Aquatic Insects and Benthic Diatoms: A History of Biotic Relationships in Freshwater Ecosystems

 , , , and
, , , and
Abstract
:1. Introduction
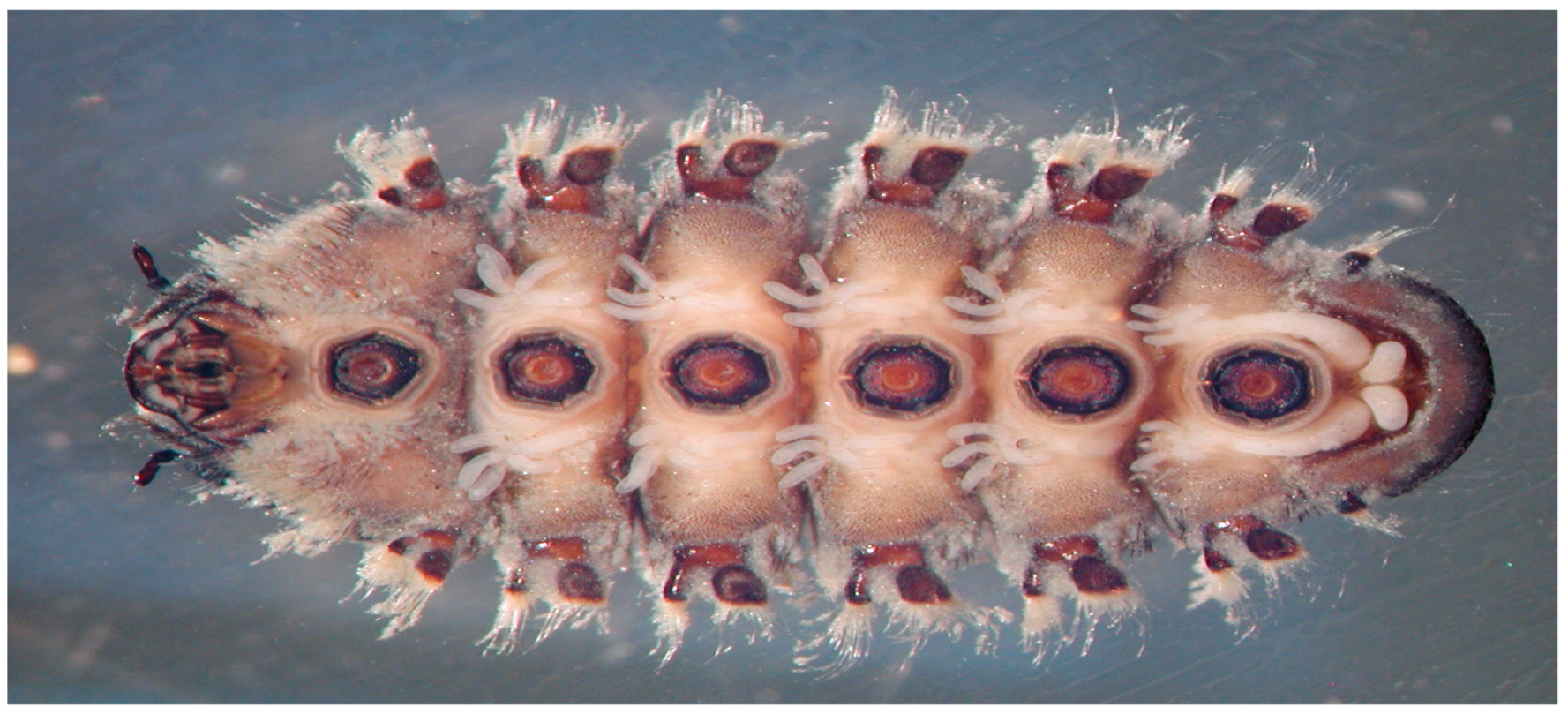
2. Ability to Withstand Elevated Velocity Current Environments
3. Ability to Scrape Algae from the Substrate
4. Adaptation to Live in The Open
5. Direct and Indirect Effects of Scrapers in Shaping Diatom Communities
6. Epizoic Relationships
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Minshall, G.W. Aquatic insect-substratum relationships. In The Ecology of Aquatic Insects; Resh, V.H., Rosenberg, D.M., Eds.; Praeger Publishers: New York, NY, USA, 1984; pp. 358–400. [Google Scholar]
- Poff, N.L.; Allan, J.D.; Bain, M.B.; Karr, J.R.; Prestegaard, K.L.; Richter, B.D.; Sparks, R.E.; Stromberg, J.C. The natural flow regime. BioScience 1997, 47, 769–784. [Google Scholar] [CrossRef]
- Bellinger, E.G.; Sigee, D.C. Freshwater Algae: Identification, Enumeration and Use as Bioindicators; John Wiley Sons: London, UK, 2015; p. 290. [Google Scholar]
- Chepurnov, V.A.; Mann, D.G.; Sabbe., K.; Vyverman, W. Experimental studies on sexual reproduction in diatoms. Int. Rev. Cytol. 2004, 237, 91–154. [Google Scholar] [PubMed]
- Torres-Ruiz M., J.D.; Wehr, J.D.; Perrone, A.A. Trophic relations in a stream food web: Importance of fatty acids for macroinvertebrate consumers. J. North. Am. Benthol. Soc. 2007, 26, 509–522. [Google Scholar] [CrossRef]
- Guo, F.; Kainz, M.J.; Sheldon, F.; Bunn, S.E. The importance of high-quality algal food sources in stream food webs–current status and future perspectives. Freshw. Biol. 2016, 61, 815–831. [Google Scholar] [CrossRef]
- Guo, F.; Bunn, S.E.; Brett, M.T.; Fry, B.; Hager, H.; Ouyang, X.; Kainz, M.J. Feeding strategies for the acquisition of high-quality food sources in stream macroinvertebrates: Collecting, integrating, and mixed feeding. Limnol. Oceanogr. 2018, 63, 1964–1978. [Google Scholar] [CrossRef]
- Groendahl, S.; Fink, P. High dietary quality of non-toxic cyanobacteria for a benthic grazer and its implications for the control of cyanobacterial biofilms. BMC Ecol. 2017, 17, 20. [Google Scholar] [CrossRef]
- Merritt, R.W.; Wallace, J.B. Aquatic habitats. In Encyclopedia of Insects; Resh, V.H., Cardi, R.T., Eds.; Academic Press: Burlington, MA, USA, 2009; pp. 38–48. [Google Scholar]
- Balian, E.V.; Segers, H.; Martens, K.; Lévéque, C. The freshwater animal diversity assessment: An overview of the results. Hydrobiologia 2008, 595, 627–637. [Google Scholar] [CrossRef]
- Resh, V.H.; Rosenberg, D.M. The Ecology of Aquatic Insects; Praeger Publisher: New York, NY, USA, 1984; p. 625. [Google Scholar]
- Cummins, K.W.; Merritt, R.W. Ecology and distribution of aquatic insects. In An Introduction to the Aquatic Insects of North. America; Merritt, R.W., Cummins, K.W., Eds.; Kendall Hunt Publishing Company: Dubuque, IA, USA, 1996; pp. 74–86. [Google Scholar]
- Cummins, K.W. Trophic relations of aquatic insects. Annu. Rev. Entomol. 1973, 18, 183–206. [Google Scholar] [CrossRef]
- Hauer, F.R.; Lamberti, G.A. Methods in Stream Ecology; Academic Press: New York, NY, USA, 2011; p. 506. [Google Scholar]
- Fenoglio, S.; Merritt., R.W.; Cummins, K.W. Why do no specialized necrophagous species exist among aquatic insects? Freshw. Sci. 2014, 33, 711–715. [Google Scholar] [CrossRef]
- Pritchard, G.; McKee, M.H.; Pike, E.M.; Scrimgeour, G.J.; Zloty, J. Did the first insects live in water or in air? Biol. J. Linn. Soc. 1993, 49, 31–44. [Google Scholar] [CrossRef]
- Wichard, W.; Arens, W.; Eisenbeis, G. Biological Atlas of Aquatic Insects; Apollo Books: Aamosen, Denmark, 2002; p. 339. [Google Scholar]
- López-Rodríguez, M.J.; Tierno de Figueroa, J.M.; Fenoglio, S.; Bo, T.; Alba-Tercedor, J. Life strategies of 3 Perlodidae species (Plecoptera) in a Mediterranean seasonal stream in southern Europe. J. North. Am. Benthol. Soc. 2009, 28, 611–625. [Google Scholar] [CrossRef]
- Wooton, R.J. The historical ecology of aquatic insects: An overview. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1988, 62, 477–492. [Google Scholar] [CrossRef]
- Lancaster, J.; Downes, B.J. Aquatic Entomology; Oxford University Press: Oxford, UK, 2013; p. 296. [Google Scholar]
- Allan, J.D.; Castillo, M.M. Stream Ecology: Structure and Function of Running Waters, 2nd ed.; Springer: Dordrecht, The Netherlands, 2007. [Google Scholar]
- Vannote, R.L.; Minshall, G.W.; Cummins, K.W.; Sedell, J.R.; Cushing, C.E. The river continuum concept. Can. J. Fish. Aquat. Sci. 1980, 37, 130–137. [Google Scholar] [CrossRef]
- Huryn, A.D. Aquatic insects—Ecology, feeding, and life history. In Encyclopedia of Inland Waters; Likens, G., Benbow, M.E., Burton, T.M., Van Donk, E., Downing, J.A., Gulati, R.D., Eds.; Academic Press: New York, NY, USA, 2009; pp. 25–50. [Google Scholar]
- Brittain, J.E.; Sartori, M. Ephemeroptera. In Encyclopedia of Insects; Resh, V.H., Cardé, R.T., Eds.; Academic Press: Burlington, MA, USA, 2009; pp. 328–333. [Google Scholar]
- Fenoglio, S.; Bo, T.; López-Rodríguez, M.J.; Tierno de Figueroa, J.M. Nymphal biology of Brachyptera risi (Morton, 1896) (Plecoptera: Taeniopterygidae) in a North Apennine stream (Italy). Entomol. Fenn. 2008, 19, 228–231. [Google Scholar] [CrossRef] [Green Version]
- Tierno de Figueroa, J.M.; López-Rodríguez, M.J. Trophic ecology of Plecoptera (Insecta): A review. Eur. Zool. J. 2019, 86, 79–102. [Google Scholar] [CrossRef] [Green Version]
- Haedicke, C.W.; Redei, D.; Kment, P. The diversity of feeding habits recorded for water boatmen (Heteroptera: Corixoidea) world-wide with implications for evaluating information on the diet of aquatic insects. Eur. J. Entomol. 2017, 114, 147–159. [Google Scholar] [CrossRef] [Green Version]
- Holzenthal, R.W.; Thomson, R.E.; Ríos-Touma, B. Order Trichoptera. In Thorp and Covich’s Freshwater Invertebrates: Ecology and General Biology; Thorp, J.H., Rogers, D.C., Eds.; Elsevier: London, UK, 2015; Volume 1, pp. 965–1002. [Google Scholar]
- Courtney, G.W.; Cranston, P.S. Diptera. In Thorp and Covich’s Freshwater Invertebrates: Ecology and General Biology; Thorp, J.H., Rogers, D.C., Eds.; Elsevier: London, UK, 2015; Volume 1, pp. 1043–1058. [Google Scholar]
- Biggs, B.J.F. Patterns in benthic algae of streams. In Algal Ecology: Freshwater Benthic Ecosystems; Stevenson, R.J., Bothwell, M.L., Lowe, R.L., Eds.; Academic Press: San Diego, CA, USA, 1996; pp. 31–56. [Google Scholar]
- Statzner, B.; Holm, T.F. Morphological adaptations of benthic invertebrates to stream flow—An old question studied by means of a new technique (Laser Doppler Anemometry). Oecologia 1982, 53, 290–292. [Google Scholar] [CrossRef]
- Courtney, G.W. Life history patterns of Nearctic mountain midges (Diptera: Deuterophlebiidae). J. North. Am. Benthol. Soc. 1991, 10, 177–197. [Google Scholar] [CrossRef]
- Ditsche-Kuru, P.; Barthlott, W.; Koop, J.H. At which surface roughness do claws cling? Investigations with larvae of the running water mayfly Epeorus assimilis (Heptageniidae, Ephemeroptera). Zoology 2012, 115, 379–388. [Google Scholar] [CrossRef]
- Higgins, M.J.; Crawford, S.A.; Mulvaney, P.; Wetherbee, R. Characterization of the adhesive mucilages secreted by live diatom cells using atomic force microscopy. Protist 2002, 153, 25–38. [Google Scholar] [CrossRef]
- Gebeshuber, I.C.; Thompson, J.B.; Del Amo, Y.; Stachelberger, H.; Kindt, J.H. In vivo nanoscale atomic force microscopy investigation of diatom adhesion properties. Mater. Sci. Tech. Lond. 2002, 18, 763–766. [Google Scholar] [CrossRef]
- Arens, W. Wear and tear of mouthparts: A critical problem in stream animals feeding on epilithic algae. Can. J. Zool. 1990, 68, 1896–1914. [Google Scholar] [CrossRef]
- McShaffrey, D.; McCafferty, W.P. Feeding behavior of Rhithrogena pellucida (Ephemeroptera: Heptageniidae). J. North. Am. Benthol. Soc. 1988, 7, 87–99. [Google Scholar] [CrossRef] [Green Version]
- Pančić, M.; Torres, R.R.; Almeda, R.; Kiørboe, T. Silicified cell walls as a defensive trait in diatoms. Proc. R. Soc. B Biol. Sci. 2019, 286, 20190184. [Google Scholar]
- Arens, W. Striking convergence in the mouthpart evolution of stream-living algae grazers. J. Zool. Syst. Evol. Res. 1994, 32, 319–343. [Google Scholar] [CrossRef]
- Lamberti, G.A.; Moore, J.W. Aquatic insects as primary consumers. In The Ecology of Aquatic Insects; Resh, V.H., Rosenberg, D.M., Eds.; Praeger: New Yok, NY, USA, 1984; pp. 164–195. [Google Scholar]
- Peterson, C.G. Gut passage and insect grazer selectivity of lotic diatoms. Freshwat. Biol. 1987, 18, 455–460. [Google Scholar] [CrossRef]
- Díaz Villanueva, V.; Albariño, R. Algal Ingestion and Digestion by Two Ephemeropteran Larvae from a Patagonian Andean Stream; Research update on Ephemeroptera & Plecoptera; University of Perugia: Perugia, Italy, 2003; pp. 468–475. [Google Scholar]
- Schmid-Araya, J.M.; Figueroa Hernández, D.; Schmid, P.E.; Drouot, C. Algivory in food webs of three temperate Andean rivers. Austral. Ecol. 2012, 37, 440–451. [Google Scholar] [CrossRef]
- Krebs, J.R. Optimal foraging: Decision rules for predators. In Behavioural Ecology: An Evolutionary Approach; Krebs, J.R., Davies, N.B., Eds.; Blackwell: Oxford, UK, 1978; pp. 23–63. [Google Scholar]
- Bo, T.; Fenoglio, S.; López-Rodríguez, M.J.; Tierno de Figueroa, J.M. Trophic behavior of two Perlidae species (Insecta, Plecoptera) in a river in southern Spain. Int. Rev. Hydrobiol. 2008, 93, 167–174. [Google Scholar] [CrossRef]
- Bo, T.; Fenoglio, S.; Malacarne, G. Diet of Dinocras cephalotes and Perla marginata (Plecoptera: Perlidae) in an Apennine stream (northwestern Italy). Can. Entomol. 2007, 139, 358–364. [Google Scholar] [CrossRef]
- Gregory, S.V. Plant-herbivore interactions in stream systems. In Stream Ecology. Application and Testing of General Ecological Theory; Barnes, J.R., Minshall, G.W., Eds.; Springer: Boston, MA, USA, 1983; pp. 157–189. [Google Scholar]
- Stevenson, R.J.; Bothwell, M.L.; Lowe, R.L.; Thorp, J.H. Algal Ecology: Freshwater Benthic Ecosystem; Academic press: Dubuque, MA, USA, 1996; p. 753. [Google Scholar]
- Graça, M.A.; Callisto, M.; Barbosa, J.E.L.; Firmiano, K.R.; França, J.; Júnior, J.F.G. Top-down and bottom-up control of epilithic periphyton in a tropical stream. Freshwat. Sci. 2018, 37, 857–869. [Google Scholar] [CrossRef]
- Holomuzki, J.R.; Feminella, J.W.; Power, M.E. Biotic interactions in freshwater benthic habitats. J. North. Am. Benthol. Soc. 2010, 29, 220–244. [Google Scholar] [CrossRef] [Green Version]
- Vilmi, A.; Tolonen, K.T.; Karjalainen, S.M.; Heino, J. Metacommunity structuring in a highly-connected aquatic system: Effects of dispersal, abiotic environment and grazing pressure on microalgal guilds. Hydrobiologia 2017, 790, 125–140. [Google Scholar] [CrossRef]
- Steinman, A.D.; McIntire, C.D.; Gregory, S.V.; Lamberti, G.A. Effects of irradiance and grazing on lotic algal assemblages. J. Phycol. 1989, 25, 478–485. [Google Scholar] [CrossRef]
- MacArthur, R.H. Population ecology of some warblers of north eastern coniferous forests. Ecology 1958, 39, 599–619. [Google Scholar] [CrossRef]
- Piano, E.; Doretto, A.; Falasco, E.; Gruppuso, L.; Fenoglio, S.; Bona, F. The role of recurrent dewatering events in shaping ecological niches of scrapers in intermittent Alpine streams. Hydrobiologia 2019, 841, 177–189. [Google Scholar] [CrossRef]
- Tall, L.; Cattaneo, A.; Cloutier, L.; Dray, S.; Legendre, P. Resource partitioning in a grazer guild feeding on a multilayer diatom mat. J. North. Am. Benthol. Soc. 2006, 25, 800–810. [Google Scholar] [CrossRef]
- Merritt, R.; Cummins, K.; Berg, M.N. An Introduction to the Aquatic Insects of North. America, 5th ed; Kendall Hunt: Dubuque, IA, USA, 2019; p. 498. [Google Scholar]
- Vadeboncoeur, Y.; Power, M.E. Attached algae: The cryptic base of inverted trophic pyramids in freshwaters. Annu. Rev. Ecol. Evol. S. 2017, 48, 255–279. [Google Scholar] [CrossRef]
- Herren, C.M.; Webert, K.C.; Drake, M.D.; Jake Vander Zanden, M.; Einarsson, Á.; Ives, A.R.; Gratton, C. Positive feedback between chironomids and algae creates net mutualism between benthic primary consumers and producers. Ecology 2017, 98, 447–455. [Google Scholar] [CrossRef]
- Hamm, C.E.; Merkel, R.; Springer, O.; Jurkojc, P.; Maier, C.; Prechtel, K.; Smetacek, V. Architecture and material properties of diatom shells provide effective mechanical protection. Nature 2003, 421, 841–843. [Google Scholar] [CrossRef] [Green Version]
- Fenoglio, S.; Bo, T.; Tierno de Figueroa, J.M.; Cucco, M. Nymphal growth, life cycle, and feeding habits of Potamanthus luteus (Linnaeus, 1767) (Insecta: Ephemeroptera) in the Bormida River, Northwestern Italy. Zool. Stud. 2008, 47, 185–190. [Google Scholar]
- Botana, L.M. Seafood and Freshwater Toxins. Pharmacology, Physiology and Detection; CRC Press: New York, NY, USA, 2000; p. 1215. [Google Scholar]
- Dhar, B.C.; Cimarelli, L.; Singh, K.S.; Brandi, L.; Brandi, A.; Puccinelli, C.; Marcheggiani, S.; Spurio, R. Molecular detection of a potentially toxic diatom species. Int. J. Environ. Res. Public Health 2015, 12, 4921–4941. [Google Scholar] [CrossRef] [Green Version]
- Violi, J.P.; Facey, J.A.; Mitrovic, S.M.; Colville, A.; Rodgers, K.J. Production of β-methylamino-L-alanine (BMAA) and its isomers by freshwater diatoms. Toxins 2019, 11, 512. [Google Scholar] [CrossRef] [Green Version]
- Falasco, E.; Bona, F. Recent findings regarding non-native or poorly known diatom taxa in north-western Italian rivers. J. Limn. 2013, 201, 35–51. [Google Scholar] [CrossRef]
- Ladrera, R.; Goma, J.; Prat, N. Effects of Didymosphenia geminata massive growth on stream communities: Smaller organisms and simplified food web structure. PLoS ONE 2018, 13, e0193545. [Google Scholar] [CrossRef]
- Bray, J.P.; Kilroy, C.; Gerbeaux, P.; Burdon, F.J.; Harding, J.S. Ecological processes mediate the effects of the invasive bloom-forming diatom Didymosphenia geminata on stream algal and invertebrate assemblages. Hydrobiologia 2020, 847, 177–190. [Google Scholar] [CrossRef]
- Wahl, M. Epibiosis. In Marine Hard Bottom Communities; Wahl, L., Ed.; Springer: Boston, MA, USA, 2009; pp. 61–72. [Google Scholar]
- Falasco, E.; Bo, T.; Ghia, D.; Gruppuso, L.; Bona, F.; Fenoglio, S. Diatoms prefer strangers: Non-indigenous crayfish host completely different epizoic algal diatom communities from sympatric native species. Biol. Inv. 2018, 20, 2767–2776. [Google Scholar] [CrossRef]
- Wahl, M.; Goecke, F.; Labes, A.; Dobretsov, S.; Weinberger, F. The second skin: Ecological role of epibiotic biofilms on marine organisms. Front. Microbiol. 2012, 3, 292. [Google Scholar] [CrossRef] [Green Version]
- Robinson, N.J.; Majewska, R.; Lazo-Wasem, E.A.; Nel, R.; Paladino, F.V.; Rojas, L.; Zardus, J.D.; Pinou, T. Epibiotic diatoms are universally present on all sea turtle species. PLoS ONE 2016, 11, e0157011. [Google Scholar] [CrossRef] [Green Version]
- Wetzel, C.E.; Van de Vijver, B.; Cox, E.J.; Bicudo, D.D.C.; Ector, L. Tursiocola podocnemicola sp. nov., a new epizoic freshwater diatom species from the Rio Negro in the Brazilian Amazon Basin. Diatom Res. 2012, 27, 1–8. [Google Scholar] [CrossRef]
- Abbott, L.L.; Bergey, E.A. Why are there few algae on snail shells? The effects of grazing, nutrients and shell chemistry on the algae on shells of Helisoma trivolvis. Freshwat. Biol. 2007, 52, 2112–2120. [Google Scholar] [CrossRef]
- Wujek, D.E. The first occurrence of the chrysophyte alga Amphirhiza epizootica from North America. Mich. Bot. 2006, 45, 197–200. [Google Scholar]
- Winemiller, K.; Winsborough, B. Occurrence of epizoic communities on the parasitic copepod Lernaea carassii (Lernaeidae). Southwest. Nat. 1990, 35, 206–210. [Google Scholar] [CrossRef]
- Fuelling, L.J.; Adams, J.A.; Badik, K.J.; Bixby, R.J.; Caprette, C.L.; Caprette, H.E.; Hall, M.M. An unusual occurrence of Thorea hispida (Thore) Desvaux chantransia on rusty crayfish in West Central Ohio. Nova Hedwig. 2012, 94, 355–366. [Google Scholar] [CrossRef]
- Bergey, E.A.; Resh, V.H. Interactions between a stream caddisfly and the algae on its case: Factors affecting algal quantity. Freshwat. Biol. 1994, 31, 153–163. [Google Scholar] [CrossRef]
- Wujek, D.E. Epizooic diatoms on the cerci of Ephemeroptera (Caenidae) naiads. Great Lakes Entomol. 2013, 46, 117–120. [Google Scholar]
- Stewart, K.W.; Schlichting, H.E., Jr. Dispersal of algae and protozoa by selected aquatic insects. J. Ecol. 1966, 54, 551–562. [Google Scholar] [CrossRef]
- Revill, D.L.; Stewart, K.W.; Schlichting, H.E. Passive dispersal of viable algae and protozoa by certain craneflies and midges. Ecology 1967, 48, 1023–1027. [Google Scholar] [CrossRef]
- Göthe, E.; Angeler, D.G.; Gottschalk, S.; Löfgren, S.; Sandin, L. The influence of environmental, biotic and spatial factors on diatom metacommunity structure in Swedish headwater streams. PLoS ONE 2013, 8, e72237. [Google Scholar]
- Burgazzi, G.; Bolpagni, R.; Laini, A.; Racchetti, E.; Viaroli, P. Algal biomass and macroinvertebrate dynamics in intermittent braided rivers: New perspectives from instream pools. River Res. Appl. 2020. [Google Scholar] [CrossRef]
- Wisz, M.S.; Pottier, J.; Kissling, W.D.; Pellissier, L.; Lenoir, J.; Damgaard, C.F.; Dormann, C.F.; Forchhammer, M.C.; Grytnes, J.-A.; Guisan, A.; et al. The role of biotic interactions in shaping distributions and realised assemblages of species: Implications for species distribution modelling. Biol. Rev. 2013, 88, 15–30. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| FFG | Type of Trophic Resources |
|---|---|
| Shredders | Coarse (>1 mm) particulate organic matter (CPOM): fragments of leaves, plant tissue and wood debris |
| Scrapers | Attached benthic algae (i.e., diatoms, green algae, and cyanobacteria) |
| Collector-gatherers | Fine (50 µm –1 mm) particulate organic matter (FPOM) |
| Collector-filterers | Very fine (<50 µm) particulate organic matter and micro-organisms in the water column |
| Predators | Small animals |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fenoglio, S.; Tierno de Figueroa, J.M.; Doretto, A.; Falasco, E.; Bona, F. Aquatic Insects and Benthic Diatoms: A History of Biotic Relationships in Freshwater Ecosystems. Water 2020, 12, 2934. https://doi.org/10.3390/w12102934
Fenoglio S, Tierno de Figueroa JM, Doretto A, Falasco E, Bona F. Aquatic Insects and Benthic Diatoms: A History of Biotic Relationships in Freshwater Ecosystems. Water. 2020; 12(10):2934. https://doi.org/10.3390/w12102934
Chicago/Turabian StyleFenoglio, Stefano, José Manuel Tierno de Figueroa, Alberto Doretto, Elisa Falasco, and Francesca Bona. 2020. "Aquatic Insects and Benthic Diatoms: A History of Biotic Relationships in Freshwater Ecosystems" Water 12, no. 10: 2934. https://doi.org/10.3390/w12102934
APA StyleFenoglio, S., Tierno de Figueroa, J. M., Doretto, A., Falasco, E., & Bona, F. (2020). Aquatic Insects and Benthic Diatoms: A History of Biotic Relationships in Freshwater Ecosystems. Water, 12(10), 2934. https://doi.org/10.3390/w12102934