
Physiological Activity, Nutritional Composition, and Gene Expression in Apple (Malus domestica Borkh) Influenced by Different ETc Levels of Irrigation at Distinct Development Stages


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Material
2.2. Location and Weather
2.3. Leaf Physiological Characteristics
2.4. Leaf Nutrient Status
2.5. Gene Expression Studies
2.5.1. RNA Extraction and cDNA Synthesis
2.5.2. RT-qPCR Analysis
2.6. Statistical Analysis
3. Results
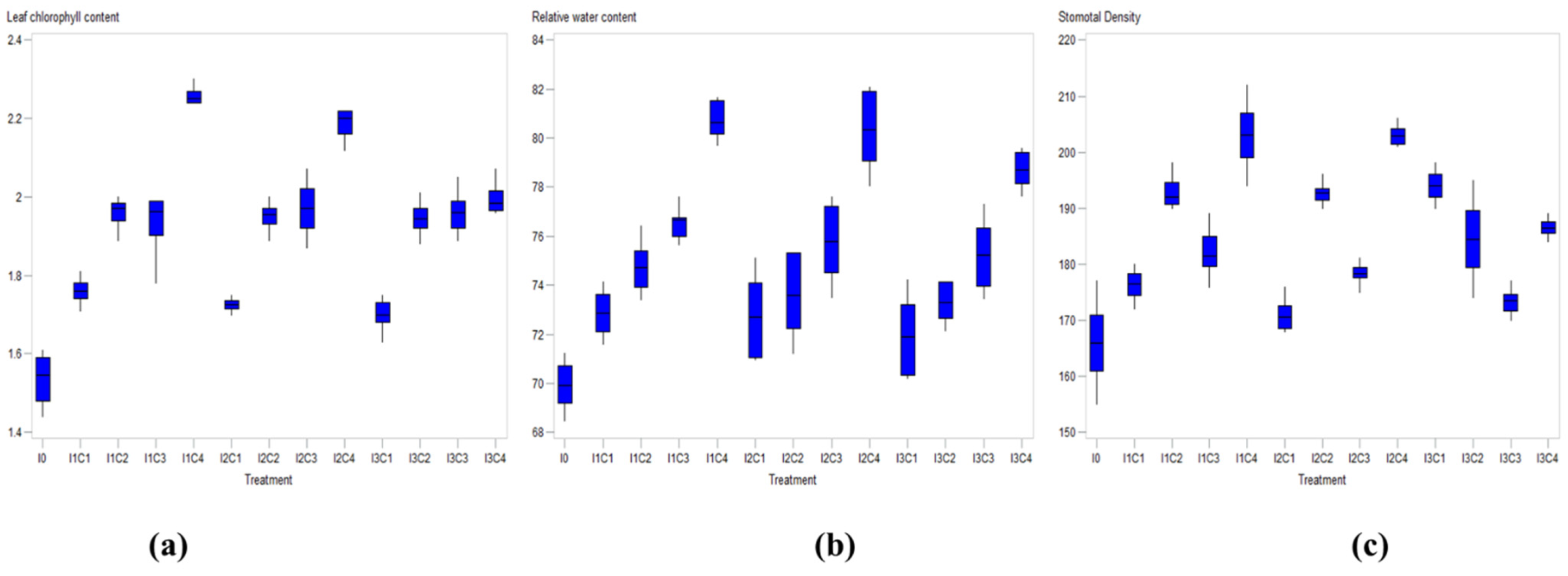
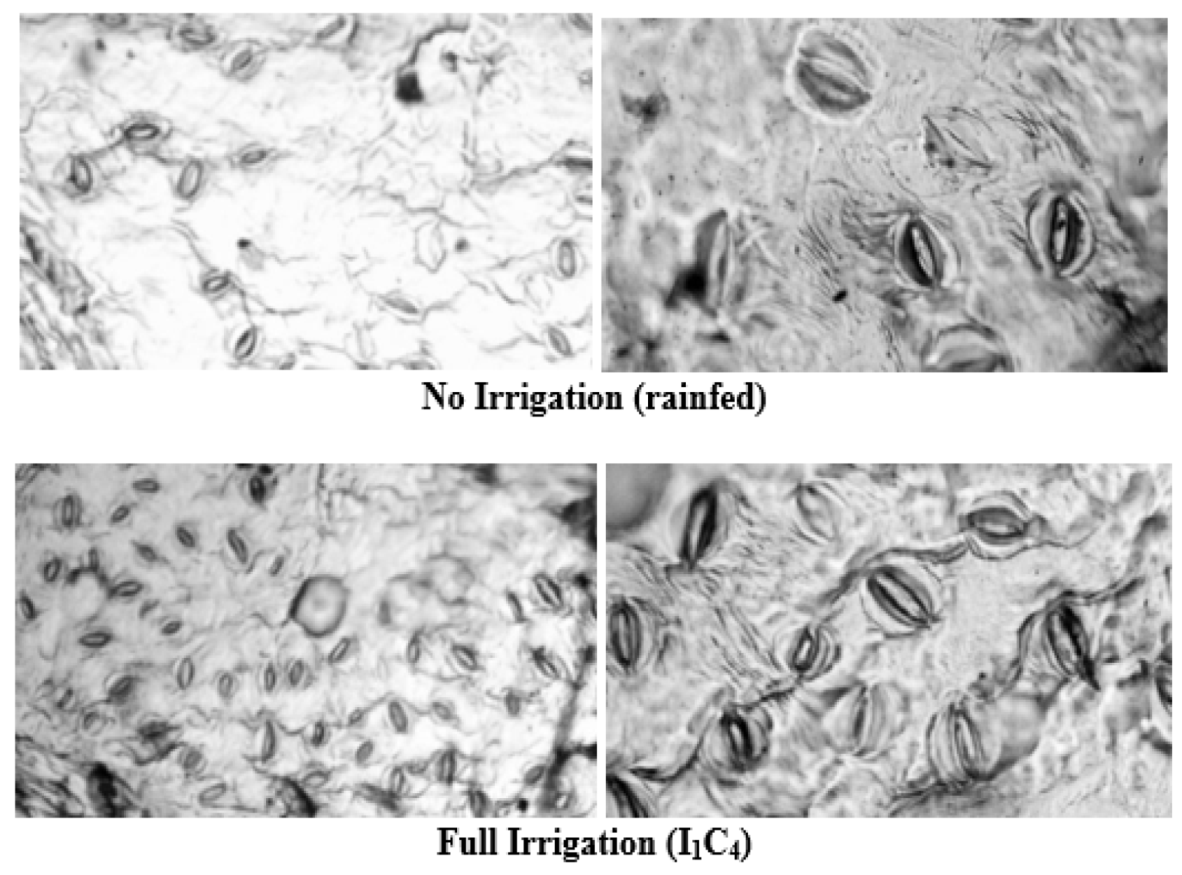
3.1. Leaf Physiological Characteristics
3.2. Leaf Nutrient Status
3.3. Expression of Nutrient Uptake Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seckler, D.; Amarasinghe, U.; Molden, D.; de Silva, R.; Barker, R. World Water Demand and Supply, 1990 to 2025: Scenarios and Issues; IWMI Research Report 19; IWMI: Colombo, Sri Lanka, 1998. [Google Scholar]
- Blanco, V.; Martínez-Hernández, G.B.; Artés-Hernández, F.; Blaya-Ros, P.J.; Torres-Sánchez, R.; Domingo, R. Water relations and quality changes throughout fruit development and shelf life of sweet cherry grown under regulated deficit irrigation. Agric. Water Manag. 2019, 217, 243–254. [Google Scholar] [CrossRef]
- Blanco, V.; Blaya-Ros, P.J.; Torres-Sánchez, R.; Domingo, R. Influence of Regulated Deficit Irrigation and Environmental Conditions on Reproductive Response of Sweet Cherry Trees. Plants 2020, 9, 94. [Google Scholar] [CrossRef] [Green Version]
- Zanotelli, D.; Montagnani, L.; Andreotti, C.; Tagliavini, M. Evapotranspiration and crop coefficient patterns of an apple orchard in a sub-humid environment. Agric. Water Manag. 2019, 226, 105756. [Google Scholar] [CrossRef]
- Fallahi, E. Influence of Rootstock and Irrigation Methods on Water Use, Mineral Nutrition, Growth, Fruit Yield, and Quality in ‘Gala’ Apple. HortTechnology 2012, 22, 731–737. [Google Scholar] [CrossRef]
- Ozturk, Z.N.; Talamè, V.; Deyholos, M.; Michalowski, C.B.; Galbraith, D.W.; Gozukirmizi, N.; Tuberosa, R.; Bohnert, H.J. Monitoring large-scale changes in transcript abundance in drought- and salt-stressed barley. Plant Mol. Biol. 2002, 48, 551–573. [Google Scholar] [CrossRef]
- Peter, F.; Waterman, P.A. Fertigation Guidelines in High Density Apples and Apple Nurseries in the Okanagan-Similkameen; Ministry of Agriculture, Food and Fisheries: Vancouver, BC, Canada, 2001.
- Fallahi, E.; Fallahi, B.; Shafii, B.; Morales, B. Water use, tree growth and leaf mineral nutrients of young ‘Fuji’ apples as influenced by different ırrigatıon systems. In Proceedings of the International Symposium on Mineral Nutrition of Fruit Plants, Talca, Chile, 31 October 2006. [Google Scholar]
- Fallahi, E.; Fallahi, B.; Neilsen, G.H.; Neilsen, D.; Peryea, F.J. Effects of mineral nutrition on fruit quality and nutritional disorders in apples. Acta Hortic. 2010, 868, 49–60. [Google Scholar] [CrossRef]
- Zegbe, J.A.; Serna-Pérez, A.; González-Fuentes, J.A. Nutrient status of apple leaves not affected by three years of irrigation using partial rootzone drying. J. Plant Nutr. Soil Sci. 2011, 174, 459–464. [Google Scholar] [CrossRef]
- Neilsen, G.H.; Neilsen, D. Nutritional requirements of apple. In Apples: Botany, Production and Uses; Ferree, D.C., Warrington, I.J., Eds.; CABI Publishing: Cambridge, MA, USA, 2003; pp. 267–302. [Google Scholar]
- Grabov, A. Plant KT/KUP/HAK Potassium Transporters: Single Family—Multiple Functions. Ann. Bot. 2007, 99, 1035–1041. [Google Scholar] [CrossRef] [Green Version]
- Zhao, D.; Oosterhuis, D.M.; Bednarz, C.W. Influence of Potassium Deficiency on Photosynthesis, Chlorophyll Content, and Chloroplast Ultrastructure of Cotton Plants. Photosynthetica 2001, 39, 103–109. [Google Scholar] [CrossRef]
- Ashley, M.K.; Grant, M.; Grabov, A. Plant responses to potassium deficiencies: A role for potassium transport proteins. J. Exp. Bot. 2006, 57, 425–436. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.Z.; Su, Y.H. Distinctive potassium-accumulation capability of alligator weed (Alternanthera philoxeroides) links to high-affinity potassium transport facilitated by K+ uptake systems. Weed Sci. 2013, 61, 77–84. [Google Scholar] [CrossRef]
- Song, Z.Z.; Yang, S.Y.; Zhu, H.; Jin, M.; Su, Y.H. Heterologous expression of an alligator weed high-affinity potassium transporter gene enhances salinity tolerance in Arabidopsis. Am. J. Bot. 2014, 101, 840–850. [Google Scholar] [CrossRef] [PubMed]
- Piper, C.S. Soil and Plant Analysis; Hans Publication: Bombay, India, 1966; p. 164. [Google Scholar]
- Arnon, Z.I. Determination of chlorophyll. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, A.D.; Duigan, S.P.; Berlyn, G.P. An evaluation of noninvasive methods to estimate foliar chlorophyll content. New Phytol. 2002, 153, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Slavik, B. Methods of Studying Plant Water Relations; Springer: Berlin/Heidelberg, Germany, 1974; Volume XVIII, p. 452. [Google Scholar]
- Beakbane, A.B.; Majumdar, P.K. A relationship between stomatal density and growth potential in apple rootstocks. J. Hortic. Sci. 1975, 50, 285–289. [Google Scholar] [CrossRef]
- Chapman, H.D. Suggested foliar sampling and handling techniques for determining the nutrient status of some field, horticultural and plantation crops. Indian J. Hortic. 1964, 21, 97–117. [Google Scholar]
- Jackson, M.L. Soil Chemical Analysis; Asia Publishing House: Bombay, India, 1967. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Gigova, A.; Moteva, M.; Mitova, T.; Kostadinov, G. Irrigation impact on leaf area and net photosynthetic productivity of Canola. In Proceedings of the 5th International Conference on Trends in Agricultural Engineering (TEA-2013), Prague, Czech Republic, 3–6 September 2013; pp. 173–180. [Google Scholar]
- Kucukyumuk, C.; Kacal, E. The Effects of different irrigation programmes in drip irrigation on leaf properties of Starkrimson delicious apple variety. Bull. UASVM Hortic. 2010, 67, 297–300. [Google Scholar]
- Yuste, J.; Rubioy, J.A.; Perez, M.A. Influence of plant density and water regime on soil water use, water relations and productivity of trellis trained tempranillo grape vines. Acta Hortic. 2004, 646, 187–193. [Google Scholar] [CrossRef]
- Klamkowski, K.; Treder, W. Response to drought stress of three strawberry cultivars grown under greenhouse conditions. J. Fruit Ornam. Plant Res. 2008, 16, 179–188. [Google Scholar]
- Eid, T.A.; Fatama, I.; Grah, A.; Hussein, S.M. Effect of soil moisture regimes and potassium application on growth, yield and fruit quality of “Canino” apricot (Prunus armeniaca L.). J. Plant Prod. 2014, 4, 621–640. [Google Scholar] [CrossRef]
- Trigo-Córdoba, E.; Bouzas-Cid, Y.; Orriols-Fernández, I.; Mirás-Avalos, J.M. Effects of deficit irrigation on the performance of grapevine (Vitis vinifera L.) cvs. Godello and Treixadura in Ribeiro, NW Spain. Agric. Water Manag. 2015, 161, 20–30. [Google Scholar] [CrossRef]
- Javadi, T.; Arzani, K.; Ebrahimzadeh, H. Study of proline, soluble sugar and chlorophyll A and B changes in nine Asian and one European pear cultivar under drought stress. Acta Hortic. 2008, 769, 241–246. [Google Scholar] [CrossRef]
- Haifeng, G.; Cuina, F.; Xinnan, L. Effects of drought stress on antioxidant system of leaves from different citrus rootstocks. J. Agric. Sci. Technol. 2011, 1, 32–35. [Google Scholar]
- Gholami, M.; Rahemi, M.; Kholdebarin, B.; Rastegar, S. Biochemical responses in leaves of four fig cultivars subjected to water stress and recovery. Sci. Hortic. 2012, 148, 109–117. [Google Scholar] [CrossRef]
- Flower, D.J.; Ludlow, M.M. Contribution of osmotic adjustment to the dehydration tolerance of water stressed pigeon pea (Cajanas cajan (L) Milsp) leaves. Plant Cell Environ. 1986, 9, 33–40. [Google Scholar]
- Gadallah, M.A.A. Effects of indole-3-acetic acid and zinc on the growth, osmotic potential and soluble carbon and nitrogen components of soybean plants growing under water deficit. J. Arid. Environ. 2000, 44, 451–467. [Google Scholar] [CrossRef]
- Romero, P.; Botia, P.; Garcia, F. Effects of regulated deficit irrigation under subsurface drip irrigation conditions on vegetative development and yield of mature almond trees. Plant Soil 2004, 260, 169–181. [Google Scholar] [CrossRef]
- Satisha, J.; Prakash, G.S.; Bhatt, R.M.; Sampathkumar, P. Effect of soil moisture stress on physiological response in grape (Vitis vinifera L.) varieties. J. Hortic. Sci. 2006, 1, 99–103. [Google Scholar]
- Alejendro, P.P.; Domingo, R.; Torrecillas, A.; Ruiz-Sanchez, M.C. Response of apricot trees to deficit irrigation strategies. Irrig. Sci. 2009, 27, 231. [Google Scholar]
- Elias, P. Stomata density and size of apple trees growing in irrigated and non-irrigated conditions. Biol. Bratisl. 1995, 50, 115–118. [Google Scholar]
- Kour, G.; Bakshi, P. Irrigation Management Practices and Their Influence on Fruit Agroecosystem. In Irrigation in Agroecosystems; BoD—Books on Demand: Norderstedt, Germany, 2018; pp. 21–36. [Google Scholar] [CrossRef] [Green Version]
- Basiouny, F.M. Response of peach seedlings to water stress and saturation conditions. Proc. Fla. State Hortic. Soc. 1977, 90, 261–263. [Google Scholar]
- Mısırlı, A.; Aksoy, U. The researches on leaf properties and stoma distrubition of Sarılop fig clone. Ege Univ. J. Agric. Fac. 1994, 31, 57–63. [Google Scholar]
- Klamkowski, K.; Treder, W. Morphological and physiological response of strawberry plants to water stress. Agric. Conspec. Sci. 2006, 71, 159–165. [Google Scholar]
- Kachwaya, D.S. Studies on Drip Irrigation and Fertigation in Strawberry (Fragaria × ananassa Duch.). Ph.D. Thesis, Dr Yashwant Singh Parmar University of Horticulture and Forestry, Nauni, India, 2014. [Google Scholar]
- Garg, B.K. Nutrient uptake and management under drought: Nutrient-moisture interaction. Curr. Agric. 2003, 27, 1–8. [Google Scholar]
- Havlin, J.L.; Beaton, J.D.; Tisdale, S.L.; Nelson, W.L. Soil Fertility and Fertilizers, an Introduction to Nutrient Management, 6th ed.; Macmillan, Inc.: New York, NY, USA, 1999; p. 409. [Google Scholar]
- Kacar, B.; Katkat, A.V. Gübreler ve Gübreleme Tekniği, 2nd ed.; Nobel Yayin No: 1119; Nobel Basımevi, 560 s; Fen ve Biyoloji Yayınları Dizisi: Ankara, Turkey, 2007. [Google Scholar]
- Thakur, J.; Sharma, J.C.; Mohit, S.; Verma, P. Vegetative Growth and Foliage Nutrient Content of Super Chief Apple under Different Irrigation and Fertigation Schedules in NW Himalayan Region. Commun. Soil Sci. Plant Anal. 2020, 51, 2174–2188. [Google Scholar] [CrossRef]
- Shirgure, P.S.; Srivastava, A.K.; Singh, S.; Pimpale, A.R. Drip irrigation scheduling, growth, yield and quality of acid Lime (Citrus aurantifolia). Indian J. Agric. Sci. 2004, 74, 92–94. [Google Scholar]
- Chauhan, N.; Chandel, J.S. Growth, productivity, leaf nutrient contents and water-use efficiency of kiwifruit (Actinidia deliciosa) under drip and basin irrigation system. Indian J. Agric. Sci. 2010, 80, 584–587. [Google Scholar]
- Küçükyumuk, Z.; Küçükyumuk, C.; İbrahim, E.I.; Eraslan, F. Effects of different sweet cherry rootstocks and drought stress on nutrient concentrations. J. Agric. Sci. 2015, 21, 431–438. [Google Scholar] [CrossRef]
- Zhong, Y.; Fei, L.; Li, Y.; Zeng, J.; Dai, Z. Response of fruit yield, fruit quality, and water use efficiency to water deficits for apple trees under surge-root irrigation in the loess plateau of China. Agric. Water Manag. 2019, 222, 221–230. [Google Scholar] [CrossRef]
- Dechorgnat, K.L.; Francis, K.L.; Dhagga, K.S.; Rafalski, J.A.; Tyermal, S.D.; Kaiser, B.N. Tissue and nitrogen linked expression profiles of ammonium and nitrate transporters in maize. BMC Plant Biol. 2019, 19, 206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, F.Q.; Young, J.; Crawford, N.M. The nitrate transporter AtNRT1. 1 (CHL1) functions in stomatal opening and contributes to drought susceptibility in Arabidopsis. Plant Cell 2003, 15, 107–111. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.Z.; Yang, Y.; Ma, R.J.; Xu, J.L.; Yu, X.L. Transcription of potassium transporter genes of KT/HAK/KUP family in peach seedlings and responses to abiotic stresses. Biol. Plant. 2015, 59, 65–73. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No. | Particulars | Status | Textural Class | Method Followed |
|---|---|---|---|---|
| 1. | Coarse sand (%) | 0.46% | Medium loam | International Pipette Method (Piper, 1966) |
| 2. | Fine sand (%) | 46.33% | ||
| 3. | Silt (%) | 35.00% | ||
| 4. | Clay (%) | 18.00% |
| Treatment | Treatment Combination | Irrigation Applied (Litre/Plant) | ||
|---|---|---|---|---|
| 2017 | 2018 | |||
| T1 | Rainfed | I0 | - | - |
| T2 | Drip Irrigation at 100% ETc during flowering and fruit set | I1 C1 | 95.80 | 87.95 |
| T3 | Drip Irrigation at 75% ETc during flowering and fruit set | I2 C1 | 72.30 | 65.75 |
| T4 | Drip Irrigation at 50% ETc during flowering and fruit set | I3 C1 | 47.60 | 43.78 |
| T5 | Drip Irrigation at 100% ETc during fruit growth stage | I1 C2 | 320.61 | 350.25 |
| T6 | Drip Irrigation at 75% ETc during fruit growth stage | I2 C2 | 240.08 | 262.10 |
| T7 | Drip Irrigation at 50% ETc during fruit growth stage | I3 C2 | 158.90 | 174.36 |
| T8 | Drip Irrigation at 100% ETc during pre-harvest stage | I1 C3 | 214.35 | 182.18 |
| T9 | Drip Irrigation at 75% ETc during pre-harvest stage | I2 C3 | 160.78 | 136.23 |
| T10 | Drip Irrigation at 50% ETc during pre-harvest stage | I3 C3 | 106.80 | 89.90 |
| T11 | Drip Irrigation at 100% ETc throughout the growing season | I1 C4 | 631.30 | 638.20 |
| T12 | Drip Irrigation at 75% ETc throughout the growing season | I2 C4 | 472.80 | 464.00 |
| T13 | Drip Irrigation at 50% ETc throughout the growing season | I3 C4 | 306.70 | 307.90 |
| Month | ETc | Total Irrigation (Liter) 2017 | ||
|---|---|---|---|---|
| 100% ETc | 75% ETc | 50% ETc | ||
| April | 44.2 | 17.4 | 13.2 | 8.6 |
| May | 103.2 | 78.4 | 59.1 | 32.4 |
| June | 152.0 | 133.8 | 100.3 | 66.1 |
| July | 201.2 | 186.8 | 139.7 | 92.8 |
| August | 167.0 | 159.4 | 119.1 | 79.5 |
| September | 62.5 | 55.5 | 41.4 | 27.3 |
| Total | 730.3 | 631.3 | 472.8 | 306.7 |
| Month | ETc | Total Irrigation (Liter) 2018 | ||
| 100% ETc | 75% ETc | 50% ETc | ||
| April | 37.6 | 13.4 | 10 | 6.7 |
| May | 90.8 | 74.5 | 55.7 | 37 |
| June | 181.3 | 170.5 | 127.6 | 84.8 |
| July | 219.6 | 197.7 | 134.5 | 89.5 |
| August | 169.0 | 109.7 | 82.3 | 54.1 |
| September | 72.6 | 72.4 | 53.9 | 35.8 |
| Total | 770.9 | 638.2 | 464.0 | 307.9 |
| Primer | Description | Forward Primer (5′–3′) | Reverse Primer 3′–5′ | Amplicon Size (bp) |
|---|---|---|---|---|
| MdNFPr | Nitrogen transporter gene | GGCCAACCAGGTTTCACTAA | ACCCACCAGGAAGAACTGTG | 170 |
| POTT | Potassium transporter gene | CAGAGTTCATTCGGTCAGA | CTTGCTTCCATCAGTTCCT | 277 |
| Irrigation Level (I) | Leaf Area (cm2) | Leaf Chlorophyll (mg/g) | ||||
|---|---|---|---|---|---|---|
| 2017 | 2018 | Pooled | 2017 | 2018 | Pooled | |
| I0 | 19.12 b | 19.58 c | 18.85 c | 1.59 b | 1.48 b | 1.53 b |
| I1 | 25.45 a | 25.75 a | 25.60 a | 2.00 a | 1.96 a | 1.96 a |
| I2 | 25.02 a | 25.16 a | 25.09 a | 1.97 a | 1.94 a | 1.94 a |
| I3 | 23.78 b | 22.37 b | 23.57 b | 1.95 a | 1.85 ab | 1.90 a |
| Pr > F | 0.0182 | 0.0184 | 0.013 | 0.041 | 0.002 | <0.0001 |
| Crop Phenological Stage (C) | ||||||
| C1 | 23.67 b | 26.83 b | 24.99 b | 1.74 b | 1.71 c | 1.71 c |
| C2 | 23.15 b | 22.03 c | 23.03 c | 1.97 ab | 1.93 b | 1.95 b |
| C3 | 21.00 c | 20.30 d | 20.65 d | 1.97 ab | 1.91 b | 1.94 b |
| C4 | 30.52 a | 30.84 a | 30.68 a | 2.21 a | 2.10 a | 2.16 a |
| Pr > F | <0.0001 | <0.0001 | <0.0001 | 0.035 | 0.002 | <0.0001 |
| Irrigation Level × Crop Phenological Stage (IC) | ||||||
| I0 | 19.12 i | 19.58 h | 18.85 g | 1.59 c | 1.48 e | 1.53 d |
| I1 C1 | 25.64 d | 27.40 c | 26.04 c | 1.78 abc | 1.74 bcd | 1.76 cd |
| I2 C1 | 23.84 ef | 26.95 cd | 25.19 cd | 1.73 abc | 1.72 cd | 1.72 cd |
| I3 C1 | 20.61 hi | 26.14 d | 23.73 de | 1.71 bc | 1.69 d | 1.70 cd |
| I1 C2 | 24.68 de | 23.14 e | 24.39 cde | 1.98 abc | 1.94 bc | 1.96 ab |
| I2 C2 | 23.44 f | 22.08 f | 22.96 ef | 1.97 abc | 1.93 bc | 1.95 ab |
| I3 C2 | 21.33 gh | 21.99 f | 21.30 fg | 1.97 abc | 1.91 bc | 1.94 ab |
| I1 C3 | 20.88 ghi | 20.75 g | 21.82 g | 1.96 abc | 1.82 bcd | 1.90 bc |
| I2 C3 | 21.86 g | 20.19 gh | 20.02 g | 1.99 abc | 1.97 b | 1.97 c |
| I3 C3 | 20.27 hi | 19.96 gh | 20.11 g | 1.97 abc | 1.95 bc | 1.96 c |
| I1 C4 | 31.96 a | 31.70 a | 31.85 a | 2.26 a | 2.22 a | 2.23 a |
| I2 C4 | 30.70 b | 31.43 a | 31.06 a | 2.21 ab | 2.19 a | 2.18 a |
| I3 C4 | 28.91 c | 29.40 b | 29.15 b | 1.97 abc | 1.99 b | 1.98 ab |
| Pr > F | <0.0001 | <0.0001 | <0.0001 | 0.008 | <0.0001 | <0.0001 |
| Irrigation Level (I) | Leaf Relative Water Content (%) | Stomatal Density (mm) | ||||
|---|---|---|---|---|---|---|
| 2017 | 2018 | Pooled | 2017 | 2018 | Pooled | |
| I0 | 70.20 c | 69.63 c | 69.91 c | 167.00 b | 165.00 b | 166.20 c |
| I1 | 76.14 a | 76.44 a | 76.29 a | 188.00 a | 191.00 a | 189.50 a |
| I2 | 75.48 ab | 75.73 ab | 75.60 ab | 187.50 a | 186.50 a | 187.12 a |
| I3 | 74.84 b | 74.78 b | 74.79 b | 179.00 ab | 177.50 ab | 178.75 b |
| Pr > F | 0.038 | 0.032 | 0.0002 | 0.036 | 0.039 | 0.0003 |
| Crop Phenological Stage (C) | ||||||
| C1 | 72.80 c | 72.20 c | 72.50 d | 174.66 | 173.00 | 173.83 b |
| C2 | 73.60 c | 74.16 b | 73.88 c | 190.33 | 190.66 | 190.50 a |
| C3 | 75.76 b | 75.19 b | 75.83 b | 178.00 | 177.00 | 177.50 b |
| C4 | 79.78 a | 80.28 a | 80.03 a | 196.66 | 197.00 | 196.83 a |
| Pr>F | <0.0001 | <0.0001 | <0.0001 | (NS) | (NS) | 0.0002 |
| Irrigation Level × Crop Phenological Stage (IC) | ||||||
| I0 | 70.20 g | 69.63 h | 69.91 h | 167.00 d | 165.00 d | 166.20 e |
| I1 C1 | 73.13 ef | 72.62 g | 72.87 fg | 176.00 c | 175.00 c | 176.50 cd |
| I2 C1 | 73.07 ef | 72.36 g | 72.71 fg | 177.00 c | 175.00 c | 176.00 cd |
| I3 C1 | 72.21 f | 71.62 gh | 71.91 g | 171.00 cd | 169.00 cd | 170.00 d |
| I1 C2 | 74.41 de | 75.08d ef | 74.74 de | 194.00 ab | 195.00 ab | 194.50 ab |
| I2 C2 | 73.25ef | 73.93 efg | 73.59 ef | 193.00 ab | 192.00 b | 192.50 b |
| I3 C2 | 73.14 ef | 73.47 gf | 73.30 fg | 184.00 bc | 185.00 bc | 184.50 bc |
| I1 C3 | 76.36 c | 76.60 cd | 76.51 c | 180.00 bc | 179.00 bcd | 182.30 bc |
| I2 C3 | 75.54 cd | 75.99 de | 75.76 cd | 180.00 bc | 178.00 bcd | 179.00 cd |
| I3 C3 | 75.39 cd | 75.09d ef | 75.24 cd | 174.00 cd | 174.00 c | 174.00 cd |
| I1 C4 | 80.67 a | 81.41 a | 81.04 a | 202.00 a | 204.00 a | 203.00 a |
| I2 C4 | 80.07 ab | 80.63 ab | 80.35 a | 201.00 a | 201.00 ab | 201.00 ab |
| I3 C4 | 78.62 b | 78.81 bc | 78.71 b | 187.00 b | 186.00 bc | 186.50 bc |
| Pr > F | <0.0001 | <0.0001 | <0.0001 | 0.045 | 0.008 | 0.016 |
| Treatment | Length (L) × Width (W) | |
|---|---|---|
| 2017 | 2018 | |
| I0 | 25.2 f× 16.8 de | 21.0 g × 16.8 f |
| I1C1 | 29.4 e×15.6 de | 29.4 e × 17.8 3 |
| I2C1 | 29.4 e×16.8 f | 28.2 e× 16.8 f |
| I3C1 | 28.4 e×16.7 e | 25.2 f × 21.00 c |
| I1C2 | 33.6 c × 16.8 e | 33.8 c × 21.7 c |
| I2C2 | 32.8 c × 19.8 e | 31.5 d × 21.7 d |
| I3C2 | 30.8 d ×17.8 d | 30.8 d × 16.8 d |
| I1C3 | 35.2 b × 21.7 b | 35.7 b × 25.2 b |
| I2C3 | 35.2 b × 19.3 c | 33.8 c × 21.00 c |
| I3C3 | 33.8 c × 19.2 c | 33.8 c × 19.2 c |
| I1C4 | 37.8 a × 25.20 a | 37.8 a × 29.4 a |
| I2C4 | 37.1 a × 24.7 a | 35.7 a × 25.2 a |
| I3 C4 | 35.4 a × 24.2 a | 35.7 a × 21.7 a |
| Pr > F | <0.0001 (L) <0.0001 (W) | <0.0001 (L) <0.0001 (W) |
| Irrigation Level (I) | N | P | K | Ca | Mg |
|---|---|---|---|---|---|
| I0 | 1.71 b | 0.20 b | 1.21 b | 1.28 b | 0.28 b |
| I1 | 2.03 a | 0.28 a | 1.35 a | 1.45 a | 0.33 a |
| I2 | 1.94 a | 0.26 a | 1.33 ab | 1.41 a | 0.31 a |
| I3 | 1.87 ab | 0.26 a | 1.31 ab | 1.40 a | 0.30 a |
| Pr > F | 0.012 | 0.004 | 0.0011 | <0.0001 | 0.0003 |
| Crop Phenological Stage (C) | |||||
| C1 | 1.81 ab | 0.25 b | 1.30 b | 1.40 c | 0.32 b |
| C2 | 1.85 ab | 0.27 b | 1.32 b | 1.44 b | 0.32 b |
| C3 | 1.76 b | 0.23 c | 1.26 c | 1.33 d | 0.27 b |
| C4 | 2.34 a | 0.31 a | 1.45 a | 1.51 a | 0.34 a |
| Pr > F | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.0002 |
| Irrigation Level × Crop Phenological Stage (IC) | |||||
| I0 | 1.71 c | 0.20 e | 1.21 e | 1.28 f | 0.25 d |
| I1 C1 | 1.92 b | 0.26 abcd | 1.32 cd | 1.43 cd | 0.34 bc |
| I2 C1 | 1.88 b | 0.25 bcde | 1.30 cde | 1.40 cde | 0.32 cd |
| I3 C1 | 1.77 bc | 0.25 cde | 1.29 de | 1.38 de | 0.32 cd |
| I1 C2 | 1.87 b | 0.28 abcd | 1.36 cd | 1.46 ef | 0.33 bc |
| I2 C2 | 1.81 bc | 0.27 abcd | 1.33 cd | 1.43 ef | 0.32 bc |
| I3 C2 | 1.75 c | 0.24 cde | 1.27 de | 1.43 f | 0.30 cd |
| I1 C3 | 1.78 bc | 0.24 cde | 1.27 de | 1.34 bc | 0.29 bc |
| I2 C3 | 1..76 bc | 0.23 de | 1.26 de | 1.34 cd | 0.28 bc |
| I3 C3 | 1.75 bc | 0.23 de | 1.26 de | 1.31 cd | 0.27 bc |
| I1 C4 | 2.48 a | 0.31 a | 1.48 a | 1.58 a | 0.36 a |
| I2 C4 | 2.35 a | 0.31 ab | 1.46 b | 1.51 b | 0.34 ab |
| I3 C4 | 2.19 ab | 0.30 abc | 1.42 bc | 1.45 bc | 0.33 abc |
| Pr > F | 0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.008 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mushtaq, R.; Sharma, M.K.; Mir, J.I.; Mansoor, S.; Mushtaq, K.; Popescu, S.M.; Malik, A.R.; El-Serehy, H.A.; Hefft, D.I.; Bhat, S.A.; et al. Physiological Activity, Nutritional Composition, and Gene Expression in Apple (Malus domestica Borkh) Influenced by Different ETc Levels of Irrigation at Distinct Development Stages. Water 2021, 13, 3208. https://doi.org/10.3390/w13223208
Mushtaq R, Sharma MK, Mir JI, Mansoor S, Mushtaq K, Popescu SM, Malik AR, El-Serehy HA, Hefft DI, Bhat SA, et al. Physiological Activity, Nutritional Composition, and Gene Expression in Apple (Malus domestica Borkh) Influenced by Different ETc Levels of Irrigation at Distinct Development Stages. Water. 2021; 13(22):3208. https://doi.org/10.3390/w13223208
Chicago/Turabian StyleMushtaq, Rafiya, Mahinder Kumar Sharma, Javid Iqbal Mir, Sheikh Mansoor, Khalid Mushtaq, Simona Mariana Popescu, Abdul Raouf Malik, Hamed A. El-Serehy, Daniel Ingo Hefft, Sajad Ahmad Bhat, and et al. 2021. "Physiological Activity, Nutritional Composition, and Gene Expression in Apple (Malus domestica Borkh) Influenced by Different ETc Levels of Irrigation at Distinct Development Stages" Water 13, no. 22: 3208. https://doi.org/10.3390/w13223208
APA StyleMushtaq, R., Sharma, M. K., Mir, J. I., Mansoor, S., Mushtaq, K., Popescu, S. M., Malik, A. R., El-Serehy, H. A., Hefft, D. I., Bhat, S. A., & Narayan, S. (2021). Physiological Activity, Nutritional Composition, and Gene Expression in Apple (Malus domestica Borkh) Influenced by Different ETc Levels of Irrigation at Distinct Development Stages. Water, 13(22), 3208. https://doi.org/10.3390/w13223208