
Modeling Chlorophyll a with Use of the SWAT Tool for the Nielba River (West-Central Poland) as an Example of an Unmonitored Watercourse


Abstract
:
1. Introduction
2. Materials and Methods
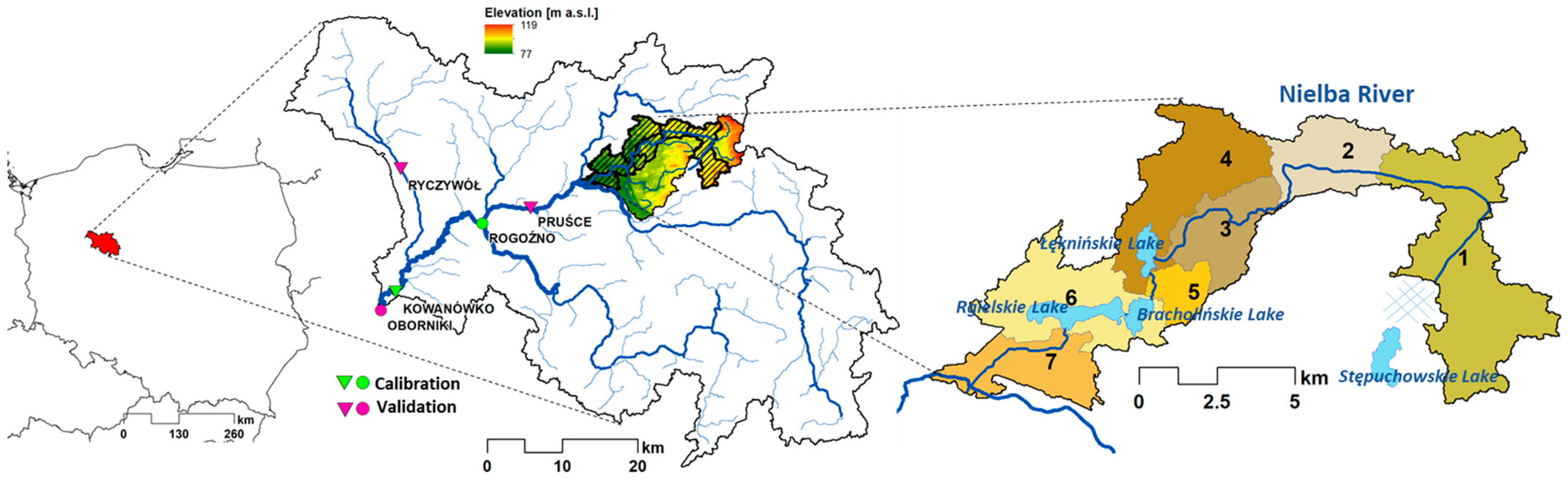
2.1. Study Area
2.2. Model Calibration and Validation
2.3. Eutrophication Parameters Simulation
2.4. Data Selection and Treatment
- Water temperature (°C);
- Solar radiation (MJ/m2);
- Surface runoff (mm);
- Chlorophyll a land delivery (µg chla/l);
- Flow (m3/s);
- Total nitrogen transported with the water into the reach (kg N/day);
- Total phosphorus transported with the water into the reach (kg P/day).
3. Results
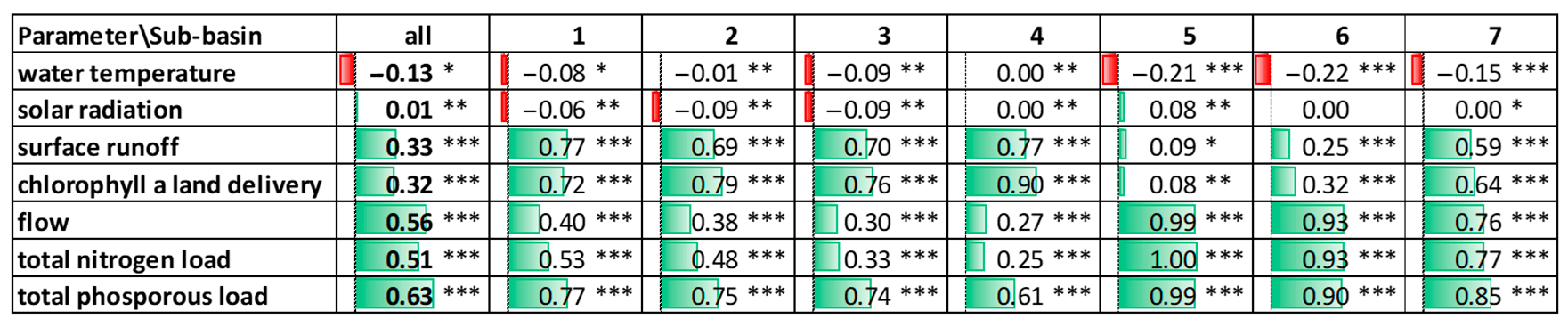
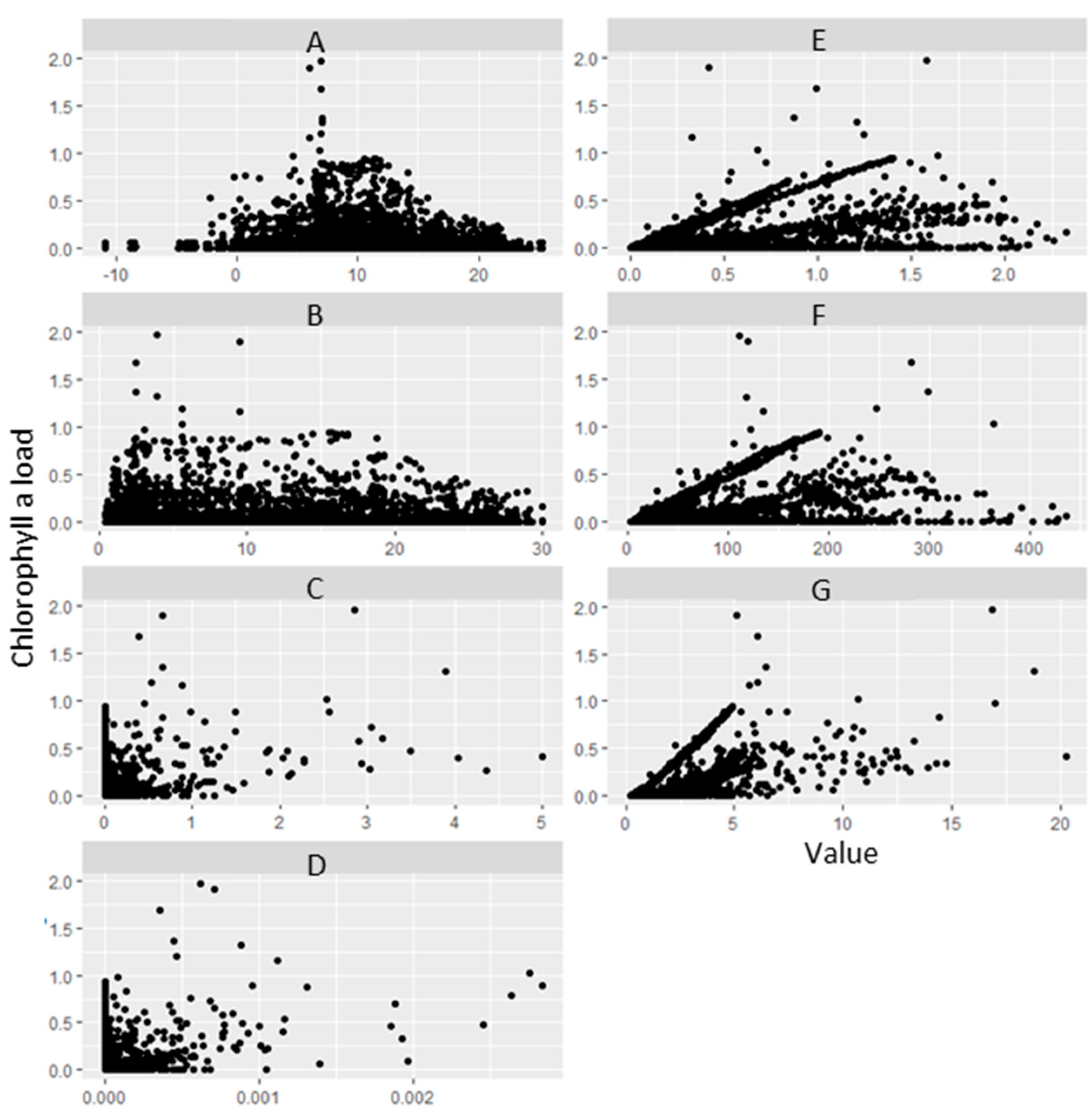
3.1. General Correlation Analysis
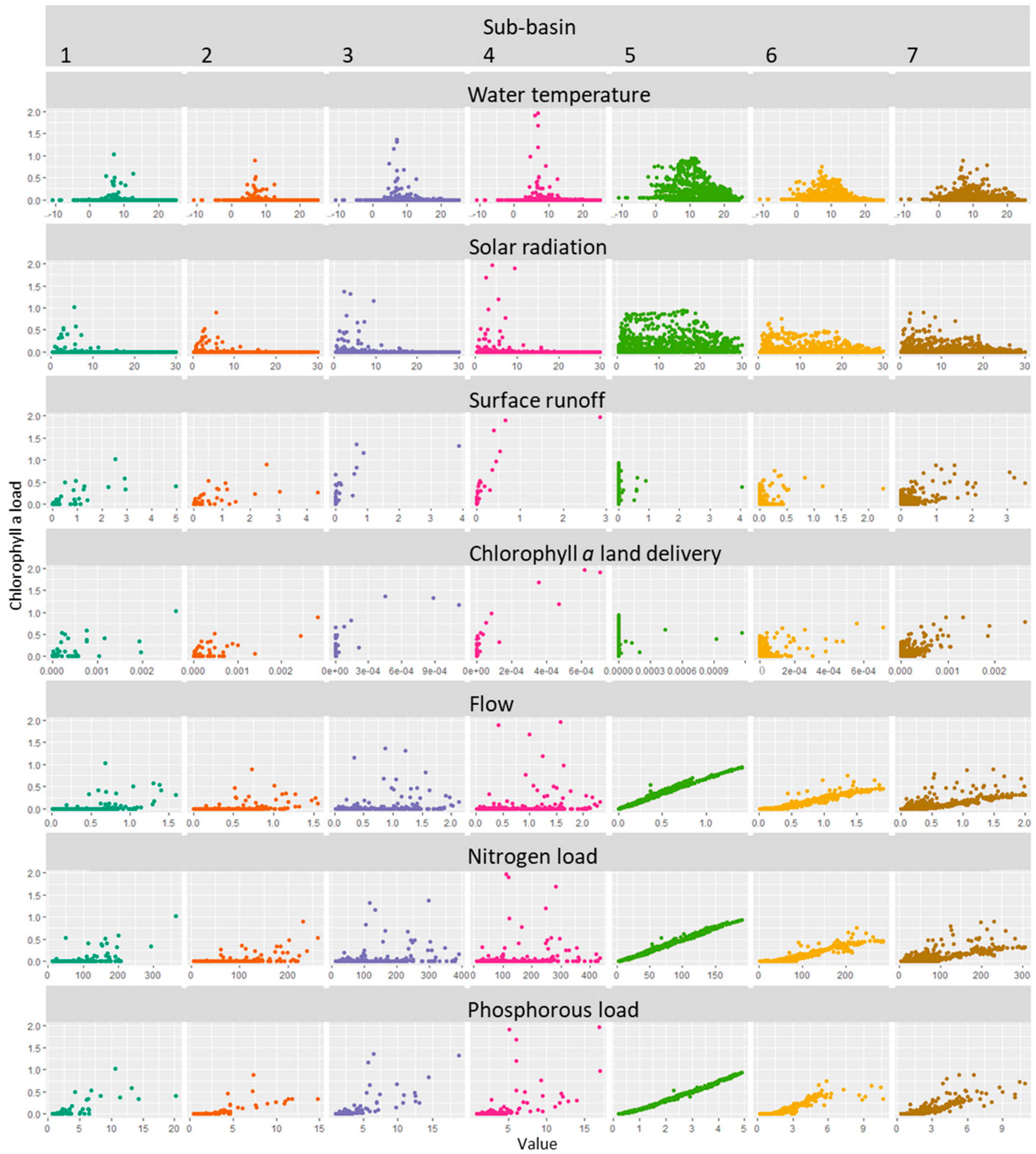
3.2. River Sections Correlation Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Boynton, W.R.; Kemp, W.M.; Keefe, C.W.A. Comparative Analysis of Nutrients and Other Factors Influencing Estuarine Phytoplankton Production. In Estuarine Comparisons; Kennedy, V.S., Ed.; Academic Press: Cambridge, MA, USA, 1982; pp. 69–90. [Google Scholar] [CrossRef]
- Cluis, D.; Couture, P.; Bégin, R.; Visser, S.A. Potential Eutrophication Assessment in Rivers; Relationship between Produced and Exported Loads. Swiss J. Hydrol. 1988, 50, 166–181. [Google Scholar] [CrossRef]
- Minaudo, C.; Meybeck, M.; Moatar, F.; Gassama, N.; Curie, F. Eutrophication Mitigation in Rivers: 30 Years of Trends in Spatial and Seasonal Patterns of Biogeochemistry of the Loire River (1980–2012). Biogeosciences 2015, 12, 2549–2563. [Google Scholar] [CrossRef] [Green Version]
- Dodds, W.K.; Smith, V.H. Nitrogen, Phosphorus, and Eutrophication in Streams. Inland Waters 2016, 6, 155–164. [Google Scholar] [CrossRef]
- Ménesguen, A.; Desmit, X.; Dulière, V.; Lacroix, G.; Thouvenin, B.; Thieu, V.; Dussauze, M. How to Avoid Eutrophication in Coastal Seas? A New Approach to Derive River-Specific Combined Nitrate and Phosphate Maximum Concentrations. Sci. Total Environ. 2018, 628–629, 400–414. [Google Scholar] [CrossRef] [Green Version]
- Le Moal, M.; Gascuel-Odoux, C.; Ménesguen, A.; Souchon, Y.; Étrillard, C.; Levain, A.; Moatar, F.; Pannard, A.; Souchu, P.; Lefebvre, A.; et al. Eutrophication: A New Wine in an Old Bottle? Sci. Total Environ. 2019, 651, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Li, R.; Han, D.; Scholz, M. Response of Eutrophication Development to Variations in Nutrients and Hydrological Regime: A Case Study in the Changjiang River (Yangtze) Basin. Water 2020, 12, 1634. [Google Scholar] [CrossRef]
- Kováčová, V. Assessment of Surface Water Eutrophication at Žitný Ostrov Region. Acta Hydrol. Slovaca 2020, 21, 65–73. [Google Scholar] [CrossRef]
- Boeykens, S.P.; Piol, M.N.; Samudio Legal, L.; Saralegui, A.B.; Vázquez, C. Eutrophication Decrease: Phosphate Adsorption Processes in Presence of Nitrates. J. Environ. Manag. 2017, 203, 888–895. [Google Scholar] [CrossRef]
- Dalu, T.; Wasserman, R.J.; Magoro, M.L.; Froneman, P.W.; Weyl, O.L.F. River Nutrient Water and Sediment Measurements Inform on Nutrient Retention, with Implications for Eutrophication. Sci. Total Environ. 2019, 684, 296–302. [Google Scholar] [CrossRef]
- Reynolds, C.S. The Ecology of Phytoplankton; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Paerl, H.W.; Valdes, L.M.; Peierls, B.L.; Adolf, J.E.; Harding, L.W., Jr. Anthropogenic and Climatic Influences on the Eutrophication of Large Estuarine Ecosystems. Limnol. Oceanogr. 2006, 51, 448–462. [Google Scholar] [CrossRef] [Green Version]
- Degobbis, D.; Precali, R.; Ivancic, I.; Smodlaka, N.; Fuks, D.; Kveder, S. Long-Term Changes in the Northern Adriatic Ecosystem Related to Anthropogenic Eutrophication. Int. J. Environ. Pollut. 2000, 13, 495–533. [Google Scholar] [CrossRef]
- Nazari-Sharabian, M.; Ahmad, S.; Karakouzian, M. Climate Change and Eutrophication: A Short Review. Eng. Technol. Appl. Sci. Res. 2018, 8, 3668–3672. [Google Scholar] [CrossRef]
- Charlton, M.B.; Bowes, M.J.; Hutchins, M.G.; Orr, H.G.; Soley, R.; Davison, P. Mapping Eutrophication Risk from Climate Change: Future Phosphorus Concentrations in English Rivers. Sci. Total Environ. 2018, 613–614, 1510–1526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krasa, J.; Dostal, T.; Jachymova, B.; Bauer, M.; Devaty, J. Soil Erosion as a Source of Sediment and Phosphorus in Rivers and Reservoirs—Watershed Analyses Using WaTEM/SEDEM. Environ. Res. 2019, 171, 470–483. [Google Scholar] [CrossRef]
- Strokal, M.; Kahil, T.; Wada, Y.; Albiac, J.; Bai, Z.; Ermolieva, T.; Langan, S.; Ma, L.; Oenema, O.; Wagner, F.; et al. Cost-Effective Management of Coastal Eutrophication: A Case Study for the Yangtze River Basin. Resour. Conserv. Recycl. 2020, 154, 104635. [Google Scholar] [CrossRef]
- Boyer, J.N.; Kelble, C.R.; Ortner, P.B.; Rudnick, D.T. Phytoplankton Bloom Status: Chlorophyll a Biomass as an Indicator of Water Quality Condition in the Southern Estuaries of Florida, USA. Ecol. Indic. 2009, 9 (Suppl. S6), S56–S67. [Google Scholar] [CrossRef]
- Hu, W.; Li, C.; Ye, C.; Wang, J.; Wei, W.; Deng, Y. Research Progress on Ecological Models in the Field of Water Eutrophication: CiteSpace Analysis Based on Data from the ISI Web of Science Database. Ecol. Model. 2019, 410, 108779. [Google Scholar] [CrossRef]
- Burigato Costa, C.M.d.s.; da Silva Marques, L.; Almeida, A.K.; Leite, I.R.; de Almeida, I.K. Applicability of Water Quality Models around the World—A Review. Environ. Sci. Pollut. Res. 2019, 26, 36141–36162. [Google Scholar] [CrossRef]
- Costa, C.M.d.S.B.; Leite, I.R.; Almeida, A.K.; de Almeida, I.K. Choosing an Appropriate Water Quality Model—a Review. Environ. Monit. Assess. 2021, 193, 38. [Google Scholar] [CrossRef]
- Park, Y.; Cho, K.H.; Park, J.; Cha, S.M.; Kim, J.H. Development of Early-Warning Protocol for Predicting Chlorophyll-a Concentration Using Machine Learning Models in Freshwater and Estuarine Reservoirs, Korea. Sci. Total Environ. 2015, 502, 31–41. [Google Scholar] [CrossRef]
- Cole, T.M.; Wells, S.A. CE-QUAL-W2: A Two-Dimensional, Laterally Averaged, Hy-Drodynamic and Water Quality Model, Version 3.1; Instruction Report EL-03-1; US Army Engi-neering and Research Development Center: Vicksburg, MS, USA, 2003. [Google Scholar]
- Walstra, D.J.R. Description of TRANSPOR2004 and Implementation in Delft3D-ONLINE: Interim Report; Rijkswaterstaat Institute for Coast and Sea: Rotterdam, The Netherlands, 2004. [Google Scholar]
- Park, R.A.; Clough, J.S.; Wellman, M.C. AQUATOX: Modeling Environmental Fate and Ecological Effects in Aquatic Ecosystems. Ecol. Model. 2008, 213, 1–15. [Google Scholar] [CrossRef]
- Parveen, N.; Singh, S. Application of Qual2e Model for River Water Quality Modelling. Int. J. Adv. Res. Innov. 2016, 4, 429–432. [Google Scholar]
- Schwarz, G.; Hoos, A.B.; Alexander, R.B.; Smith, R.A. Section 3. The SPARROW Surface Water-Quality Model—Theory, Application and User Documentation; Techniques and Methods; USGS Numbered Series 6-B3; U.S. Geological Survey: Reston, VA, USA, 2006; Volume 6-B3. [CrossRef] [Green Version]
- Srinivas, R.; Singh, A.P. An Integrated Fuzzy-Based Advanced Eutrophication Simulation Model to Develop the Best Management Scenarios for a River Basin. Environ. Sci. Pollut. Res. 2018, 25, 9012–9039. [Google Scholar] [CrossRef] [PubMed]
- Duda, P.; Hummel, P.; Donigian, J.A.; Imhoff, J. BASINS/HSPF: Model Use, Calibration, and Validation. Trans. ASABE 2012, 55, 1523–1547. [Google Scholar] [CrossRef]
- Arnold, J.; Moriasi, D.; Gassman, P.; Abbaspour, K.; White, M.; Srinivasan, R.; Santhi, C.; Harmel, R.; van Griensven, A.; Van Liew, M.; et al. SWAT: Model Use, Calibration, and Validation. Trans. ASABE 2012, 55, 1491–1508. [Google Scholar] [CrossRef]
- Morelli, B.; Hawkins, T.R.; Niblick, B.; Henderson, A.D.; Golden, H.E.; Compton, J.E.; Cooter, E.J.; Bare, J.C. Critical Review of Eutrophication Models for Life Cycle Assessment. Environ. Sci. Technol. 2018, 52, 9562–9578. [Google Scholar] [CrossRef]
- Debele, B.; Srinivasan, R.; Parlange, J.-Y. Coupling Upland Watershed and Downstream Waterbody Hydrodynamic and Water Quality Models (SWAT and CE-QUAL-W2) for Better Water Resources Management in Complex River Basins. Environ. Model. Assess. 2008, 13, 135–153. [Google Scholar] [CrossRef]
- Grizzetti, B.; Bouraoui, F.; Granlund, K.; Rekolainen, S.; Bidoglio, G. Modelling Diffuse Emission and Retention of Nutrients in the Vantaanjoki Watershed (Finland) Using the SWAT Model. Ecol. Model. 2003, 169, 25–38. [Google Scholar] [CrossRef]
- Santhi, C.; Arnold, J.G.; Williams, J.R.; Dugas, W.A.; Srinivasan, R.; Hauck, L.M. Validation of the Swat Model on a Large Rwer Basin with Point and Nonpoint Sources1. JAWRA J. Am. Water Resour. Assoc. 2001, 37, 1169–1188. [Google Scholar] [CrossRef]
- Krysanova, V.; Arnold, J.G. Advances in Ecohydrological Modelling with SWAT—A Review. Hydrol. Sci. J. 2008, 53, 939–947. [Google Scholar] [CrossRef]
- Imani, S.; Delavar, M.; Niksokhan, M.H. Identification of Nutrients Critical Source Areas with SWAT Model under Limited Data Condition. Water Resour. 2019, 46, 128–137. [Google Scholar] [CrossRef]
- Lee, J.; Woo, S.-Y.; Kim, Y.-W.; Kim, S.-J.; Pyo, J.; Cho, K.H. Dynamic Calibration of Phytoplankton Blooms Using the Modified SWAT Model. J. Clean. Prod. 2022, 343, 131005. [Google Scholar] [CrossRef]
- Orlińska-Woźniak, P.; Szalińska, E.; Jakusik, E.; Bojanowski, D.; Wilk, P. Biomass Production Potential in a River under Climate Change Scenarios. Environ. Sci. Technol. 2021, 55, 11113–11124. [Google Scholar] [CrossRef]
- Gąbka, M.; Jakubas, E.; Joniak, T.; Golski, J. Rzeki Wełna i Flinta—Charakterystyka Obiektów Badań, Ich Położenie i Granice Zlewni; Bogucki Wydawnictwo Naukowe: Poznań, Poland, 2014; pp. 21–33. [Google Scholar]
- Budzynska, A.; Dondajewska-Pielka, R.; Rosinska, J.; Kowalczewska-Madura, K.; Kozak, A.; Bogucki Wydawnictwo Naukowe. Ekosystemy Wodne: Funkcjonowanie, Znaczenie, Ochrona i Rekultywacja: Monografia Wydana z Okazji Jubileuszu 70. Urodzin prof. dr. hab. Ryszarda Goldyna; Bogucki Wydawnictwo Naukowe: Poznan, Poland, 2019. [Google Scholar]
- Dondajewska, R.; Gołdyn, R.; Kowalczewska-Madura, K.; Kozak, A.; Romanowicz-Brzozowska, W.; Rosińska, J.; Budzyńska, A.; Podsiadłowski, S. Hypertrophic Lakes and the Results of Their Restoration in Western Poland. In Polish River Basins and Lakes—Part II: Biological Status and Water Management; Korzeniewska, E., Harnisz, M., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 373–399. [Google Scholar] [CrossRef]
- GIOŚ. Monitoring Wód. Available online: https://www.gios.gov.pl/pl/stan-srodowiska/monitoring-wod (accessed on 5 April 2022).
- Messyasz, B.; Szczuka, E.; Kaznowski, A.; Burchardt, L. Phytoseston and Heterotrophic Bacteria in the Assessment of the Waters in the Wełna and Nielba Rivers. Oceanol. Hydrobiol. Stud. 2010, 39, 45–63. [Google Scholar] [CrossRef]
- O Wodach Polskich. Available online: https://www.wody.gov.pl/ (accessed on 5 April 2022).
- Strona główna|Instytut Meteorologii i Gospodarki Wodnej—Państwowy Instytut Badawczy. Available online: https://www.imgw.pl/ (accessed on 5 April 2022).
- Abbaspour, K.C.; Rouholahnejad, E.; Vaghefi, S.; Srinivasan, R.; Yang, H.; Kløve, B. A Continental-Scale Hydrology and Water Quality Model for Europe: Calibration and Uncertainty of a High-Resolution Large-Scale SWAT Model. J. Hydrol. 2015, 524, 733–752. [Google Scholar] [CrossRef] [Green Version]
- Park, G.A.; Park, J.Y.; Joh, H.K.; Lee, J.W.; Ahn, S.R.; Kim, S.J. Evaluation of Mixed Forest Evapotranspiration and Soil Moisture Using Measured and Swat Simulated Results in a Hillslope Watershed. KSCE J. Civ. Eng. 2014, 18, 315–322. [Google Scholar] [CrossRef]
- Khoi, D.N.; Thom, V.T. Parameter Uncertainty Analysis for Simulating Streamflow in a River Catchment of Vietnam. Glob. Ecol. Conserv. 2015, 4, 538–548. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Kronvang, B.; Audet, J.; Trolle, D.; Andersen, H.E.; Thodsen, H.; van Griensven, A. Modelling Sediment and Total Phosphorus Export from a Lowland Catchment: Comparing Sediment Routing Methods. Hydrol. Process. 2015, 29, 280–294. [Google Scholar] [CrossRef]
- Ostojski, M.S.; Gębala, J.; Orlińska-Woźniak, P.; Wilk, P. Implementation of Robust Statistics in the Calibration, Verification and Validation Step of Model Evaluation to Better Reflect Processes Concerning Total Phosphorus Load Occurring in the Catchment. Ecol. Model. 2016, 332, 83–93. [Google Scholar] [CrossRef]
- Bauwe, A.; Eckhardt, K.-U.; Lennartz, B. Predicting Dissolved Reactive Phosphorus in Tile-Drained Catchments Using a Modified SWAT Model. Ecohydrol. Hydrobiol. 2019, 19, 198–209. [Google Scholar] [CrossRef]
- Neitsch, S.; Arnold, J.; Kiniry, J.; Williams, J.R. Soil and Water Asessment Tool Theoritical Documentation: Version 2009; Texas Water Resources Institute Technical Report No. 406; Texas Water Resources Institute, Texas A&M University: College Station, TX, USA, 2011. [Google Scholar]
- Monod, J. Recherches sur la Croissance des Cultures Bactériennes; Hermann: Paris, France, 1942. [Google Scholar]
- Kiniry, J.R.; Tischler, C.R.; Van Esbroeck, G.A. Radiation Use Efficiency and Leaf CO2 Exchange for Diverse C4 Grasses. Biomass Bioenergy 1999, 17, 95–112. [Google Scholar] [CrossRef]
- Eppley, R.W.; Thomas, W.H. Comparison of Half-Saturation Constants for Growth and Nitrate Uptake of Marine Phytoplankton 2. J. Phycol. 1969, 5, 375–379. [Google Scholar] [CrossRef]
- Benedict, A.H.; Carlson, D.A. Rational Assessment of the Streeter-Phelps Temperature Coefficient. J. Water Pollut. Control Fed. 1974, 46, 1792–1799. [Google Scholar]
- Dragoni, D.; Schmid, H.P.; Wayson, C.A.; Potter, H.; Grimmond, C.S.B.; Randolph, J.C. Evidence of Increased Net Ecosystem Productivity Associated with a Longer Vegetated Season in a Deciduous Forest in South-Central Indiana, USA. Glob. Chang. Biol. 2011, 17, 886–897. [Google Scholar] [CrossRef]
- Tylkowski, J. The Variability of Climatic Vegetative Seasons and Thermal Resources at the Polish Baltic Sea Coastline in the Context of Potential Composition of Coastal Forest Communities. Balt. For. 2015, 21, 73–82. [Google Scholar]
- Wüest, A.; Piepke, G.; Van Senden, D.C. Turbulent Kinetic Energy Balance as a Tool for Estimating Vertical Diffusivity in Wind-Forced Stratified Waters. Limnol. Oceanogr. 2000, 45, 1388–1400. [Google Scholar] [CrossRef]
- Czikowsky, M.J.; MacIntyre, S.; Tedford, E.W.; Vidal, J.; Miller, S.D. Effects of Wind and Buoyancy on Carbon Dioxide Distribution and Air-Water Flux of a Stratified Temperate Lake. J. Geophys. Res. Biogeosci. 2018, 123, 2305–2322. [Google Scholar] [CrossRef]
- Boogaard, H.L.; Kroes, J.G. Leaching of Nitrogen and Phosphorus from Rural Areas to Surface Waters in the Netherlands. In Soil and Water Quality at Different Scales: Proceedings of the Workshop “Soil and Water Quality at Different Scales” held 7–9 August 1996, Wageningen, The Netherlands; Finke, P.A., Bouma, J., Hoosbeek, M.R., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 1998; pp. 321–324. [Google Scholar] [CrossRef]
- Pikosz, M.; Messyasz, B.; Gąbka, M. Functional Structure of Algal Mat (Cladophora Glomerata) in a Freshwater in Western Poland. Ecol. Indic. 2017, 74, 1–9. [Google Scholar] [CrossRef]
- Patil, S.D.; Stieglitz, M. Comparing spatial and temporal transferability of hydrological model parameters. J. Hydrol. 2015, 525, 409–417. [Google Scholar] [CrossRef] [Green Version]
- Libera, D.A.; Sankarasubramanian, A. Multivariate bias corrections of mechanistic water quality model predictions. J. Hydrol. 2018, 564, 529–541. [Google Scholar] [CrossRef]
- Moriasi, D.N.; Gitau, M.W.; Pai, N.; Daggupati, P. Hydrologic and water quality models: Performance measures and evaluation criteria. Trans. ASABE 2015, 58, 1763–1785. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orlińska-Woźniak, P. Modeling Chlorophyll a with Use of the SWAT Tool for the Nielba River (West-Central Poland) as an Example of an Unmonitored Watercourse. Water 2022, 14, 1528. https://doi.org/10.3390/w14101528
Orlińska-Woźniak P. Modeling Chlorophyll a with Use of the SWAT Tool for the Nielba River (West-Central Poland) as an Example of an Unmonitored Watercourse. Water. 2022; 14(10):1528. https://doi.org/10.3390/w14101528
Chicago/Turabian StyleOrlińska-Woźniak, Paulina. 2022. "Modeling Chlorophyll a with Use of the SWAT Tool for the Nielba River (West-Central Poland) as an Example of an Unmonitored Watercourse" Water 14, no. 10: 1528. https://doi.org/10.3390/w14101528
APA StyleOrlińska-Woźniak, P. (2022). Modeling Chlorophyll a with Use of the SWAT Tool for the Nielba River (West-Central Poland) as an Example of an Unmonitored Watercourse. Water, 14(10), 1528. https://doi.org/10.3390/w14101528