
Effects of In-Channel Structure on Chinook Salmon Spawning Habitat and Embryo Production


Abstract
:1. Introduction
2. Materials and Methods
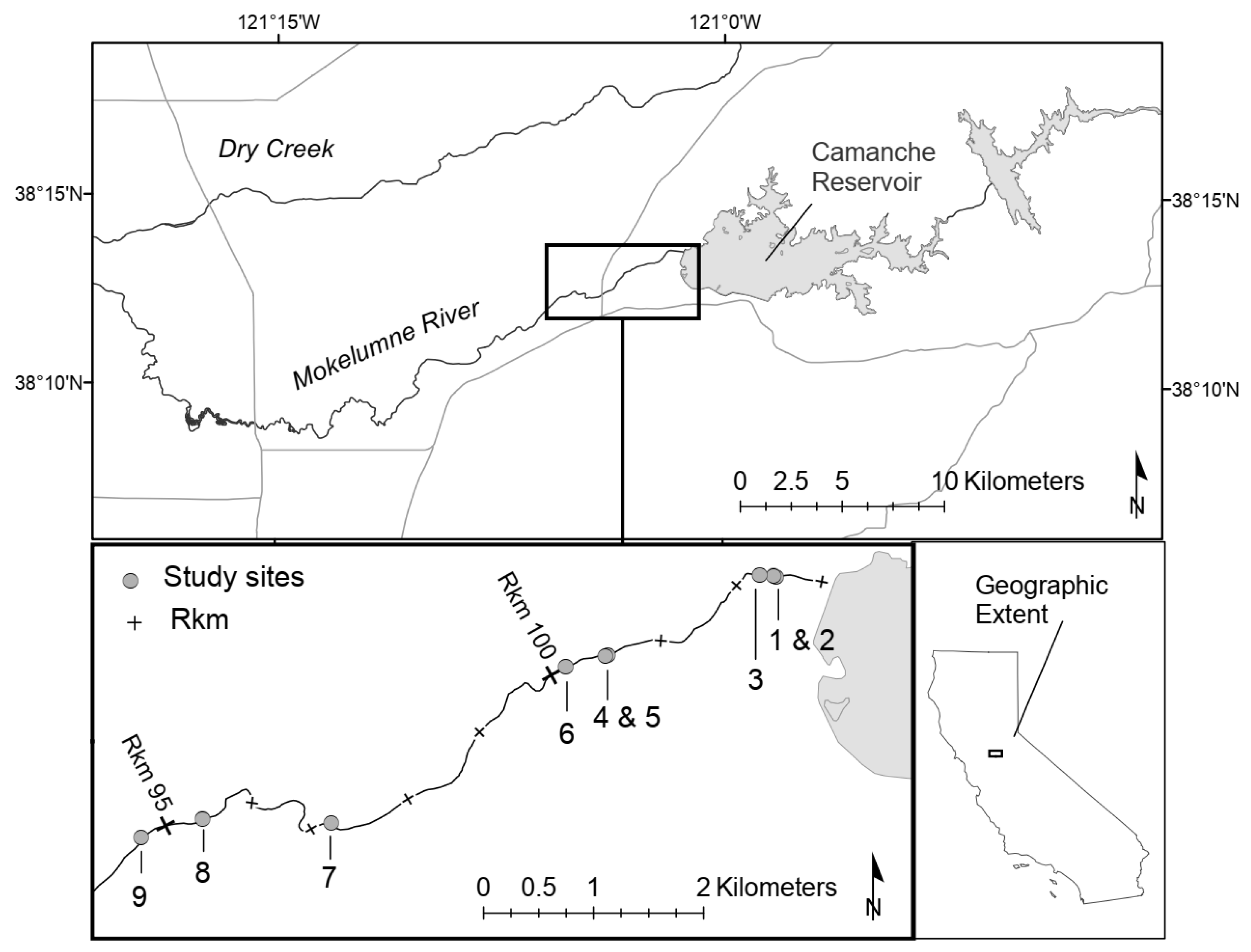
2.1. Study Area
2.2. Site Selection
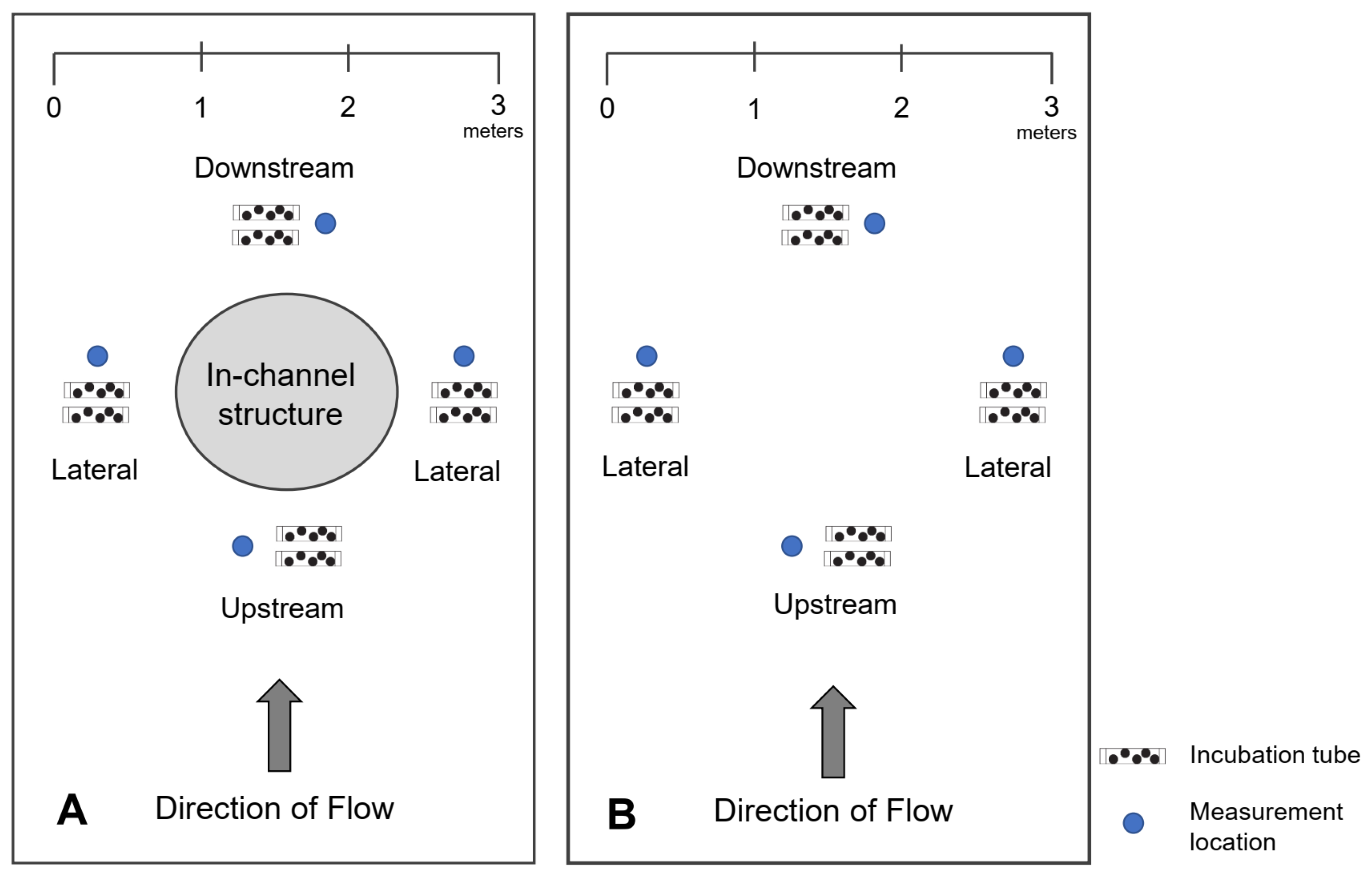
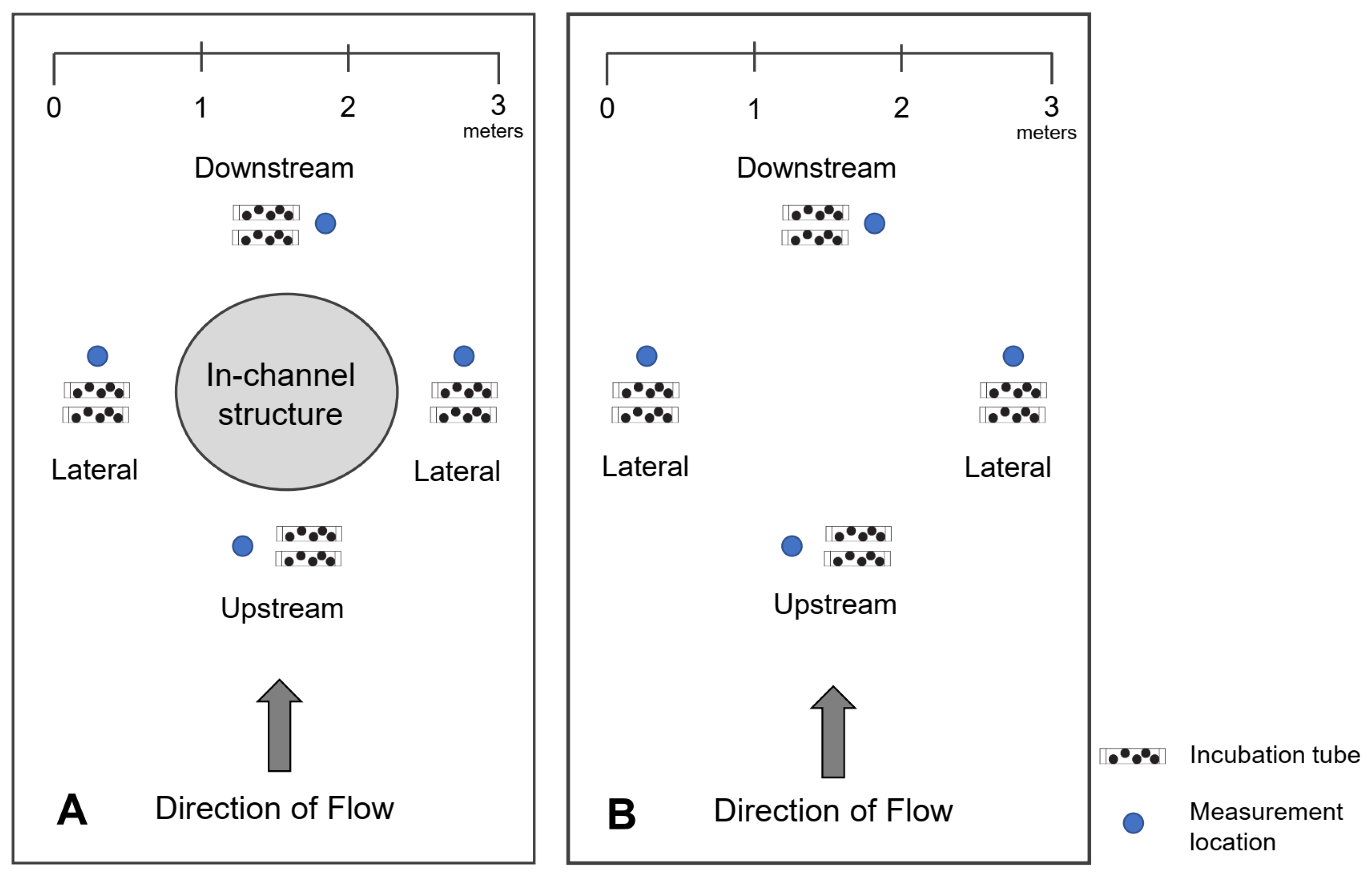
2.3. Hydraulics—Surface Water Depth and Velocity and River Flow
2.4. Hyporheic Flow (Vertical Hydraulic Gradient)
2.5. Hyporheic and Surface Water Quality
2.6. Embryo Survival, Development, and Growth
2.7. Statistical Analyses
3. Results
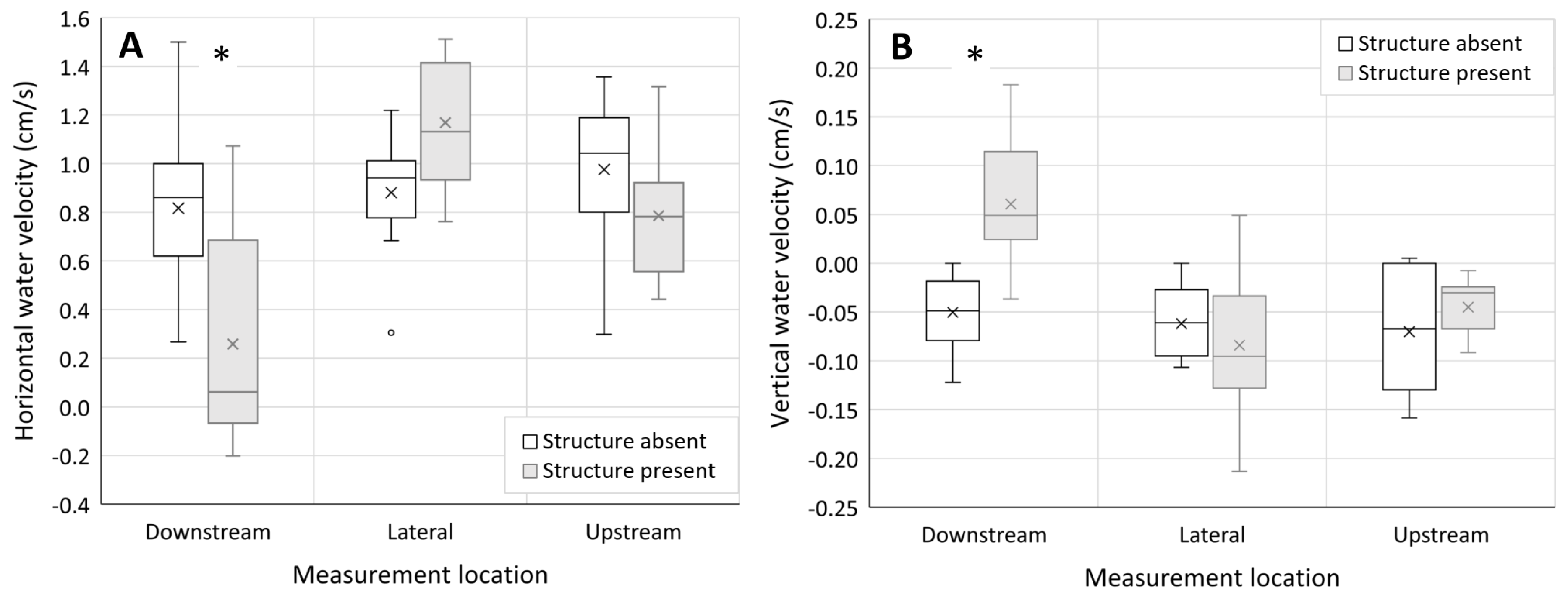
3.1. Hydraulics—Surface Water Velocity
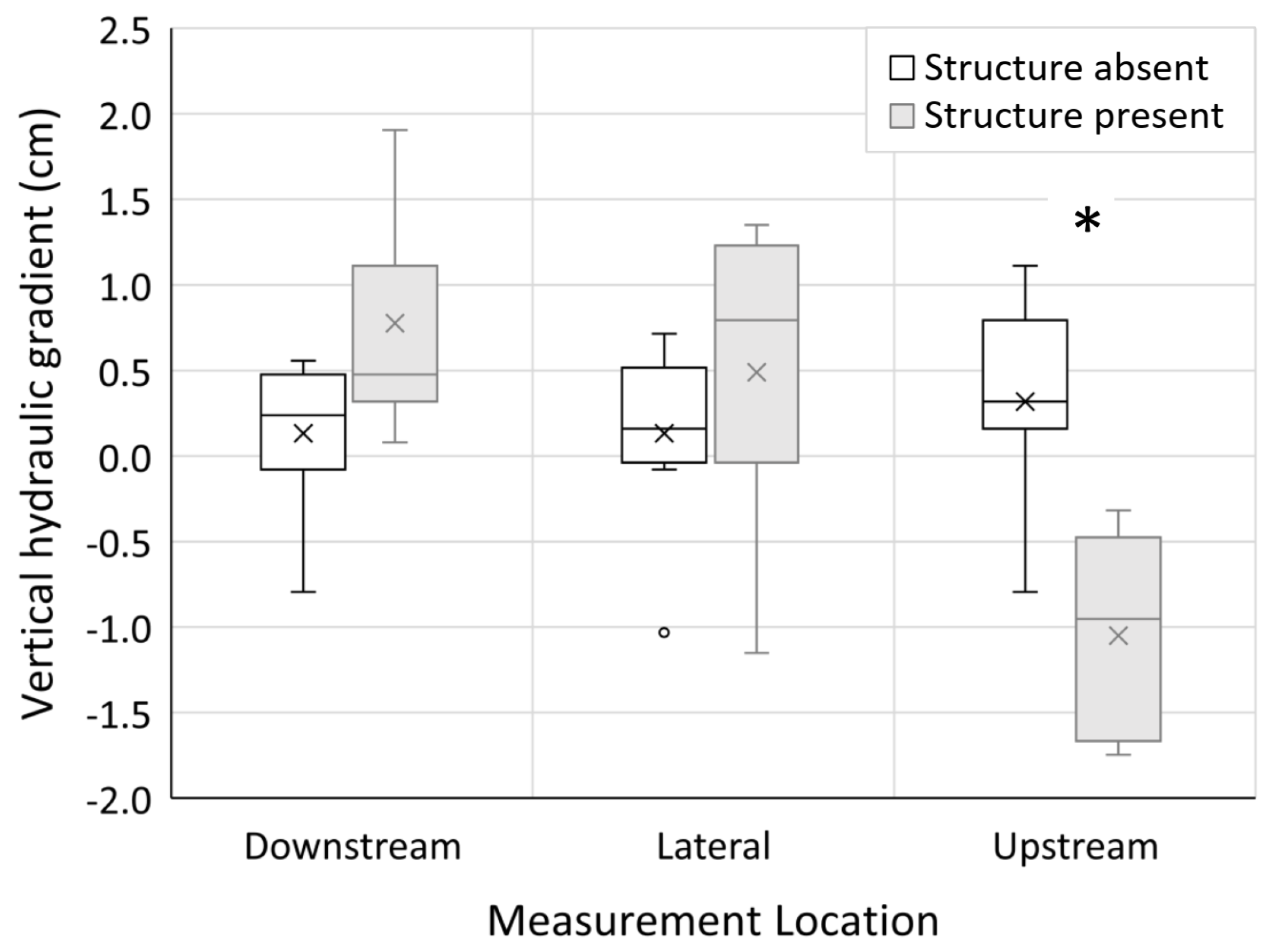
3.2. Hyporheic Flow
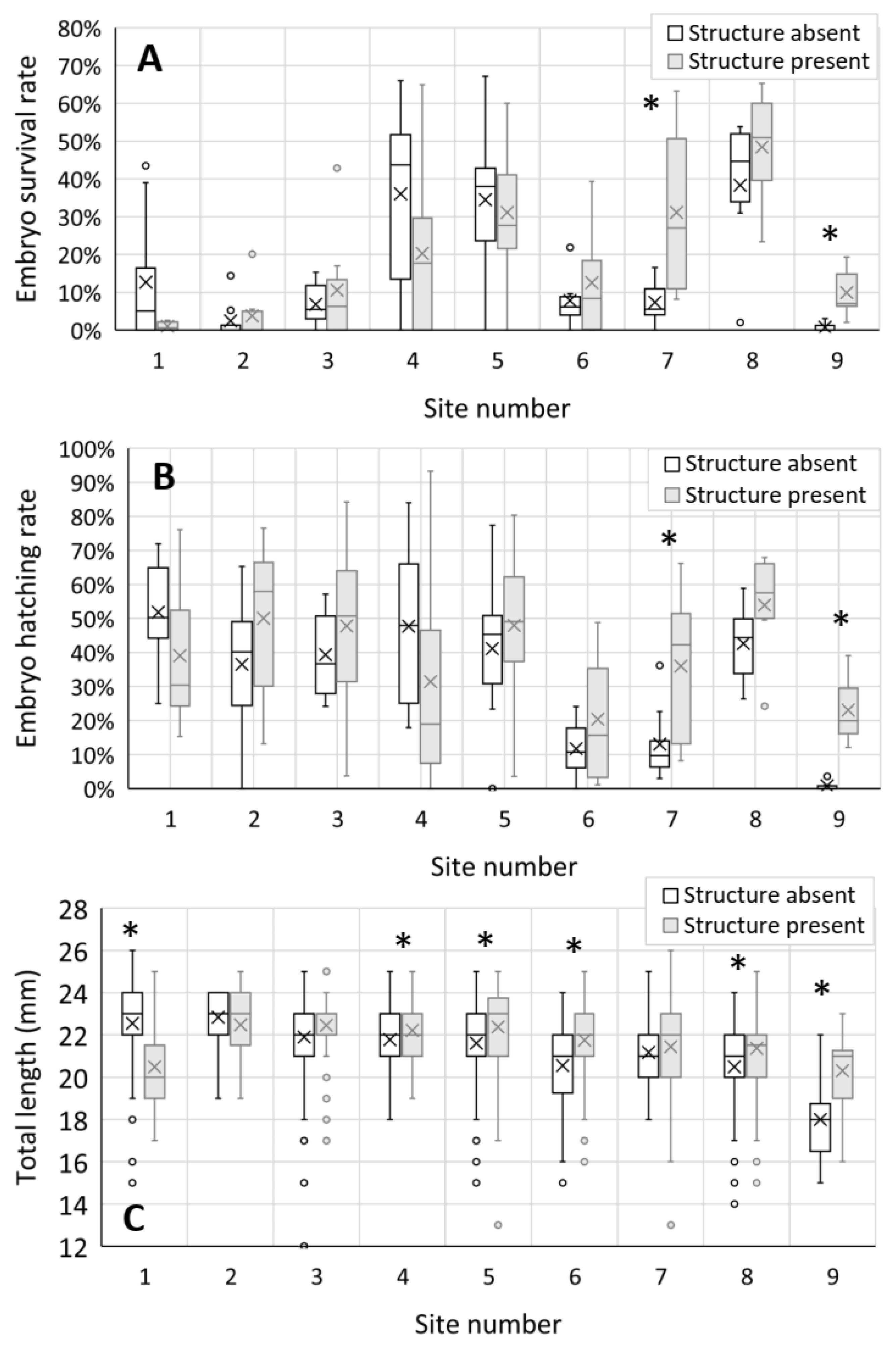
3.3. Embryo Survival, Development, and Growth
3.4. Physical and Chemical Conditions Associated with Embryo Survival, Development, and Growth
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bravard, J.; Amoros, C.; Pautou, G.; Bornette, G.; Bournaud, M.; Creuze Des Chantelliers, M.; Gilbert, J.; Peiry, J.; Perrin, J.; Tachet, H. River incision in south-east France: Morphological phenomena and ecological effects. Regul. Rivers Res. Manag. 1997, 13, 75–90. [Google Scholar] [CrossRef]
- House, R.A.; Boehne, P.L. Effects of instream structures on salmonid habitat and populations in Tobe Creek, Oregon. N. Am. J. Fish. Manag. 1986, 6, 38–46. [Google Scholar] [CrossRef]
- Langler, G.J.; Smith, C. Effects of habitat enhancement on 0-group fishes in a lowland river. Regul. Rivers Res. Manag. 2001, 17, 677–686. [Google Scholar] [CrossRef]
- Buffington, J.M.; Montgomery, D.R. Effects of hydraulic roughness on surface textures of gravel-bed rivers. Water Resour. Res. 1999, 35, 3507–3522. [Google Scholar] [CrossRef]
- Negishi, J.N.; Richardson, J.S. Responses of organic matter and macroinvertebrates to placements of boulder clusters in a small stream of southwestern British Columbia, Canada. Can. J. Fish. Aquat. Sci. 2003, 60, 247–258. [Google Scholar] [CrossRef]
- House, R. An evaluation of stream restoration structures in a coastal Oregon stream, 1981–1993. N. Am. J. Fish. Manag. 1996, 16, 272–281. [Google Scholar] [CrossRef]
- Gurnell, A.M.; Petts, G.E. Island-dominated landscapes of large floodplain rivers, a European perspective. Freshw. Biol. 2002, 47, 581–600. [Google Scholar] [CrossRef]
- Merz, J.E.; Pasternack, G.B.; Wheaton, J.M. Sediment budget for salmonid spawning habitat rehabilitation in a regulated river. Geomorphology 2006, 76, 207–228. [Google Scholar] [CrossRef]
- Harrison, L.R.; Keller, E.A. Modeling forced pool-riffle hydraulics in a boulder-bed stream, southern California. Geomorphology 2007, 83, 232–248. [Google Scholar] [CrossRef]
- Yarnell, S.M.; Mount, J.F.; Larsen, E.W. The influence of relative sediment supply on riverine habitat heterogeneity. Geomorphology 2006, 80, 310–324. [Google Scholar] [CrossRef]
- Abbe, T.B.; Montgomery, D.R. Large woody debris jams, channel hydraulics and habitat formation in large rivers. Regul. Rivers Res. Manag. 1996, 12, 201–221. [Google Scholar] [CrossRef]
- Faustini, J.M.; Jones, J.A. Influence of large woody debris on channel morphology and dynamics in steep, boulder-rich mountain streams, western Cascades, Oregon. Geomorphology 2003, 51, 187–205. [Google Scholar] [CrossRef]
- Fausch, K.D. Experimental analysis of microhabitat selection by juvenile steelhead (Oncorhynchus mykiss) and coho salmon (O. kisutch) in a British Columbia stream. Can. J. Fish. Aquat. Sci. 1993, 50, 1198–1207. [Google Scholar] [CrossRef]
- Lonzarich, D.G.; Quinn, T.P. Experimental evidence for the effect of depth and structure on the distribution, growth, and survival of stream fishes. Can. J. Zool. 1995, 73, 2223–2230. [Google Scholar] [CrossRef] [Green Version]
- Shirvell, C.S. Role of instream rootwads as juvenile coho salmon (Oncorhynchus kisutch) and steelhead trout (O. mykiss) cover habitat under varying streamflows. Can. J. Fish. Aquat. Sci. 1990, 47, 852–861. [Google Scholar] [CrossRef]
- Hafs, A.; Harrison, L.; Utz, R.; Dunne, T. Quantifying the role of woody debris in providing bioenergetically favorable habitat for juvenile salmon. Ecol. Model. 2014, 285, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Buffington, J.M.; Montgomery, D.R.; Greenberg, H.M. Basin-scale availability of salmonid spawning gravel as influenced by channel type and hydraulic roughness in mountain catchments. Can. J. Fish. Aquat. Sci. 2004, 61, 2085–2096. [Google Scholar] [CrossRef]
- Merz, J.E. Association of fall-run Chinook salmon redds with woody debris in the lower Mokelumne River, California. Calif. Fish Game 2001, 87, 51–60. [Google Scholar]
- Senter, A.E.; Pasternack, G.B. Large wood aids spawning Chinook salmon (Oncorhynchus tshawytscha) in marginal habitat on a regulated river in California. River Res. Appl. 2011, 27, 550–565. [Google Scholar] [CrossRef]
- MacInnis, C.; Floyd, T.; Taylor, B. Large woody debris structures and their influence on Atlantic salmon spawning in a stream in Nova Scotia, Canada. N. Am. J. Fish. Manag. 2008, 28, 781–791. [Google Scholar] [CrossRef]
- Wheaton, J.; Pasternack, G.; Merz, J. Use of habitat heterogeneity in salmonid spawning habitat rehabilitation design. In Fifth International Symposium on Ecohydraulics: Aquatic habitats: Analysis & Restoration; Garcia, D., Martinez, P., Eds.; IAHR-AIRH: Madrid, Spain, 2004; pp. 791–796. [Google Scholar]
- Thibodeaux, L.J.; Boyle, J.D. Bedform-generated convective transport in bottom sediment. Nature 1987, 325, 341–343. [Google Scholar] [CrossRef]
- Tonina, D.; Buffington, J.M. Hyporheic exchange in gravel bed rivers with pool-riffle morphology: Laboratory experiments and three-dimensional modeling. Water Resour. Res. 2007, 43, W01421. [Google Scholar] [CrossRef] [Green Version]
- Tonina, D.; Buffington, J.M. Hyporheic exchange in mountain rivers I: Mechanics and environmental effects. Geogr. Compass 2009, 3, 1063–1086. [Google Scholar] [CrossRef]
- Hendricks, S.P.; White, D.S. Hummocking by lotic Chara: Observations on alterations of hyporheic temperature patterns. Aquat. Bot. 1988, 31, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Crispell, J.K.; Endreny, T.A. Hyporheic exchange flow around constructed in-channel structures and implications for restoration design. Hydrol. Processes 2009, 23, 1158–1168. [Google Scholar] [CrossRef]
- Esteve, M. Observations of spawning behaviour in Salmoninae: Salmo, Oncorhynchus and Salvelinus. Rev. Fish Biol. Fish. 2005, 15, 1–21. [Google Scholar] [CrossRef]
- Youngson, A.F.; Malcolm, I.A.; Thorley, J.L.; Bacon, P.J.; Soulsby, C. Long- residence groundwater effects on incubating salmonid eggs: Low hyporheic oxygen impairs embryo development. Can. J. Fish. Aquat. Sci. 2004, 60, 2278–2287. [Google Scholar] [CrossRef]
- Malcolm, I.A.; Soulsby, C.; Youngston, A.F.; Hannah, D.M. Catchment-scale controls on groundwater-surface water interactions in the hyporheic zone: Implications for salmon embryo survival. River Res. Appl. 2005, 21, 977–989. [Google Scholar] [CrossRef]
- Mouw, J.; Tappenbeck, T.; Stanford, J. Spawning tactics of summer chum salmon Oncorhynchus keta in relation to channel complexity and hyporheic exchange. Environ. Biol. Fishes 2014, 97, 1095–1107. [Google Scholar] [CrossRef]
- Geist, D.R.; Hanrahan, T.P.; Arntzen, E.V.; McMichael, G.A.; Murray, C.J.; Chien, Y. Physicochemical characteristics of the hyporheic zone affect redd site selection by chum salmon and fall Chinook salmon in the Columbia River. N. Am. J. Fish. Manag. 2002, 22, 1077–1085. [Google Scholar] [CrossRef]
- Gordon, R.P.; Lautz, L.K.; Daniluk, T.L. Spatial patterns of hyporheic exchange and biogeochemical cycling around cross-vane restoration structures: Implications for stream restoration design. Water Resour. Res. 2013, 49, 2040–2055. [Google Scholar] [CrossRef]
- Hester, E.T.; Doyle, M.W. In-stream geomorphic structures as drivers of hyporheic exchange. Water Resour. Res. 2008, 44, W03417. [Google Scholar] [CrossRef]
- Sawyer, A.H.; Bayani Cardenas, M.; Buttles, J. Hyporheic exchange due to channel-spanning logs. Water Resour. Res. 2011, 47, W08502. [Google Scholar] [CrossRef]
- Kondolf, G.M. Geomorphological Observations on Spawning Gravel, Channel Change, and Riparian Vegetation on the Lower Mokelumne River, California; Report to the Environmental Sciences Division, Oak Ridge National Laboratory: Oak Ridge, TN, USA, 1993.
- Pasternack, G.B.; Wang, C.L.; Merz, J.E. Application of a 2D hydrodynamic model to design of reach-scale spawning gravel replenishment on the Mokelumne River, California. River Res. Appl. 2004, 20, 205–225. [Google Scholar] [CrossRef]
- Merz, J.E. Diet of juvenile fall-run Chinook salmon in the lower Mokelumne River, California. Calif. Fish Game 2001, 87, 102–111. [Google Scholar]
- Hurlburt, S.H. Pseudoreplication and the design of ecological field experiments. Ecol. Monogr. 1984, 54, 187–211. [Google Scholar] [CrossRef] [Green Version]
- Ahearn, D.S.; Dahlgren, R.A. Sediment and nutrient dynamics following a low-head dam removal at Murphy Creek, California. Limnol. Oceanogr. 2005, 50, 1752–1762. [Google Scholar] [CrossRef]
- Merz, J.E.; Setka, J.D.; Pasternack, G.B.; Wheaton, J.M. Predicting benefits of spawning-habitat rehabilitation to salmonid (Oncorhynchus spp.) fry production in a regulated California river. Can. J. Fish. Aquat. Sci. 2004, 61, 1433–1446. [Google Scholar] [CrossRef] [Green Version]
- Washburn, K.; Kratville, D.; Merz, J. Assessing the Distribution, Density, and Potential Impacts of Potamopyrgus antipodarum on Resident Aquatic Invertebrate Assemblages in the Lower Mokelumne River, California; Report to the United States Fish and Wildlife Service: Lodi, CA, USA, 2007.
- Merz, J.E.; Setka, J.D. Evaluation of a spawning habitat enhancement site for Chinook salmon in a regulated California river. N. Am. J. Fish. Manag. 2004, 24, 397–407. [Google Scholar] [CrossRef]
- Cederholm, C.J.; Bilby, R.E.; Bisson, P.A.; Bumstead, T.W.; Fransen, B.R.; Scarlett, W.J.; Ward, J.W. Responses of juvenile coho salmon and steelhead to placement of large woody debris in a coastal Washington stream. N. Am. J. Fish. Manag. 1997, 17, 947–963. [Google Scholar] [CrossRef]
- Biron, P.M.; Robson, C.; LaPointe, M.F.; Gaskin, S.J. Three-dimensional flow dynamics around deflectors. River Res. Appl. 2005, 21, 961–975. [Google Scholar] [CrossRef]
- California Data Exchange Center. Sacramento (CA): Department of Water Resources. 2021. Available online: http://cdec.water.ca.gov/ (accessed on 13 December 2021).
- Horner, T.C. Site Assessment at the Lower Sunrise Side Channel: Surface Water, Pore Water, and Intergravel Flow; Report to the Sacramento Water Forum: Sacramento, CA, USA, 2005.
- Devries, P. Riverine salmonid egg burial depths: Review of published data and implications for scour studies. Can. J. Fish. Aquat. Sci. 1997, 54, 1685–1698. [Google Scholar] [CrossRef]
- Kondolf, G.; Williams, J.; Horner, T.; Milan, D. Assessing physical quality of spawning habitat. Am. Fish. Soc. Symp. 2008, 65, 249–274. [Google Scholar]
- Silver, S.J.; Warren, C.E.; Doudoroff, P. Dissolved oxygen requirements of developing steelhead trout and Chinook salmon embryos at different water velocities. Trans. Am. Fish. Soc. 1963, 92, 327–343. [Google Scholar] [CrossRef]
- Geen, G.H.; Neilson, J.D.; Bradford, M. Effects of pH on the early development and growth and otolith microstructure of Chinook salmon, Oncorhynchus tshawytscha. Can. J. Zool. 1985, 63, 22–27. [Google Scholar] [CrossRef]
- Beacham, T.D.; Murray, C.B. Temperature, egg size, and development of embryos and alevins of five species of Pacific salmon: A comparative analysis. Trans. Am. Fish. Soc. 1990, 119, 927–945. [Google Scholar] [CrossRef]
- Greig, S.M.; Sear, D.A.; Carling, P.A. A review of factors influencing the availability of dissolved oxygen to incubating salmonid embryos. Hydrol. Processes 2007, 21, 323–334. [Google Scholar] [CrossRef]
- Malcolm, I.A.; Youngson, A.F.; Soulsby, C. Survival of salmonid eggs in a degraded gravel-bed stream: Effects of groundwater-surface water interactions. River Res. Appl. 2003, 19, 303–316. [Google Scholar] [CrossRef]
- Leitritz, E.; Lewis, R.C. California Department of Fish and Game Fish Bulletin 164 Trout and Salmon Culture (Hatchery Methods); University of California: San Diego, CA, USA, 1976; pp. 160–167. [Google Scholar]
- Murray, C.B.; McPhail, J.D. Effect of incubation temperature on the development of five species of Pacific salmon (Oncorhynchus) embryos and alevins. Can. J. Zool. 1988, 66, 266–273. [Google Scholar] [CrossRef]
- Lapointe, M.F.; Bergeron, N.E.; Berube, F.; Pouliot, M.A.; Johnston, P. Interactive effects of substrate, sand and silt contents, redd-scale hydraulic gradients, and interstitial velocities on egg-to-emergence survival of Atlantic salmon (Salmo salar). Can. J. Fish. Aquat. Sci. 2004, 61, 2271–2277. [Google Scholar] [CrossRef] [Green Version]
- Geist, D.R.; Dauble, D.D. Redd site selection and spawning habitat use by fall Chinook salmon: The importance of geomorphic features in large rivers. Environ. Manag. 1998, 22, 655–669. [Google Scholar] [CrossRef]
- Saltveit, S.; Brabrand, A. Incubation, hatching and survival of eggs of Atlantic salmon (Salmo salar) in spawning redds influenced by groundwater. Limnologica 2013, 43, 325–331. [Google Scholar] [CrossRef]
- Kasahara, T.; Hill, A.R. Hyporheic exchange flows induced by constructed riffles and steps in lowland streams in southern Ontario, Canada. Hydrol. Processes 2006, 20, 4287–4305. [Google Scholar] [CrossRef]
- Malcolm, I.A.; Soulsby, C.; Youngson, A.F.; Hannah, D.M.; McLaren, I.S.; Thorne, A. Hydrological influences on hyporheic water quality: Implications for salmon egg survival. Hydrol. Processes 2004, 18, 1543–1560. [Google Scholar] [CrossRef]
- Coble, D.W. Influence of water exchange and dissolved oxygen in redds on survival of steelhead trout embryos. Trans. Am. Fish. Soc. 1961, 90, 469–474. [Google Scholar] [CrossRef] [Green Version]
- Shumway, D.L.; Warren, C.E.; Doudoroff, P. Influence of oxygen concentration and water movement on the growth of steelhead trout and coho salmon embryos. Trans. Am. Fish. Soc. 1964, 93, 342–356. [Google Scholar] [CrossRef]
- Del Rio, A.M.; Davis, B.E.; Fangue, N.A.; Todgham, A.E. Combined effects of warming and hypoxia on early life stage Chinook salmon physiology and development. Conserv. Physiol. 2019, 7, coy078. [Google Scholar] [CrossRef] [Green Version]
- Martin, B.; Dudley, P.; Kashef, N.; Stafford, D.; Reeder, W.; Tonina, D.; Del Rio, A.; Foott, J.; Danner, E. The biophysical basis of thermal tolerance in fish eggs. Proc. R. Soc. B Biol. Sci. 2020, 287, jeb235150. [Google Scholar] [CrossRef]
- Geist, D.; Abernethy, C.; Hand, K.; Cullinan, V.; Chandler, J. Survival, development, and growth of fall Chinook salmon embryos, alevins, and fry exposed to variable thermal and dissolved oxygen regimes. Trans. Am. Fish. Soc. 2006, 135, 1462–1477. [Google Scholar] [CrossRef]
- Groves, P.A.; Chandler, J.A. Habitat quality of historic Snake River fall Chinook salmon spawning locations and implications for incubation survival. Part 2: Intragravel water quality. River Res. Appl. 2005, 21, 469–483. [Google Scholar] [CrossRef]
- Rombough, P.J. Effects of low pH on eyed embryos and alevins of Pacific salmon. Can. J. Fish. Aquat. Sci. 1982, 40, 1575–1582. [Google Scholar] [CrossRef]
- Malcolm, I.A.; Soulsby, C.; Youngson, A.F.; Petry, J. Heterogeneity in groundwater interactions in the hyporheic zone of a salmonid spawning stream. Hydrol. Processes 2003, 17, 601–617. [Google Scholar] [CrossRef]
- Yoshiyama, R.M.; Fisher, F.W.; Moyle, P.B. Historical abundance and decline of Chinook salmon in the Central Valley region of California. N. Am. J. Fish. Manag. 1998, 18, 487–521. [Google Scholar] [CrossRef]
- Yoshiyama, R.M.; Gerstung, E.R.; Fisher, F.W.; Moyle, P.B. Chinook salmon in the California Central Valley: An assessment. Fisheries 2000, 25, 6–20. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site Number | Form of In-Channel Structure | River Kilometer | Year(s) Enhanced |
|---|---|---|---|
| 1 | 1 Boulder | 102.5 | 1999, 2005 |
| 1 | Control | 102.5 | 1999, 2005 |
| 2 | 1 LWD | 102.4 | 1999, 2005 |
| 2 | Control | 102.4 | 1999, 2005 |
| 3 | 1 Boulder | 102.3 | 1992, 1993, 2006 |
| 3 | Control | 102.2 | 1992, 1993, 2006 |
| 4 | 2 Boulders | 100.5 | 2002 |
| 4 | Control | 100.5 | 2002 |
| 5 | 2 Boulders | 100.4 | 2002 |
| 5 | Control | 100.4 | 2002 |
| 6 | 1 LWD | 100.1 | None |
| 6 | Control | 100.3 | None |
| 7 | 1 LWD | 97.2 | None |
| 7 | Control | 97.2 | None |
| 8 | 1 LWD | 95.4 | None |
| 8 | Control | 95.4 | None |
| 9 | 1 Boulder | 94.8 | 1997 |
| 9 | Control | 94.8 | 1997 |
| Source | Sum of Squares | df | F | p-Value |
|---|---|---|---|---|
| Vertical surface water velocity | ||||
| Main effects | ||||
| Site number | 0.045 | 8 | 1.93 | 0.089 |
| Structure presence | 0.019 | 1 | 6.70 | 0.014 |
| Measurement location | 0.061 | 2 | 10.56 | 0.000 |
| Interactions | ||||
| Structure presence · measurement location | 0.041 | 2 | 7.05 | 0.003 |
| Structure presence · site number | 0.018 | 8 | 0.76 | 0.641 |
| Error | 0.093 | 32 | ||
| Horizontal surface water velocity | ||||
| Main effects | ||||
| Site number | 2.836 | 8 | 5.59 | 0.000 |
| Structure presence | 0.317 | 1 | 5.00 | 0.032 |
| Measurement location | 2.260 | 2 | 17.83 | <0.001 |
| Interactions | ||||
| Structure presence · measurement location | 1.618 | 2 | 12.76 | <0.001 |
| Structure presence · site number | 0.580 | 8 | 1.14 | 0.362 |
| Error | 2.028 | 32 | ||
| Source | Sum of Squares | df | F | p-Value |
|---|---|---|---|---|
| Directional vertical hydraulic gradient | ||||
| Main effects | ||||
| Site number | 2.936 | 8 | 1.72 | 0.131 |
| Structure presence | 0.201 | 1 | 0.94 | 0.339 |
| Measurement location | 6.909 | 2 | 16.20 | <0.0001 |
| Interactions | ||||
| Structure presence · measurement location | 10.648 | 2 | 24.97 | <0.0001 |
| Structure presence · site number | 8.601 | 8 | 5.04 | 0.000 |
| Error | 6.823 | 32 | ||
| Vertical hydraulic gradient magnitude | ||||
| Main effects | ||||
| Site number | 0.883 | 8 | 0.89 | 0.540 |
| Structure presence | 1.662 | 1 | 13.33 | 0.001 |
| Measurement location | 0.500 | 2 | 2.00 | 0.151 |
| Interactions | ||||
| Structure presence · measurement location | 0.004 | 2 | 0.01 | 0.986 |
| Structure presence · site number | 8 | 2.74 | 0.020 | |
| Error | 3.991 | 32 | ||
| Site | S | Z | p-Value |
|---|---|---|---|
| Embryo survival | |||
| 1 | 34.5 | −1.344 | 0.179 |
| 2 | 72.0 | 0.447 | 0.655 |
| 3 | 68.0 | 0.000 | 1.000 |
| 4 | 45.5 | −1.168 | 0.243 |
| 5 | 63.5 | −0.420 | 0.674 |
| 6 | 39.0 | 0.000 | 1.000 |
| 7 | 93.0 | 2.573 | 0.010 |
| 8 | 35.0 | −0.929 | 0.353 |
| 9 | 44.0 | 2.523 | 0.012 |
| Embryo hatching rate | |||
| 1 | 36.0 | −1.097 | 0.273 |
| 2 | 78.0 | 0.998 | 0.318 |
| 3 | 77.0 | 0.893 | 0.372 |
| 4 | 45.0 | −1.216 | 0.224 |
| 5 | 75.0 | 0.683 | 0.495 |
| 6 | 43.0 | 0.560 | 0.575 |
| 7 | 91.0 | 2.363 | 0.018 |
| 8 | 34.0 | −1.071 | 0.284 |
| 9 | 45.0 | 2.710 | 0.007 |
| Embryo total length | |||
| 1 | 212.0 | −2.307 | 0.021 |
| 2 | 1219.0 | 0.521 | 0.603 |
| 3 | 7337.5 | −1.268 | 0.205 |
| 4 | 32,127.5 | 2.490 | 0.013 |
| 5 | 101,812.0 | 5.948 | <0.0001 |
| 6 | 6019.5 | −4.427 | <0.0001 |
| 7 | 21,444.0 | −1.919 | 0.055 |
| 8 | 97,459.0 | −6.370 | <0.0001 |
| 9 | 102.0 | −2.356 | 0.019 |
| Sites →/ | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Parameters ↓ | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 |
| Surface water | |||||||||
| DO (mg/L) | 9.4 | 9.6 | 8 | 10.2 | 9 | 10.2 | 8.9 | 9.2 | 10.4 |
| pH | 6.6 | 6.4 | 6.7 | 6.8 | 6.7 | 7 | 6.8 | 6.8 | 6.8 |
| COND (µS/cm) | 38 | 37.6 | 37.4 | 38.3 | 38.7 | 37.9 | 38 | 37.7 | 37.9 |
| TEMP (°C) | 12.2 | 11.9 | 11.9 | 11.4 | 11.4 | 11.3 | 11.8 | 11.8 | 11.7 |
| Hyporheic water—Structure present | |||||||||
| DO (mg/L) | 8.2 (1.0) | 8.6 (0.6) | 8.1 (1.9) | 8.9 (1.6) | 9.7 (1.3) | 8.2 (2.6) | 6.7 (2.1) | 7.3 (1.0) | 8.5 (1.6) |
| pH | 6.7 (0.2) | 6.8 (0.1) | 6.7 (0.2) | 6.5 (0.2) | 6.8 (0.3) | 6.6 (0.2) | 6.5 (0.2) | 6.5 (0.1) | 6.5 (0.3) |
| COND (µS/cm) | 37.4 (0.5) | 37.5 (0.6) | 37.4 (0.6) | 38.2 (0.6) | 39.2 (2.0) | 36.5 (1.0) | 37.4 (1.9) | 37.8 (0.8) | 37.8 (0.7) |
| TEMP (°C) | 11.8 (1.0) | 12 (0.9) | 11.9 (0.9) | 11.9 (1.0) | 12 (1.0) | 11.9 (1.0) | 11.7 (1.0) | 11.2 (1.2) | 11.6 (1.0) |
| VHG (M) | 0.9 (0.7) | 1.2 (0.5) | 1 (1.1) | 1.2 (0.9) | 0.3 (0.3) | 0.8 (0.8) | 1.6 (0.7) | 0.9 (0.5) | 0.8 (0.4) |
| Hyporheic water—Structure absent | |||||||||
| DO (mg/L) | 8.5 (0.6) | 9.1 (0.7) | 8.7 (1.4) | 9.9 (0.7) | 6.1 (1.9) | 3 (2.2) | 7.8 (2.4) | 8.6 (0.6) | 6.7 (0.9) |
| pH | 6.8 (0.1) | 6.8 (0.2) | 6.8 (0.2) | 6.7 (0.2) | 6.4 (0.2) | 6.5 (0.1) | 6.2 (0.4) | 6.7 (0.1) | 6.5 (0.1) |
| COND (µS/cm) | 37.5 (0.2) | 37.7 (0.2) | 37.3 (0.2) | 38.1 (0.3) | 39.0 (1.9) | 58.5 (12.8) | 38.5 (2.7) | 37.2 (0.9) | 38.1 (1.2) |
| TEMP (°C) | 11.9 (0.9) | 12.0 (0.9) | 12.0 (0.9) | 11.9 (1.0) | 11.9 (1.0) | 11.8 (0.9) | 11.7 (1.0) | 11.6 (1.0) | 11.7 (1.1) |
| VHG (M) | 0.5 (0.5) | 0.6 (0.4) | 0.2 (0.2) | 0.5 (0.4) | 0.5 (0.2) | 0.7 (0.2) | 0.5 (0.2) | 0.7 (0.5) | 0.9 (0.2) |
| GLM | AIC | Term | Estimate | SE | Chi Square | Pr > ChiSq |
|---|---|---|---|---|---|---|
| Embryo survival | 44.650 | |||||
| Intercept | 8.523 | 10.294 | 0.546 | 0.460 | ||
| ATEMP | −0.863 | 0.877 | 0.747 | 0.387 | ||
| Embryo hatching rate | 65.842 | |||||
| Intercept | −5.461 | 11.652 | 0.234 | 0.629 | ||
| pH | 1.017 | 1.233 | 0.722 | 0.396 | ||
| ATEMP | −0.189 | 0.886 | 0.043 | 0.836 | ||
| Embryo growth | 131.528 | |||||
| Intercept | −1.772 | 8.056 | 0.048 | 0.826 | ||
| pH | 2.759 | 0.774 | 11.067 | 0.001 | ||
| SWV-V | 3.751 | 2.294 | 2.590 | 0.108 | ||
| ATEMP | 0.680 | 0.622 | 1.176 | 0.278 | ||
| COND | −0.071 | 0.034 | 4.210 | 0.040 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bilski, R.L.; Wheaton, J.M.; Merz, J.E. Effects of In-Channel Structure on Chinook Salmon Spawning Habitat and Embryo Production. Water 2022, 14, 83. https://doi.org/10.3390/w14010083
Bilski RL, Wheaton JM, Merz JE. Effects of In-Channel Structure on Chinook Salmon Spawning Habitat and Embryo Production. Water. 2022; 14(1):83. https://doi.org/10.3390/w14010083
Chicago/Turabian StyleBilski, Robyn L., Joseph M. Wheaton, and Joseph E. Merz. 2022. "Effects of In-Channel Structure on Chinook Salmon Spawning Habitat and Embryo Production" Water 14, no. 1: 83. https://doi.org/10.3390/w14010083
APA StyleBilski, R. L., Wheaton, J. M., & Merz, J. E. (2022). Effects of In-Channel Structure on Chinook Salmon Spawning Habitat and Embryo Production. Water, 14(1), 83. https://doi.org/10.3390/w14010083