopulmonary aspergillosis in cystic fibrosis: Systematic review and meta-analysis. Clin. Exp. Allergy 2015, 45, 1765–1778. [Google Scholar] [CrossRef] [PubMed]
- Alsante, A.N.; Thornton, D.C.O.; Brooks, S.D. Ocean aerobiology. Front. Microbiol. 2021, 12, 764178. [Google Scholar] [CrossRef]
- Purić, J.; Vieira, G.; Cavalca, L.B.; Sette, L.D.; Ferreira, H.; Vieira, M.L.C.; Sass, D.C. Activity of Antarctic fungi extracts against phytopathogenic bacteria. Lett. Appl. Microbiol. 2018, 66, 530–536. [Google Scholar] [CrossRef]
- Baricz, A.; Teban, A.; Chiriac, C.M.; Szekeres, E.; Farkas, A.; Nica, M.; Dascălu, A.; Oprișan, C.; Lavin, P.; Coman, C. Investigating the potential use of an Antarctic variant of Janthinobacterium lividum for tackling antimicrobial resistance in a One Health approach. Sci. Rep. 2018, 8, 15272. [Google Scholar] [CrossRef]
- Corte, A.M. Antibacterial activity of Penicillium spp. strains isolated in extreme environments. Polar Biol. 2000, 23, 294–297. [Google Scholar] [CrossRef]
- Giudice, A.L.; Fani, R. Antimicrobial potential of cold-adapted bacteria and fungi from Polar Regions. In Biotechnology of Extremophiles: Advances and Challenges; Rampelotto, P.H., Ed.; Springer International Publishing: Berlin/Heidelberg, Germany, 2016; pp. 83–115. [Google Scholar] [CrossRef]
- Krishnan, A.; Alias, S.A.; Wong, C.M.; Pang, K.L.; Convey, P. Extracellular hydrolase enzyme production by soil fungi from King George Island, Antarctica. Polar Biol. 2011, 34, 1535–1542. [Google Scholar] [CrossRef]
- Liu, J.T.; Lu, X.L.; Liu, X.Y.; Gao, Y.; Hu, B.; Jiao, B.H.; Zheng, H. Bioactive natural products from the Antarctic and Arctic organisms. Mini Rev. Med. Chem. 2013, 13, 617–626. [Google Scholar] [CrossRef] [PubMed]
- Abrashev, R.; Feller, G.; Kostadinova, N.; Krumova, E.; Alexieva, Z.; Gerginova, M.; Spasova, B.; Miteva-Staleva, J.; Vassilev, S.; Angelova, M. Production, purification, and characterization of a novel cold-active superoxide dismutase from the Antarctic strain Aspergillus glaucus 363. Fungal Biol. 2016, 120, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Martinez, A.; Cavello, I.; Garmendia, G.; Rufo, C.; Cavalitto, S.; Veroet, S. Yeasts from sub-Antarctic region: Biodiversity, enzymatic activities and their potential as oleaginous microorganisms. Extremophiles 2016, 20, 759–769. [Google Scholar] [CrossRef] [PubMed]
- Bratchkova, A.; Ivanova, V. Bioactive metabolites produced by microorganisms collected in Antarctica and the Arctic. Biotechnol. Biotechnol. Equip. 2011, 25, 1–7. [Google Scholar] [CrossRef]
- Kirtsideli, I.Y.; Vlasov, D.Y.; Krylenkov, V.A.; Rolle, N.N.; Barantsevich, E.P.; Sokolov, V.T. Comparative study of airborne fungi at Arctic stations near water area of the Northern Sea Route. Hum. Ecol. 2018, 4, 16–21. (In Russian) [Google Scholar] [CrossRef]
- Kellogg, C.A.; Griffin, D.W. Aerobiology and the global transport of desert dust. Trends Ecol. Evol. 2006, 21, 638–644. [Google Scholar] [CrossRef]
- Barrie, L.A. Arctic air pollution: An overview of current knowledge. Atmos. Environ. 1986, 20, 643–663. [Google Scholar] [CrossRef]
- Brown, J.K.M.; Hovmøller, M.S. Aerial dispersal of pathogens on the global and continental scales and its impact on plant disease. Science 2002, 297, 537–541. [Google Scholar] [CrossRef]
- Garcıa-Pando, C.P.; Stanton, M.C.; Diggle, P.J.; Trzaska, S.; Miller, R.L.; Perlwitz, J.P.; Baldasano, J.M.; Cuevas, E.; Ceccato, P.; Yaka, P.; et al. Soil dust aerosols and wind as predictors of seasonal meningitis incidence in Niger. Environ. Health Perspect. 2014, 122, 679–686. [Google Scholar] [CrossRef]
- Kollath, D.R.; Miller, K.J.; Barker, B.M. The mysterious desert dwellers: Coccidioides immitis and Coccidioides posadasii, causative fungal agents of coccidioidomycosis. Virulence 2019, 10, 222–233. [Google Scholar] [CrossRef] [PubMed]
- Fahlgren, C.; Hagström, Å.; Nilsson, D.; Zweifel, U.L. Annual variations in the diver-sity, viability, and origin of airborne bacteria. Appl. Environ. Microbiol. 2010, 76, 3015–3025. [Google Scholar] [CrossRef] [PubMed]
- Pearce, D.A.; Bridge, P.D.; Hughes, K.A.; Sattler, B.; Psenner, R.; Russell, N.J. Microorganisms in the atmosphere over Antarctica. FEMS Microbiol. Ecol. 2009, 69, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Gritsenko, L.Z.; Kolokolova, Y.V.; Kolesnikova, A.G.; Mishin, V.V.; Ananyeva, M.N. Role of acinetobacter in the occurrence of problematic infections. Med. Soc. Probl. Fam. 2014, 19, 122–127. (In Russian) [Google Scholar]
- Moreira, J.S.; Riccetto, A.G.L.; da Silva, M.T.N.; dos Santos Vilela, M.M. Endocarditis by Kocuria rosea in an immunocompetent child. Braz. J. Infect. Dis. 2015, 19, 82–84. [Google Scholar] [CrossRef] [PubMed]
- Chou, Y.-J.; Chou, J.-H.; Lin, K.-Y.; Lin, M.-C.; Wei, Y.-H.; Arun, A.B.; Young, C.-C.; Chen, W.-M. Rothia terrae sp. nov. isolated from soil in Taiwan. Int. J. Syst. Evol. Microbiol. 2008, 58, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Arda, B.; Aydemir, S.; Yamazhan, T.; Hassan, A.; Tunger, A.; Serter, D. Comamonas testosteroni meningitis in a patient with recurrent cholesteatoma. APMIS 2003, 111, 474–476. [Google Scholar] [CrossRef]
- Lee, M.R.; Huang, Y.T.; Liao, C.H.; Chuang, T.Y.; Lin, C.K.; Lee, S.W.; Lai, C.C.; Yu, C.J.; Hsueh, P.R. Bacteremia caused by Brevundimonas species at a tertiary care hospital in Taiwan, 2000–2010. Eur. J. Clin. Microbiol. Infect. Dis. 2011, 30, 1185–1191. [Google Scholar] [CrossRef]
- Chen, Y.-G.; Zhang, Y.-Q.; Shi, J.-X.; Xiao, H.-D.; Tang, S.-K.; Liu, Z.-X.; Huang, K.; Cui, X.-L.; Li, W.-J. Jeotgalicoccus marinus sp. nov., a marine bacterium isolated from a sea urchin. Int. J. Syst. Evol. Microbiol. 2009, 59, 1625–1629. [Google Scholar] [CrossRef]
- Ghannoum, M.A. Potential role of phospholipases in virulence and fungal pathogenesis. Clin. Microbiol. Rev. 2000, 13, 122–143. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).