Zooplankton Community Responses to Oxygen Stress

 ,
,  ,
,
Abstract
:1. Introduction
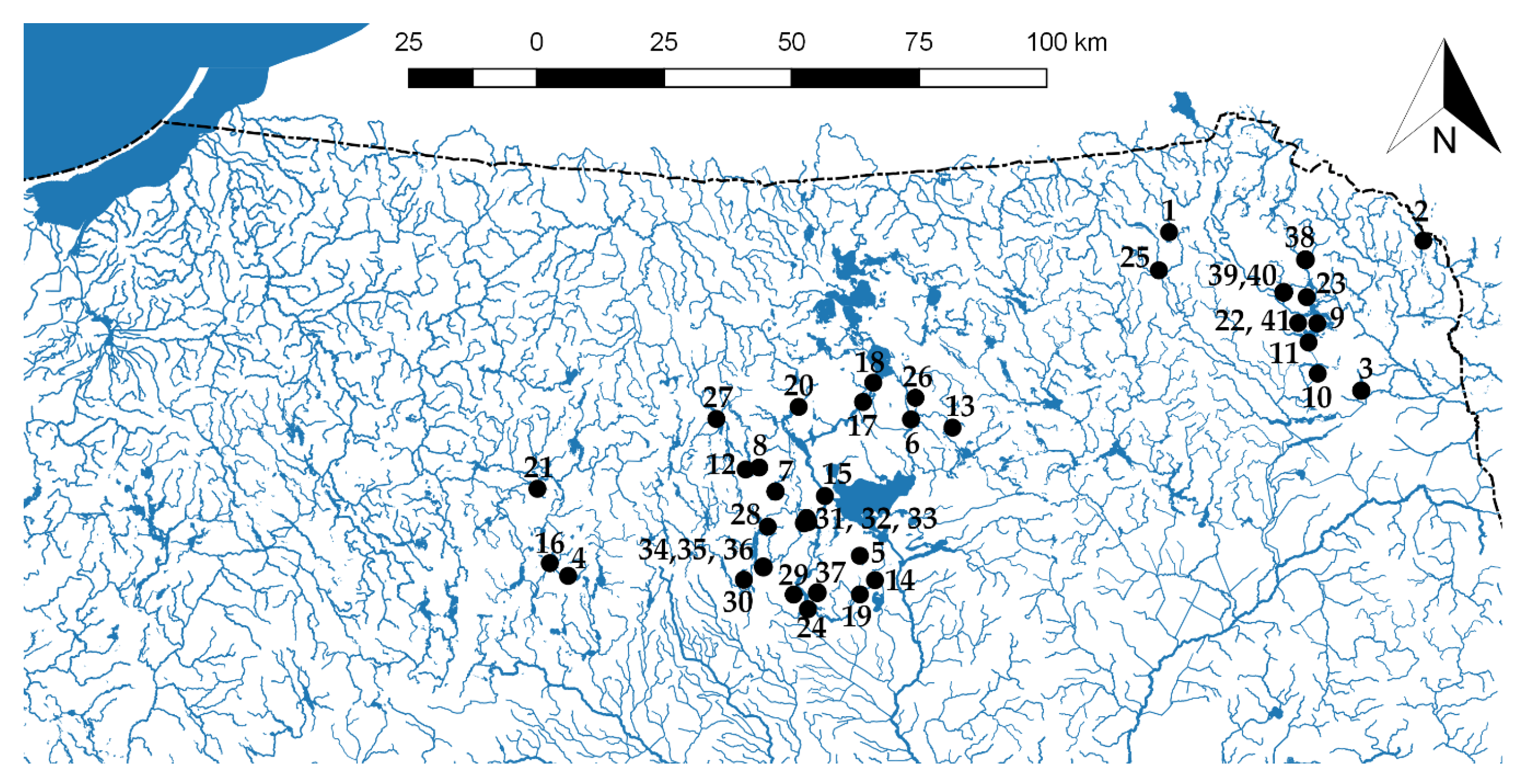
2. Study Area and Methods
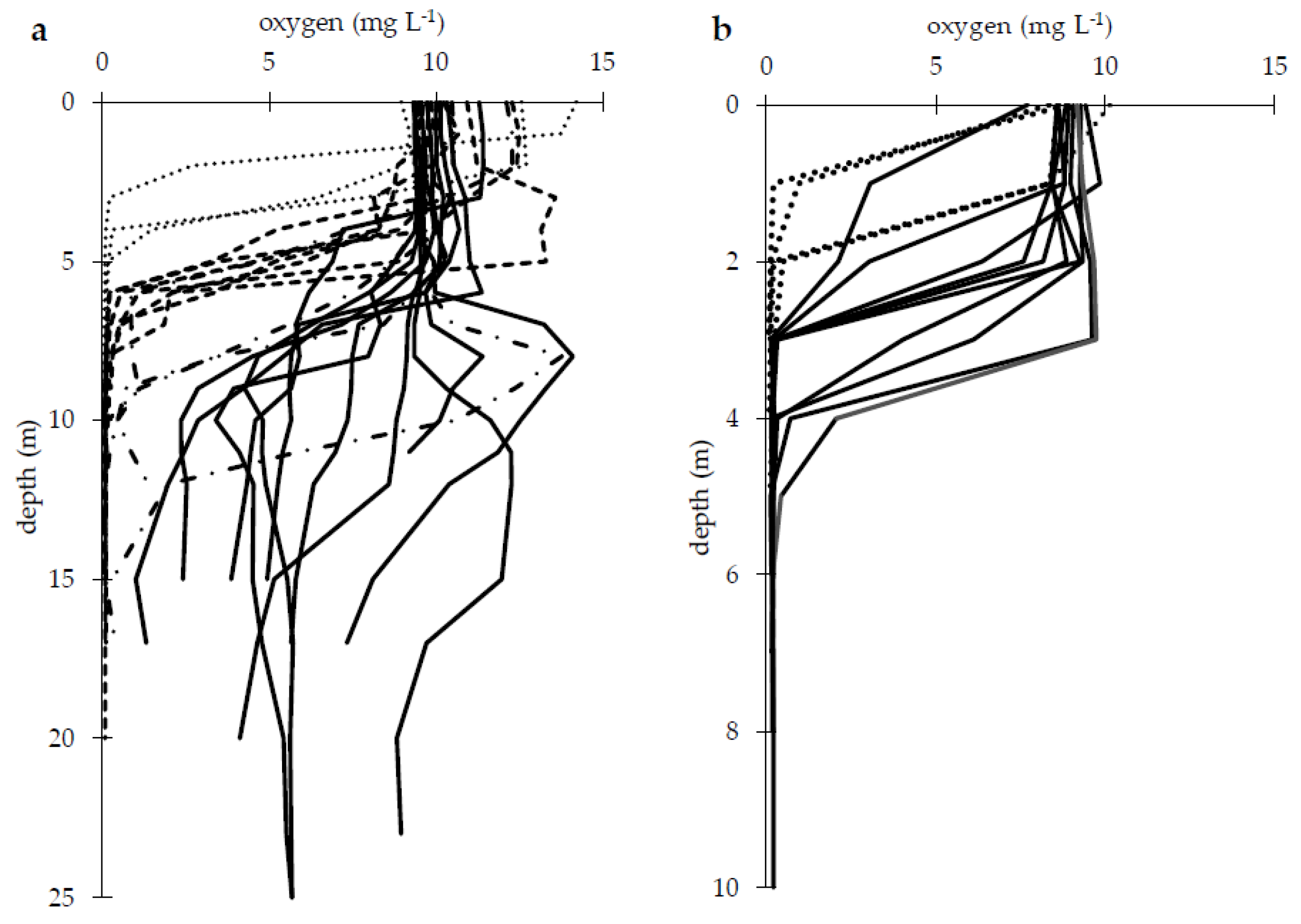
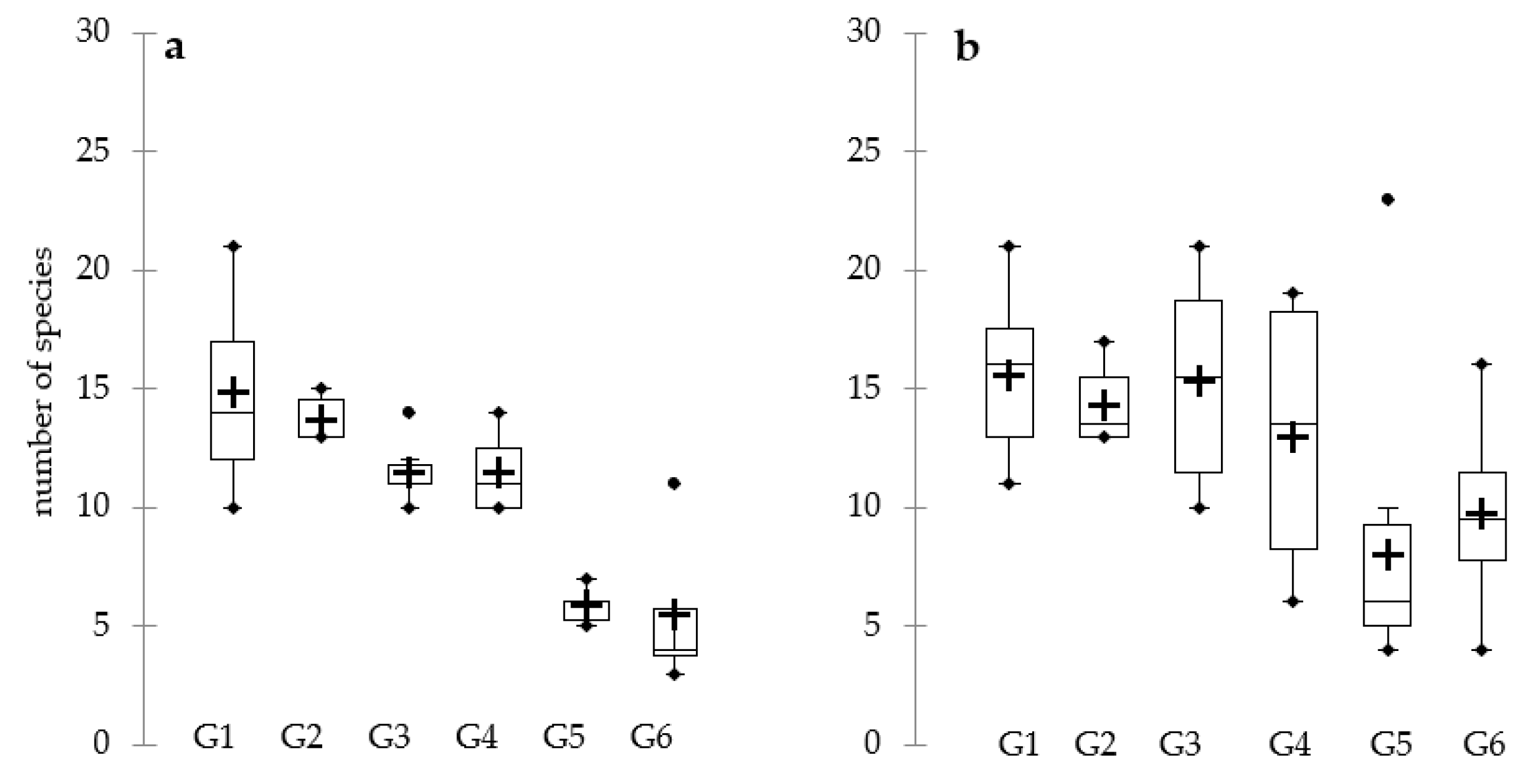
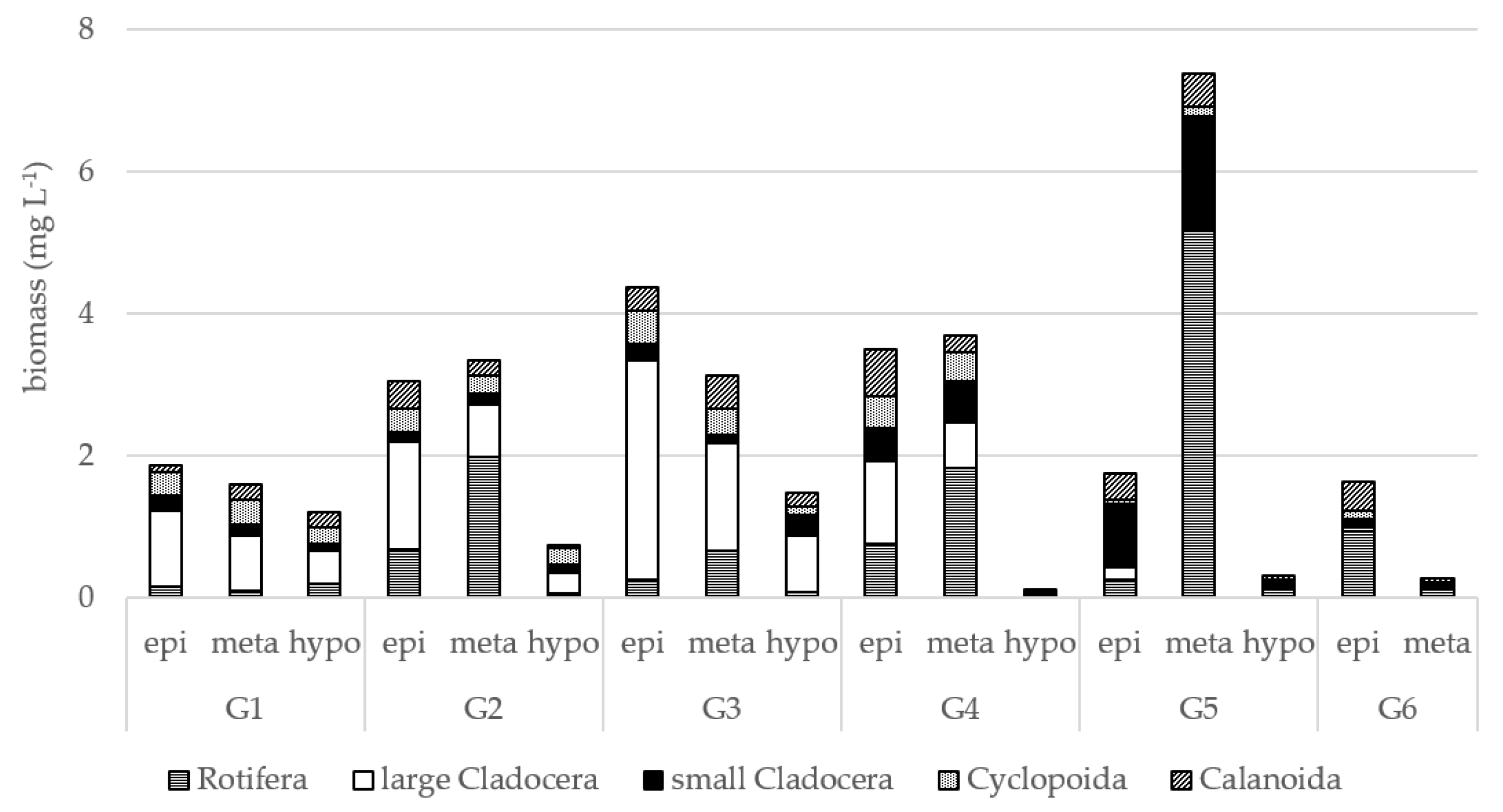
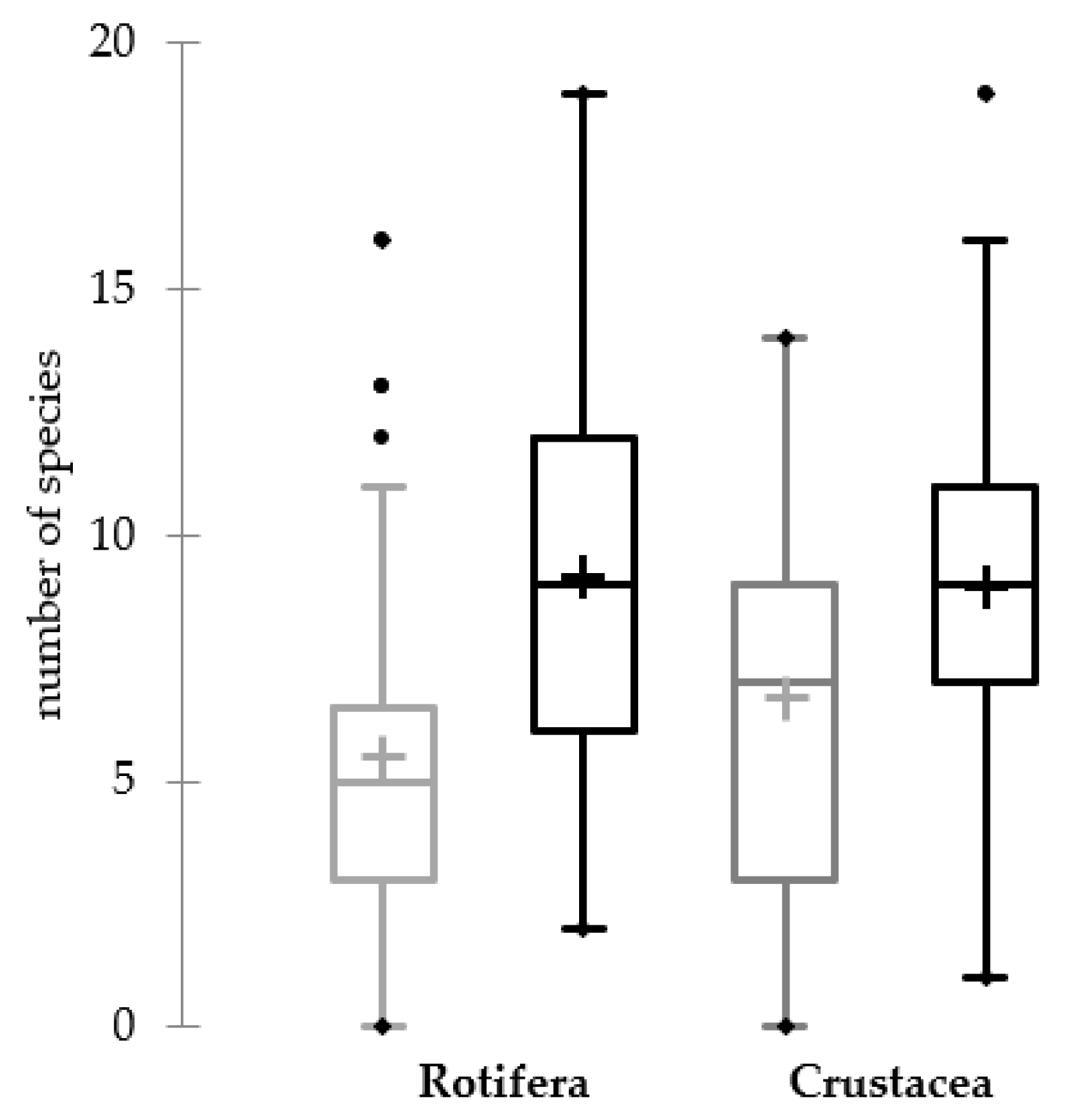
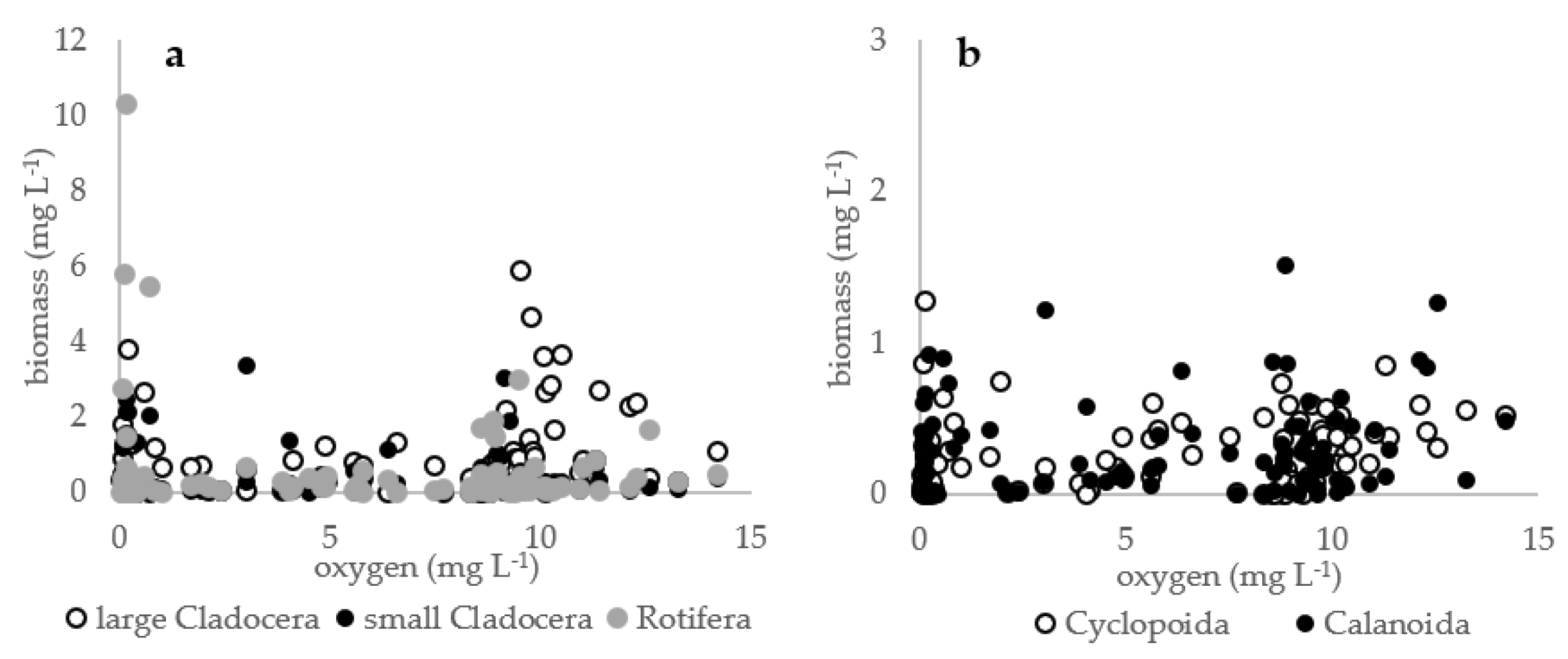
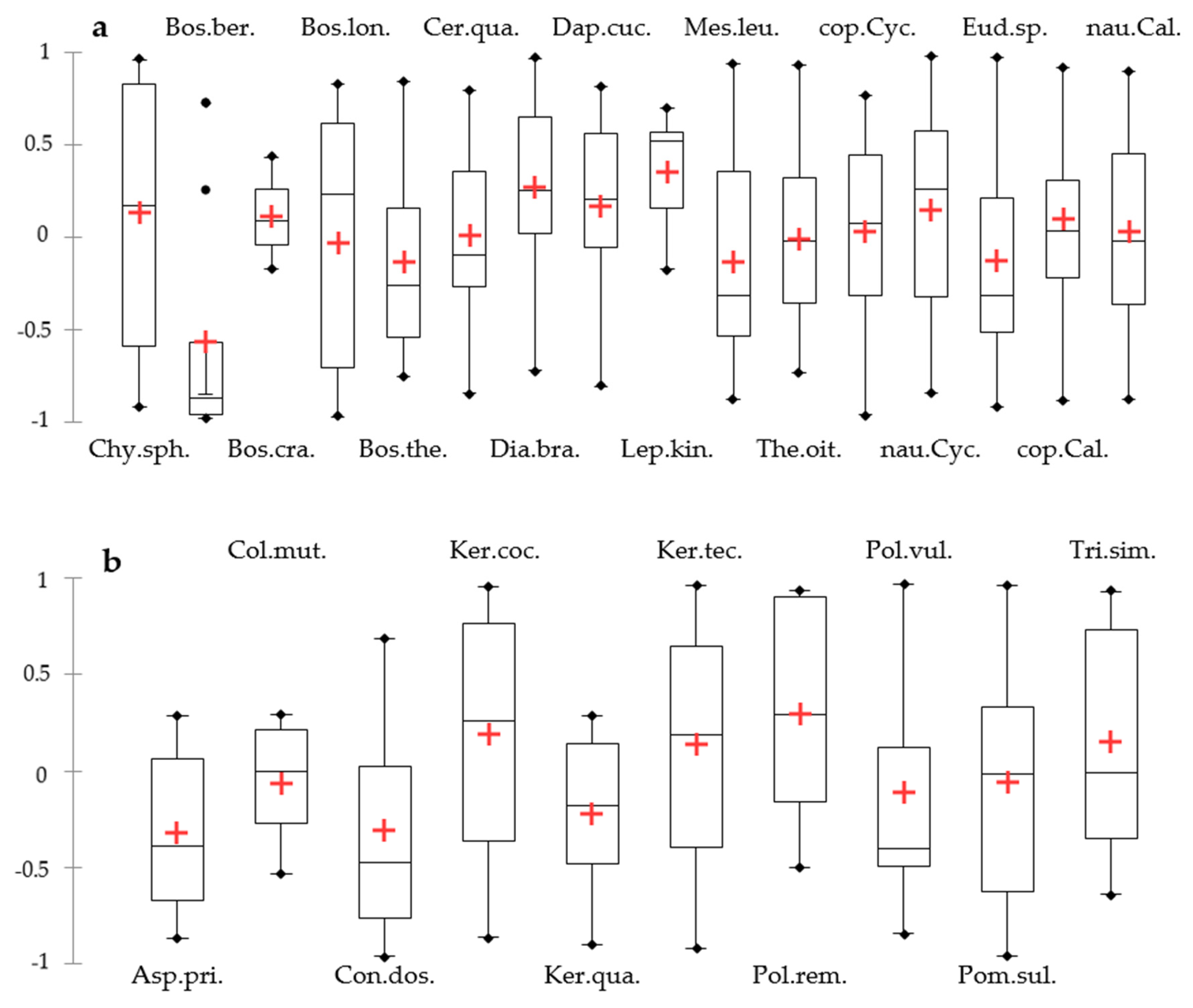
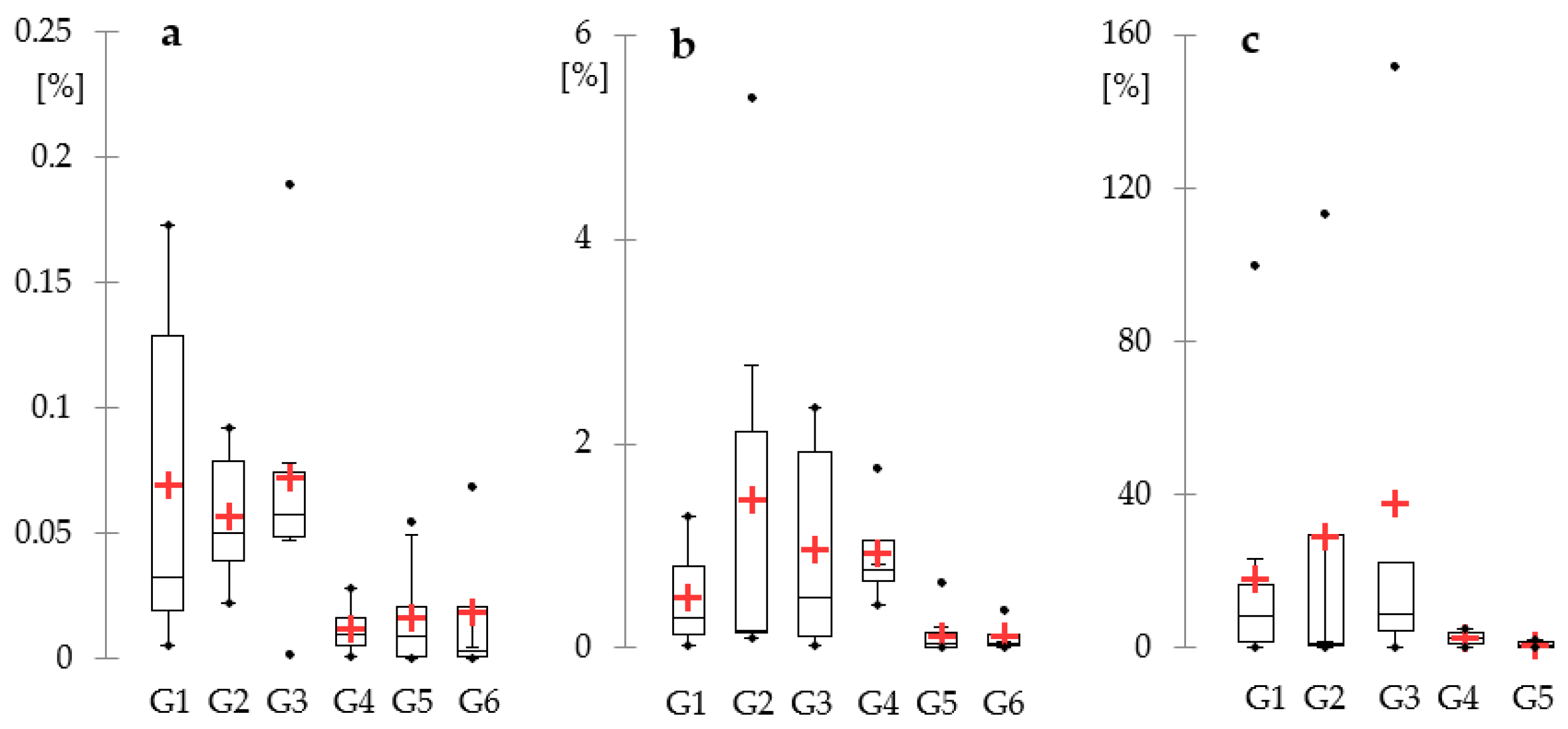
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ekau, W.; Auel, H.; Pörtner, H.-O.; Gilbert, D. Impacts of hypoxia on the structure and processes in pelagic communities (zooplankton, macro-invertebrates and fish). Biogeosciences 2010, 7, 1669–1699. [Google Scholar] [CrossRef] [Green Version]
- Doubek, J.P.; Campbell, K.L.; Lofton, M.E.; McClure, R.P.; Carey, C.C. Hypolimnetic Hypoxia Increases the Biomass Variability and Compositional Variability of Crustacean Zooplankton Communities. Water 2019, 11, 2179. [Google Scholar] [CrossRef] [Green Version]
- Górniak, A.; Kajak, Z. Hydrobiologia. Limnologia; Wydawnictwo Naukowe PWN: Warszawa, Poland, 2019; pp. 45–108. ISBN 978-83-01-20721-2. (In Polish) [Google Scholar]
- Winder, M.; Schindler, D.E. Climatic effects on the phenology of lake processes. Glob. Chang. Boil. 2004, 10, 1844–1856. [Google Scholar] [CrossRef] [Green Version]
- Jankowski, T.; Livingstone, D.M.; Bührer, H.; Forster, R.; Niederhauser, P. Consequences of the 2003 European heat wave for lake temperature profiles, thermal stability, and hypolimnetic oxygen depletion: Implications for a warmer world. Limnol. Oceanogr. 2006, 51, 815–819. [Google Scholar] [CrossRef] [Green Version]
- Butcher, J.B.; Nover, D.; Johnson, T.E.; Clark, C.M. Sensitivity of lake thermal and mixing dynamics to climate change. Clim. Chang. 2015, 129, 295–305. [Google Scholar] [CrossRef] [Green Version]
- Diaz, R.J.; Rosenberg, R. Spreading dead zones and consequences for marine ecosystems. Science 2008, 321, 926–929. [Google Scholar] [CrossRef]
- Jenny, J.P.; Francus, P.; Normandeau, A.; Lapointe, F.; Perga, M.E.; Ojala, A.; Schimmelmann, A.; Zolitschka, B. Global spread of hypoxia in freshwater ecosystems during the last three centuries is caused by rising local human pressure. Glob. Chang. Biol. 2016, 22, 1481–1489. [Google Scholar] [CrossRef]
- Doubek, J.P.; Campbell, K.L.; Doubek, K.M.; Hamre, K.D.; Lofton, M.E.; McClure, R.P.; Ward, N.K.; Carey, C.C. The effects of hypolimnetic anoxia on the diel vertical migration of freshwater crustacean zooplankton. Ecosphere 2018, 9, e02332. [Google Scholar] [CrossRef] [Green Version]
- Kolar, C.S.; Rahel, F.J. Interaction of a biotic factor (predator presence) and an abiotic factor (low oxygen) as an influence on benthic invertebrate communities. Oecologia 1993, 95, 210–219. [Google Scholar] [CrossRef]
- Ludsin, S.A.; Zhang, X.; Brandt, S.B.; Roman, M.R.; Boicourt, W.C.; Mason, D.M.; Costantini, M. Hypoxia-avoidance by planktivorous fish in Chesapeake Bay: Implications for food web interactions and fish recruitment. J. Exp. Mar. Boil. Ecol. 2009, 381, S121–S131. [Google Scholar] [CrossRef]
- Miller, D.; Poucher, S.; Coiro, L.L. Determination of lethal dissolved oxygen levels for selected marine and estuarine fishes, crustaceans, and a bivalve. Mar. Biol. 2002, 140, 287–296. [Google Scholar] [CrossRef]
- Vaquer-Sunyer, R.; Duarte, C.M. Thresholds of hypoxia for marine biodiversity. Proc. Natl. Acad. Sci. USA 2008, 105, 15452–15457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, K.H.; Imai, H.; Ayukawa, K.; Sugahara, S.; Nakano, S.I.; Seike, Y. Impact of improved bottom hypoxia on zooplankton community in shallow eutrophic lake. Knowl. Manag. Aquat. Ecosyst. 2013, 408, 1–8. [Google Scholar] [CrossRef]
- Karpowicz, M.; Ejsmont-Karabin, J. Influence of environmental factors on vertical distribution of zooplankton communities in humic lakes. Ann. Limnol. Int. J. Limnol. 2018, 54, 17. [Google Scholar] [CrossRef]
- Karpowicz, M.; Ejsmont-Karabin, J.; Więcko, A.; Górniak, A.; Cudowski, A. A place in space - the horizontal vs. vertical factors that influence zooplankton (Rotifera, Crustacea) communities in a mesotrophic lake. J. Limnol. 2019, 78, 243–258. [Google Scholar] [CrossRef]
- Stalder, L.C.; Marcus, N.H. Zooplankton responses to hypoxia: Behavioral patterns and survival of three species of calanoid copepods. Mar. Biol. 1997, 127, 599–607. [Google Scholar] [CrossRef]
- Wright, D.; Shapiro, J. Refuge availability: A key to understanding the summer disappearance of Daphnia. Freshw. Biol. 1990, 24, 43–62. [Google Scholar] [CrossRef]
- Field, K.M.; Prepas, E.E. Increased abundance and depth distribution of pelagic crustacean zooplankton during hypolimnetic oxygenation in a deep, eutrophic Alberta lake. Can. J. Fish. Aquat. Sci. 1997, 54, 2146–2156. [Google Scholar] [CrossRef]
- Weider, L.J.; Lampert, W. Differential response of Daphnia genotypes to oxygen stress: Respiration rates, hemoglobin content and low-oxygen tolerance. Oecologia 1985, 65, 487–491. [Google Scholar] [CrossRef]
- Klumb, R.A.; Bunch, K.L.; Mills, E.L.; Rudstam, L.G.; Brown, G.; Knauf, C.; Burton, R.; Arrhenius, F. Establishment of a metalimnetic oxygen refuge for zooplankton in a productive Lake Ontario embayment. Ecol. Appl. 2004, 14, 113–131. [Google Scholar] [CrossRef]
- Stanley, D.R.; Wilson, C.A. Effect of hypoxia on the distribution of fishes associated with a petroleum platform off coastal Louisiana. N. Am. J. Fish. Manag. 2004, 24, 662–671. [Google Scholar] [CrossRef]
- Vanderploeg, H.A.; Ludsin, S.A.; Cavaletto, J.F.; Höök, T.O.; Pothoven, S.A.; Brandt, S.B.; Liebig, J.R.; Lang, G.A. Hypoxic zones as habitat for zooplankton in Lake Erie: Refuges from predation or exclusions zones? J. Exp. Mar. Biol. Ecol. 2009, 381, 108–120. [Google Scholar] [CrossRef]
- Vanderploeg, H.A.; Ludsin, S.A.; Ruberg, S.A.; Hook, T.O.; Pothoven, S.A.; Brandt, S.B.; Lang, G.A.; Liebig, J.R. Hypoxia affects spatial distributions and overlap of pelagic fish, zooplankton, and phytoplankton in Lake Erie. J. Exp. Mar. Biol. Ecol. 2009, 381, 92–107. [Google Scholar] [CrossRef]
- Larsson, P.; Lampert, W. Experimental evidence of a low oxygen refuge for large zooplankton. Limnol. Oceanogr. 2011, 56, 1682–1688. [Google Scholar] [CrossRef]
- Karpowicz, M.; Ejsmont-Karabin, J. Effect of metalimnetic gradient on phytoplankton and zooplankton (Rotifera, Crustacea) communities in different trophic conditions. Environ. Monit. Assess. 2017, 189, 367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karpowicz, M.; Kalinowska, K. Vertical distribution of the relic species Eurytemora lacustris (Copepoda, Calanoida) in stratified mesotrophic lakes. Biologia 2018, 73, 1197–1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, M.; Hoshi, T. Analysis of respiratory role of haemoglobin in Daphnia magna. Zool. Sci. 1984, 1, 523–532. [Google Scholar]
- Hanazato, T.; Dodson, S.I. Synergistic effects of low oxygen concentration, predator kairomone, and a pesticide on the cladoceran Daphnia pulex. Limnol. Oceanogr. 1995, 40, 700–709. [Google Scholar] [CrossRef]
- Larsson, P.; Lampert, W. Finding the optimal vertical distribution: Behavioural responses of Daphnia pulicaria to gradients of environmental factors and the presence of fish. Freshw. Biol. 2012, 57, 2514–2525. [Google Scholar] [CrossRef]
- Wilczyński, W.; Dynak, P.; Babkiewicz, E.; Bernatowicz, P.; Maszczyk, P. The combined effects of hypoxia and fish kairomones on several physiological and life history traits of Daphnia. Freshw. Biol. 2019, 64, 2204–2220. [Google Scholar] [CrossRef]
- Maszczyk, P.; Babkiewicz, E.; Ciszewski, K.; Dabrowski, K.; Dynak, P.; Krajewski, K.; Urban, P.; Żebrowski, M.; Wilczynski, W. Combined effects of elevated epilimnetic temperature and metalimnetic hypoxia on the predation rate of planktivorous fish. J. Plankton Res. 2019, 41, 709–722. [Google Scholar] [CrossRef] [PubMed]
- Sorensen, M.L.; Branstrator, D.K. The North American invasive zooplanktivore Bythotrephes longimanus is less hypoxia-tolerant than the native Leptodora kindtii. Can. J. Fish. Aquat. Sci. 2017, 74, 824–832. [Google Scholar] [CrossRef] [Green Version]
- Berzins, B.; Pejler, B. Rotifer occurrence in relation to oxygen content. Hydrobiologia 1989, 183, 165–172. [Google Scholar] [CrossRef]
- Jekatierynczuk-Rudczyk, E.; Zieliński, P.; Grabowska, M.; Ejsmont-Karabin, J.; Karpowicz, M.; Więcko, A. The trophic status of Suwałki Landscape Park lakes based on selected parameters (NE Poland). Environ. Monit. Assess. 2014, 186, 5101–5121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Górniak, A. A new version of the Hydrochemical Dystrophy Index to evaluate dystrophy in lakes. Ecol. Indic. 2017, 78, 566–573. [Google Scholar] [CrossRef]
- Carlson, R.E. A trophic state index for lakes. Limnol. Oceanogr. 1977, 22, 361–369. [Google Scholar] [CrossRef] [Green Version]
- Drzymulska, D.; Zieliński, P. Developmental changes in the historical and present-day trophic status of brown water lakes. Are humic water bodies a uniform aquatic ecosystem? Wetlands 2013, 33, 909–919. [Google Scholar] [CrossRef] [Green Version]
- Kring, S.A.; Figary, S.E.; Boyer, G.L.; Watson, S.B.; Twiss, M.R. Rapid in situ measures of phytoplankton communities using the bbe FluoroProbe: Evaluation of spectral calibration, instrument intercompatibility, and performance range. Can. J. Fish. Aquat. Sci. 2014, 71, 1087–1095. [Google Scholar] [CrossRef]
- Neal, C.; Neal, M.; Wickham, H. Phosphate measurement in natural waters: Two examples of analytical problems associated with silica interference using phosphomolybdic acid methodologies. Sci. Total Environ. 2000, 251–252, 513–542. [Google Scholar] [CrossRef]
- Błędzki, L.A.; Rybak, J.I. Freshwater Crustacean Zooplankton of Europe; Springer: Cham, Switzerland, 2016; pp. 1–918. [Google Scholar] [CrossRef]
- Ejsmont-Karabin, J. Empirical equations for biomass calculation of planktonic rotifers. Pol. Arch. Hydrobiol. 1998, 45, 513–522. [Google Scholar]
- Hart, R.C.; Bychek, E.A. Body size in freshwater planktonic crustaceans: An overview of extrinsic determinants and modifying influences of biotic interactions. Hydrobiologia 2011, 668, 61–108. [Google Scholar] [CrossRef]
- Feniova, I.; Dawidowicz, P.; Ejsmont-Karabin, J.; Gladyshev, M.I.; Kalinowska, K.; Karpowicz, M.; Kostrzewska-Szlakowska, I.; Majsak, N.; Petrosyan, V.; Razlutskij, V.; et al. Effects of zebra mussels on cladoceran communities under eutrophic conditions. Hydrobiologia 2018, 822, 37–54. [Google Scholar] [CrossRef] [Green Version]
- Karpowicz, M.; Feniova, I.; Gladyshev, M.I.; Ejsmont-Karabin, J.; Górniak, A.; Zieliński, P.; Dawidowicz, P.; Kolmakova, A.A.; Dzialowski, A.R. The stoichiometric ratios (C:N:P) in a pelagic food web under experimental conditions. Limnologica 2019, 77, 125690. [Google Scholar] [CrossRef] [Green Version]
- Ivlev, V.S. Experimental Ecology of the Feeding of Fishes; Yale University Press: New Haven, CT, USA, 1961; pp. 1–302. [Google Scholar]
- Gaevsky, N.A.; Kolmakov, V.I.; Popelnitsky, V.A.; Gold, V.M.; Dubovskaya, O.P. Evaluation of the effect of light intensity on the measurement of the photosynthetic rate in plankton microalgae by the chlorophyll fluorescence method. Russ. J. Plant Physiol. 2000, 47, 820–825. [Google Scholar] [CrossRef]
- Ishimaru, T.; Mimuro, M.; Fujita, Y. Estimation of phytoplankton photosynthesis using a fluorescence induction technique. J. Plankton Res. 1985, 7, 679–689. [Google Scholar] [CrossRef]
- Alimov, A.F. An Introduction to Production Hydrobiology; Gidrometeoizdat: Leningrad, Russia, 1989; pp. 1–152. [Google Scholar]
- Stockwell, J.D.; Johansson, O.E. Temperature-dependent allometric models to estimate zooplankton production in temperate freshwater lakes. Can. J. Fish. Aquat. Sci. 1997, 54, 2350–2360. [Google Scholar] [CrossRef]
- Bottrell, H.H.; Duncan, A.; Gliwicz, Z.M.; Grygierek, E.; Herzig, A.; Hillbricht-Ilkowska, A.; Kurasawa, H.; Larsson, P.; Węgleńska, T. A review of some problems in zooplankton production studies. Nor. J. Zool. 1976, 24, 419–456. [Google Scholar]
- Ter Braak, C.J.F. Canonical correspondence analysis: A new eigenvector technique for multivariate direct gradient analysis. Ecology 1986, 67, 1167–1179. [Google Scholar] [CrossRef] [Green Version]
- Brooks, J.L.; Dodson, S.I. Predation, body size and composition of plankton. Science 1965, 150, 28–35. [Google Scholar] [CrossRef]
- Gliwicz, Z.M. Between Hazards of Starvation and Risk of Predation: The Ecology of Off-Shore Animals; International Ecology Institute: Oldendorf (Luhe), Germany, 2003; ISBN 978-3-946729-12-9. [Google Scholar]
- Sikora, A.; Dawidowicz, P. Do the presence of filamentous cyanobacteria and an elevated temperature favor small-bodied Daphnia in interspecific competitive interactions? Fundam. Appl. Limnol. 2014, 185, 307–314. [Google Scholar] [CrossRef]
- Henning, M.; Hertel, W.; Wall, H.; Kohl, J.G. Strain-specific influence of Microcystis aeruginosa on food ingestion and assimilation of some cladocerans and copepods. Int. Rev. Ges. Hydrobiol. 1991, 76, 37–45. [Google Scholar] [CrossRef]
- Mayer, J.; Dokulil, M.T.; Salbrechter, M.; Berger, M.; Posch, T.; Pfister, G.; Kirschner, A.K.T.; Velimorov, B.; Steitz, A.; Ulbricht, T. Seasonal succession and trophic relations between phytoplankton, zooplankton, ciliate and bacteria in a hypertrophic shallow lake in Vienna, Austria. Hydrobiologia 1997, 342–343, 165–174. [Google Scholar] [CrossRef]
- Cyr, H. Cladoceran- and copepod-dominated zooplankton communities graze at similar rates in low-productivity lakes. Can. J. Fish. Aquat. Sci. 1998, 55, 414–422. [Google Scholar] [CrossRef]
- Górniak, A.; Karpowicz, M. Development of crustacean plankton in a shallow, polyhumic reservoir in the first 20 years after impoundment (northeast Poland). Inland Waters 2014, 4, 311–318. [Google Scholar] [CrossRef]
- Hutorowicz, A.; Szeląg-Wasielewska, E.; Grabowska, M.; Owsianny, P.M.; Pęczuła, W.; Luscińska, M. The occurrence of Gonyostomum semen (Raphidophyceae) in Poland. Flor. Geobot. Pol. 2006, 13, 399–407. [Google Scholar]
- Pęczuła, W.; Grabowska, M.; Zieliński, P.; Karpowicz, M.; Danilczyk, M. Vertical distribution of expansive, bloom-forming algae Gonyostomum semen vs. plankton community and water chemistry in four small humic lakes. Knowl. Manag. Aquat. Ecosyst. 2018, 419, 28. [Google Scholar] [CrossRef]
- DeMott, W.R.; Kerfoot, W.C. Competition among cladocerans: Nature of the interactions between Bosmina and Daphnia. Ecology 1982, 6, 1949–1966. [Google Scholar] [CrossRef]
- Esparcia, A.; Miracle, M.R.; Serra, M. Brachionus plicatilis tolerance to low oxygen concentrations. Hydrobiologia 1989, 186, 331–337. [Google Scholar] [CrossRef]
- Dimante-Deimantovica, I.; Skute, A.; Skute, R. Vertical variability of pelagic zooplankton fauna in deep Latvian lakes, with some notes on changes in ecological conditions. Est. J. Ecol. 2012, 61, 247–264. [Google Scholar] [CrossRef] [Green Version]
- Snell, T.W.; Johnston, R.K.; Jones, B.L. Hypoxia extends lifespan of Brachionus manjavacas (Rotifera). Limnetica 2019, 38, 159–166. [Google Scholar] [CrossRef]
- Gilbert, J.J. Suppression of rotifer populations by Daphnia: A review of the evidence, the mechanisms, and the effects on zooplankton community structure. Limnol. Oceanogr. 1988, 33, 1286–1303. [Google Scholar] [CrossRef]
- Gilbert, J.J. Susceptibilities of ten rotifer species to interference from Daphnia pulex. Ecology 1988, 69, 1826–1838. [Google Scholar] [CrossRef]
- Kizito, Y.S.; Nauwerck, A. Temporal and vertical distribution of planktonic rotifers in a meromictic crater lake, Lake Nyahirya (Western Uganda). Hydrobiologia 1995, 312–314, 303–312. [Google Scholar] [CrossRef]
- Bertilsson, J.; Berzins, B.; Pejler, B. Occurrence of limnic micro-crustaceans in relation to temperature and oxygen. Hydrobiologia 1995, 299, 163–167. [Google Scholar] [CrossRef]
- Elliott, J.I. Seasonal changes in the abundance and distribution of planktonic rotifers in Grasmere (English Lake District). Freshw. Biol. 1977, 7, 147–166. [Google Scholar] [CrossRef]
- Czarnoleski, M.; Ejsmont-Karabin, J.; Angilletta, M.J., Jr.; Kozlowski, J. Colder rotifers grow larger but only in oxygenated waters. Ecosphere 2015, 6, 1–5. [Google Scholar] [CrossRef]
- Mikschi, E. Rotifer distribution in relation to temperature and oxygen content. Hydrobiologia 1989, 186–187, 209–214. [Google Scholar] [CrossRef]
- Lass, S.; Boersma, M.; Spaak, P. How do migrating daphnids cope with fish predation risk in the epilimnion under anoxic conditions in the hypolimnion? J. Plankton Res. 2000, 22, 1411–1418. [Google Scholar] [CrossRef]
- Kasprzak, P.; Reese, C.; Koschel, R.; Schulz, M.; Hambaryan, I.; Mathes, J. Habitat characteristics of Eurytemora lacustris (Poppe, 1887) (Copepoda, Calanoida): The role of lake depth, temperature, oxygen concentration and light intensity. Int. Rev. Hydrobiol. 2005, 90, 292–309. [Google Scholar] [CrossRef]
- Maier, G.; Speth, B.; Wolfgang, A.R.P.; Bahnwart, M.; Kasprzak, P. New records of the rare glacial relict Eurytemora lacustris (Poppe 1887) (Copepoda; Calanoida) in atypical lake habitats of northern Germany. J. Limnol. 2011, 70, 145–148. [Google Scholar] [CrossRef] [Green Version]
- Tinson, S.; Laybourn-Parry, J. The behavioural responses and tolerance of freshwater benthic cyclopoid copepods to hypoxia and anoxia. Hydrobiologia 1985, 127, 257–263. [Google Scholar] [CrossRef]
- Roman, M.R.; Gauzens, A.L.; Rhinehart, W.K.; White, J.R. Effects of low oxygen waters on Chesapeake Bay zooplankton. Limnol. Oceanogr. 1993, 38, 1603–1614. [Google Scholar] [CrossRef]
- Gliwicz, Z.M.; Pijanowska, J. Effects of predation and resource depth distribution on vertical migration of zooplankton. Bull. Mar. Sci. 1988, 43, 695–709. [Google Scholar]
- Loose, C.J.; Dawidowicz, P. Trade-offs in diel vertical migration by zooplankton: The costs of predator avoidance. Ecology 1994, 75, 2255–2263. [Google Scholar] [CrossRef]
- Pannard, A.; Planas, D.; Beisner, B.E. Macrozooplankton and the persistence of the deep chlorophyll maximum in a stratified lake. Freshw. Biol. 2015, 60, 1717–1733. [Google Scholar] [CrossRef]
- Sługocki, Ł.; Czerniawski, R. Trophic state (TSISD) and mixing type significantly influence pelagic zooplankton biodiversity in temperate lakes (NW Poland). PeerJ 2018, 6, e5731. [Google Scholar] [CrossRef] [Green Version]
- Nolan, S.; Bollens, S.M.; Rollwagen-Bollens, G. Diverse taxa of zooplankton inhabit hypoxic waters during both day and night in a temperate eutrophic lake. J. Plankton Res. 2019, 41, 431–447. [Google Scholar] [CrossRef]
- Roman, M.R.; Brandt, S.B.; Houde, E.D.; Pierson, J.J. Interactive Effects of Hypoxia and Temperature on Coastal Pelagic Zooplankton and Fish. Front. Mar. Sci. 2019, 6, 139. [Google Scholar] [CrossRef] [Green Version]
- Suthers, I.M.; Gee, J.H. Role of hypoxia in limiting summer distribution of juvenile yellow perch (Perca flavescens) in a prairie marsh. Can. J. Fish. Aquat. Sci. 1986, 43, 1562–1570. [Google Scholar] [CrossRef]
- Eby, L.A.; Crowder, L.B. Hypoxia-based habitat compression in the Neuse River Estuary: Context-dependent shifts in behavioral avoidance thresholds. Can. J. Fish. Aquat. Sci. 2002, 59, 952–965. [Google Scholar] [CrossRef]
- Rautio, M.; Tartarotti, B. UV radiation and freshwater zooplankton: Damage, protection and recovery. Freshw. Rev. 2010, 3, 105–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robidoux, M.; del Giorgio, P.; Derry, A. Effects of humic stress on the zooplankton from clear and DOC-rich lakes. Freshw. Biol. 2015, 60, 1263–1278. [Google Scholar] [CrossRef]
- Taipale, S.J.; Vuorio, K.; Strandberg, U.; Kahilainen, K.K.; Järvinen, M.; Hiltunen, M.; Peltomaa, E.; Kankaala, P. Lake eutrophication and brownification downgrade availability and transfer of essential fatty acids for human consumption. Environ. Int. 2016, 96, 156–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Podshivalina, V.N.; Sheveleva, N.G.; Bayanov, N. Biology and Ecology of Holopedium gibberum (Branchiopoda: Cladocera: Ctenopoda) in the Palearctic. Hydrobiol. J. 2012, 48, 28–36. [Google Scholar] [CrossRef]
- Kuczyńska-Kippen, N.; Klimaszyk, P.; Piotrowicz, R. Zooplankton communities in three adjacent softwater lobelia lakes of slightly differentiated morphology and trophic state. Limnol. Rev. 2017, 17, 207–214. [Google Scholar] [CrossRef]
- Ejsmont-Karabin, J. Rotifer invasion? On appearance and abundance of tropical species in lakes of North-Eastern Poland. Pol. J. Ecol. 2014, 62, 727–733. [Google Scholar] [CrossRef]
- Brzeziński, T.; Fronk, J.; Trzcińska-Danielewicz, J.; Dawidowicz, P. Interspecific hybridization in sympatric species of Daphnia inhabiting lakes in north-eastern Poland. Oceanol. Hydrobiol. Stud. 2012, 41, 1–6. [Google Scholar] [CrossRef]
- Sługocki, Ł.; Rymaszewska, A.; Kirczuk, L. Insights into the morphology and molecular characterization of glacial relict Eurytemora lacustris (Poppe, 1887) (Crustacea, Copepoda, Calanoida, Temoridae). ZooKeys 2019, 864, 15–33. [Google Scholar] [CrossRef]
- Ejsmont-Karabin, J.; Kalinowska, K.; Karpowicz, M. Structure of Ciliate, Rotifer, and Crustacean Communities in Lake Systems of Northeastern Poland. In Polish River Basins and Lakes—Part II; Springer: Cham, Switzerland, 2020; Volume 87, pp. 77–101. [Google Scholar] [CrossRef]
- Rybak, J.I.; Błędzki, L.A. Słodkowodne Skorupiaki Planktonowe. Klucz do Oznaczania Gatunków; Wydawnictwa Uniwersytetu: Warszawskiego, Poland, 2010; (In Polish). [Google Scholar] [CrossRef]
- Litvinchuk, L.F.; Litvinchuk, S.N. Morphological diversity and widespread hybridization in the genus Bythotrephes Leydig, 1860 (Branchiopoda, Onychopoda, Cercopagidae). Arch. Biol. Sci. 2016, 68, 67–79. [Google Scholar] [CrossRef]
- Korovchinsky, N.M. Further revision of the genus Bythotrephes Leydig (Crustacea: Cladocera: Onychopoda): Redescription of B. brevimanus Lilljeborg, reevaluation of B. cederströmii Schödler, and description of a new species of the genus. Zootaxa 2018, 4379, 347–387. [Google Scholar] [CrossRef]
- Korovchinsky, N.M. Morphological assessment of the North Eurasian interspecific hybrid forms of the genus Bythotrephes Leydig, 1860 (Crustacea: Cladocera: Cercopagididae). Zootaxa 2019, 4550, 340–356. [Google Scholar] [CrossRef] [PubMed]
- Pauly, D.; Christensen, V. Primary production required to sustain global fisheries. Nature 1995, 374, 255–257. [Google Scholar] [CrossRef]
- Schulz, M.; Koschel, R.; Reese, C.; Mehner, T. Pelagic trophic transfer efficiency in an oligotrophic, dimictic deep lake (Lake Stechlin, Germany) and its relation to fisheries yield. Limnologica 2004, 34, 264–273. [Google Scholar] [CrossRef] [Green Version]
- Lindeman, R.L. The trophic–dynamic aspect of ecology. Ecology 1942, 23, 399–418. [Google Scholar] [CrossRef]
- Gladyshev, M.I.; Sushchik, N.N.; Anishchenko, O.V.; Makhutova, O.N.; Kolmakov, V.I.; Kalachova, G.S.; Kolmakova, A.A.; Dubovskaya, O.P. Efficiency of transfer of essential polyunsaturated fatty acids versus organic carbon from producers to consumers in a eutrophic reservoir. Oecologia 2011, 165, 521–531. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, G.; Lescher-Moutoue, F.; Bertolo, A. Biomass and production of plankton in shallow and deep lakes: Are there general patterns? Ann. Limnol. Int. J. Limnol. 1999, 35, 111–122. [Google Scholar] [CrossRef] [Green Version]
- Sherr, E.B.; Sherr, B.F. High rates of consumption of bacteria by pelagic ciliates. Nature 1987, 325, 710–711. [Google Scholar] [CrossRef]
- Havens, K.E.; Work, K.A.; East, T.L. Relative efficiencies of carbon transfer from bacteria and algae to zooplankton in a subtropical lake. J. Plankton Res. 2000, 22, 1801–1809. [Google Scholar] [CrossRef] [Green Version]
- Ducklow, H.W.; Purdie, D.A.; Williams, P.J.L.; Davies, J.M. Bacterioplankton: A sink for carbon in a coastal marine plankton community. Science 1986, 232, 865–867. [Google Scholar] [CrossRef]
- Koshikawa, H.; Shigeki, H.; Watanabe, M.; Sato, K.; Akehatu, K. Relative contribution of bacterial and photosynthetic production to metazooplankton as carbon sources. J. Plankton Res. 1996, 18, 2269–2281. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Lakes | Surface (ha) | Max Depth (m) | Avg. Depth (m) | Meta Depth (m) | SDV (m) | Max chl a (µg L−1) | TSI | HDI | Trophic Status | Oxygen Group |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Białe Filipowskie | 132.4 | 52 | 17.1 | 7–10 | 7.5 | 2.6 | 29.6 | 35.1 | oligo | G1 |
| 2 | Gaładuś | 728.6 | 54.8 | 12.7 | 7–10 | 5.7 | 1.6 | 33.8 | 35.3 | oligo | G1 |
| 3 | Serwy | 460.3 | 41.5 | 14.6 | 6–10 | 3.8 | 5.6 | 38.5 | 35 | oligo | G1 |
| 4 | Leleskie | 423.5 | 49.5 | 12.2 | 7–10 | 5.5 | 3.5 | 39.7 | 37.3 | oligo | G1 |
| 5 | Jegocin | 127.4 | 36.1 | 9 | 6.5–9 | 7 | 32.7 | 39.7 | 37.7 | oligo | G2 |
| 6 | Buwełno | 360.3 | 49.1 | 12.4 | 7.5–10 | 2.4 | 7.3 | 42.4 | 30.7 | meso | G1 |
| 7 | Majcz Wielki | 163.5 | 16.4 | 6 | 6–9 | 2.7 | 4.1 | 42.8 | 34.1 | meso | G2 |
| 8 | Kuc | 98.8 | 28 | 8 | 6–9.5 | 4 | 8.8 | 43.5 | 34.7 | meso | G1 |
| 9 | Białe Wigierskie | 100.2 | 34 | 13.2 | 6.5–11 | 5.7 | 2.3 | 44.3 | 33.3 | meso | G1 |
| 10 | Busznica | 49.4 | 48 | 6.8 | 4–10 | 6.6 | 4.8 | 44.3 | 34.8 | meso | G1 |
| 11 | Wigry | 2118.3 | 73 | 15.8 | 7.5–10 | 5.2 | 3.8 | 45.4 | 30.9 | meso | G1 |
| 12 | Probarskie | 201.4 | 31 | 9.2 | 6.5–9.5 | 3.8 | 7.6 | 46.3 | 33.6 | meso | G1 |
| 13 | Okrągłe | 13.7 | 13 | 5.6 | 3.5–8 | 3.1 | 6.8 | 49.7 | 31.4 | meso | G3 |
| 14 | Brzozolasek | 155.9 | 17.2 | 5.1 | 6–9 | 1.2 | 6.4 | 49.4 | 36 | meso | G3 |
| 15 | Mikołajskie | 497.9 | 25.9 | 11.2 | 7–10 | 2 | 10.8 | 50 | 34.8 | eu | G2 |
| 16 | Kalwa | 562.2 | 31.7 | 7 | 5–8 | 2 | 26 | 50 | 34.8 | eu | G3 |
| 17 | Jagodne | 942.7 | 37.4 | 8.7 | 6.5–9 | 2.2 | 11.9 | 50.3 | 29.6 | eu | G2 |
| 18 | Boczne | 183.3 | 17 | 8.7 | 7–9.5 | 1.4 | 11.4 | 51.1 | 32.2 | eu | G2 |
| 19 | Wiartel | 178.6 | 29 | 4.5 | 7–10 | 1.6 | 15.6 | 51.7 | 37.7 | eu | G3 |
| 20 | Ryńskie | 670.8 | 50.8 | 10 | 6–9 | 1 | 9 | 52.5 | 34.3 | eu | G3 |
| 21 | Kierźlińskie | 92.8 | 44.5 | 11.7 | 6–9 | 1.5 | 23.5 | 53.1 | 32.7 | eu | G2 |
| 22 | Długie Wigierskie | 80 | 14.8 | 6.4 | 4–8 | 1.65 | 26.8 | 53.5 | 30.3 | eu | G3 |
| 23 | Leszczewek | 21 | 6.5 | 3.6 | >3 | 2.3 | 97.8 | 54.4 | 32.1 | eu | G4 |
| 24 | Nidzkie | 1818 | 23.7 | 6.2 | 6–9 | 0.9 | 34.4 | 55.4 | 35.4 | eu | G4 |
| 25 | Garbaś | 152.5 | 48 | 20.9 | 4–9 | 2 | 34.5 | 58 | 30.1 | eu | G1 |
| 26 | Miłkowskie | 23.7 | 15 | 4.2 | 5–7 | 0.5 | 73.9 | 59.4 | 32.3 | eu | G4 |
| 27 | Juno | 380.7 | 33 | 11.9 | 6–9 | 0.7 | 33.7 | 62.9 | 32.7 | eu | G4 |
| 28 | Klimunt | 11.8 | 5.5 | 2.7 | >1.5 | 0.3 | 85.9 | 63.8 | 60.4 | dy | G6 |
| 29 | Wesołek | 7 | 3 | nd. | >1.5 | 1.2 | 18.7 | 50.7 | 66.9 | dy | G5 |
| 30 | Zdrużno | 6.8 | 5 | nd. | >2.5 | 1.8 | 94.6 | 51.2 | 69.8 | dy | G5 |
| 31 | Gryżlewskie | 4.3 | 5 | nd. | >2 | 1.8 | 65.2 | 50.7 | 71.4 | dy | G5 |
| 32 | Sęczek | 3.8 | 3.5 | nd. | >2 | 0.9 | 118.7 | 56 | 72.2 | dy | G6 |
| 33 | Borkowskie | 2.9 | 5 | nd. | 2–5 | 1.2 | 47.3 | 53.7 | 73.2 | dy | G5 |
| 34 | Kruczy Staw | 2.1 | 8 | nd. | 2–5 | 2 | 36 | 47.7 | 73.3 | dy | G5 |
| 35 | Kruczek | 4.2 | 4 | nd. | >2 | 1.5 | 35.3 | 51.3 | 74.2 | dy | G5 |
| 36 | Kruczek Mały | 2.6 | 9 | nd. | 1–5 | 1.7 | 134 | 46.7 | 74.6 | dy | G5 |
| 37 | Konopniak | 9.5 | 4 | nd. | >1.5 | 1.2 | 64.5 | 50.4 | 79.9 | dy | G6 |
| 38 | Wądołek | 1.09 | 15 | 8.9 | >1 | 1.2 | 61.9 | 70 | 68.6 | dy | G5 |
| 39 | SI | 0.96 | 4 | 2.3 | >1 | 1.1 | 135 | 62 | 66.5 | dy | G6 |
| 40 | SII | 2.52 | 10 | 4.6 | 1.5–5.5 | 1.9 | 52.8 | 53.7 | 69.4 | dy | G5 |
| 41 | Suchar Wielki | 8.9 | 9.6 | 3.9 | 3.5–6 | 2.2 | 58.5 | 56.7 | 68.2 | dy | G5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karpowicz, M.; Ejsmont-Karabin, J.; Kozłowska, J.; Feniova, I.; Dzialowski, A.R. Zooplankton Community Responses to Oxygen Stress. Water 2020, 12, 706. https://doi.org/10.3390/w12030706
Karpowicz M, Ejsmont-Karabin J, Kozłowska J, Feniova I, Dzialowski AR. Zooplankton Community Responses to Oxygen Stress. Water. 2020; 12(3):706. https://doi.org/10.3390/w12030706
Chicago/Turabian StyleKarpowicz, Maciej, Jolanta Ejsmont-Karabin, Joanna Kozłowska, Irina Feniova, and Andrew R. Dzialowski. 2020. "Zooplankton Community Responses to Oxygen Stress" Water 12, no. 3: 706. https://doi.org/10.3390/w12030706