1. Introduction
Soil erosion is a serious issue which endangers sustainable land use strategies, especially in arid and semiarid regions [
1,
2,
3]. Water and wind as erosive factors degrade the land, their importance varies depending on the region [
4,
5]. In Morocco, water is often the crucial erosive force [
5]. Most of the time only a few strong events are responsible for the majority of soil losses [
6,
7]. Due to the small amount of precipitation coupled with its great variability, rangeland is more common in these arid regions than farmland. Therefore, agrosilvopastoral systems are the most typical land uses. These forms of land use often become established in the dryland forests which occur naturally in these areas [
3,
7]. Forest degradation as a consequence of erosion and mismanagement is a common phenomenon in these dryland forests. It has been recorded all over the world (e.g., in Spanish dehesas [
8], Algerian oak forests [
9], Moroccan argan woodlands [
3], and Mongolian forest steppes [
10]). While the degradation dynamics are well understood in some regions, there is a lack of knowledge elsewhere. For example, in the Spanish dehesas, there are studies regarding the interrelations between rainfall intensity, vegetation cover, grazing and soil loss [
6], the influence of vegetation on moisture conditions in the soil [
7] and the connection between soil conditions, vegetation cover, and pasture production [
2]. Furthermore, the influence of patchy vegetation on soil conditions beneath is highlighted around the world [
11,
12,
13,
14,
15,
16]. Typically, there is an enrichment of organic matter and nutrients under plants. Therefore the term “fertile islands” is often used for patchy vegetation in dry regions [
14,
15]. It is likely that these “fertile islands” are better protected against soil loss, because the input of organic matter in the soil is crucial for its resistance against erosion [
17,
18,
19,
20].
Corresponding studies for the Moroccan argan woodlands investigating the interaction between soil, vegetation, and erosion processes are missing, although protection of argan trees and land-use strategies around them have been stipulated by UNESCO since 2014. Reasons for this are the complex socioeconomic structures behind the argan economy and argan oil which is in growing demand around the world [
21]. A massive decline of argan tree density from 27 to 15 trees ha
−1 between 1970 and 2007 shows that protection of these dryland forests is necessary to preserve them [
3].
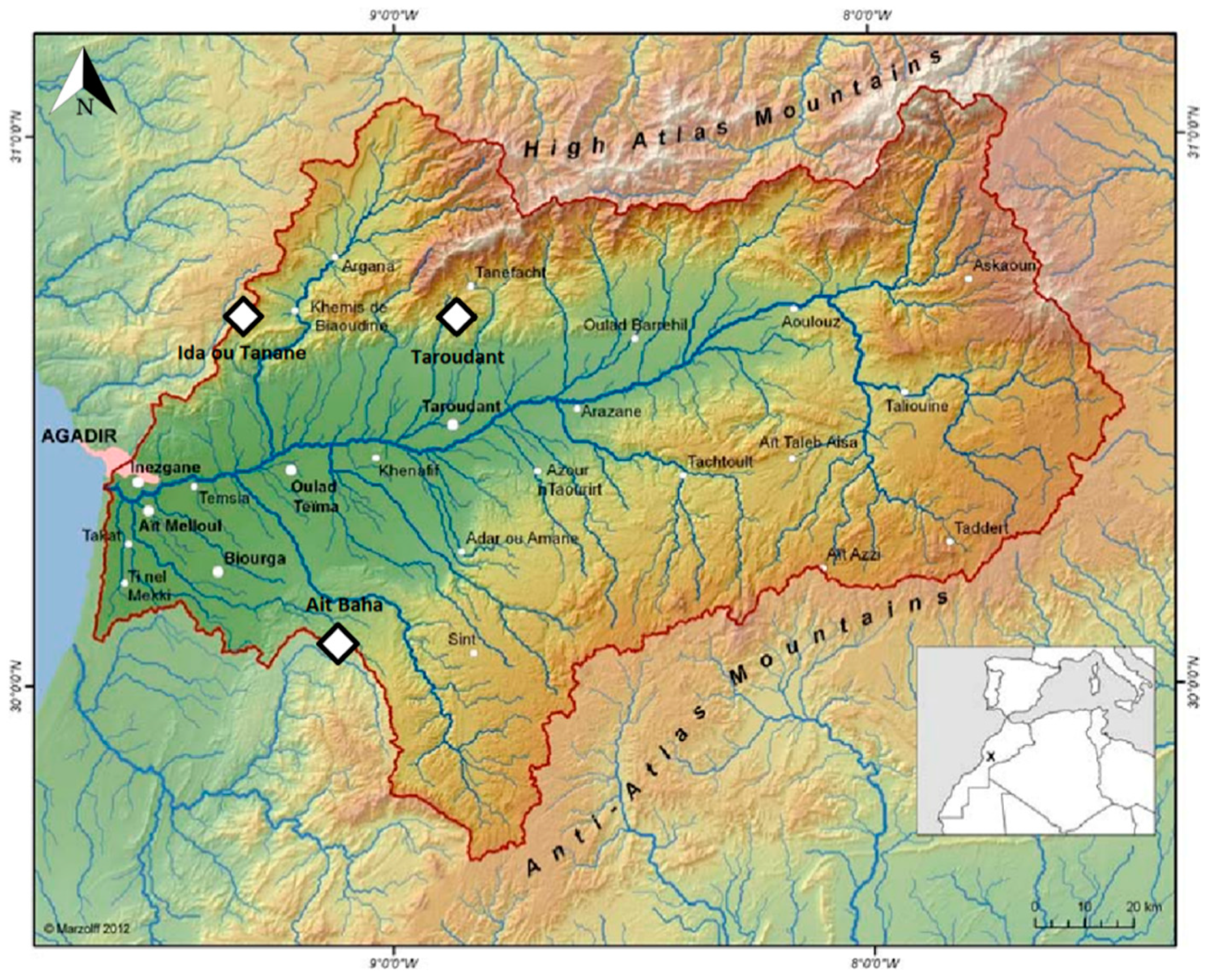
Argania spinosa is endemic in southern Morocco and has a small dispersal area of ~950,000 ha [
3] which is mostly limited to the Souss basin [
22]. Although
Argania spinosa is well adapted to the dry conditions of Morocco’s southwest, the population is endangered because it is the only source of forage in dry periods [
3,
22]. In consequence, there is a high pressure of use by the local population and their goat herds, as well as by nomadic tribes and their goat and dromedary herds [
23]. In addition, the fruits of
Argania spinosa are collected to produce argan oil, which is sold for cosmetic and culinary usage [
3]. In other areas, access for animals is not permitted and speculative rainfed agriculture is practiced [
23]. Especially overgrazing and deforestation lead to varying tree-crown sizes and growth forms, smaller numbers of trees per hectare, and a lower proportion of covered soil [
23,
24] (
Figure 1). As a result, there is less protection against surface runoff, desiccation is smaller, soils become more degraded, young sprouts cannot grow, and thus, the vegetation cover further declines.
While various studies have investigated the degradation of argan trees [
3,
22,
25,
26], there is a lack of knowledge regarding the connection between vegetation degradation and soil degradation as well as soil loss. Other studies have already shown that there are strong erosive processes in the Souss valley [
27]. Usage on the sites is not only limited to the area between the trees. The soil is often cultivated between the trees and under the tree crowns. Herds of goats, sheep, and dromedaries graze and browse the tree area most of all, thus possibly compacting the soil beneath the crown as was demonstrated by Mulholland and Fullen on loamy sands and Stavi et al. [
28,
29]. It is unknown if the soils beneath argan trees are already so degraded that, should the tree be removed, young sprouts would not be able to grow. If this is already the case in the intertree areas, a regeneration even by afforestation is unlikely to be successful. The main aim of this study is to analyze the interaction between argan trees and multiple soil parameters, such as C-content and N-content, as indicators for erosion stability. Our main hypothesis is that the soils in the tree area are more protected from rain, splash erosion, and surface wash and thus show less degraded soils than the unprotected intertree areas. Consequently, the securing of adequate tree density would be an important step towards sustainable land management in southern Morocco. Yet, the soils beneath the trees have been the subject of potentially degrading factors. If a difference between the soil parameters exists, the extent to which they differ is an important indicator for the degradation of soils in the intertree and tree areas. To test this hypothesis, measurements of erodibility and soil characteristics were taken under trees and in corresponding intertree areas at 30 test sites of 1 ha each and statistically analyzed.
4. Discussion
A comparison between tree and intertree areas in argan woodlands was carried out using rainfall simulations, infiltration measurements, as well as soil analyses. Since this study did not focus on specific tree degradation stages but tried to encompass a multitude of different land uses as well as tree and soil degradation stages, the range of values is not surprising. Disturbance of vegetation, such as loss of herbaceous cover, can lead to an increase in sediment transport and erosion. This is especially true when there is a decrease in woody plant canopy cover from woodlands to shrublands [
68], which is partly the case in the argan woodlands with different stages of degradation and a change in canopy cover due to overgrazing [
69,
70]. The argan woodlands have been degraded because of grazing pressure, rainfed agriculture in silvo-agricultural land uses, as well as fuelwood extraction [
3,
71], leading to a wider and more disturbed intertree area with less and wider spaced trees [
3].
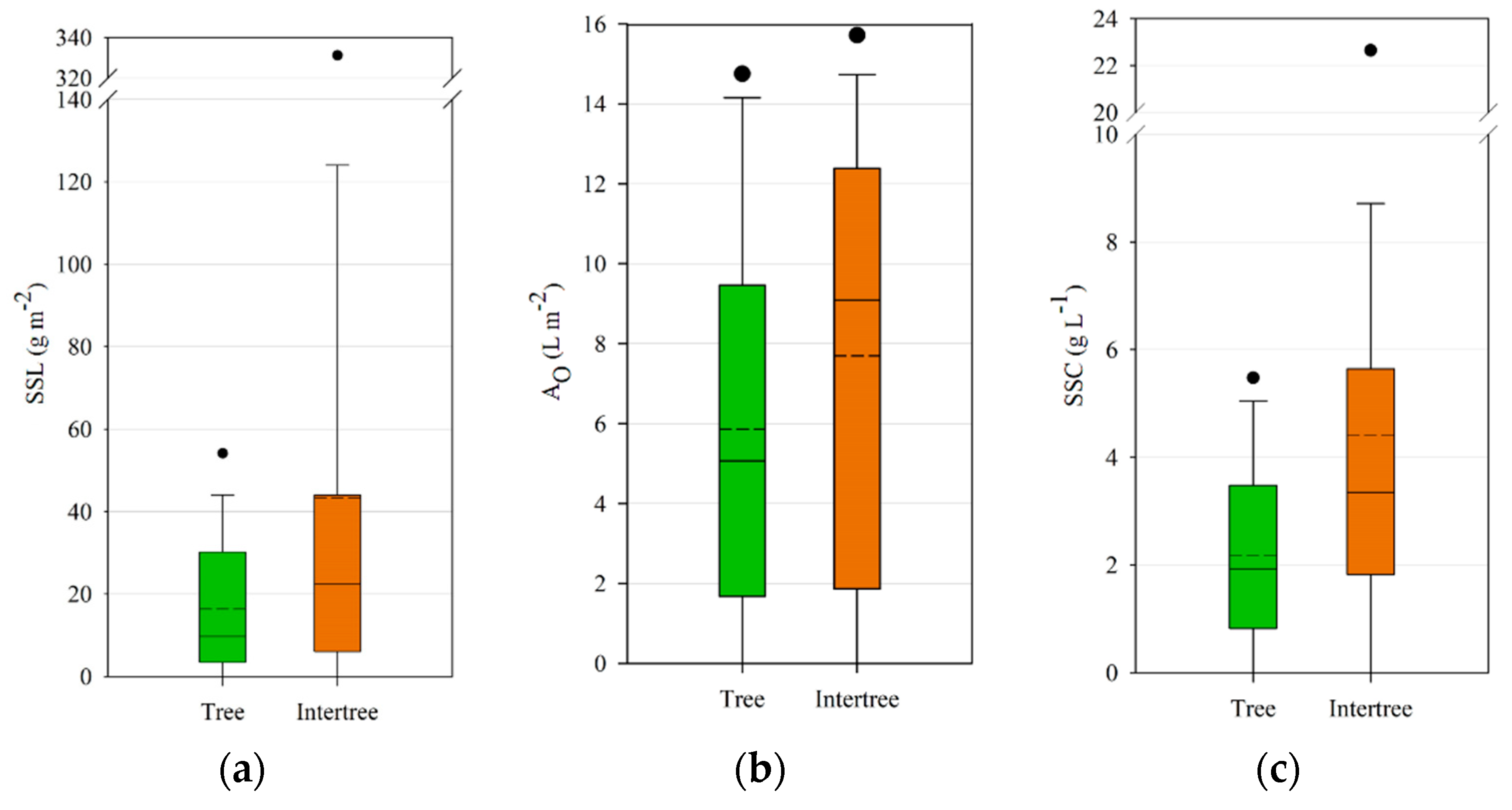
Although significant differences in the
t-tests could not be found, most of the test sites showed lower SSC in the tree areas compared to the neighboring intertree areas. Other studies showed higher runoff and erosion rates on bare intercanopy patches compared to canopy-covered areas [
72,
73], whereas vegetated intercanopy patches showed medium rates of runoff and erosion [
72]. Ceballos et al. obtained differing results, where vegetated grassy areas showed little runoff and erosion rates, yet tree-covered areas showed much higher rates due to soil hydrophobicity hindering infiltration [
74]. Since rainfall simulations in our study were carried out the same way under trees as in the intertree areas, the rainfall intensity was much higher than under natural conditions, when rainfall may be reduced by 6.6% to 82.7% due to interception [
75]. Nevertheless, splash erosion by throughfall drops may be 2.59 times higher than on open fields [
76], so the rainfall simulations under the trees should not be equated with natural conditions. It is possible, however, to compare the erodibility [
77,
78] between tree and intertree areas. The cluster analyses in
Table 3 and
Table 6 show the tendency of clusters being attributed to tree or intertree areas. Intertree areas with lower SSC, SSL, and A
O could all be assigned to active agricultural use with better infiltration potential due to ploughing. The three tree areas belonging to cluster 2 showed degraded tree areas with little to no vegetation cover or organic litter.
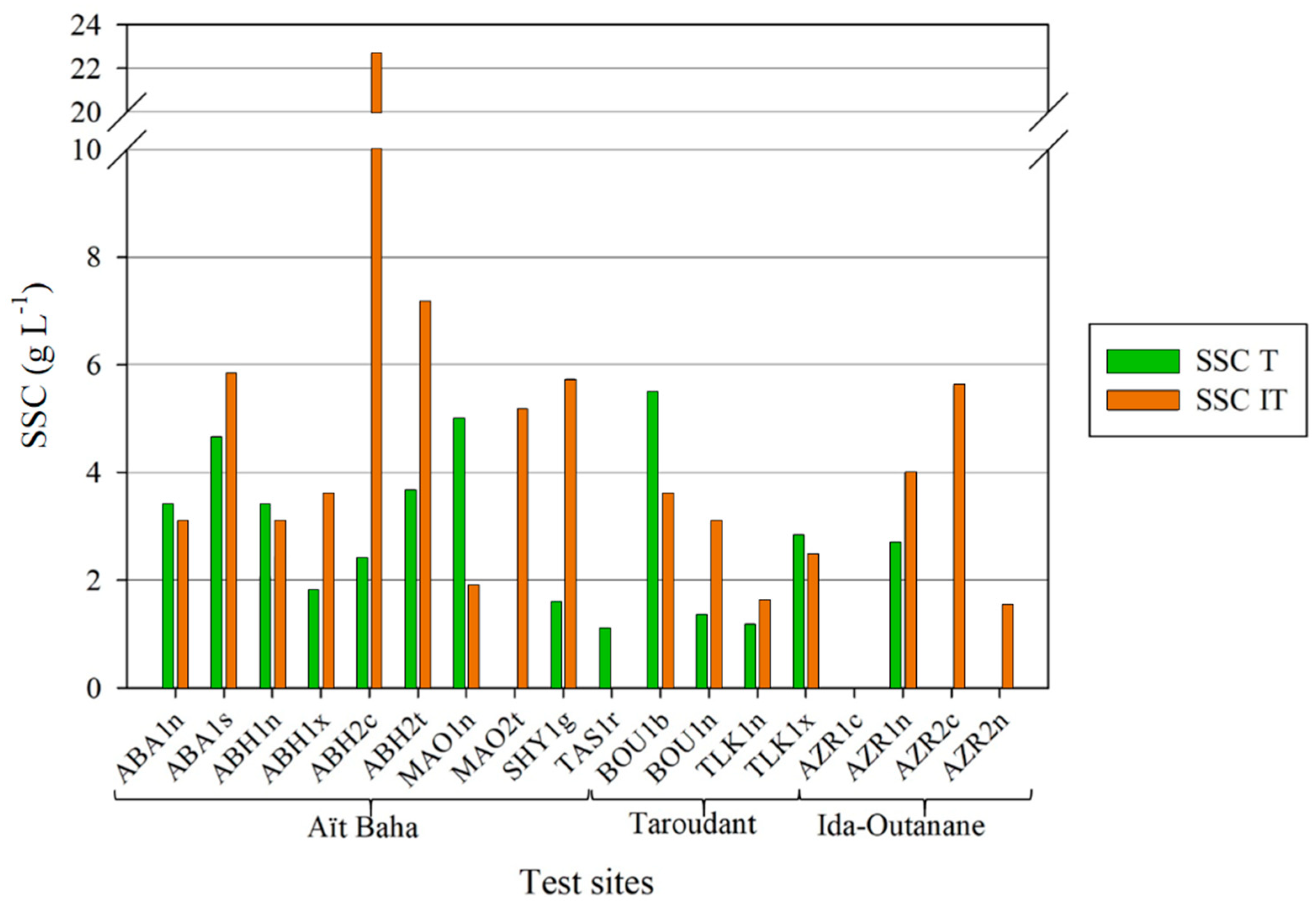
Other studies in Morocco, also in the study area of Taroudant, found mean suspended sediment concentrations on fallow land and orange plantations of 2.7 g L
−1 [
37] compared to lower tree area and higher intertree area means of 2.18 g L
−1 and 4.42 g L
−1, respectively. Ceballos et al. also showed suspended sediment concentrations for tree-covered areas in the Spanish dehesas which were lower both under dry (mean 0.37 g L
−1) and wet conditions (mean 0.65 g L
−1) than either tree or intertree areas. Even sheep trails showed lower average SSC (dry: 0.45 g L
−1, wet: 0.76 g L
−1) [
74]. This indicates the higher erodibility of the soils in the Moroccan study areas, especially in the intertree areas that are not protected by vegetation or leaf litter. This extra layer has been found to strongly reduce runoff and sediment loss [
79]; it is missing in some test sites such as MAO1n, where higher SSC was measured in the tree area (see
Figure 7). Since some test sites are more disturbed and show little to no vegetation in the intertree and tree areas (undergrowth), there are cases where SSC can also be higher in tree areas. Ludwig et al. [
80] showed higher runoffs and sediment yields on disturbed areas as compared to undisturbed or only slightly grazed areas. Wind erosion has not yet been studied in these regions, but due to a high crust cover and little loose sediment on the surface, it is assumed to be secondary.
Soil erodibility is, of course, also dependent on aggregate stability. The results differ significantly between the tree and intertree areas. This could also be due to the higher carbon contents in the tree areas, which can lead to the formation of larger aggregates [
81,
82]. Under agricultural use organic matter content and aggregate stability can decline [
83]. This was the case on all test sites that were under agricultural use in the intertree areas, which partly explains the higher SSC on the intertree areas. In some cases, there was nearly no surface runoff until the end of the experiment, which was partly due to infiltration into the loosened soil or enhanced surface roughness by ploughing. As Mbagwu and Auerswald noted, high interrill erosion rates should be expected on soils with a PS <250 mL 10 min
−1 [
64]. The measured PS was mostly below this value in tree or intertree areas. This could explain the much higher SSC compared to the values of Ceballos et al. [
74]. In comparison to the values published by Mbagwu and Auerswald the measured PS ranked very low in many of the studied test sites, with a mean of 43.23 mL 10 min
−1 in the intertree area, which is just slightly higher than the minimum value published by the aforementioned authors. Their values for fallows, pastures or (secondary) forests (>250 mL 10 min
−1) cannot be repeated in our measurements for most intertree and tree areas. Intertree areas are partly cultivated or unmulched plots which should show medium (250–150 mL 10 min
−1) to low PS (<150 mL 10 min
−1). They could also be compared to secondary forest subsoil with low PS (<150 mL 10 min
−1), since topsoil could have been eroded, thus explaining the low percolation stabilities [
64]. Goebel et al. [
84] obtained similar results regarding higher aggregate stability in topsoil, and also found higher aggregate stabilities in forest soils than arable soils, which matches the results with higher aggregate stabilities under trees compared to the crusted and mostly eroded intertree areas. Since aggregate breakdown leads to crusting of soil, the infiltration rate decreases with lower aggregate stability and runoff, and sediment detachment occurs [
85,
86].
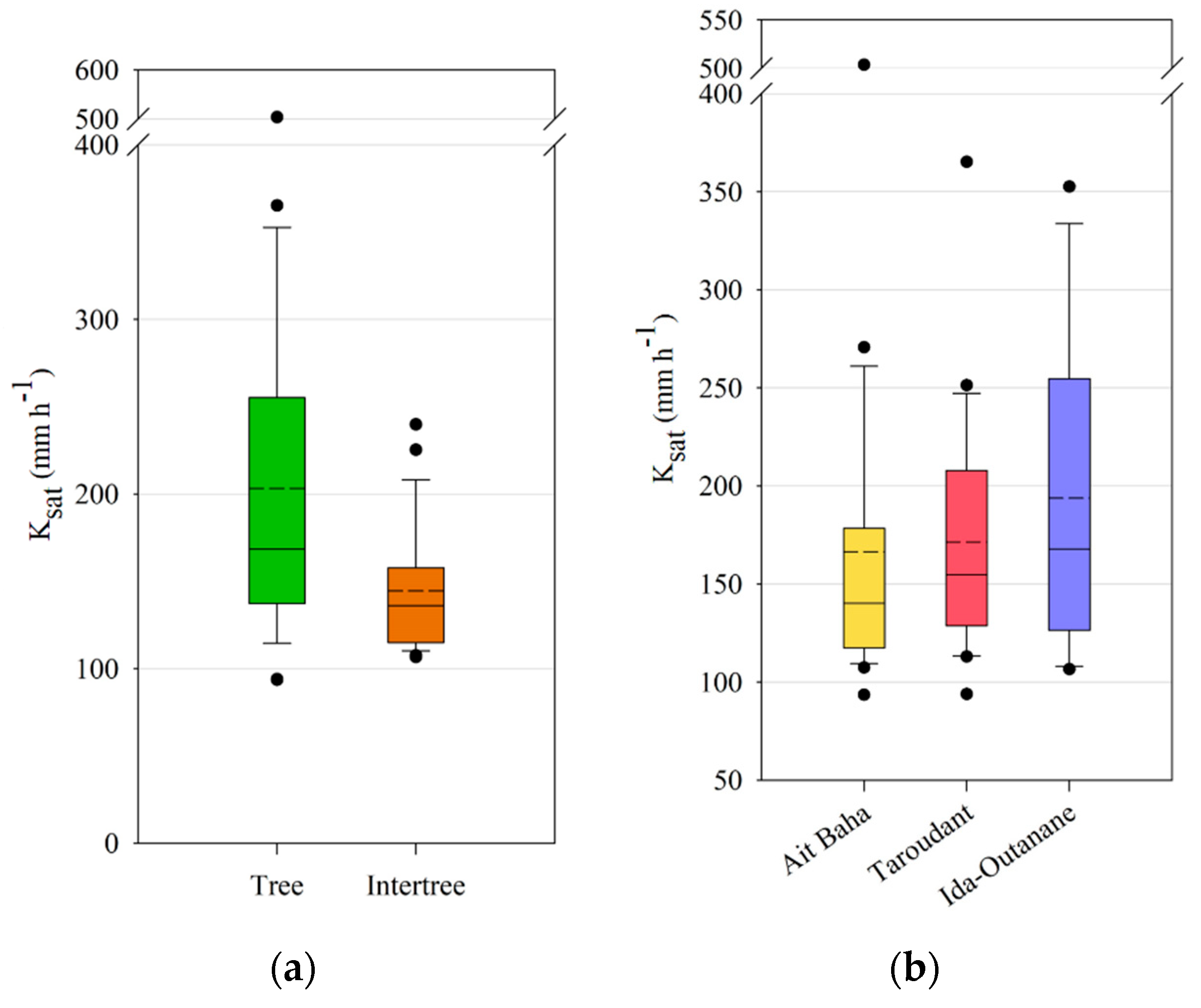
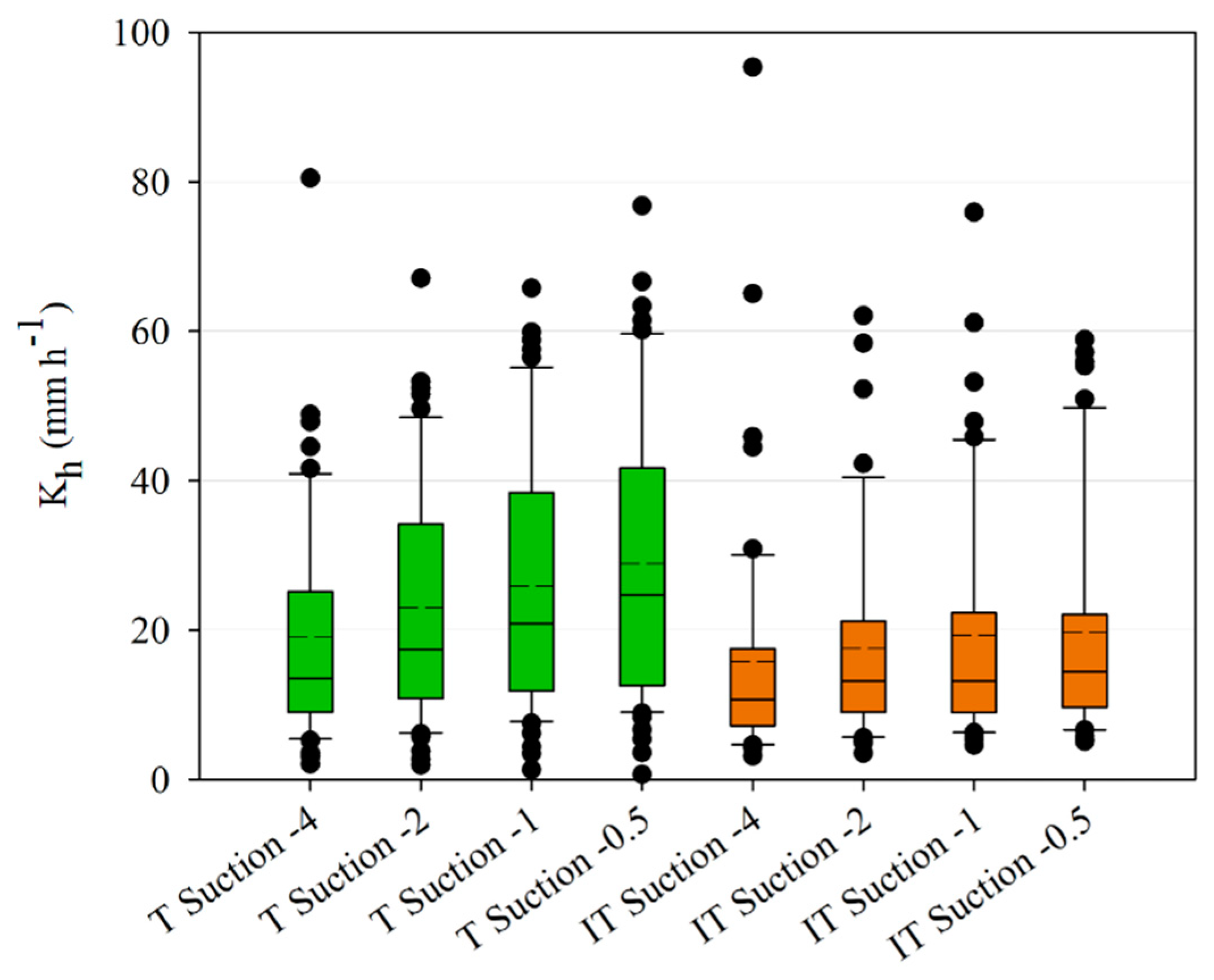
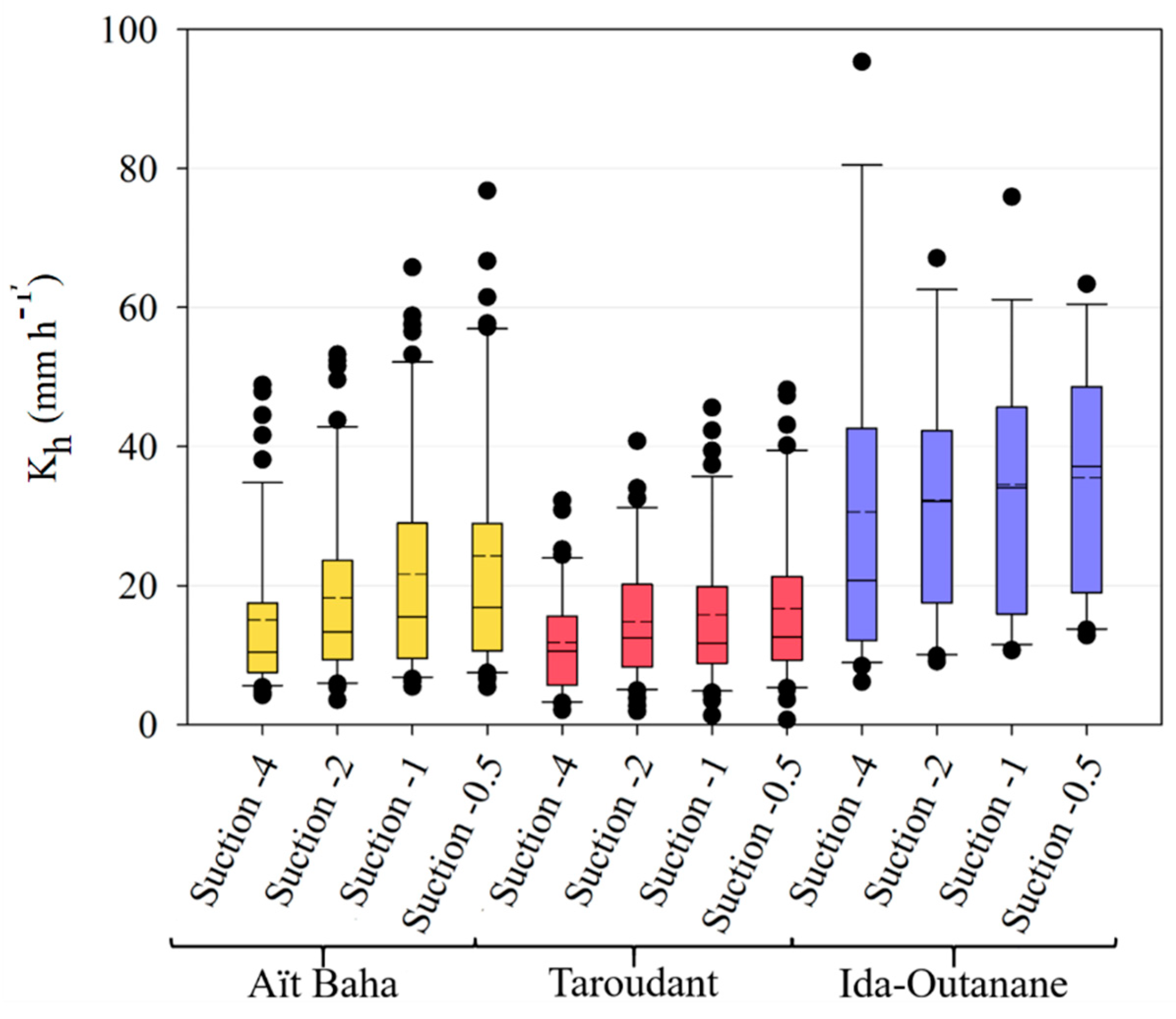
The infiltration measurements have shown a significant difference between tree and intertree areas for both unsaturated and saturated hydraulic conductivity, as well as infiltration rates, indicating a better infiltrability into the soil in vegetation patches [
80,
87]. Lichner et al. [
88] noted high K
sat values in forest soils due to the greater presence of macropores, yet found the lowest K
h values for forest sites possibly due to a higher water repellency caused by the coating of soil particles by pine-needle waxes. A study conducted in the southern part of the study area of Taroudant found much lower infiltration rates than the values presented here with a mean of 84 mm h
−1 and a maximum of 265 mm h
−1 [
49]. This study focused on cultivated and leveled landscapes with high soil-crust cover in contrast to the less crusted tree areas or the intertree areas with a higher stone-fragment cover on the surface. The influence of stone cover on infiltration rates may be either positive or negative, depending on the measurement context [
89,
90]. If the measurements occur between large shrubs, or in this case trees, the influence is positive, yet when shrub and intershrub are compared, the correlation with stone cover becomes negative since the finer sediments under shrubs have a positive effect on infiltration [
89]. Thus, compared to the results of Peter and Ries [
49], stone cover influences the infiltration in the intertree areas positively, yet tree area infiltration is still mostly higher. Although there are high percentages of sand, mostly in the coarse fraction, in both tree and intertree areas, differences in infiltration rates and hydraulic conductivities could also be due to the plant cover which could maintain macropores on the surface and conduct water into the soil [
91], while in the intertree areas vegetation cover is rather low (10.17 ± 14.10%). A higher density of soil underneath the argan trees due to compaction by livestock was not measured in this study but would affect infiltration into the soil negatively [
92]. Since the measured infiltration values are higher in the tree area, it suggests that the soil is rather stable against compaction by livestock. The grain size distribution for corresponding tree and intertree areas does not vary much, they are mostly classified as loam, sandy loam, or sandy clay loam.
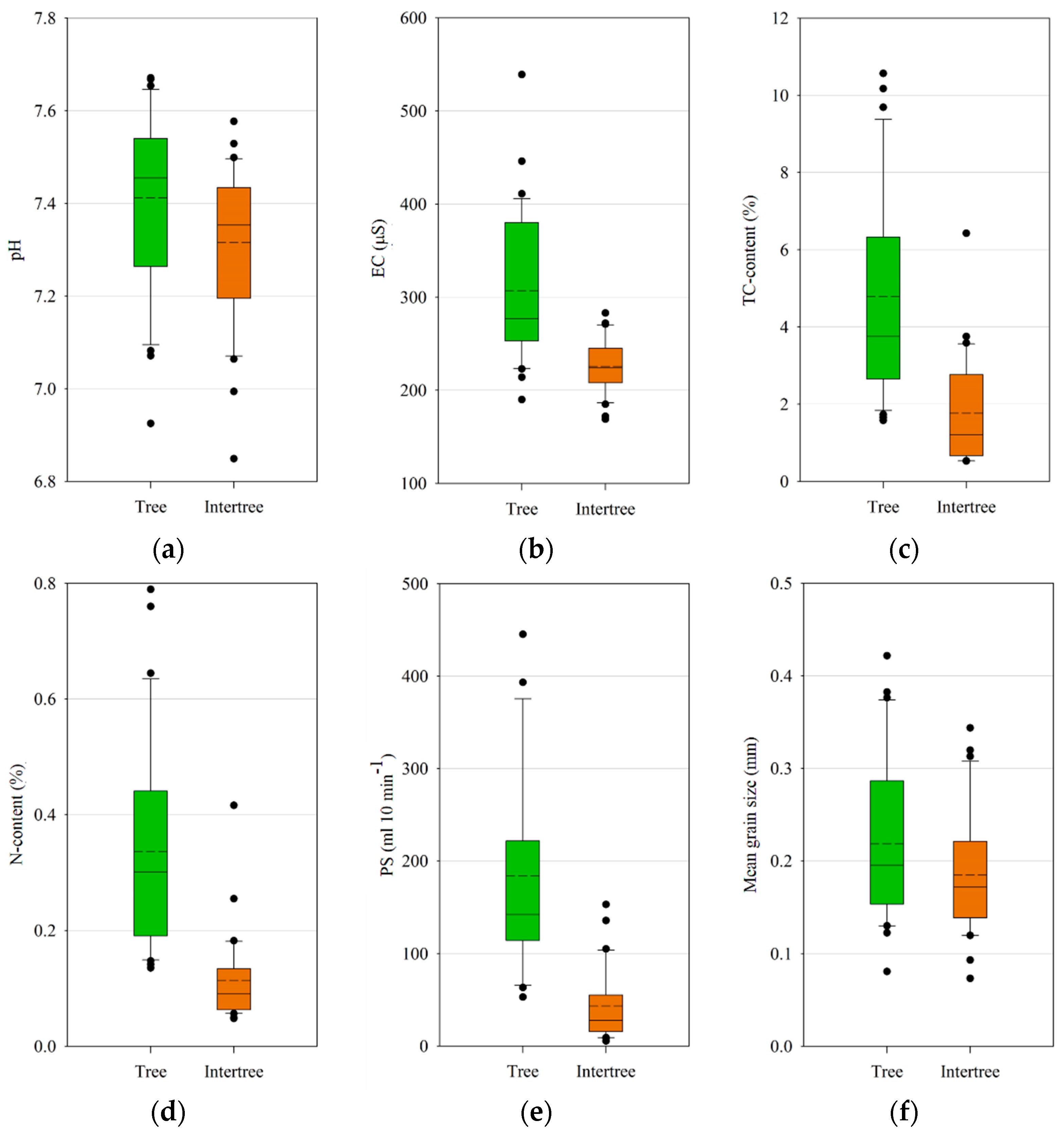
With a higher vegetation cover and more leaf litter in the tree areas in most cases, the difference in the content of total carbon between tree and intertree areas can be explained. The same is true for the content of total nitrogen, which could also be influenced by the droppings of goats and sheep. During dry years the goats and sheep feed from the trees, which are the only biomass storage in these areas, and are shepherded from one tree to another. Thus, they spend most time in the tree area, possibly influencing the N-content in the soil. Soil microbial communities profit from the higher N-content under the plants [
15]. Micro-, meso-, and macro-fauna are drawn to canopy-covered areas, since they are the places with the lowest temperatures in the daytime and the highest soil moisture, even in these very dry climates (see
Table 2), and therefore the places where food is most abundant [
13]. The higher carbon contents under the trees originate from the litter as well as the undergrowth. Tree or shrub litter has a positive influence on soil carbon as well as on aggregate stability, shear strength, as well as infiltration rates, which in turn leads to greater resistance against runoff and erosion [
14,
93]. The removal or absence of litter can therefore have a negative influence on the studied soil parameters.
The cluster analysis classifying soil parameters shows that an attribution of tree and intertree areas to specific clusters is in many cases possible. Since total carbon content is the most influential predictor, areas with low total carbon content are often classified into the cluster associated with lower values. On the other hand, agriculturally used intertree areas where organic litter was ploughed into the soil can be classed into the cluster with higher values. Yet, the majority of tree areas is classed into the cluster with generally higher values, while the majority of intertree sites can be associated with the cluster with lower values.
Many studies focus on shrubs and intershrub differences [
14,
15,
94] where the shrub covers the soil well and there is no great distance between the soil surface and the leaf cover of the shrub. It can thus act as an obstacle for runoff and transported sediment. The distance between the canopy of trees and the soil surface is much higher though, resulting in splash erosion due to throughfall and interception, as noted. The influence of leaf litter can prevent soil erosion, yet without litter the canopy-covered area is unprotected against splash erosion as well as from runoff from further upslope, leading to a degradation of the soil in the tree area. Here, different degradation stages of the trees could show a high influence on the results. This depends on the tree architecture. Round, healthy-looking crowns (see
Figure 1c) produce a lot of leaf litter, yet the thickness of the litter layer also depends on the height of the tree. The litter from higher trees might be scattered more widely by the wind and therefore not as concentrated as litter from lower trees. Rather low trees can lead to a greater number of sheep (and goats) that could erode the soil further with their hooves [
95]. Since sheep do not climb the trees as goats do, they browse only the trees they can reach from the ground. With more open crowns throughfall is higher, while litter concentration is lower. In the case of the argan as a cushion-type shrub with a very dense network of branches and twigs (
Figure 1e), the most degraded ‘tree’, the distance between canopy and soil surface is lowest, thus the litter and soil underneath are most protected, although only in a very small area.
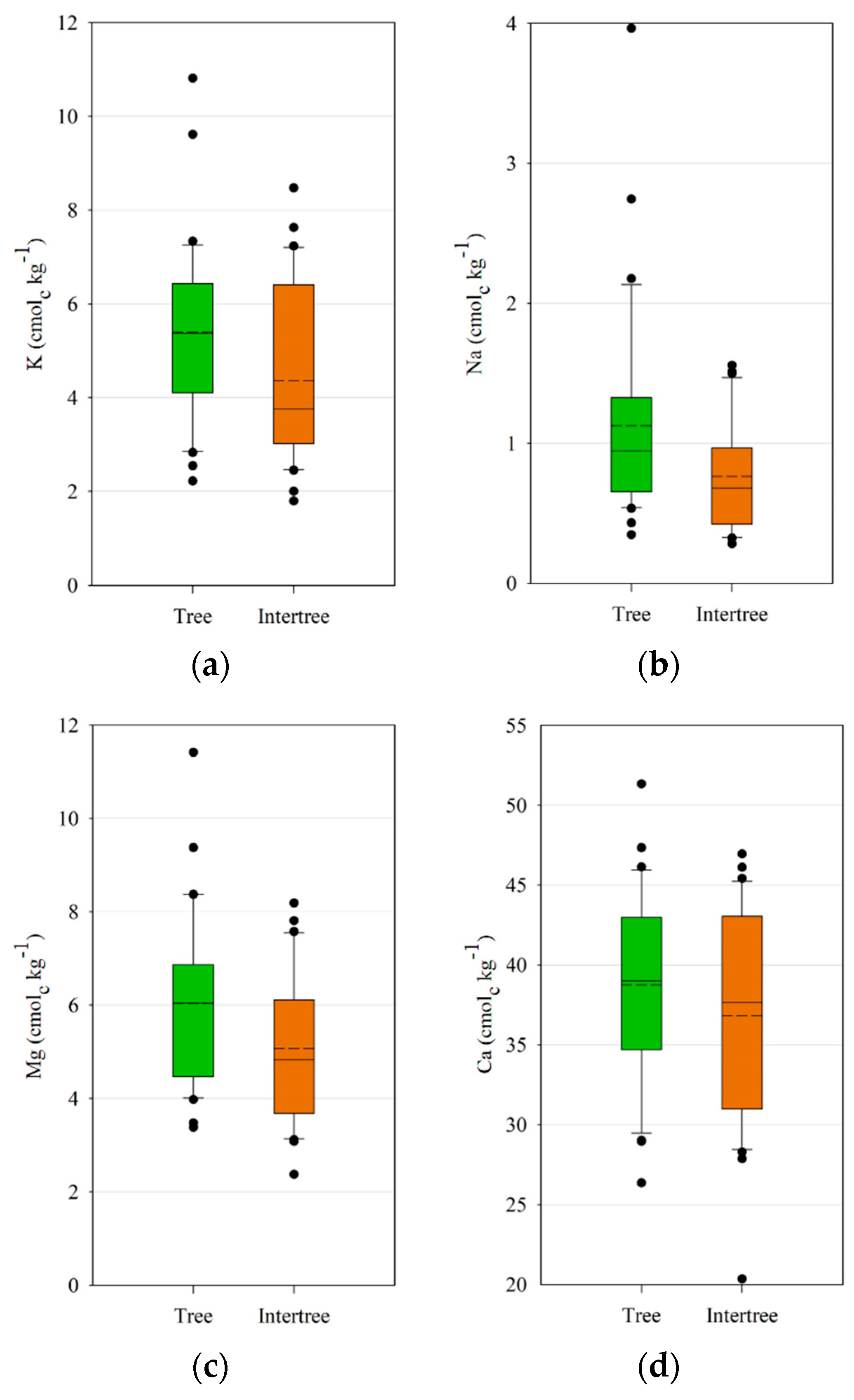
Significant differences between tree and intertree areas were also observed for the cation concentrations of K, Mg, and Na. Berthrong et al. [
96] noted decreases of Ca, Mg, and K as well as pH and an increase of Na in response to afforestation. For argan afforestation projects in Morocco this potential decrease should be kept in mind. Organic matter also increases the cation exchange capacity and thus the available cations [
14].
However, another possible explanation for the differences could be due to coastal fog which is caught by the argan trees [
97] and precipitates in the tree area. It is a major source of precipitation for the argan trees, since rainfall is very variable in the study areas [
22]. Although fog has lower concentration of nutrients than rain, as shown by Azevedo and Morgan [
98], it could still lead to a higher accumulation of cations in the tree area, since precipitation by rainfall influences both tree and intertree areas, but fog predominantly precipitates in the tree area. The occult deposition rates of nutrients by fog decrease with the distance from the ocean [
99]. This is shown in the results. Ida-Outanane is the closest study area to the sea and has the highest cation concentrations for K, Ca, and Na. Although the study areas of Taroudant and Aït Baha are situated further from the ocean, fog can occur on the foothills of the High Atlas and Anti-Atlas and be collected by argan plants, as shown in
Figure 13; this can therefore lead to a higher concentration of the cations K, Ca, and Na in the tree areas.
The differences in the pH between tree and intertree areas are potentially caused by the source of nitrogen in the soil. If a lot of nitrate (NO
3−) is available, the plants will absorb more anions and thus the pH rises. If cations are absorbed the pH decreases in the vicinity of the roots [
100,
101].
Inter-plant soil properties such as pH, EC, and C:N ratio are all influenced by the adjacent vegetation [
15]. This shows the importance of a closer distance between trees rather than an expanding intertree area. Since the distances between the studied trees and their intertree areas are all different depending on the tree density of each test site, variabilities in the values could stem from these differences. The reintroduction of specific tree species into the degraded intertree areas could lead to an increase in carbon and nutrients [
102]. Afforestation projects are in process, yet only with limited success, since young argan sprouts need to be protected from grazing animals. This is mostly done by fenced exclusion areas, although the exclusion is in many cases not successful. Break-ins are punishable, but in dry years there is no other food source for the many herds. In addition, young sprouts need a lot of water for several years to develop their roots and be able to reach water deeper in the soil. Dry years often lead to withering of the sprouts, since planting companies are often only engaged for two to three years, which does not give the young sprouts enough time to grow.


 ,
, 













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}