Chorological and Ecological Differentiation of the Commonest Leech Species from the Suborder Erpobdelliformes (Arhynchobdellida, Hirudinea) on the Balkan Peninsula



 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Statistical Analyses
3. Results
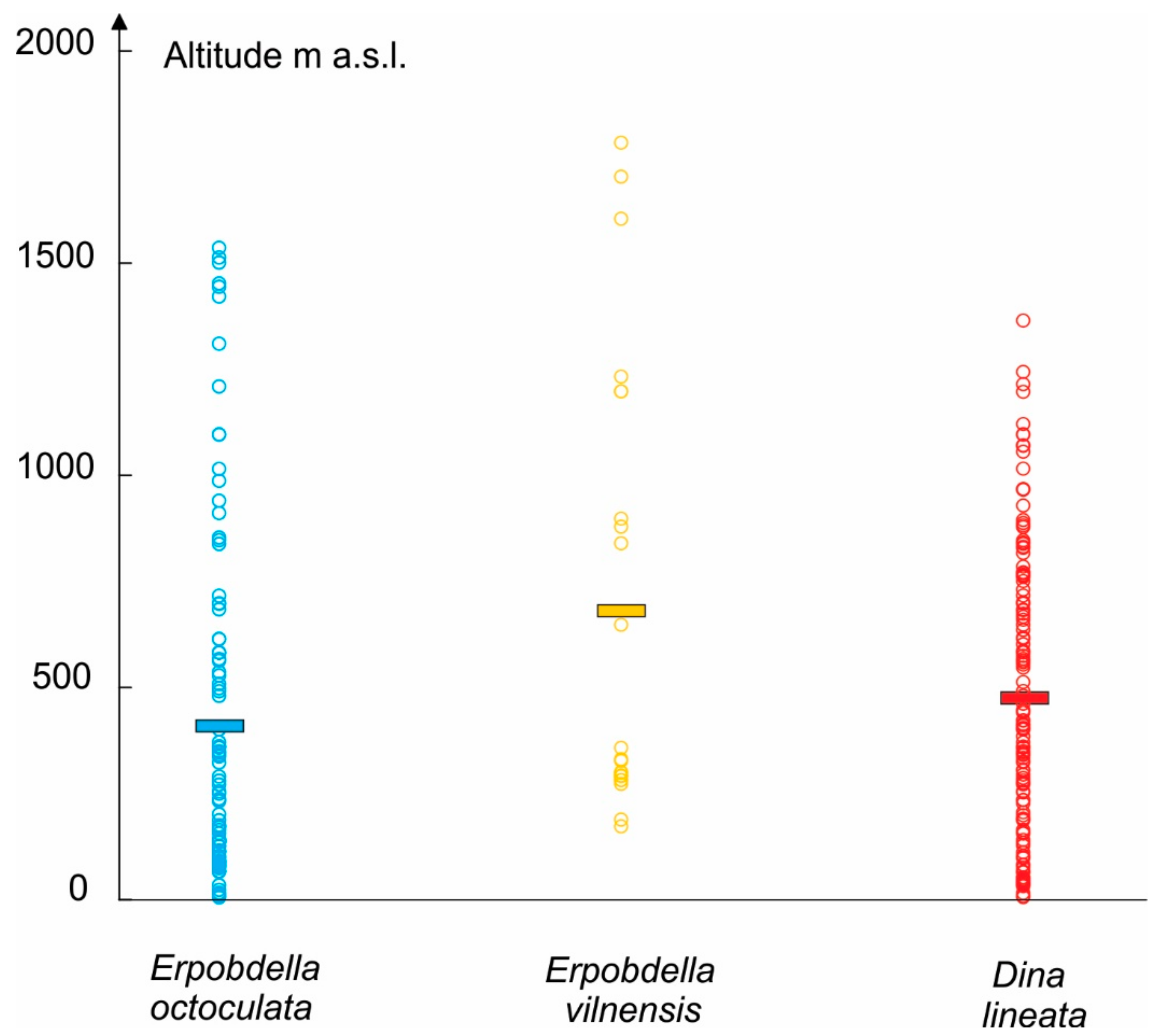
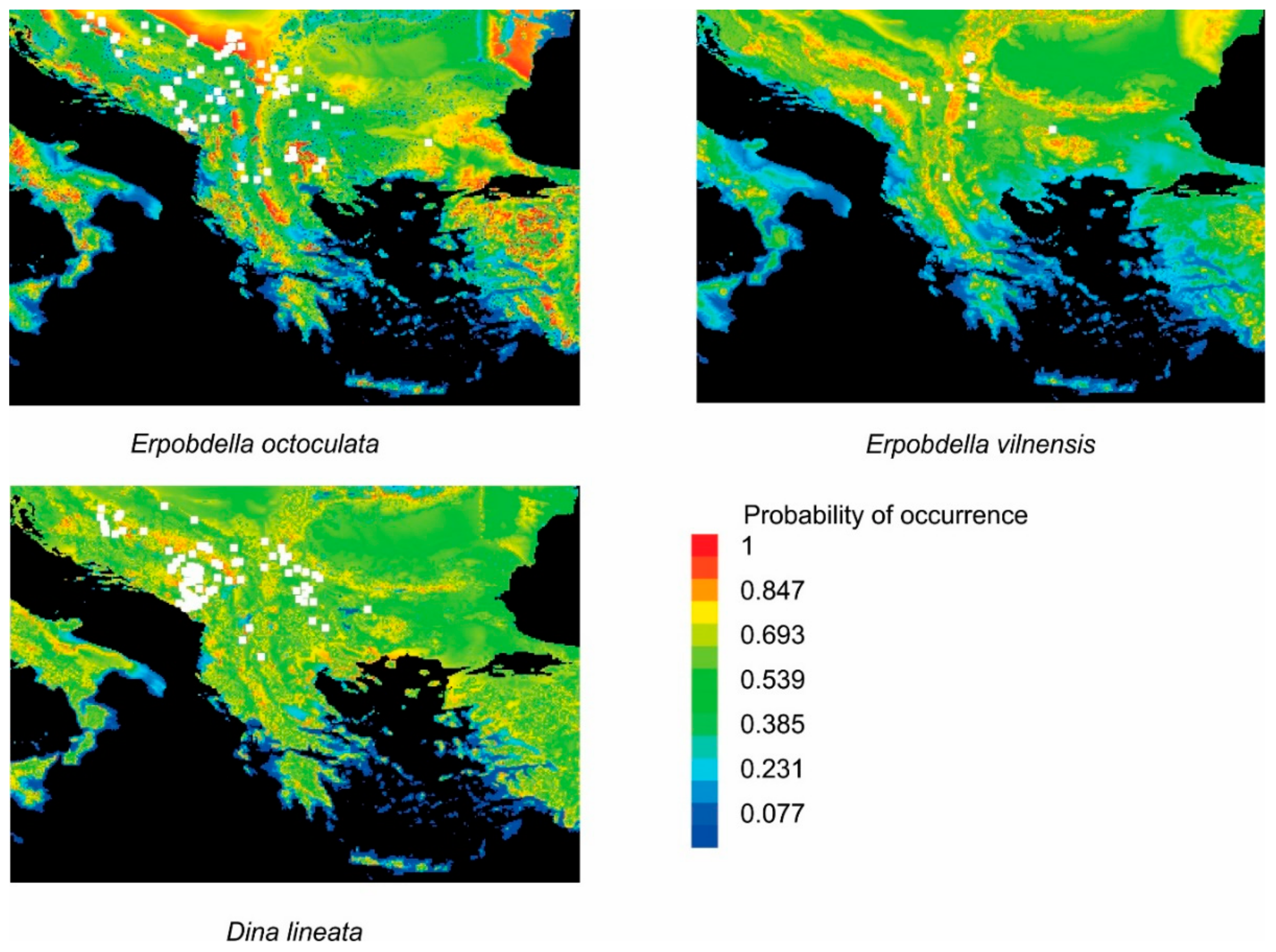
3.1. The Actual Distribution of the Analyzed Species
3.2. The Potential Distribution of the Analyzed Species
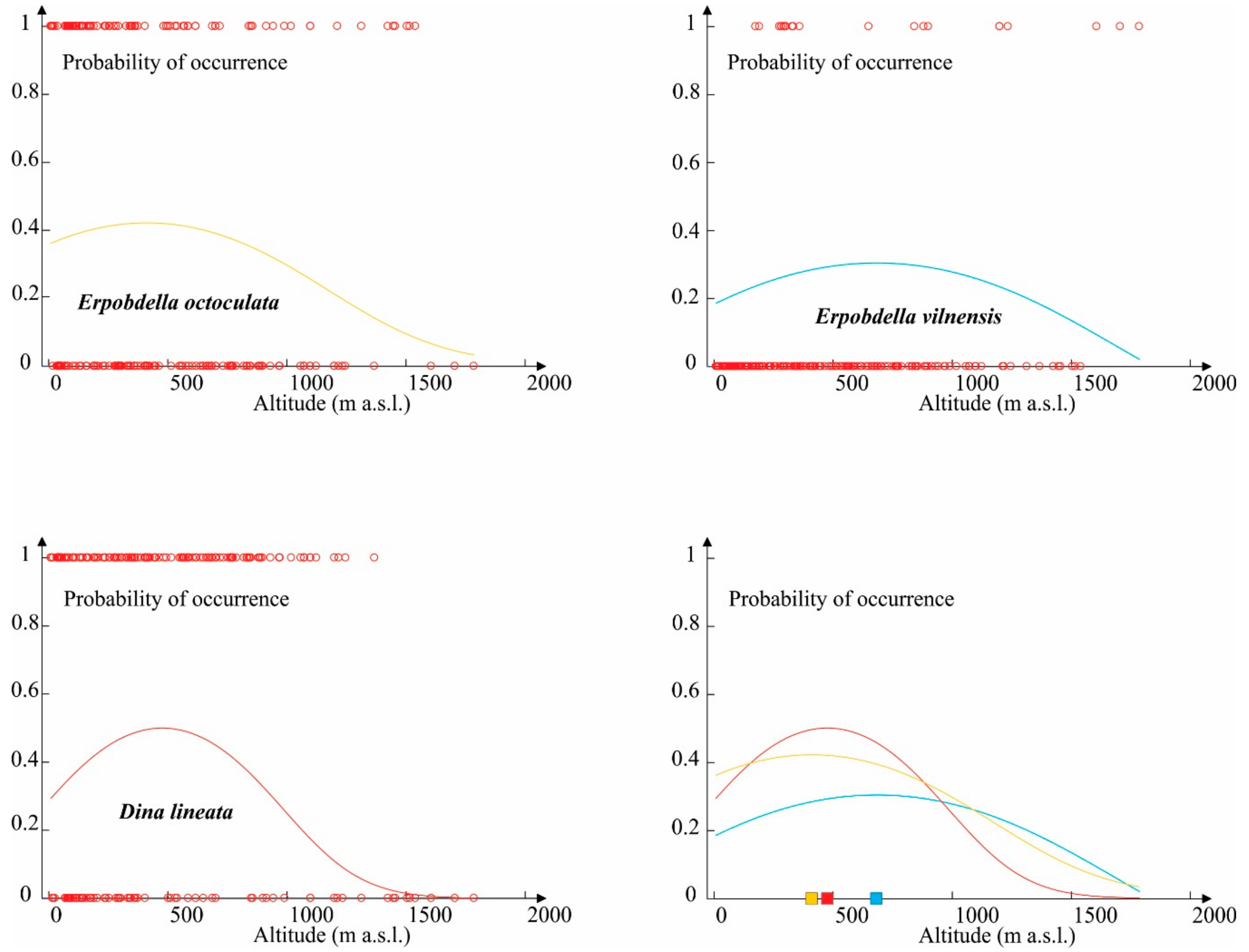
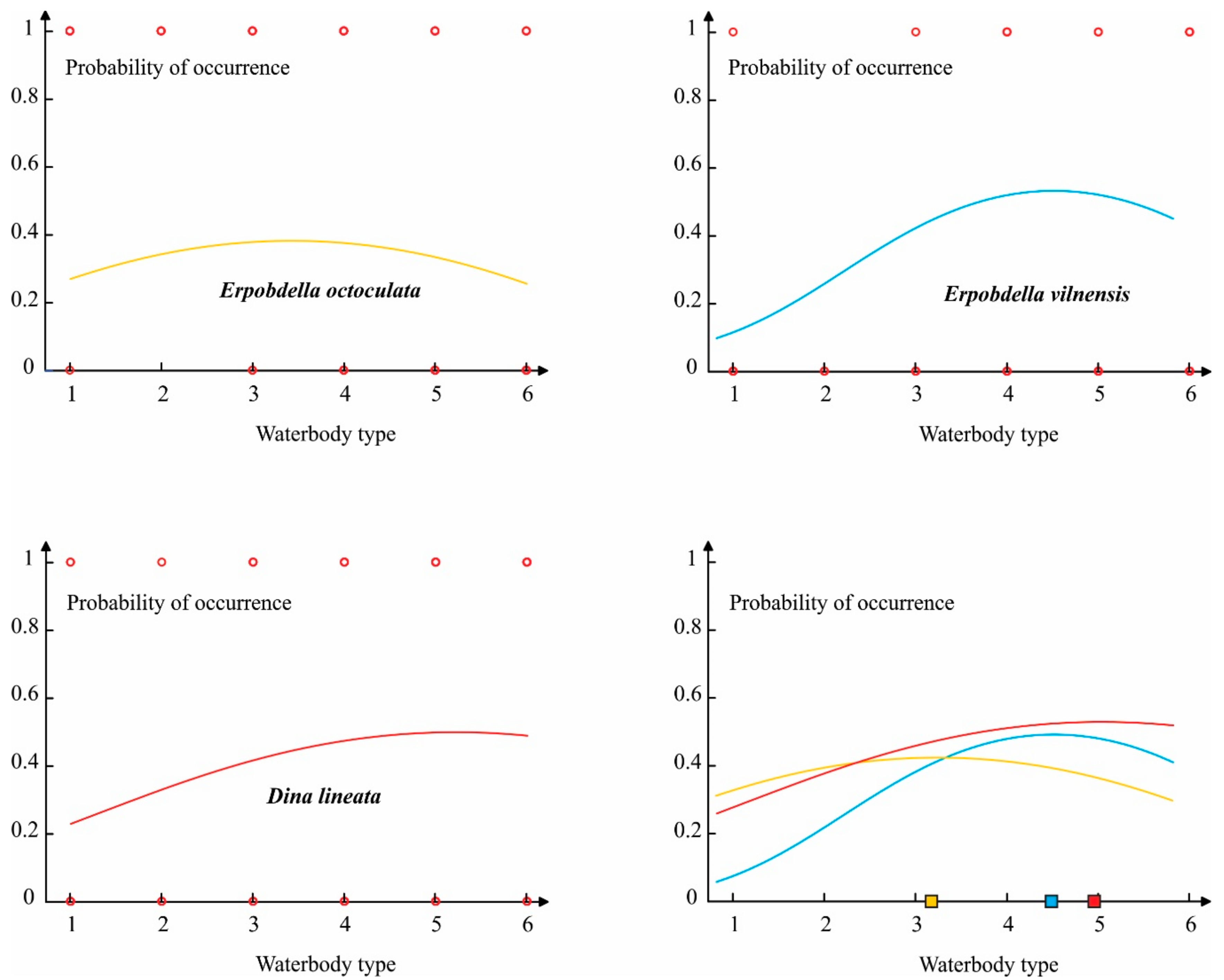
3.3. The Ecological Differentiation of Analyzed Species
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Blanchard, R. Hirudineen aus Montenegro. Sitzs-Ber. Königl. Böhm. Ges. Wiss. Prag. 1905, 1–3. [Google Scholar]
- Augener, H. Hirudineen aus jugoslavischen Seen. Festschr. Zum 1937, 60, 403–413. [Google Scholar]
- Rémy, P. Sangsues de Yougoslavie. Bull. Soc. Zool. Fr. 1937, 62, 140–148. [Google Scholar]
- Sket, B. K Poznavanju Favne Pijavk (Hirudinea) v Jugoslaviji, Zur Kenntnis der Egelfauna (Hirudinea) Jugoslawiens. Acad. Sci. Artium Slov. Cl. IV Hist. Nat. Med. Diss. Ljubl. 1968, 9, 127–197. [Google Scholar]
- Šapkarev, J.A. The fauna of Hirudinea of Macedonia. The taxonomy and distribution of leeches of Aegean lakes. Int. Rev. Ges. Hydrobiol. Hydrogr. 1970, 55, 317–324. [Google Scholar] [CrossRef]
- Šapkarev, J. Contribution to the knowledge the earthworms (Lumbricidae) and leeches (Hirudinea) of Kosovo, Yugoslavia. Annu. Fac. Sci. L’université Skopje 1975, 27, 39–54. [Google Scholar]
- Šapkarev, J.A. Composition and dynamics of the bottom animals in the littoral zone of Dojran Lake, Macedonia: With 11 figures and 4 tables in the text. Verh. Int. Verein. Theor. Angew. Limnol. 1975, 19, 1339–1350. [Google Scholar] [CrossRef]
- Sket, B.; Šapkarev, J. Dina lepinja sp. n. (Hirudinea, Erpobdellidae), a new endemic leech from the ancient lake Ohridsko Ezero. Biološki Vestn. 1986, 34, 89–92. [Google Scholar]
- Sket, B.; Šapkarev, J. Distribution of Hirudinea (Annelida) in the ancient Ohrid Lake region. Arch. Hydrobiol. 1992, 124, 225–237. [Google Scholar]
- Sket, B.; Dovč, P.; Jalžić, B.; Kerovec, M.; Kučinić, M.; Trontelj, P. A cave leech (Hirudinea, Erpobdellidae) from Croatia with unique morphological features. Zool. Scr. 2001, 30, 223–229. [Google Scholar] [CrossRef]
- Trontelj, P.; Sket, B. Molecular re-assessment of some phylogenetic, taxonomic and biogeographic relationships between the leech genera Dina and Trocheta (Hirudinea: Erpobdellidae). Hydrobiologia 2000, 438, 227–235. [Google Scholar] [CrossRef]
- Grosser, C.; Pešić, V. First record of Batracobdelloides moogi (Hirudinea: Glossiphoniidae) in the Balkans. Nat. Montenegr. 2005, 4, 29–32. [Google Scholar]
- Grosser, C.; Moritz, G.; Pešić, V. Dina minuoculata sp. nov. (Hirudinea: Erpobdellidae)—Eine neue Egelart aus Montenegro. Lauterbornia 2007, 59, 7–18. [Google Scholar]
- Grosser, C. First record of Trocheta haskonis Grosser, 2000 (Hirudinea: Erpobdellidae) in Serbia. Lauterbornia 2013, 76, 111–113. [Google Scholar]
- Grosser, C.; Pešić, V.; Dmitrović, D. Dina sketi n. sp., a new erpobdellid leech (Hirudinida: Erpobdellidae) from Bosnia and Herzegovina. Zootaxa 2014, 3793, 393–397. [Google Scholar] [CrossRef]
- Grosser, C.; Pešić, V.; Gligorović, B. A checklist of the Leeches (Annelida: Hirudinea) of Montenegro. Ecol. Montenegrina 2014, 2, 20–28. [Google Scholar]
- Grosser, C.; Pešić, V.; Lazarević, P. A checklist of the Leeches (Annelida: Hirudinida) of Serbia, with new records. Fauna Balk. 2015, 3, 71–86. [Google Scholar]
- Grosser, C.; Pešić, V.; Berlajolli, V.; Gligorović, B. Glossiphonia balcanica n. sp. and Dina prokletijaca n. sp. (Hirudinida: Glossiphoniidae, Erpobdellidae)—Two new leeches from Montenegro and Kosovo. Ecol. Montenegrina 2016, 8, 17–26. [Google Scholar]
- Grosser, C.; Šukalo, G.; Pešić, V. Monster from the Vault: A new finding of one of the largest European leech Trocheta haskonis Grosser, 2000 from Bosnia and Herzegovina. Ecol. Montenegrina 2018, 19, 69–72. [Google Scholar]
- Trajanovski, S.; Albrecht, C.; Schreiber, K.; Schultheiß, R.; Stadler, T.; Benke, M.; Wilke, T. Testing the spatial and temporal framework of speciation in an ancient lake species flock: The leech genus Dina (Hirudinea: Erpobdellidae) in Lake Ohrid. Biogeosciences 2010, 7, 3387–3402. [Google Scholar] [CrossRef] [Green Version]
- Utevsky, S.; Utevsky, A.; Pešić, V. First record of Glossiphonia nebulosa (Hirudinida: Glossiphoniidae) from the Skadar Lake in Montenegro. Lauterbornia 2013, 76, 123–125. [Google Scholar]
- Živić, I.; Radosavljević, T.; Stojanović, K.; Petrović, A. The first molecular characterization of the genus Hirudo on the territory of Serbia: Estimation of endangerment. Aquat. Ecol. 2015, 49, 81–90. [Google Scholar] [CrossRef]
- Živić, I.; Cvetković, A.; Božanić, M.; Radojević, A.; Stojanović, K. Checklist of Freshwater Leeches (Hirudinea) in Serbia. Water Res. Manag. 2017, 7, 35–41. [Google Scholar]
- Marinković, N.; Karadžić, B.; Pešić, V.; Gligorović, B.; Grosser, C.; Paunović, M.; Nikolić, V.; Raković, M. Faunistic patterns and diversity components of leech assemblages in karst springs of Montenegro. Knowl. Manag. Aquat. Ecosyst. 2019, 420, 26. [Google Scholar] [CrossRef]
- Sket, B.; Trontelj, P. Global diversity of leeches (Hirudinea) in freshwater. Hydrobiologia 2007, 595, 129–137. [Google Scholar] [CrossRef]
- Sket, B. Intralacustrine speciation in the genus Dina (Hirudinea, Erpobdellidae) in Lake Ohrid (Yugoslavia). Hydrobiologia 1989, 182, 49–59. [Google Scholar] [CrossRef]
- Sawyer, R.T. Leech Biology and Behavior; Clarendon Press: Oxford, UK, 1986; Volume 2, pp. 419–793. [Google Scholar] [CrossRef]
- Persson, L.; Elliott, J.M. Population variation and individual maximum size in two leech populations: Energy extraction from cannibalism or niche widening? Oecologia 2013, 172, 119–127. [Google Scholar] [CrossRef]
- Oceguera-Figueroa, A.; Phillips, A.J.; Pacheco-Chaves, B.; Reeves, W.K.; Siddall, M.E. Phylogeny of macrophagous Leeches (Hirudinea, Clitellata) based on molecular data and evaluation of the barcoding locus. Zool. Scr. 2011, 40, 194–203. [Google Scholar] [CrossRef]
- Nesemann, H.; Neubert, E. Süßwasserfauna von Mitteleuropa, Bd. 6, Annelida, 2, Clitellata: Branchiobdellida, Acanthobdellea, Hirudinea; Spektrum Akademischer Verlag: Berlin/Heidelberg, Germany, 1999; p. 178. [Google Scholar]
- Kutschera, U. The feeding strategies of the leech Erpobdella octoculata (L.): A laboratory study. Int. Rev. Hydrobiol 2003, 88, 94–101. [Google Scholar] [CrossRef]
- Schenková, J.; Sychra, J.; Kubová, B. The freshwater leeches (Clitellata: Hirudinida) of the Czech Republic–list of taxa and remarks on rare and endangered species. Dep. Zool. Bot. Fac. Sci. Masaryk Univ. Kotlarska 2005, 2, 11–37. [Google Scholar] [CrossRef]
- Koperski, P. Urban environments as habitats for rare aquatic species: The case of leeches (Euhirudinea, Clitellata) in Warsaw freshwaters. Limnologica 2010, 40, 233–240. [Google Scholar] [CrossRef] [Green Version]
- Kubová, N.; Schenková, J. Tolerance, optimum ranges and ecological requirements of free-living leech species (Clitellata: Hirudinida). Arch. Hydrobiol. 2014, 185, 167–180. [Google Scholar] [CrossRef]
- Official Gazette of the Republic of Serbia. The Parameters of Ecological and Chemical Status of Surface Waters and Parameters of the Chemical and Quantitative Status of Groundwater; Official Gazette of the Republic of Serbia: Belgrade, Serbia, 2011. [Google Scholar]
- Hijmans, R.J.; Guarino, L.; Mathur, P. 2012: DIVA-GIS Version 7.5 Manual. Available online: http://www.diva-gis.org/ (accessed on 1 June 2018).
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Dudik, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudik, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Whittaker, R.H. Evolution and measurement of species diversity. Taxon 1972, 21, 213–251. [Google Scholar] [CrossRef] [Green Version]
- Stanković, S. Ekologija Životinja; Univerzitet u Beogradu. Zavod za izdavanje udžbenika Narodne Republike Srbije: Belgrade, Serbia, 1962; pp. 374–388. [Google Scholar]
- Vannote, R.L.; Minshall, G.W.; Cummins, K.W.; Sedell, J.R.; Cushing, C.E. The river continuum concept. Can. J. Fish. Aquat. Sci. 1980, 37, 130–137. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F. Correspondence Analysis of Incidence and Abundance Data: Properties in Terms of a Unimodal Response Model. Biometrics 1985, 41, 859–873. [Google Scholar] [CrossRef]
- James, G.; Wittenmm, D.; Hastie, T.; Tibshirani, R. An Introduction to Statistical Learning with Applications in R; Springer: New York, NY, USA; Heidelberg, Germany; Dordrecht, The Netherlands; London, UK, 2013; p. 426. [Google Scholar]
- Karadžić, B. FLORA: A software package for statistical analysis of ecological data. Water Res. Manag. 2013, 3, 45–54. [Google Scholar]
- Nesemann, H.F.; Moog, O. Hirudinea. In Fauna Aquatica Austriaca, 3rd edition A Comprehensive Species Inventory of Austrian Aquatic Organisms with Ecological Notes; Abt. IV/3, Stubenring 1, A-1010; Bundesministerium für Land- und Forstwirtschaft, Umwelt und Wasserwirtschaft: Vienna, Austria, 2017. [Google Scholar]
- Kubová, N.; Schenková, J.; Horsák, M. Environmental determinants of leech assemblage patterns in lotic and lenitic habitats. Limnologica 2013, 43, 516–524. [Google Scholar] [CrossRef]
- van Haaren, T.; Hop, H.; Soes, M.; Tempelman, D. The freshwater leeches (Hirudinea) of the Netherlands. Lauterbornia 2004, 52, 113–131. [Google Scholar]
- Westendorff, M.; Kalettka, T.; Jueg, U. Occurrence of leeches (Hirudinea) in different types of water bodies in northeast Germany (Brandenburg). Lauterbornia 2008, 65, 153–162. [Google Scholar]
- Bielecki, A.; Cichocka, J.M.; Jeleń, I.; Świątek, P.; Adamiak-Brud, Ż. A checklist of leech species from Poland. Wiad. Parazytol. 2011, 57, 11–20. [Google Scholar]
- Novaković, B. Indicative ecological status assessment of the Južna Morava River based on aquatic macroinvertebrates. Water Res. Manag. 2012, 2, 45–50. [Google Scholar]
- Savić, A.; Ranđelović, V.; Branković, S.; Krpo-Ćetković, J. Mayfly (Insecta: Ephemeroptera) community structure as an indicator of the ecological status of the Nišava river (Central Balkan Peninsula). Aquat. Ecosyst. Health Manag. 2011, 14, 276–284. [Google Scholar] [CrossRef]
- Cichocka, J.; Jabłońska-Barna, I.; Bielecki, A.; Buczyńska, E.; Buczyński, P.; Stryjecki, R.; Pikuła, D. Leeches (Clitellata: Hirudinida) of an upland stream: Taxonomic composition in relation to habitat conditions. Oceanol. Hydrobiol. Stud. 2015, 44, 245–253. [Google Scholar] [CrossRef]
- Kazancı, N.; Ekingen, P.; Dügel, M.; Türkmen, G. Hirudinea (Annelida) species and their ecological preferences in some running waters and lakes. Int. J. Environ. Sci. Technol. 2015, 12, 1087–1096. [Google Scholar] [CrossRef] [Green Version]
- Nesemann, H.; Csanyi, B. On the leech fauna (Hirudinea) of the Tisza river basin in Hungary with notes on the faunal history. Lauterbornia 1993, 14, 41–70. [Google Scholar]
- Utevsky, S.Y.; Son, M.O.; Dyadichko, V.G.; Kaygorodova, J.A. New information on the geographical distribution of Erpobdella vilnensis (Liskiewicz, 1915) (Hirudinida, Erpobdellidae) in Ukraine. Lauterbornia 2012, 75, 75–78. [Google Scholar]
- Košel, V. Checklist of Hirudinea of the Czech Republic. Acta Musei Morav. Sci. Biol. 2014, 99, 1–14. [Google Scholar]
- Koperski, P. Relative importance of factors determining diversity and composition of freshwater leech assemblages (Hirudinea; Clitellata): A metaanalysis. Arch. Hydrobiol. 2006, 166, 325–341. [Google Scholar] [CrossRef]
- Aqem Consortium. Manual for the application of the AQEM system. In A Comprehensive Method to Assess European Streams Using Benthic Macroinvertebrates, Developed for the Purpose of the Water Framework Directive; Version 1; The AQEM consortium: Duisburg-Esen, Germany, 2002; pp. 619–630. [Google Scholar]
- Directive, W.F. Water Framework Directive—Directive 2000/60/EC of the European Parliament and of the Council Establishing a Framework for Community Action in the Field of Water Policy. Off. J. Eur. Communities 2000, 22, 2000. [Google Scholar]
- Paunović, M.; Grošelj, S.; Milačić, R.; Grđan, S.; Zuliani, T.; Vidaković, I.; Vučković, I.; Jelena, V.; Ščančar, J.; Makovinska, J.; et al. Steps towards integrated water management in the Sava River Basin. Water Res. Manag. 2016, 6, 3–10. [Google Scholar]
- Radinger, J.; Wolter, C.; Kail, J. Spatial scaling of environmental variables improves species-habitat models of fishes in a small, sand-bed lowland river. PLoS ONE 2015, 10, e0142813. [Google Scholar] [CrossRef] [Green Version]
- Moog, O.; Hartmann, A. Fauna Aquatica Austriaca, 3rd edition A Comprehensive Species Inventory of Austrian Aquatic Organisms with Ecological Notes; Abt. IV/3, Stubenring 1, A-1010; Bundesministerium für Land- und Forstwirtschaft, Umwelt und Wasserwirtschaft: Vienna, Austria, 2017. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Waterbody Type | Waterbodies Included | Flow Velocity | Bottom Material |
|---|---|---|---|
| T1 | lakes, ponds and reservoirs | stagnant → slow | very fine sediment (silt, clay mud and sand) |
| T2 | lower stretches of large lowland rivers | slow | very fine sediment (silt, clay mud and sand) |
| T3 | main tributaries of large lowland rivers | slow → medium | fine to medium size sediment (mud, sand and gravel) |
| T4 | medium to small (wadeable) rivers, elevation below 500 m | medium | hard substratum, large fractions (gravel, stones) |
| T5 | medium to small (wadeable) rivers, elevation above 500 m | medium → fast | hard substratum, large fractions (gravel, stones and rocks) |
| T6 | mix of small waterbodies including springs and upper stretches of streams | fast | hard substratum, large fractions (gravel, stones and rocks) |
| Number of Sampling Sites: T/Ws | ||||
|---|---|---|---|---|
| Waterbody Type | Adriatic Sea | Aegean Sea | Black Sea | Σ |
| T1 | 3 | 3 | 18 | 24 |
| T2 | 0 | 1 | 7 | 8 |
| T3 | 2 | 0 | 15 | 17 |
| T4 | 2 | 5 | 33 | 40 |
| T5 | 4 | 2 | 42 | 48 |
| T6 | 46 | 3 | 43 | 92 |
| Σ | 57 | 14 | 158 | 229 |
| Number of Records: Sp/Ws | ||||
| Species | Adriatic Sea | Aegean Sea | Black Sea | Σ |
| E. octoculata | 11 | 10 | 78 | 99 |
| E. vilnensis | 1 | 4 | 18 | 23 |
| D. lineata | 48 | 4 | 90 | 142 |
| Σ | 60 | 18 | 186 | 264 |
| Number of Records: Sp/T | ||||
| Waterbody Type | E. octoculata | E. vilnensis | D. lineata | Σ |
| T1 | 11 | 1 | 3 | 15 |
| T2 | 23 | 0 | 1 | 24 |
| T3 | 8 | 1 | 9 | 18 |
| T4 | 13 | 9 | 15 | 37 |
| T5 | 25 | 4 | 35 | 64 |
| T6 | 19 | 8 | 79 | 106 |
| Σ | 99 | 23 | 142 | 264 |
| Geographic Variable | Parameter | E. octoculata | E. vilnensis | D. lineata |
|---|---|---|---|---|
| Altitude (m) | Minimum | 9 | 175 | 9 |
| Maximum | 1540 | 1786 | 1774 | |
| Mean | 418.636 | 683.608 | 494.267 | |
| Standard deviation | 408.194 | 528.766 | 368.935 | |
| Latitude (°) | Minimum | 40.903 | 40.903 | 40.903 |
| Maximum | 45.409 | 44.225 | 45.054 | |
| Mean | 43.414 | 43.312 | 43.120 | |
| Standard deviation | 1.104 | 0.818 | 0.742 | |
| Longitude (°) | Minimum | 16.383 | 19.215 | 16.683 |
| Maximum | 25.785 | 24.009 | 24.009 | |
| Mean | 20.196 | 21.175 | 19.698 | |
| Standard deviation | 1.813 | 1.189 | 1.458 |
| Species | E. octoculata | E. vilnensis | D. lineata |
|---|---|---|---|
| E. octoculata | 1 | −0.145 | −0.625 * |
| E. vilnensis | - | 1 | −0.247 |
| D. lineata | - | - | 1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marinković, N.; Karadžić, B.; Slavevska Stamenković, V.; Pešić, V.; Nikolić, V.; Paunović, M.; Raković, M. Chorological and Ecological Differentiation of the Commonest Leech Species from the Suborder Erpobdelliformes (Arhynchobdellida, Hirudinea) on the Balkan Peninsula. Water 2020, 12, 356. https://doi.org/10.3390/w12020356
Marinković N, Karadžić B, Slavevska Stamenković V, Pešić V, Nikolić V, Paunović M, Raković M. Chorological and Ecological Differentiation of the Commonest Leech Species from the Suborder Erpobdelliformes (Arhynchobdellida, Hirudinea) on the Balkan Peninsula. Water. 2020; 12(2):356. https://doi.org/10.3390/w12020356
Chicago/Turabian StyleMarinković, Nikola, Branko Karadžić, Valentina Slavevska Stamenković, Vladimir Pešić, Vera Nikolić, Momir Paunović, and Maja Raković. 2020. "Chorological and Ecological Differentiation of the Commonest Leech Species from the Suborder Erpobdelliformes (Arhynchobdellida, Hirudinea) on the Balkan Peninsula" Water 12, no. 2: 356. https://doi.org/10.3390/w12020356
APA StyleMarinković, N., Karadžić, B., Slavevska Stamenković, V., Pešić, V., Nikolić, V., Paunović, M., & Raković, M. (2020). Chorological and Ecological Differentiation of the Commonest Leech Species from the Suborder Erpobdelliformes (Arhynchobdellida, Hirudinea) on the Balkan Peninsula. Water, 12(2), 356. https://doi.org/10.3390/w12020356