Effects of Cyanobacteria on Phosphorus Cycling and Other Aquatic Organisms in Simulated Eutrophic Ecosystems

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Physical, Chemical, and Biological Analyses
2.2.1. Water Quality
2.2.2. Phosphorus Contents
2.2.3. Algal Cell Density
2.2.4. Alkaline Phosphatase Activity
2.2.5. Microcystin Content
2.2.6. Microbial Community Analysis
2.3. Statistical Analysis
3. Results and Discussion
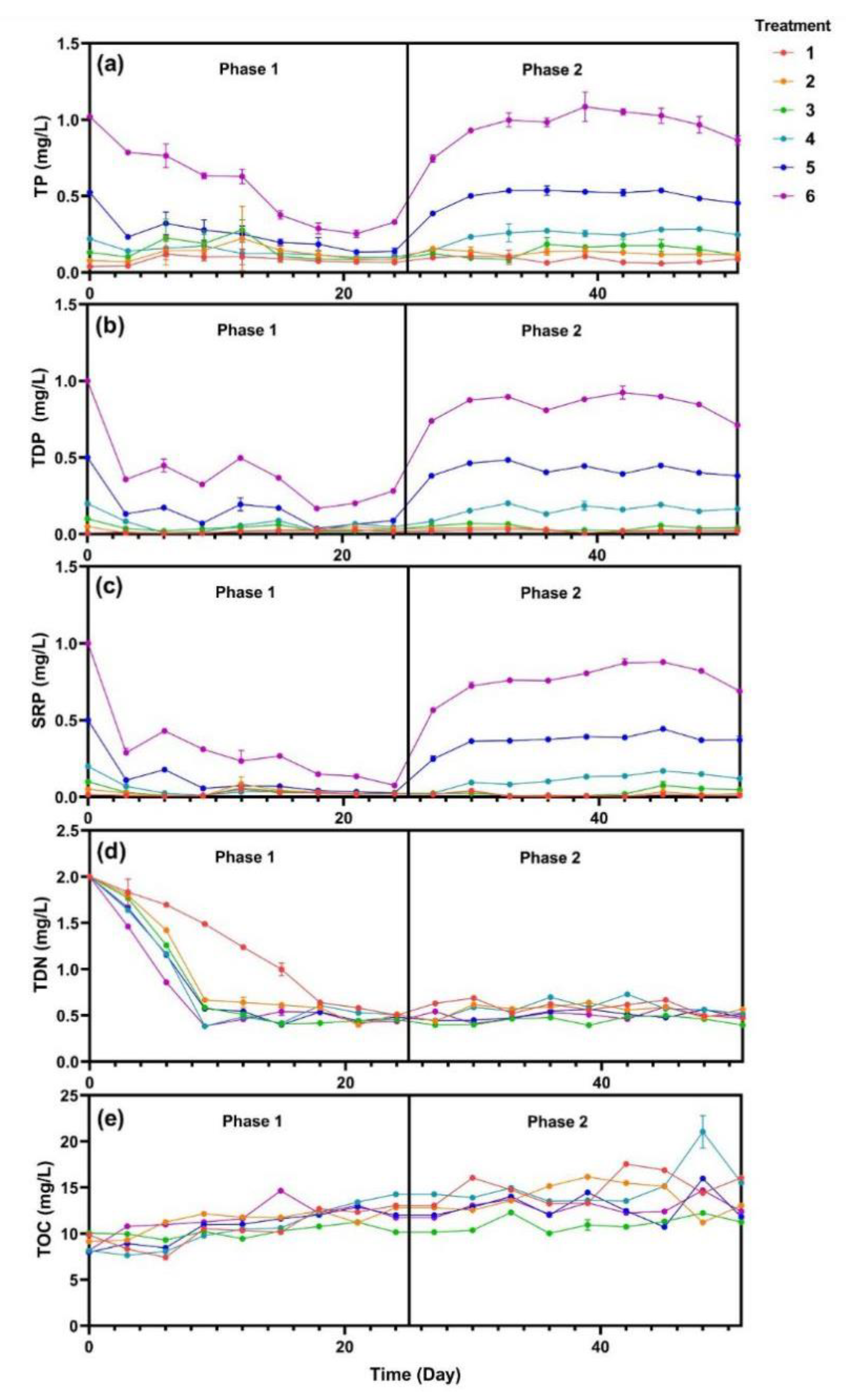
3.1. Water Quality Characterization
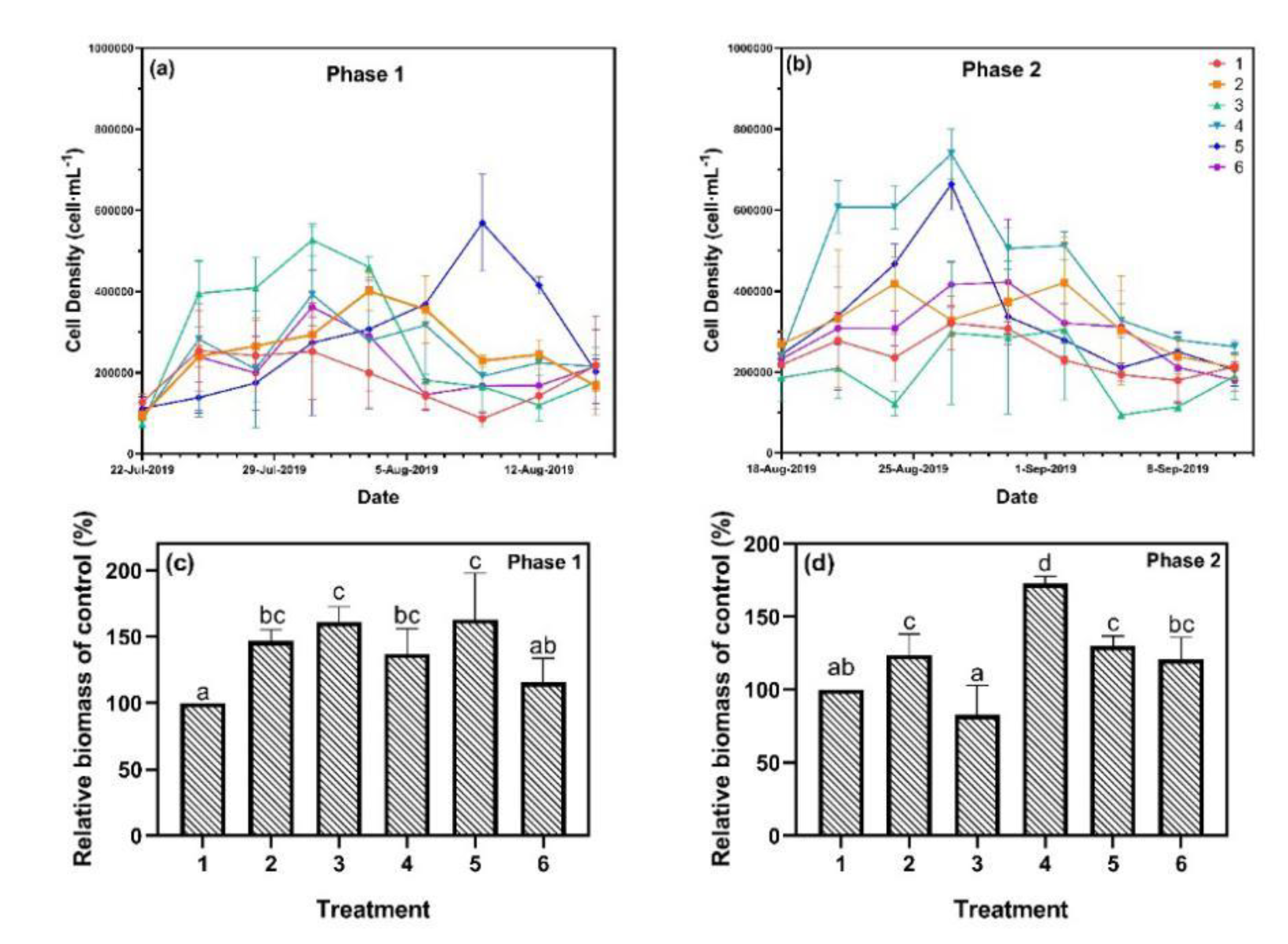
3.2. Algal Growth Analysis
3.3. Alkaline Phosphatase Activity and P Cycling
3.3.1. Alkaline Phosphatase (ALP) Activity
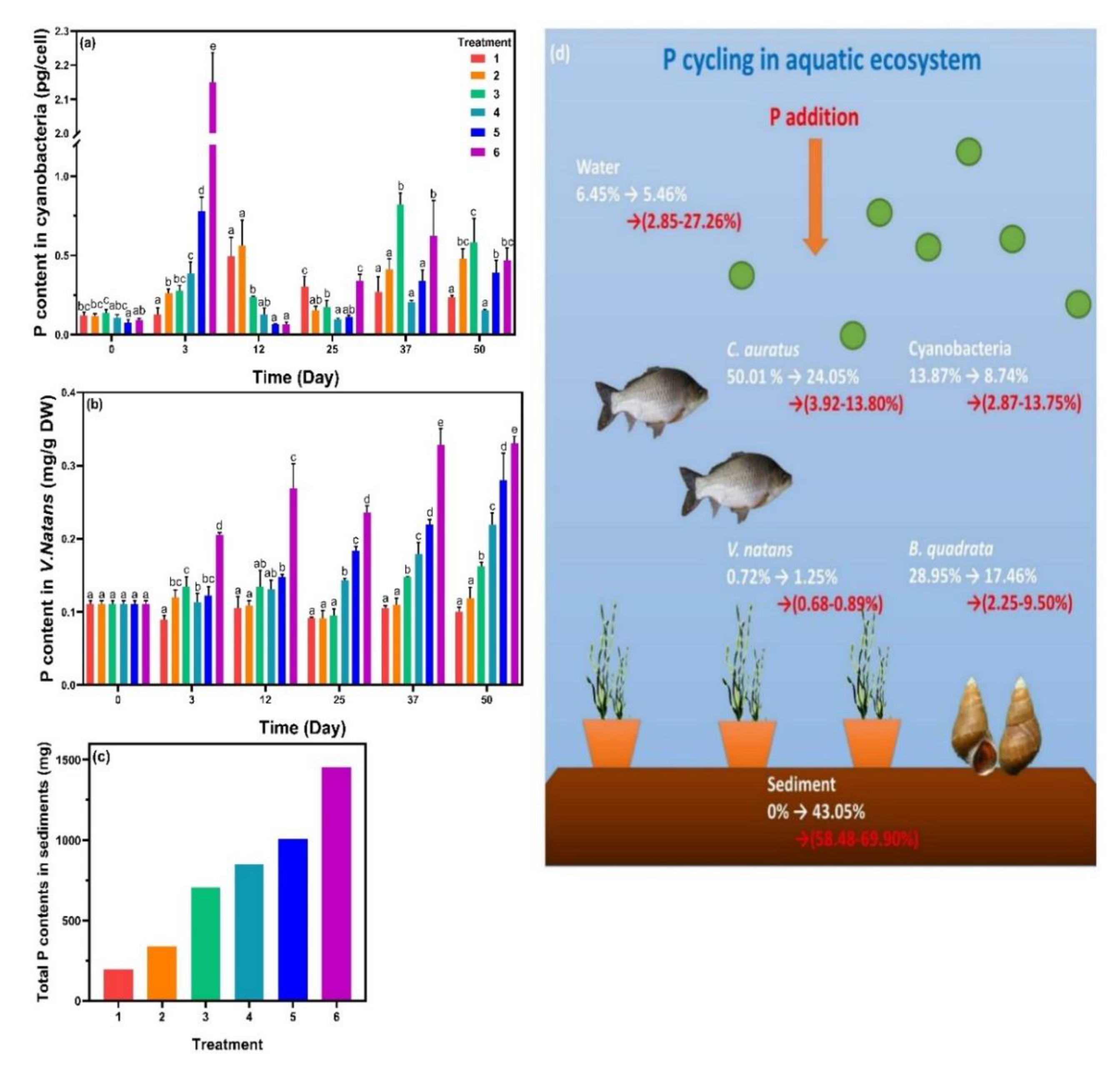
3.3.2. P Cycling
3.4. MC-LR Release and Effects on Other Organisms
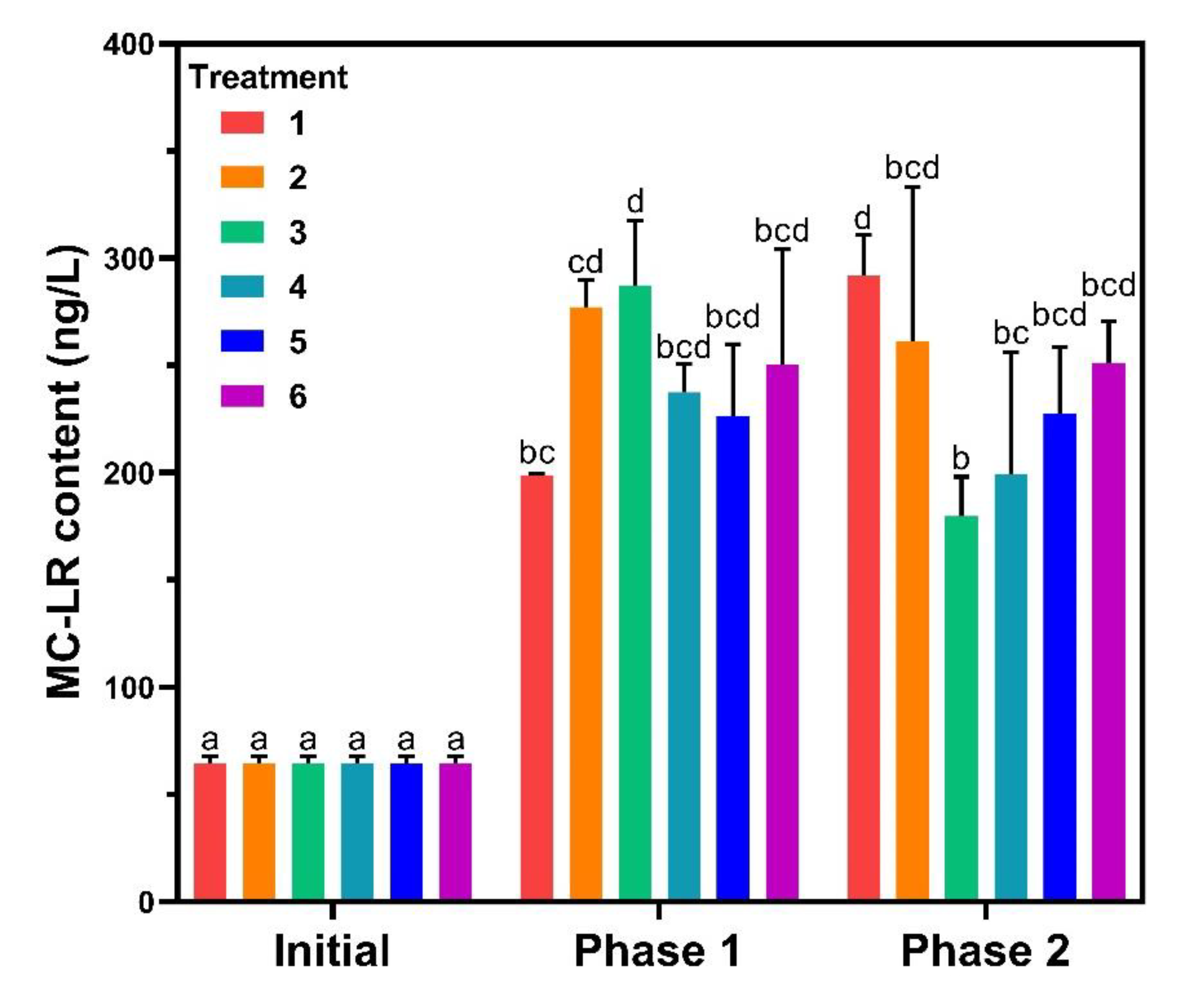
3.4.1. MC-LR Content
3.4.2. Response of Aquatic Organisms
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Li, W.; Yang, Q. Wetland utilization in Lake Taihu for fish farming and improvement of lake water quality. Ecol. Eng. 1995, 5, 107–121. [Google Scholar] [CrossRef]
- He, D.; Ren, L.; Wu, Q. Epiphytic bacterial communities on two common submerged macrophytes in Taihu Lake: Diversity and host-specificity. Chin. J. Oceanol. Limnol. 2012, 30, 237–247. [Google Scholar] [CrossRef]
- Chen, Q.; Zhang, X.; Shen, L.; Zhang, X.; Dongjiong, X.U. Community structure and species diversity of benthic macroinvertebrates in Taihu Basin of Jiangsu Province. J. Lake Sci. 2017, 29, 1398–1411. [Google Scholar]
- Xu, H.; Paerl, H.W.; Qin, B.; Zhu, G.; Hall, N.S.; Wu, Y. Determining critical nutrient thresholds needed to control harmful cyanobacterial blooms in eutrophic Lake Taihu, China. Environ. Sci. Technol. 2015, 49, 1051–1059. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.W.; Xiao-Guang, X.U.; Han, R.M.; Zhou, X.H.; Feng, D.Y.; Zhi-Chun, L.I.; Wang, G.X. Effect of nutrient loadings on the regulation of water nitrogen and phosphorus by Vallisneria natans and its photosynthetic fluorescence characteristics. Environ. Sci. 2018, 39, 1180. [Google Scholar]
- Xu, H.; Yang, G.; Zhou, J.; Qin, B.; Zhang, G.; Zou, H.; Hu, X. Effect of nitrogen and phosphorus concentration on colony growth of Microcystis flosaquae in Lake Taihu. J. Lake Sci. 2014, 26, 213–220. [Google Scholar]
- Dhote, S.; Dixit, S. Water quality improvement through macrophytes—A review. Environ. Monit. Assess. 2009, 152, 149–153. [Google Scholar] [CrossRef]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.H.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef]
- Merel, S.; Walker, D.; Chicana, R.; Snyder, S.; Thomas, O. State of knowledge and concerns on cyanobacterial blooms and cyanotoxins. Environ. Int. 2013, 59, 303–327. [Google Scholar] [CrossRef]
- Dawson, R.M. The toxicology of microcystins. Toxicon 1998, 36, 953–962. [Google Scholar] [CrossRef]
- Magalhães, V.F.D.; Soares, R.M.; Azevedo, S.M.F.O. Microcystin contamination in fish from the Jacarepagua Lagoon (Rio de Janeiro, Brazil): Ecological implication and human health risk. Toxicon 2001, 39, 1077–1085. [Google Scholar] [CrossRef]
- Yin, L.Y.; Huang, J.Q.; Li, D.H.; Hu, Z.Q.; Liu, Y. The effect of microcystin on the growth and development of submergent macrophyte Vallisneria natans Hara. Acta Hydrobiol. Sin. 2004, 28, 2115–2124. [Google Scholar]
- APHA; AWWA; WEF. Standard Methods for Examination of Water and Wastewater, 22nd ed.; American Public Health Association: Washington, DC, USA, 2012. [Google Scholar]
- Ebina, J.; Tsutsui, T.; Shirai, T. Simultaneous determination of total nitrogen and total phosphorus in water using peroxodisulfate oxidation. Water Res. 1983, 17, 1721–1726. [Google Scholar] [CrossRef]
- Griffiths, D. The direct contribution of fish to lake phosphorus cycles. Ecol. Freshw. Fish 2006, 15, 86–95. [Google Scholar] [CrossRef]
- Schwartz, M.K.; Kessler, G.; Bodansky, O. Comparison of serum alkaline phosphatase activities determined with sodium β-glycerophosphate and sodium phenylphosphate as substrates. Am. J. Clin. Pathol. 1960, 33, 275–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, P.; Hegde, K.; Brar, S.K.; Cledon, M.; Kermanshahi-Pour, A.; Roy-Lachapelle, A.; Galvez-Cloutier, R. Biodegradation of microcystin-LR using acclimatized bacteria isolated from different units of the drinking water treatment plant. Environ. Pollut. 2018, 242, 407–416. [Google Scholar] [CrossRef] [Green Version]
- Kerschbaumer, R.J.; Hirschl, S.; Kaufmann, A.; Ibl, M.; Koenig, R.; Himmler, G. Single-Chain Fv Fusion Proteins Suitable as Coating and Detecting Reagents in a Double Antibody Sandwich Enzyme-Linked Immunosorbent Assay. Anal. Biochem. 1997, 249, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Kemp, P.F.; Aller, J.Y. Bacterial diversity in aquatic and other environments: What 16S rDNA libraries can tell us. FEMS Microbiol. Ecol. 2004, 47, 161–177. [Google Scholar] [CrossRef]
- Fadrosh, D.W.; Ma, B.; Gajer, P.; Sengamalay, N.; Ott, S.; Brotman, R.M.; Ravel, J. An improved dual-indexing approach for multiplexed 16S rRNA gene sequencing on the Illumina MiSeq platform. Microbiome 2014, 2, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Schindler, D.W. Eutrophication and recovery in experimental lakes: Implications for lake management. Science 1974, 184, 897–899. [Google Scholar] [CrossRef] [Green Version]
- Conley, D.J.; Paerl, H.W.; Howarth, R.W.; Boesch, D.F.; Seitzinger, S.P.; Havens, K.E.; Lancelot, C.; Likens, G.E. Controlling eutrophication: Nitrogen and phosphorus. Science 2009, 323, 1014–1015. [Google Scholar] [CrossRef]
- Paerl, H.W.; Xu, H.; Hall, N.S.; Rossignol, K.L.; Joyner, A.R.; Zhu, G.; Qin, B. Nutrient limitation dynamics examined on a multi-annual scale in Lake Taihu, China: Implications for controlling eutrophication and harmful algal blooms. J. Freshw. Ecol. 2014, 30, 5–24. [Google Scholar] [CrossRef]
- Xu, H.; Paerl, H.W.; Qin, B.; Zhu, G.; Gaoa, G. Nitrogen and phosphorus inputs control phytoplankton growth in eutrophic Lake Taihu, China. Limnol. Oceanogr. 2010, 55, 420–432. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Zhu, G.; Qin, B.; Paerl, H.W. Growth response of Microcystis spp. to iron enrichment in different regions of Lake Taihu 2012, China. Hydrobiologia 2012, 700, 187–202. [Google Scholar] [CrossRef]
- Paerl, H.W.; Scott, J.T.; McCarthy, M.J.; Newell, S.E.; Gardner, W.S.; Havens, K.E.; Hoffman, D.K.; Wilhelm, S.W.; Wurtsbaugh, W.A. It takes two to tango: When and where dual nutrient (N & P) reductions are needed to protect lakes and downstream ecosystems. Environ. Sci. Technol. 2016, 50, 10805–10813. [Google Scholar]
- Paerl, H.W.; Xu, H.; McCarthy, M.J.; Zhu, G.; Qin, B.; Li, Y.; Gardner, W.S. Controlling harmful cyanobacterial blooms in a hyper-eutrophic lake (Lake Taihu, China): The need for a dual nutrient (N & P) management strategy. Water Res. 2011, 45, 1973–1983. [Google Scholar]
- Wen, R.; Sui, Z.; Bao, Z.; Zhou, W.; Wang, C. Isolation and characterization of calmodulin gene of Alexandrium catenella (Dinoflagellate) and its performance in cell growth and heat stress. J. Ocean Univ. China 2014, 13, 290–296. [Google Scholar] [CrossRef]
- Zhang, N.; Li, G.; Yu, J.; Ding, M.; Xu, L. Character of blue-green algal blooms outbreak in Taihu Lake. Environ. Monit. China 2009, 25, 71–74. [Google Scholar]
- Wang, S.; Xiao, J.; Wan, L.; Zhou, Z.; Wang, Z.; Song, C.; Zhou, Y.; Cao, X. Mutual dependence of nitrogen and phosphorus as key nutrient elements: One facilitates Dolichospermum flos-aquae to overcome the limitations of the other. Environ. Sci. Technol. 2018, 52, 5653–5661. [Google Scholar] [CrossRef] [PubMed]
- Harke, M.J.; Berry, D.L.; Ammerman, J.W.; Gobler, C.J. Molecular response of the bloom-forming cyanobacterium, Microcystis aeruginosa, to phosphorus limitation. Microb. Ecol. 2012, 63, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Chróst, R.J.; Overbeck, J. Kinetics of alkaline phosphatase activity and phosphorus availability for phytoplankton and bacterioplankton in lake pluβsee (North German Eutrophic Lake). Microb. Ecol. 1987, 13, 229–248. [Google Scholar] [CrossRef]
- Mazard, S.; Wilson, W.H.; Scanlan, D.J. Dissecting the physiological response to phosphorus stress in marine synechococcus isolates (Cyanophyceae). J. Phycol. 2012, 48, 94–105. [Google Scholar] [CrossRef]
- Baldia, S.F.; Evangelista, A.D.; Aralar, E.V.; Santiago, A.E. Nitrogen and phosphorus utilization in the cyanobacterium Microcystis aeruginosa isolated from Laguna de Bay, Philippines. J. Appl. Phycol. 2007, 19, 607–613. [Google Scholar] [CrossRef]
- Gras, A.F.; Koch, M.S.; Madden, C.J. Phosphorus uptake kinetics of a dominant tropical seagrass Thalassia testudinum. Aquat. Bot. 2003, 76, 299–315. [Google Scholar] [CrossRef]
- Barnabas, A.D. Thalassodendron ciliatum (Forssk.) Den Hartog: Root structure and histochemistry in relation to apoplastic transport. Aquat. Bot. 1991, 40, 129–143. [Google Scholar] [CrossRef]
- Spears, B.M.; Carvalho, L.; Perkins, R.; Kirika, A.; Paterson, D.M. Sediment phosphorus cycling in a large shallow lake: Spatio-temporal variation in phosphorus pools and release. Hydrobiologia 2007, 584, 37–48. [Google Scholar] [CrossRef]
- Cao, X.; Wang, Y.; He, J.; Luo, X.; Zheng, Z. Phosphorus mobility among sediments, water and cyanobacteria enhanced by cyanobacteria blooms in eutrophic Lake Dianchi. Environ. Pollut. 2016, 219, 580–587. [Google Scholar] [CrossRef]
- Watanabe, M.F.; Tsuji, K.; Watanabe, Y.; Harada, K.I.; Suzuki, M. Release of heptapeptide toxin (microcystin) during the decomposition process of Microcystis aeruginosa. Nat. Toxins 2006, 1, 48–53. [Google Scholar] [CrossRef]
- Tang, X.; Krausfeldt, L.; Shao, K.; LeCleir, G.; Stough, J.; Gao, G.; Boyer, G.; Zhang, Y.; Paerl, H.; Qin, B.; et al. Seasonal gene expression and the ecophysiological implications of toxic Microcystis aeruginosa blooms in Lake Taihu. Environ. Sci. Technol. 2018, 52, 11049–11059. [Google Scholar] [CrossRef]
- Yu, G.; Jiang, Y.; Song, G.; Tan, W.; Zhu, M.; Li, R.; Yu, G.; Jiang, Y.; Song, G.; Tan, W. Variation of Microcystis and microcystins coupling nitrogen and phosphorus nutrients in Lake Erhai, a drinking-water source in Southwest Plateau, China. Environ. Sci. Pollut. Res. Int. 2014, 21, 9887–9898. [Google Scholar] [CrossRef]
- Otten, T.G.; Xu, H.; Qin, B.; Zhu, G.; Paerl, H.W. Spatiotemporal Patterns and Ecophysiology of Toxigenic Microcystis Blooms in Lake Taihu, China: Implications for Water Quality Management. Environ. Sci. Technol. 2012, 46, 3480–3488. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Q.; Liang, H.; Zhang, X. Effect of cyanobacteria on immune function of crucian carp (Carassius auratus) via chronic exposure in diet. Chemosphere 2013, 90, 1167–1176. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Yu, N.; Jiang, X.; Han, Z.; Wang, S.; Zhang, X.; Wei, S.; Giesy, J.P.; Yu, H. Influence of blooms of phytoplankton on concentrations of hydrophobic organic chemicals in sediments and snails in a hyper-eutrophic, freshwater lake. Water Res. 2017, 113, 22–31. [Google Scholar] [PubMed]
- Song, Y.-Z.; Kong, F.-F.; Xue, Y.; Qin, B.-Q. Responses of chlorophyll and MDA of Vallisneria natans to nitrogen and phosphorus availability and epiphytic algae. J. Freshw. Ecol. 2014, 30, 85–97. [Google Scholar] [CrossRef]
- Qiao, F.; Lei, K.; Han, X.; Wei, Z.; Zhao, X.; An, L.; LeBlanc, G.A. No impacts of microcystins on wild freshwater snail Bellamya aeruginosa fecundity from a eutrophic lake. Environ. Toxicol. Pharm. 2018, 60, 165–168. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Gu, P.; Zhang, H.; Luo, X.; Zhang, J.; Zheng, Z. Response of submerged macrophytes and leaf biofilms to the decline phase of Microcystis aeruginosa: Antioxidant response, ultrastructure, microbial properties, and potential mechanism. Sci. Total Environ. 2019, 699, 134325. [Google Scholar] [CrossRef] [PubMed]
- Liu, E.-S. A study on diet composition of dominant fishes in Lake Taihu. J. Fish China 2008, 32, 395–401. [Google Scholar]
- He, H.; He, Y.; Ji, Y.; Guo, L.; Li, K. Effects of cyanobacterial accumulation and snail grazing on the growth of Vallisneria natans. Acta Ecol. Sin. 2012, 32, 5562–5567. [Google Scholar] [CrossRef] [Green Version]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, P.; Li, Q.; Zhang, H.; Luo, X.; Zhang, W.; Zheng, Z.; Luo, X. Effects of Cyanobacteria on Phosphorus Cycling and Other Aquatic Organisms in Simulated Eutrophic Ecosystems. Water 2020, 12, 2265. https://doi.org/10.3390/w12082265
Gu P, Li Q, Zhang H, Luo X, Zhang W, Zheng Z, Luo X. Effects of Cyanobacteria on Phosphorus Cycling and Other Aquatic Organisms in Simulated Eutrophic Ecosystems. Water. 2020; 12(8):2265. https://doi.org/10.3390/w12082265
Chicago/Turabian StyleGu, Peng, Qi Li, Hao Zhang, Xin Luo, Weizhen Zhang, Zheng Zheng, and Xingzhang Luo. 2020. "Effects of Cyanobacteria on Phosphorus Cycling and Other Aquatic Organisms in Simulated Eutrophic Ecosystems" Water 12, no. 8: 2265. https://doi.org/10.3390/w12082265