1. Introduction
During the last decades, much attention has been paid to the role of megabenthic invertebrate structures composed mainly by invertebrates in benthic-pelagic coupling processes [
1]. It is well known that these organisms, mainly active or passive suspension feeders, have a determinant role in biogeochemical cycles and near bottom seston regulation [
2]. Biogenic structures produced by sponges, cnidarians, bryozoans, etc., also known as marine animal forests [
3,
4], may even potentially change, depending on their structure, patch size, and current regime, the seston concentration [
5]. The dependence on currents and seston concentration has been claimed as an essential point to understand benthic-pelagic coupling processes [
6,
7]; nevertheless, there are few studies highlighting the role of short-time cycles as energy providers in marine animal forests.
It is well known that in a brief time (i.e., few hours), the seston concentration in near bottom water layers may suddenly change in concentration and quality [
8,
9,
10,
11]. Such changes may have an immediate response to the energy fluxes in terms of activity of the benthic community [
12,
13,
14,
15]. When the benthic activity or the abundance of meiofauna has been analyzed in the deep sea, for example, a fast response to the primary productivity have been found according to different areas of the world, suggesting that the quantity and quality of the organic material transferred from the pelagic to the benthic domain is a crucial step to understand the energetic implications of these transfer processes [
16,
17,
18,
19]. However, nor in shallow or deep waters has a short time approach been adopted in which the seston variables, the activity of the suspension feeders, and the capture rates have been simultaneously quantified to understand the importance of such food pulses in the benthic-pelagic coupling processes.
Several studies indicated that the quality and quantity of the available food would be essential in the understanding of processes determining changes in health status of marine animal forests driven by anthropogenic impacts at local scale in a wider framework of global climate change stressors [
20,
21]. However, to develop reliable models describing changes in energy inputs due to these emerging constrains, it is crucial also to know how short time cycles influence the capture rates in natural conditions, and how fluctuations in seston concentration and food availability may affect the global energy budget of these benthic suspension feeders. Every single community responds differently to the food pulses [
20] and knowing the importance of the short-term variability in the prey capture and associated carbon entry may be a key point that has been largely neglected so far [
22]. There is a baseline of seston concentration that all the benthic suspension feeders take advantage transversally (i.e., continuously capturing available particles through filtration, depending on the season), but single pulses have been also detected through the year in different marine animal forests [
23,
24,
25], demonstrating that pulse-like feeding may be essential to understand the overall budget of these invertebrates and its future constraints. In the optimal forage theory [
26], all the organisms tend to optimize the available resources for their own benefit with different strategies, including the patch structure and density [
27]. In benthic suspension feeders, such optimization depends on the feeding strategy [
2,
28], being dependent on the seston concentration, quality and the hydrodynamics of the specific area and time of the year. The comprehension of how, in coincidence with the major productivity period of the year, each benthic organism responds in a short time feeding on the available seston will be essential to understand the energetic dynamics of the whole benthic community.
Among the organisms that form the marine animal forests, gorgonians have an important role in benthic-pelagic processes due to their abundance in several habitats of the world [
29,
30,
31,
32,
33]. In this study, we examined high temporal resolution variability in the polyp expansion and plankton prey captures of three different Mediterranean passive suspension feeders:
Eunicella singularis (Esper, 1791),
Paramuricea clavata (Risso, 1826), and
Leptogorgia sarmentosa (Esper, 1791).
Eunicella singularis is a mixotrophic species that feeds on particulate organic carbon [
34] but, in shallow areas, relies also in the photosynthetic activity [
35]. Its bathymetric distribution range is wide [
36], being the morphology and the heterotrophic input different when shallow and deeper populations are compared [
37,
38].
Paramuricea clavata settles preferentially on steep walls [
36], it is exclusively heterotrophic, and has a wide diet spectrum, being the zooplankton a significant part of its energy input [
30,
39].
Leptogorgia sarmentosa prefers detrital-gravel horizontal habitats [
36], it is also exclusively heterotrophic, having a diet based in zooplankton, detritus, and picoplankton [
40,
41].
The goal of the present research is to assess how the food pulses affect the capture rates (in terms of growth and C uptake and utilization) monitoring the three species in a short-time cycle, having each species a different patch density and colony branching pattern. To do that, we studied the (i) seston quantity and quality in the near bottom water layer, (ii) the polyp activity in the field, and (iii) the prey capture composition and rates of the three different passive suspension feeders at several temporal scales by using an high-frequency temporal sampling. Thus, the present study highlights the role of passive suspension feeders in energy transferring to benthic communities in temperate littoral ecosystems depending on their distribution and also patch density.
2. Materials and Methods
The field survey was conducted in the Medes Islands, NW Mediterranean (40°02′55″ N, 3°13′30″ E). Sampling and observations were carried out at 18–20 m depth among a coralligenous reef located in a steep wall and boulders. The area was alternately influenced by northerly and southerly currents, which may reach high speeds (from 2 up to 30 cm s
−1, [
10,
15]). The underwater topography is markedly asymmetrical, with prominent slopes on the northern side and gentle slopes on the southern side. Strong, dry winds blow from the north-northwest in spring-summer. Water temperature is also highly seasonal, with a strong thermocline between April and October at a depth of 20–30 m (with differences of 7–10 °C) [
10]. Salinity is usually around 37.5–38‰, with maximum values in winter during mixing events produced by heavy storms caused by the north winds [
10].
The three species analyzed were octocorals: Paramuricea clavata, Eunicella singularis, and Leptogorgia sarmentosa. Paramuricea clavata has a mean density of 56 colonies m−2, whilst E. singularis has a mean density of 41 colonies m−2 in the study area. Leptogorgia sarmentosa has a mean density of 1.5 colonies m−2.
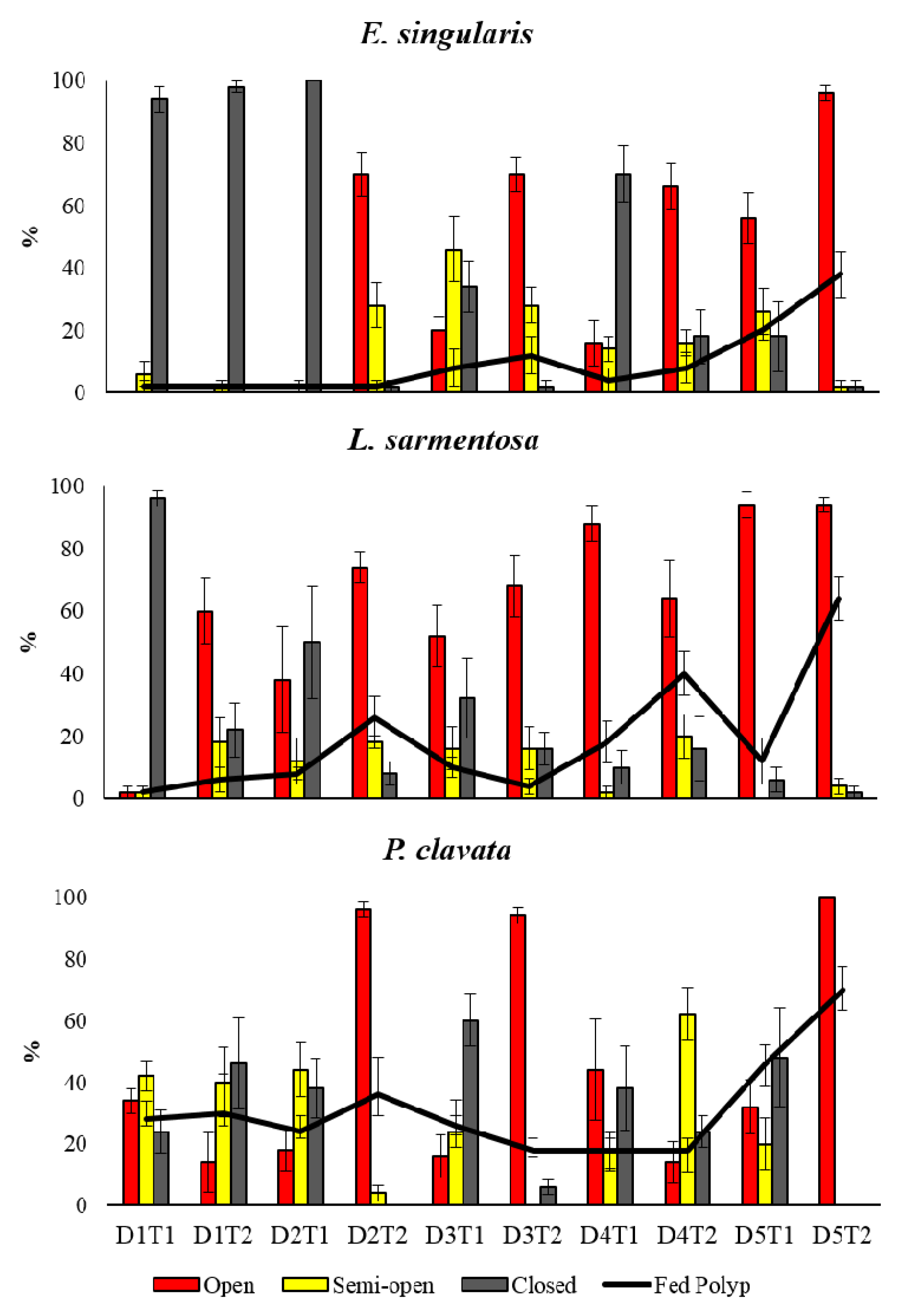
2.1. Polyp Expansion
Species polyp expansion was monitored at a high frequency (i.e., two sampling times within the day) during the late spring-early summer. This period was chosen because pelagic primary production and the frequency of seston pulses is high [
10]. Expansion is defined as the maximum aperture of polyps [
15,
42]. Polyp activity was observed in ten groups of five colonies each time by scuba divers in patches distributed along 30–40 meters. On each sampling time, the percentages of expanded semi-expanded or contracted polyps was recorded in colonies of
E. singularis,
L. sarmentosa, and
P. clavata.
2.2. Environmental and Biological Parameters of the Water Column
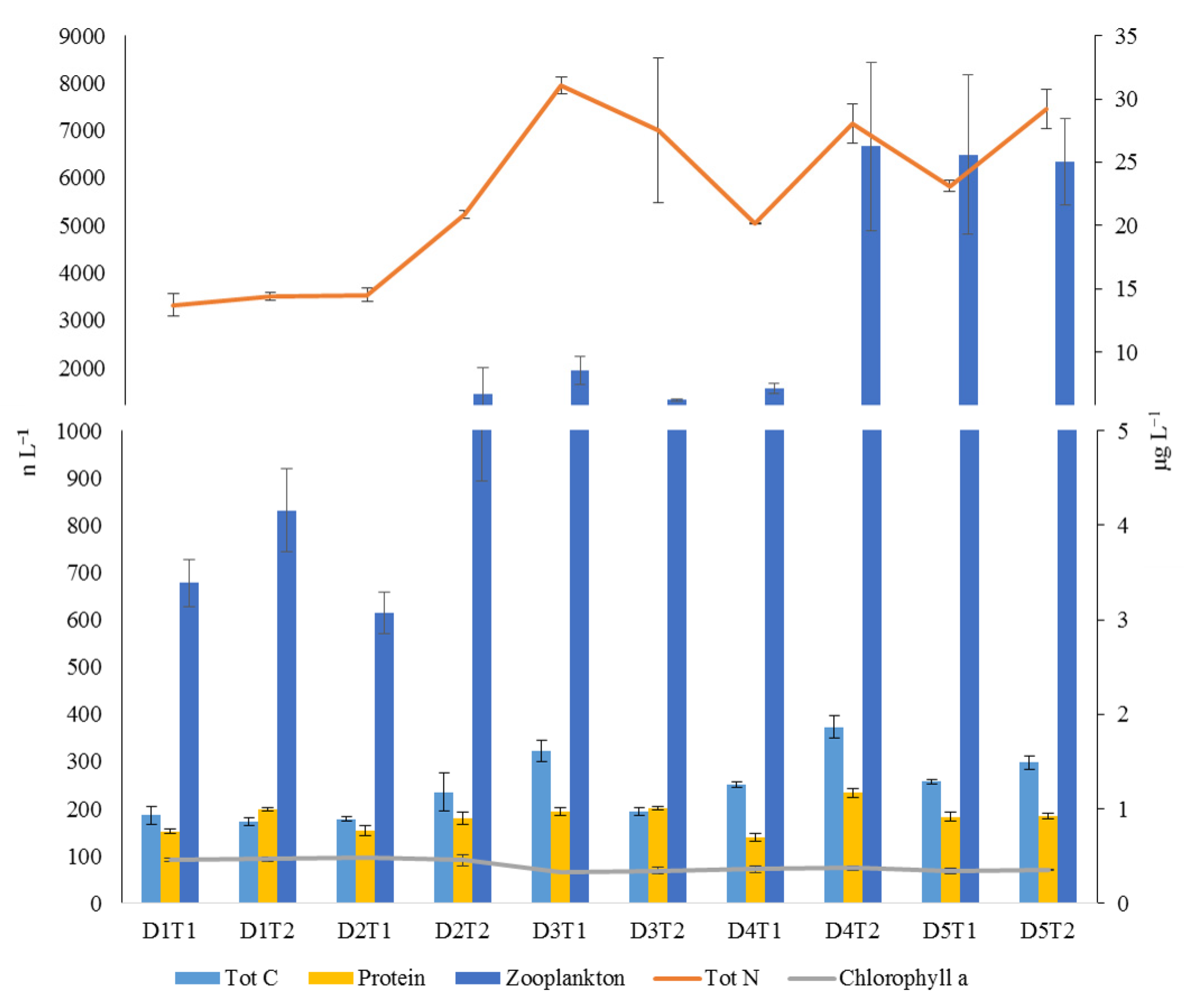
Several environmental and biological variables were concomitantly monitored: (1) seston concentration and quality, the latter determined by assessment of total particulate carbon and nitrogen, chlorophyll a and protein concentrations [
10], and (2) zooplankton concentration, determined by analyzing three samples collected by a scuba diver towing a plankton net (22 cm in diameter with a mesh size of 100 µm) for a distance of 40 m [
30,
41].
2.3. Analysis of Gut Contents
Feeding on plankton in the three species was assessed by means of gut content examinations of apical fragments [
41]. The colonies were sampled two times each day during the whole intensive cycle (5 days). Each sample consisted of one apical fragment collected from five randomly selected colonies. The species collected (
P. clavata,
E. singularis, and
L. sarmentosa) are not protected species; we have a specific permit to operate and sample in the area from the local authorities, the Parc Natural de les Illes Medes.
The fragments were immediately placed in 10% formaldehyde solution in seawater to prevent further digestion. The contents of 50 polyps selected randomly from each sample (ten from each apical fragment of five colonies in each sampling time per each species) were isolated by dissection under a binocular microscope, identified to the higher taxon level and counted. The length of all prey was measured under the microscope with a micrometer ruler.
2.4. Benthic-Pelagic Coupling Calculations
2.4.1. Prey Capture Rate
The zooplankton capture rate, expressed as the number of prey items captured per polyp and hour, was calculated using the following equation [
30]:
where C is the number of preys captured per polyp per hour, N is the number of prey items per polyp, t is time (in hours), and D is digestion time (in hours). Digestion time applied for the three species considered the temperature recorded (19.5 °C, [
10]) in the study area in this time of the year. The applied digestion time is based on the available literature of
E. singularis and
L. sarmentosa [
34,
41]; no data on
P. clavata digestion time at this temperature range has been calculated in previous studies; thus, the previous two species digestion was also used to make the
P. clavata calculations on prey capture rates.
2.4.2. Prey Biomass
Prey biomass was estimated from biovolumes [
43], using conversion factors for wet weight (1.025; [
44]), dry weight (13% of wet weight, [
45]), and carbon content (45% of dry weight, [
46]), as per Rossi et al. [
41].
2.4.3. Potential Impact on Seston
To understand how much seston is needed to satisfy the needs of a single polyp during the day, a combined approach was applied.
First, the total seston concentration, minimum and maximum total carbon values across the sampling times were estimated. The obtained values were multiplied by the coefficient 0.66, as it has been shown that in this area, the inorganic carbon represents approximately 33% of the seston [
47]. Those quantities represent the organic carbon (i.e., 105.5 and 267.9 µg C L
−1, respectively). Then, the obtained values were divided by the maximum ingesta of a single polyp throughout the entire cycle (values found in the present study, see below). This number will represent the potential quantity of polyps of each species that may feed with one litre of near bottom seawater in minimum and maximum seston concentration across the sampling times.
Second, the minimum and maximum zooplankton concentration values throughout the sampling times (i.e., 533 and 8437 zooplankters m−3, respectively; values found in the present study, see below) were estimated. Then, the obtained values were divided by the maximum ingesta of a single polyp throughout the entire cycle. This number will be the potential number of polyps of each species that may withstand with one cubic meter of near bottom water in minimum and maximum zooplankton concentration across sampling times.
2.5. Statistical Analysis
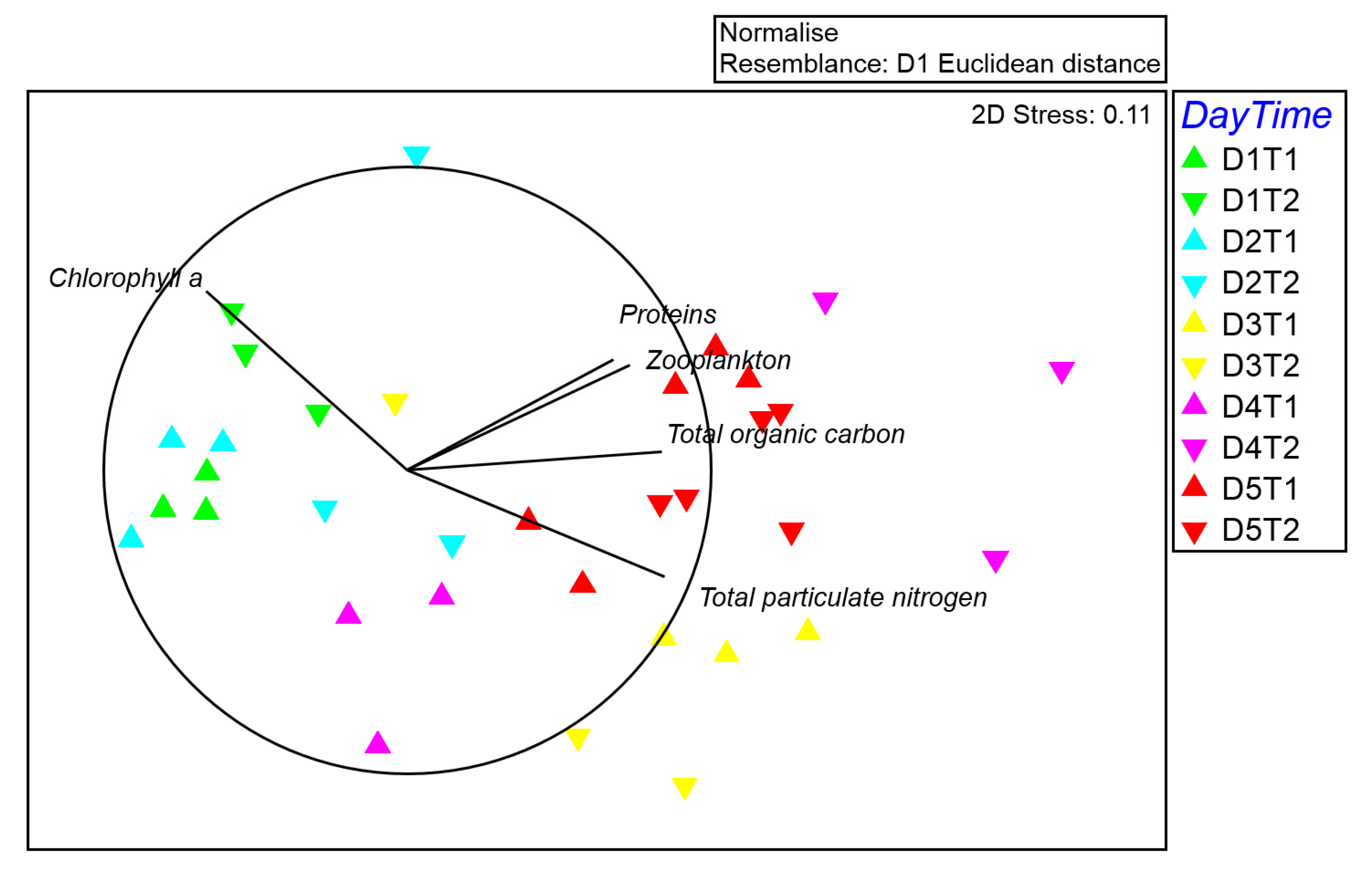
The variability in seston composition (total particulate carbon and nitrogen, chlorophyll a, and protein concentrations) and zooplankton density was assessed at several temporal scales by multivariate analyses. The design consisted of two factors: Day (D, as a random factor with 5 levels, each 24 h) and Time (T, as a random factor with 2 levels, nested in D, twice per day), with n = 3. Multivariate analyses of variance (PERMANOVA, [
47]) was performed based on Euclidean distances of previously normalized data, using 9999 random permutations of the appropriate units [
48].
To assess differences in polyp expansion, fed polyp percentages and prey biomass of fed polyps belonging to the three investigated species, univariate permutational analyses of variance (PERMANOVA, [
47]) were performed, based on Euclidean distances of untransformed data, using 9999 random permutations of the appropriate units [
48]. Differences in prey composition were assessed performing permutational analyses of variance (PERMANOVA, [
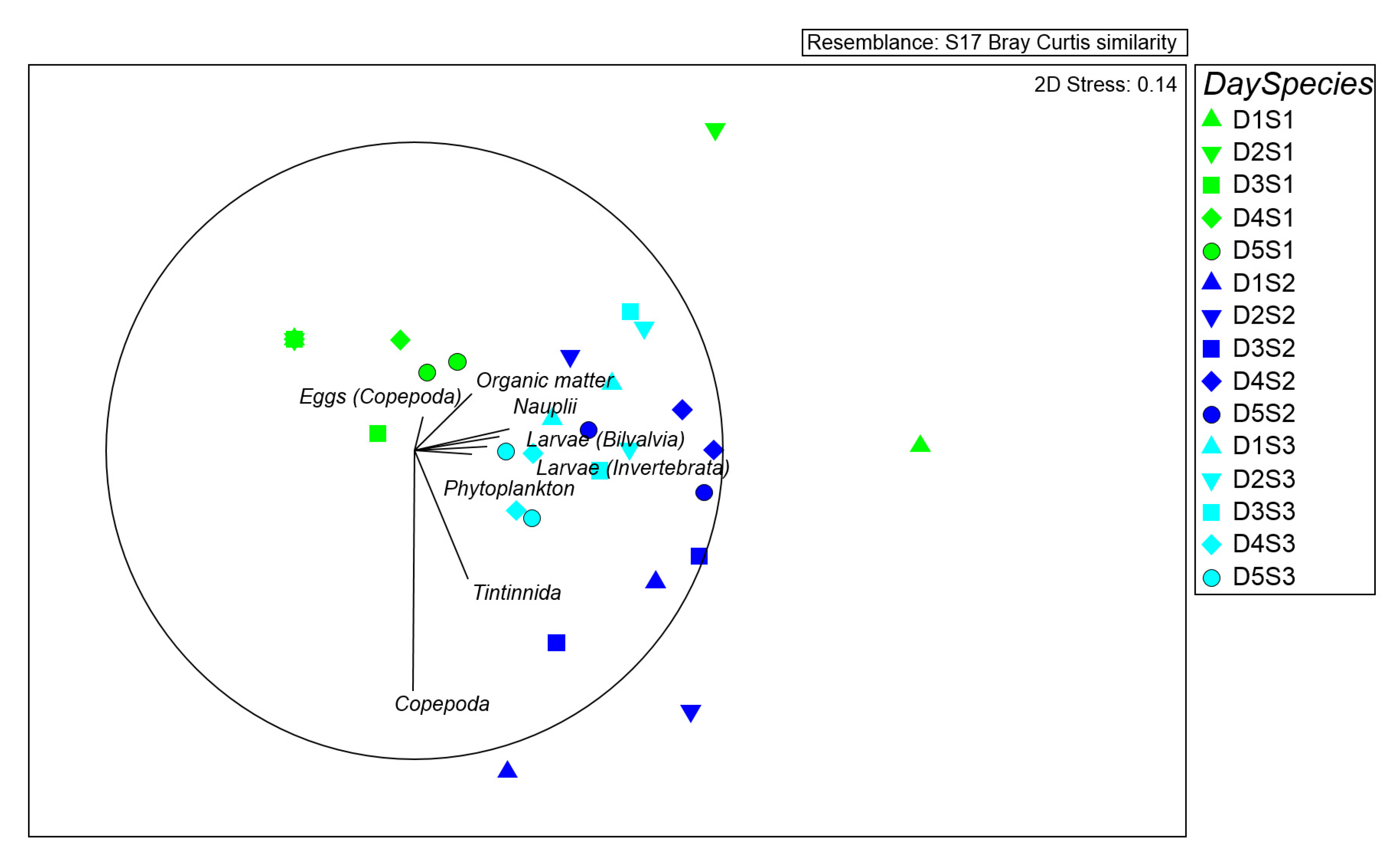
47]) based on Bray Curtis dissimilarities. The design adopted consisted of three factors: Day (D, as a random factor with 5 levels, each 24 h) Time (T, as a random factor with 2 levels, nested in D, twice per day), and Species (S, as fixed factor with 3 levels) with n = 5.
Finally, in order to assess differences in gut contents of colonies belonging to the three gorgonian species, multivariate permutational analyses of variance (PERMANOVA, [
47]) were performed based on Bray Curtis similarities of untransformed data adding a dummy variable, using 9999 random permutations of the appropriate units [
48]. The experimental design consists of four factors: Day (D, as a random factor with 5 levels, each 24 h) Time (T, as a random factor with 2 levels, nested in D, twice per day), Species (S, as fixed factor with 3 levels), and Colony (C, as random factor nested in S, with 5 levels), with n = 10.
When significant differences were encountered (
p < 0.05), post-hoc pairwise tests were carried out in order to ascertain the consistency of the differences across the different conditions tested. Because of the restricted number of unique permutations in the pairwise tests,
p-values were obtained from Monte Carlo tests. To examine the generality of patterns in seston and zooplankton densities, and prey composition of fed polyps, Multidimensional Scaling (MDS) plots were generated. The analyses were performed using PRIMER v. 6 software [
49] including the PERMANOVA + add-on package [
49,
50].
4. Discussion
In the present paper, for the first time, seston variables and prey capture rates are contrasted in different species of passive suspension feeders with different colony densities, analyzing the potential relations throughout a short time cycle. It has been reported that, to make energetic balances, short time cycles may be essential although neglected [
51].
Seston variables found in the present study (including zooplankton) are in line with the variability found in previous Mediterranean studies in which coastal or near bottom water column parameters were analyzed [
30,
41,
52,
53,
54]. Wide variations in a short time, especially in zooplankton, have been however seldom analyzed in near bottom seston in warm temperate seas and have been considered as an obstacle to understand the potential mechanisms in benthic-pelagic coupling processes [
10,
55].
The present work is also in line with previous findings on the diet and capture rates (maximum and minimum) observed in annual cycles in the three studied species [
30,
34,
41], however several peaks of ingesta have been recorded that in seasonal sampling are difficult to detect. These peaks demonstrate that, in only few hours, the energy input may be even more than an order of magnitude higher in species like
P. clavata, but also noteworthy in
E. singularis and
L. sarmentosa.
The present high-frequency monitoring does not show a clear coupling between the activity and the seston concentration. It represents, however, a first insight into the complex mechanisms among the seston variables (concentration), the polyp activity and the species diet. The seston concentration (or quality, see the protein concentration) was not clearly related to the gut contents even taking both variables contextually (seston and polyps) through a high-frequency sampling. Rossi et al. [
15] showed that the polyp opening in another octocoral,
Corallium rubrum, due to the presence of a nutritional stimulus (chemical signals or zooplankton) in controlled experiments, may be 1–3 minutes. The activity (opening polyps) of the three gorgonians studied though a field experiment seems not to be affected to the seston concentration or quality since the elapsed time between the high signals (i.e., high zooplankter concentration for example) and the polyp opening may happen quickly and non-detectable even with such an intensive cycle. At this temperature (19.5 °C), the three species seem to follow similar patterns to those found in previous studies of these or other octocorals [
15,
56]. However, the present observations highlight that the fed polyps are often in coincidence with high concentration of seston and zooplankton density. Benthic macrofaunal response to pelagic production can cover different time ranges according to physiology and ecology of the considered species, as also underlined by observations and model simulation [
57].
Digestion time, being relatively slow at 19.5 °C (the temperature found in the water column in the area during the sampling [
10]), may be essential to understand the underlining trophic dynamics. The prey abundance in gut contents could reflect the preys caught nine hours before. In fact, sudden changes of the seston concentration in only few hours are well-known [
10,
11], representing the opportunity to retain the particles limited in time for benthic suspension feeders [
51].
Our findings report a clear difference among the investigated species in the food pulse behavior in term of capture rates (and in different times of the cycle).
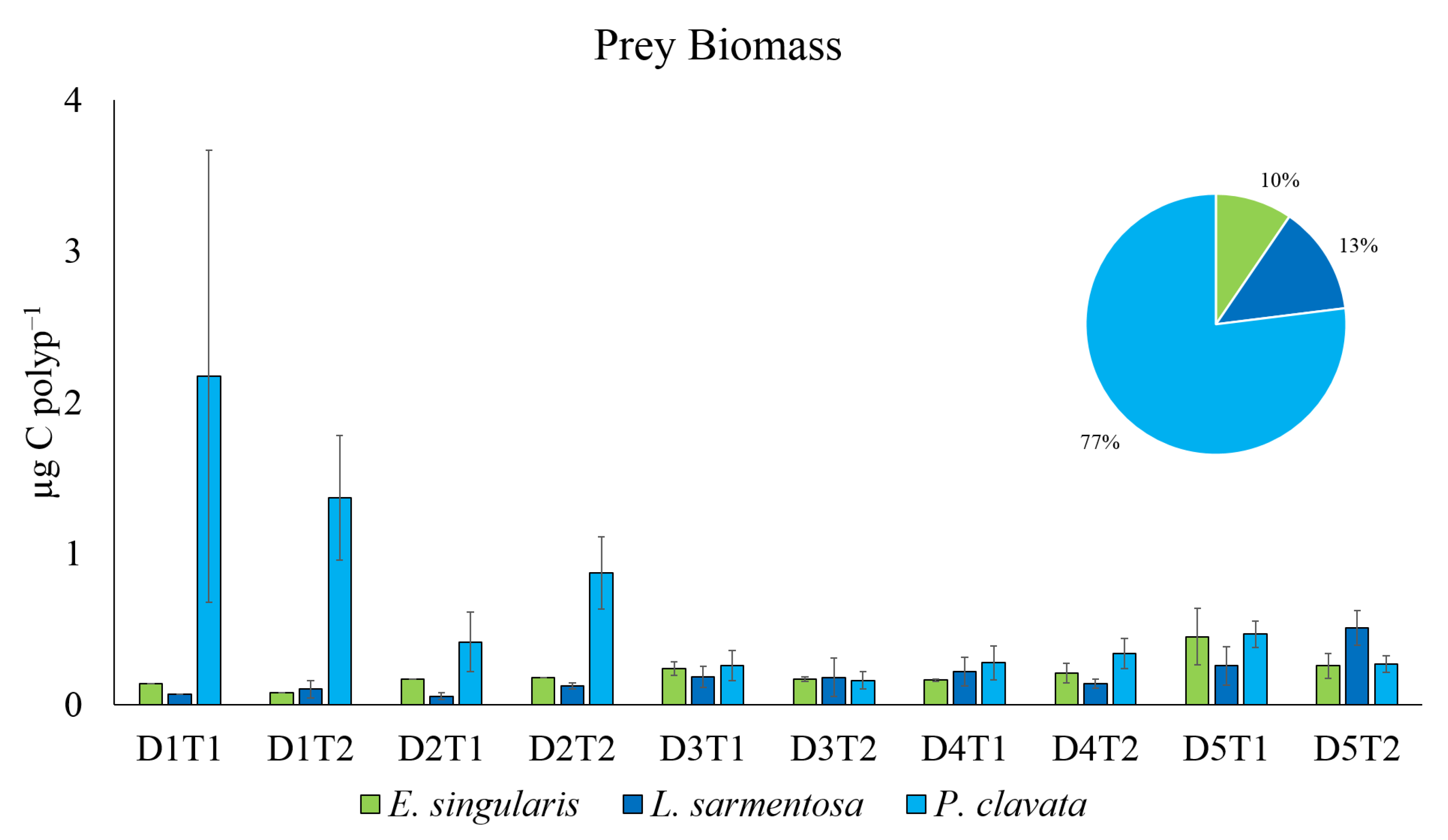
Paramuricea clavata has the highest prey abundance in gut contents that, transformed in captured organic carbon, indicates the highest energy input among the three species. In the study area,
P. clavata lives on vertical rocky walls, as reported in other studies [
36], on a large boulder [
58], forming crowded patches (around 56 colonies m
−2, [
30]). The density of colony patches of
E. singularis was lower than
P. clavata in the study area (41 colonies m
−2, [
59]), and sparse colonies were found in the case of
L. sarmantosa (1.5 colonies m
−2, [
41]). Nelson and Bramanti [
27] suggested that the density and population structure/ramification of the gorgonian forests can be crucial to understand the efficient capture rates, since the high-density patches are able to retain the particles for longer times potentially [
5]. Coma et al [
60] found that, in the rocky coasts of Menorca, after a mass mortality event, high-density patches of
E. singularis were affected by lower mortality rates when compared with patches hosting low-density colonies. This species relies on the autotrophic input to be successful in shallow waters [
35,
38]), with low number of longer primary branches in the colonies respect those found in deeper waters [
38].
The capture rates of
E. singularis are comparable with those of
L. sarmentosa: a poor branching pattern possibly structured for light harvesting [
61] and a non-effective cnidocilia density in the polyps [
34] may be essential to understand why the numbers of incoming carbon by heterotrophy are not similar to those of
P. clavata. Rossi et al. [
25] also suggested that the patch density in mixotrophic species may be one of the keys to understanding colony efficiency in the energy inputs. The species studied (
Pterogorgia anceps, Caribbean Sea) had similar capture rates to
E. singularis.
Leptogorgia sarmentosa, being in the study area the least dense among the three species investigated (1.5 colonies m
−2 [
41]) is highly branched [
62]. Thus, its capture rates are quite high if we compare with the crowded colonies of
P. clavata. It suggests that indeed the colony density can be essential to be largely efficient capturing particles in gorgonians, together with the exposition of the colonies [
58]. Further investigations applying high-frequency temporal approaches at large spatial scales will compare different ranges of the density of
L. sarmentosa to assess the capture efficiency of this sand-gravel suspension feeder. The observations on
P. clavata were made in a vertical wall exposed to the currents, a preferential settlement of the species [
36]. This position can enhance the prey capture rates, making this species particularly efficient in seston retention. We suggest that, beyond the exposition of the patch (vertical instead of horizontal [
58]), and polyp size and morphology [
63], the density of the colonies [
27] represent key factors to understand the optimization of the prey capture rates. Even if the final target of the present experimental design was not to demonstrate the effect of the patch density in the beginning, our results shed a first light on the relationship between the colony density and branching pattern and the capture rates in the intensive cycle. Campanyà-Llovet et al. [
20] stressed that food quantity and quality will be essential to understand the future of mega-benthic communities in different areas of the world. Indeed, the synergy of currents and seston concentration may be the key to understanding prey capture rates in passive suspension feeders [
15]. However, as also suggested by Nelson and Bramanti [
27], other relevant factors as patch density, position, colony branching pattern, and the mixotrophic or heterotrophic nature of the gorgonians can affect the efficiency in these capture rates with associated consequences on carbon cycling and immobilization [
64,
65]. Rossi et al. [
21] also highlighted this issue, stressing the importance of studies on the effects of primary and secondary productivity changes driven by climate change, which will shape the future seascape.











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}