

Rainfall Partitioning by Evergreen and Deciduous Broad-Leaved Xerophytic Tree Species: Influence of Rainfall, Canopy Characteristics, and Meteorology





Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Meteorological and Canopy Characteristics
2.3. Measurement of Gross Rainfall, Throughfall and Stemflow
2.4. Statistical Analysis
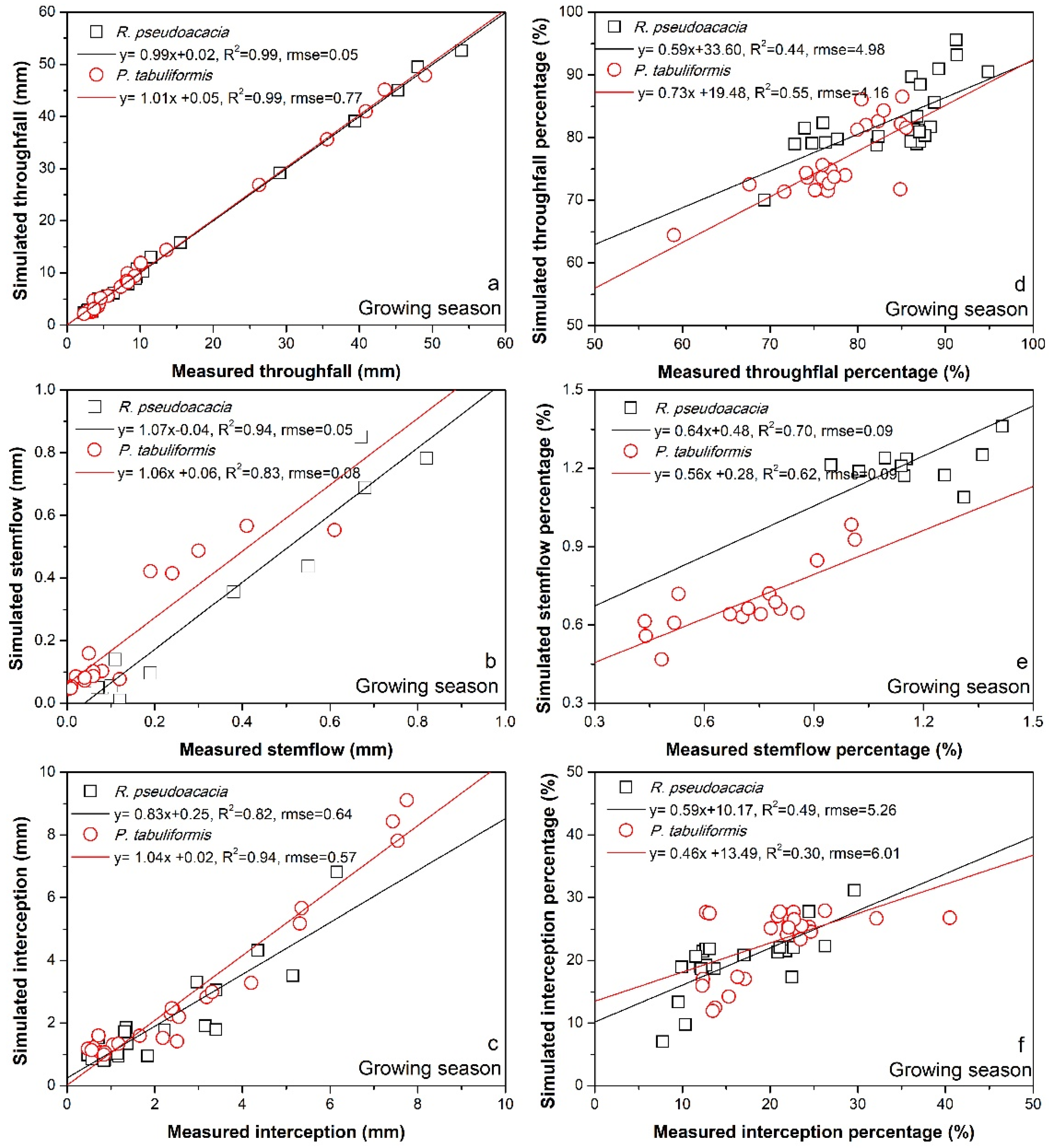
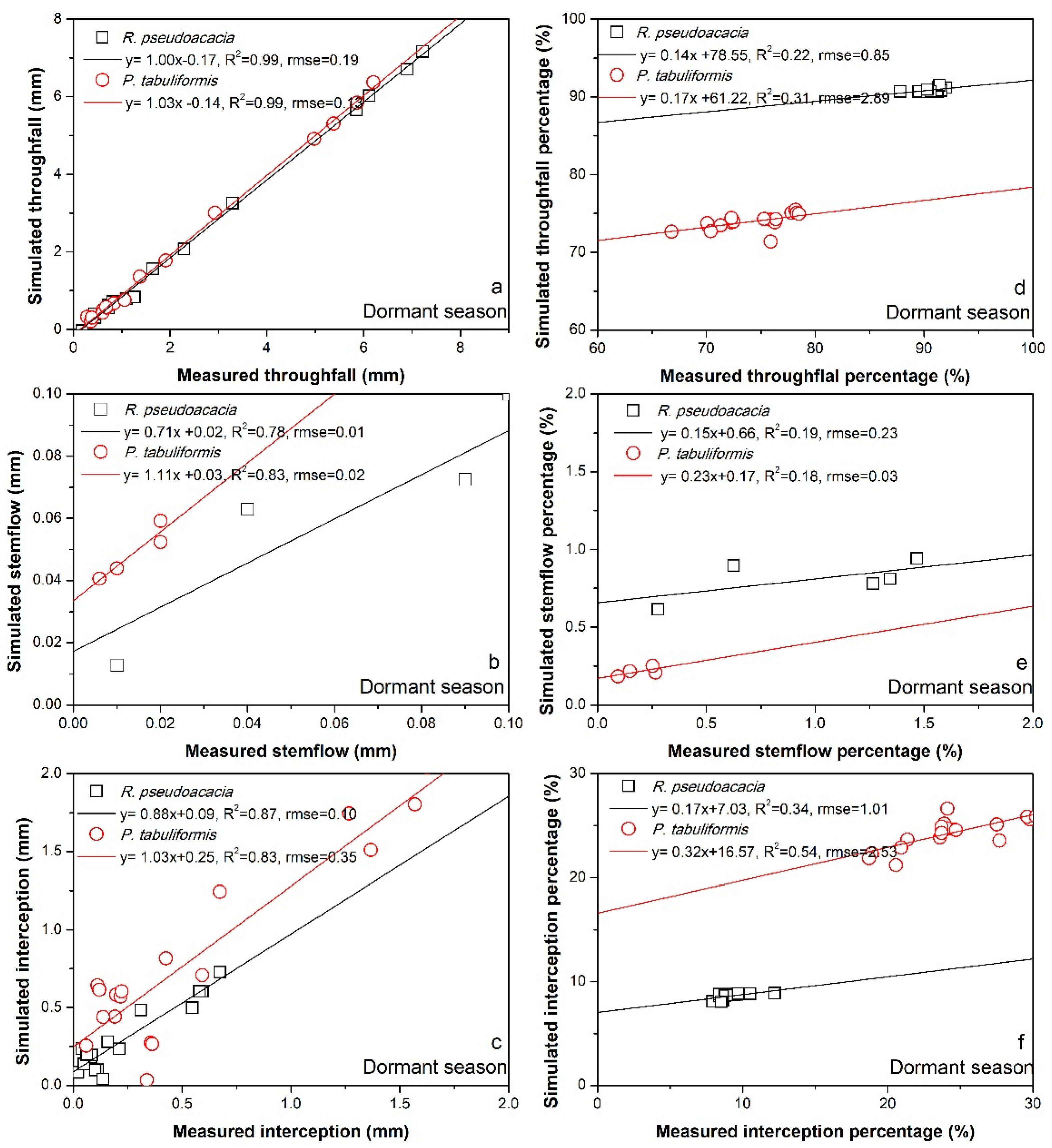
3. Results
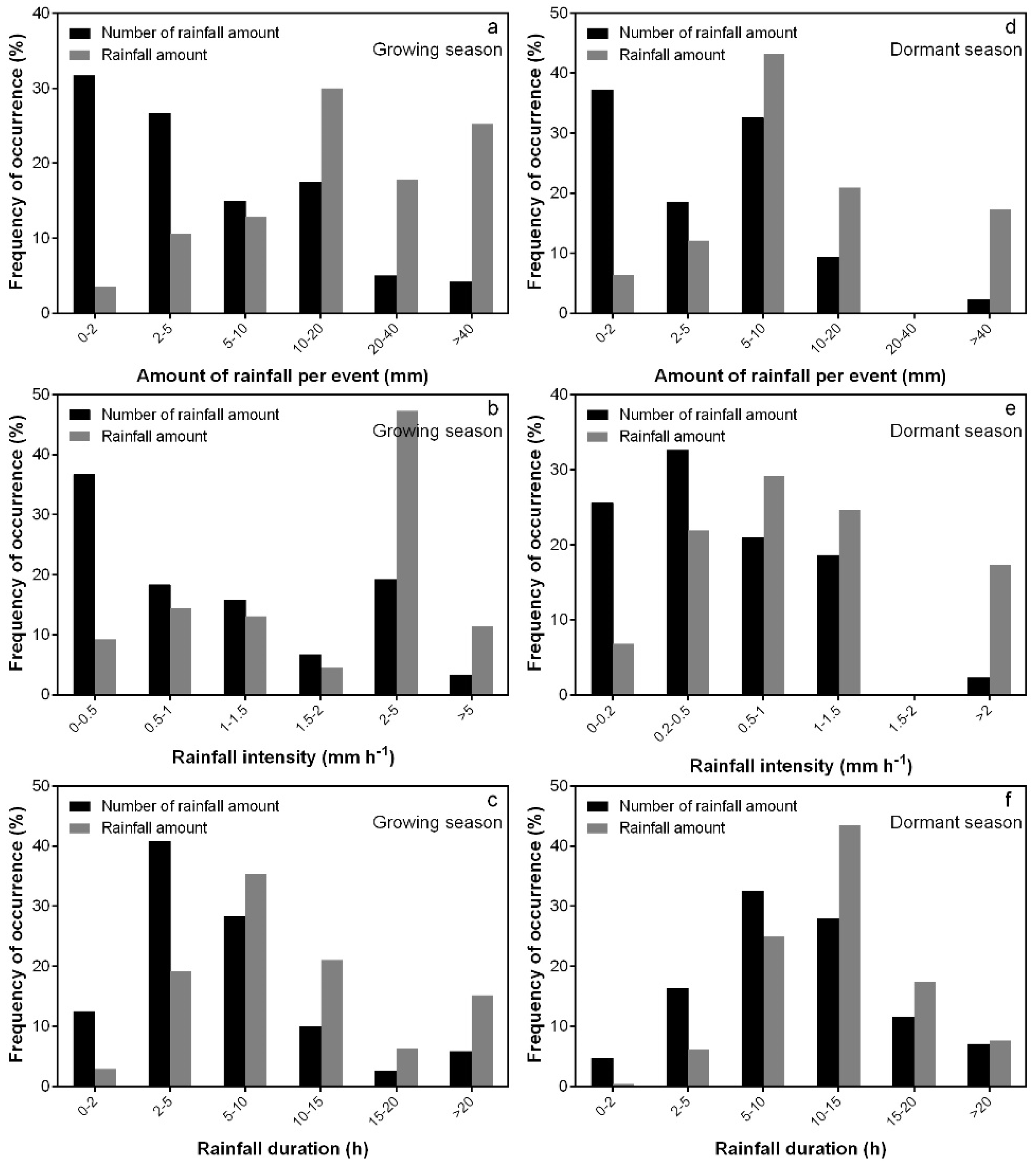
3.1. Rainfall Characteristics
3.2. Rainfall Partitioning Characteristics
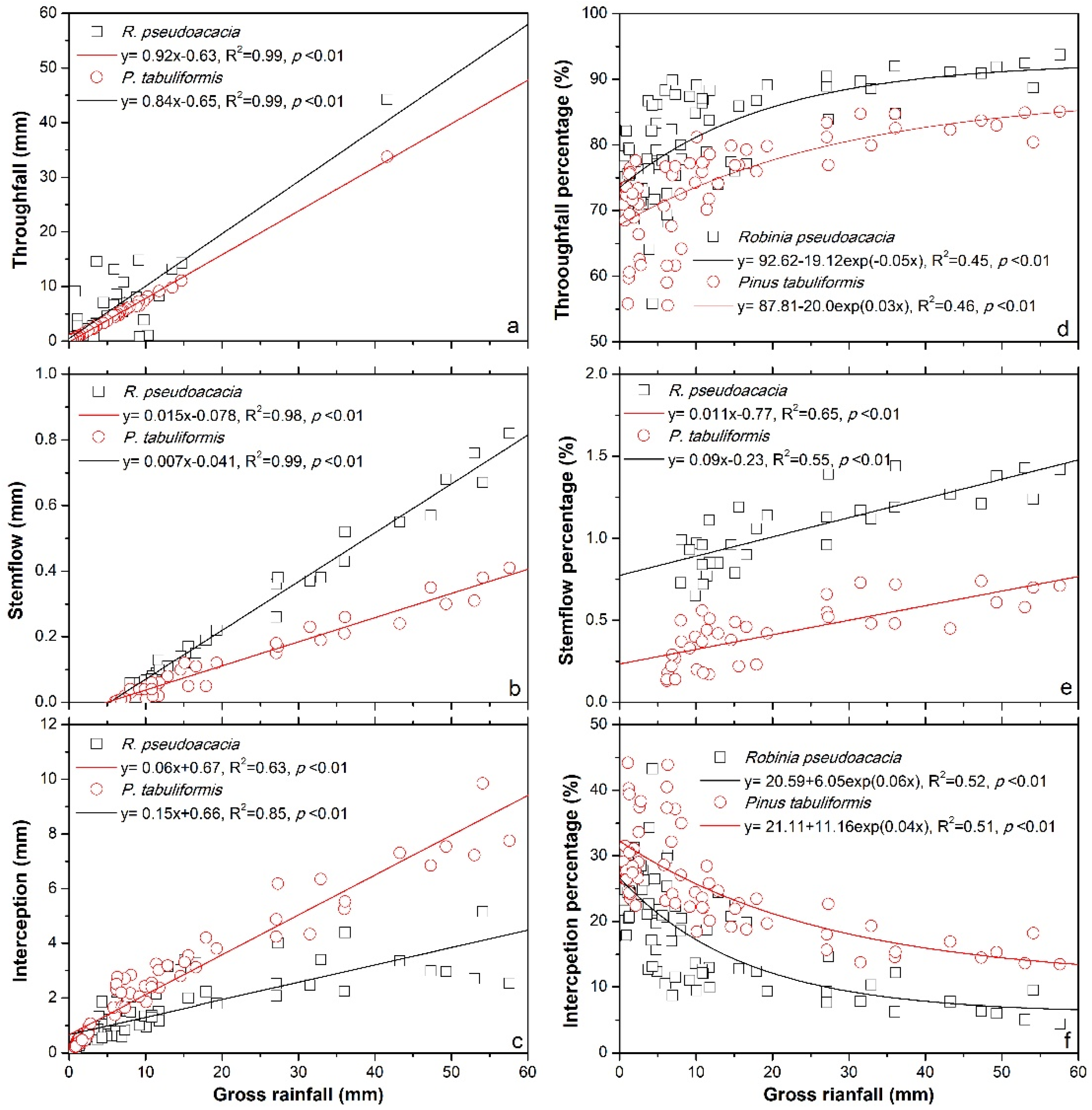
3.3. Rainfall Partitioning in Relation to Rainfall Amount
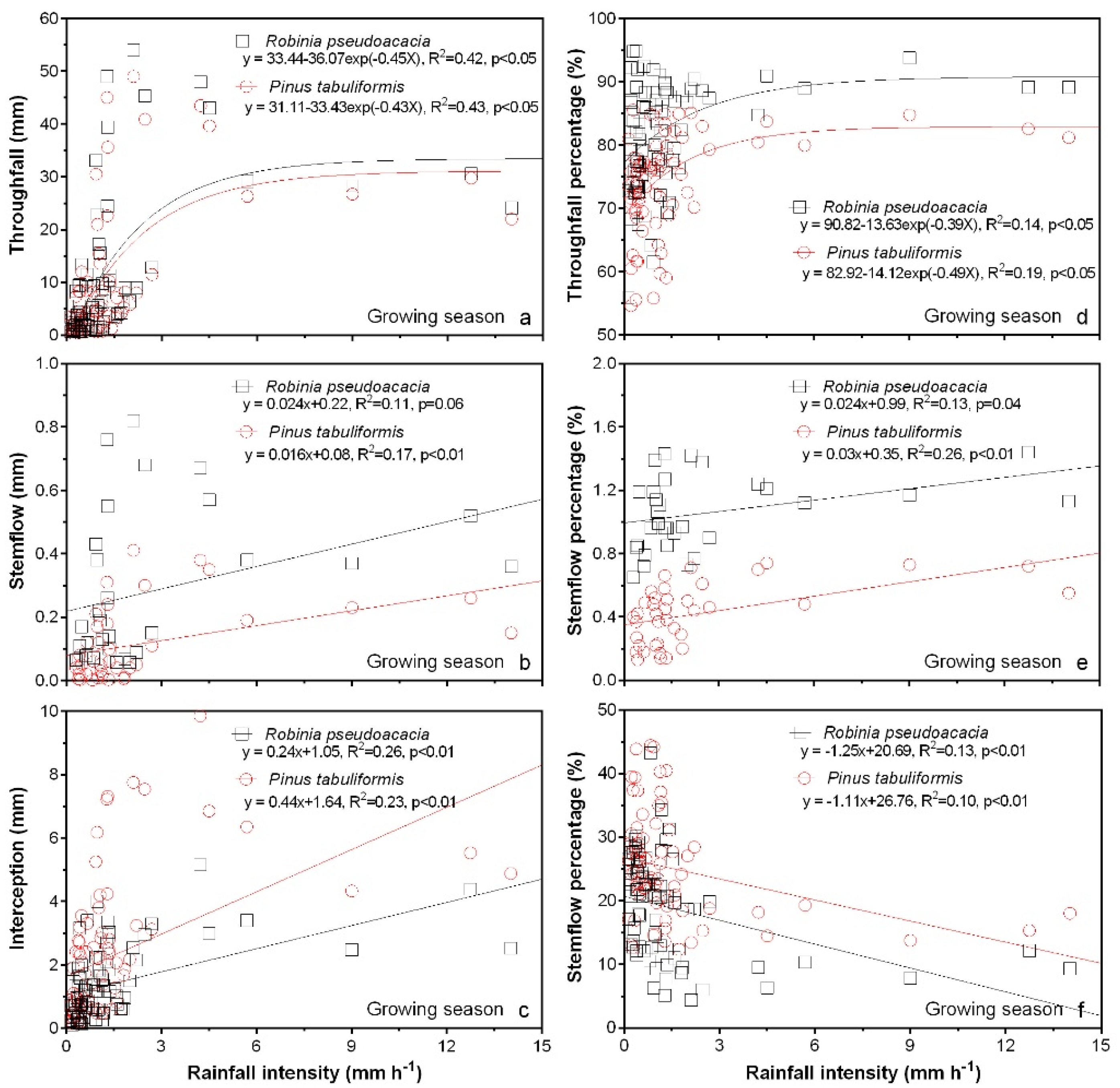
3.4. Rainfall Partitioning in Relation to Rainfall Intensity
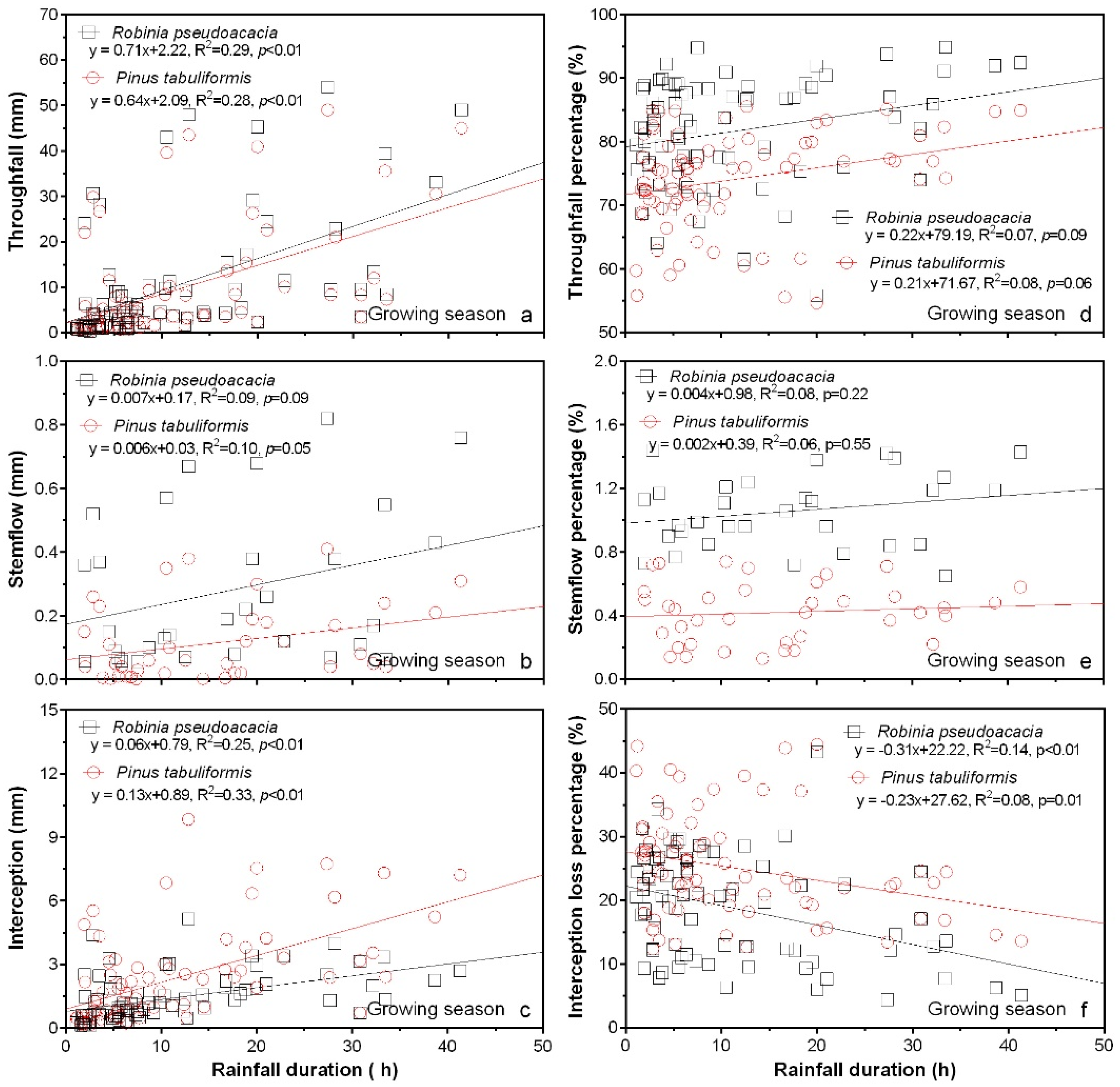
3.5. Rainfall Partitioning in Relation to Rainfall Duration
3.6. Relationship between Canopy Characteristics, Meteorological Variables and Rainfall Partitioning
3.7. Multiple Linear Regression Analysis
4. Discussion
4.1. Difference in Rainfall Partitioning between Two Study Forest Species
4.2. Rainfall Partitioning in Relation to Rainfall, Canopy Characteristics and Meteorological Variables
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Cao, S.; Chen, L.; Shankman, D.; Wang, C.; Wang, X.; Zhang, H. Excessive reliance on afforestation in China’s arid and semi-arid regions: Lessons in ecological restoration. Earth-Sci. Rev. 2011, 104, 240–245. [Google Scholar] [CrossRef]
- Malagnoux, M.; Sène, E.H.; Atzmon, N. Forests, trees and water in arid lands: A delicate balance. Unasylva 2007, 58, 24–29. [Google Scholar]
- Bastin, J.; Berrahmouni, N.; Grainger, A.; Maniatis, D.; Mollicone, D.; Moore, R.; Patriarca, C.; Picard, N.; Sparrow, B.; Abraham, E.M.; et al. The extent of forest in dryland biomes. Science 2017, 356, 635–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sexton, J.O.; Song, X.P.; Feng, M.; Noojipady, P.; Anand, A.; Huang, C.; Kim, D.H.; Collins, K.M.; Channan, S.; Dimiceli, C. Global, 30-m resolution continuous fields of tree cover: Landsat-based rescaling of MODIS vegetation continuous fields with lidar-based estimates of error. Int. J. Digit. Earth 2013, 6, 427–448. [Google Scholar] [CrossRef] [Green Version]
- Sadeghi, S.M.M.; Attarod, P.; Van Stan, J.T.; Pypker, T.G. The importance of considering rainfall partitioning in afforestation initiatives in semiarid climates: A comparison of common planted tree species in Tehran, Iran. Sci. Total Environ. 2016, 568, 845–855. [Google Scholar] [CrossRef]
- Fathizadeh, O.; Hosseini, S.M.; Keim, R.F.; Boloorani, A.D. A seasonal evaluation of the reformulated Gash interception model for semi-arid deciduous oak forest stands. For. Ecol. Manag. 2018, 409, 601–613. [Google Scholar] [CrossRef]
- Ma, C.; Luo, Y.; Shao, M.; Li, X.; Sun, L.; Jia, X. Environmental controls on sap flow in black locust forest in Loess Plateau, China. Sci. Rep. 2017, 7, 13160. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, A.D.; Giambelluca, T.W.; Nullet, M.A.; Sutherland, R.A.; Tantasarin, C.; Vogler, J.B.; Negishi, J.N. Throughfall in an evergreen-dominated forest stand in northern Thailand: Comparison of mobile and stationary methods. Agric. For. Meteorol. 2009, 149, 373–384. [Google Scholar] [CrossRef]
- Llorens, P.; Domingo, F. Rainfall partitioning by vegetation under Mediterranean conditions. A review of studies in Europe. J. Hydrol. 2007, 335, 37–54. [Google Scholar] [CrossRef]
- Zhang, Y.F.; Wang, X.P.; Hu, R.; Pan, Y.X.; Paradeloc, M. Rainfall partitioning into throughfall, stemflow and interception loss by two xerophytic shrubs within a rain-fed re-vegetated desert ecosystem, northwestern China. J. Hydrol. 2015, 527, 1084–1095. [Google Scholar] [CrossRef]
- Klaassen, W.; Bosveld, F.; de Water, E. Water storage and evaporation as constituents of rainfall interception. J. Hydrol. 1998, 212, 36–50. [Google Scholar] [CrossRef]
- Crockford, R.H.; Richardson, D.P. Partitioning of rainfall into throughfall, stemflow and interception: Effect of forest type, ground cover and climate. Hydrol. Process. 2000, 14, 2903–2920. [Google Scholar] [CrossRef]
- Muzylo, A.; Valente, F.; Domingo, F.; Llorens, P. Modelling rainfall partitioning with sparse Gash and Rutter models in a downy oak stand in leafed and leafless periods. Hydrol. Process. 2012, 26, 3161–3173. [Google Scholar] [CrossRef]
- Toba, T.; Ohta, T. An observational study of the factors that influence interception loss in boreal and temperate forests. J. Hydrol. 2005, 313, 208–220. [Google Scholar] [CrossRef]
- Levia, D.F.; Germer, S. A review of stemflow generation dynamics and stemflow-environment interactions in forests and shrublands. Rev. Geophys. 2015, 53, 673–714. [Google Scholar] [CrossRef]
- Carlyle-Moses, D.E. Throughfall, sternflow, and canopy interception loss fluxes in a semi-arid Sierra Madre Oriental matorral community. J. Arid. Environ. 2004, 58, 181–202. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, Q.; Shao, M.; Wang, Q. Rainfall interception in a robinia pseudoacacia forest stand: Estimates using gash’s analytical model. J. Hydrol. Eng. 2013, 18, 474–479. [Google Scholar] [CrossRef]
- Ghimire, C.P.; Bruijnzeel, L.A.; Lubczynski, M.W.; Bonell, M. Rainfall interception by natural and planted forests in the Middle Mountains of Central Nepal. J. Hydrol. 2012, 475, 270–280. [Google Scholar] [CrossRef]
- Keresztesi, B. The black locust. Unasylva 1980, 32, 23–33. [Google Scholar]
- Vitkova, M.; Muellerova, J.; Sadlo, J.; Pergl, J.; Pysek, P. Black locust (Robinia pseudoacacia) beloved and despised: A story of an invasive tree in Central Europe. For. Ecol. Manag. 2017, 384, 287–302. [Google Scholar] [CrossRef]
- Tsiko, C.T.; Makurira, H.; Gerrits, A.M.J.; Savenije, H.H.G. Measuring forest floor and canopy interception in a savannah ecosystem. Phys. Chem. Earth 2012, 47–48, 122–127. [Google Scholar] [CrossRef]
- Domingo, F.; Sanchez, G.; Moro, M.J.; Brenner, A.J.; Puigdefabregas, J. Measurement and modelling of rainfall interception by three semi-arid canopies. Agric. For. Meteorol. 1998, 91, 275–292. [Google Scholar] [CrossRef]
- Ma, C.; Li, X.; Luo, Y.; Shao, M.; Jia, X. The modelling of rainfall interception in growing and dormant seasons for a pine plantation and a black locust plantation in semi-arid Northwest China. J. Hydrol. 2019, 577, 123849. [Google Scholar] [CrossRef]
- LI-COR. LAI-2200C Plant Canopy Analyzer: Instruction Manual; LI-COR Inc.: Lincoln, Nebraska, 2015. [Google Scholar]
- Gash, J.H.C.; Lloyd, C.R.; Lachaud, G. Estimating sparse forest rainfall interception with an analytical model. J. Hydrol. 1995, 170, 79–86. [Google Scholar] [CrossRef]
- Bryant, M.L.; Bhat, S.; Jacobs, J.M. Measurements and modeling of throughfall variability for five forest communities in the southeastern US. J. Hydrol. 2005, 312, 95–108. [Google Scholar] [CrossRef]
- Silva, I.C.; Okumura, T. Throughfall, stemflow and interception loss in a mixed white oak forest (Quercus serrata Thunb.). J. For. Res. 1996, 1, 123–129. [Google Scholar] [CrossRef]
- Shi, Z.; Wang, Y.; Xu, L.; Xiong, W.; Yu, P.; Gao, J.; Zhang, L. Fraction of incident rainfall within the canopy of a pure stand of Pinus armandii with revised Gash model in the Liupan Mountains of China. J. Hydrol. 2010, 385, 44–50. [Google Scholar]
- Park, A.; Cameron, J.L. The influence of canopy traits on throughfall and stemflow in five tropical trees growing in a Panamanian plantation. For. Ecol. Manag. 2008, 255, 1915–1925. [Google Scholar] [CrossRef]
- Deguchi, A.; Hattori, S.; Park, H.T. The influence of seasonal changes in canopy structure on interception loss: Application of the revised Gash model. J. Hydrol. 2006, 318, 80–102. [Google Scholar] [CrossRef]
- Molina, A.J.; del Campo, A.D. The effects of experimental thinning on throughfall and stemflow: A contribution towards hydrology-oriented silviculture in Aleppo pine plantations. For. Ecol. Manag. 2012, 269, 206–213. [Google Scholar] [CrossRef] [Green Version]
- Fan, J.; Oestergaard, K.T.; Guyot, A.; Lockington, D.A. Measuring and modeling rainfall interception losses by a native Banksia woodland and an exotic pine plantation in subtropical coastal Australia. J. Hydrol. 2014, 515, 156–165. [Google Scholar] [CrossRef]
- Herbst, M.; Rosier, P.T.W.; McNeil, D.D.; Harding, R.J.; Gowing, D.J. Seasonal variability of interception evaporation from the canopy of a mixed deciduous forest. Agric. For. Meteorol. 2008, 148, 1655–1667. [Google Scholar] [CrossRef]
- Siles, P.; Vaast, P.; Dreyer, E.; Harmand, J.-M. Rainfall partitioning into throughfall, stemflow and interception loss in a coffee (Coffea arabica L.) monoculture compared to an agroforestry system with Inga densiflora. J. Hydrol. 2010, 395, 39–48. [Google Scholar] [CrossRef]
- Fang, S.; Zhao, C.; JIian, S.; Kai, Y. Canopy interception of pinus tabulaeformis plantation on Longzhong Loess Plateau, Northwest China: Characteristics and simulation. Chin. J. Appl. Ecol. 2013, 24, 1509–1516. [Google Scholar]
- Gomez, J.A.; Giraldez, J.V.; Fereres, E. Rainfall interception by olive trees in relation to leaf area. Agric. Water Manag. 2001, 49, 65–76. [Google Scholar] [CrossRef]
- Yuan, C.; Gao, G.; Fu, B. Stemflow of a xerophytic shrub (Salix psammophila) in northern China: Implication for beneficial branch architecture to produce stemflow. J. Hydrol. 2016, 539, 577–588. [Google Scholar] [CrossRef]
- Su, L.; Xu, W.; Zhao, C.; Xie, Z.; Ju, H. Inter- and intra-specific variation in stemflow for evergreen species and deciduous tree species in a subtropical forest. J. Hydrol. 2016, 537, 1–9. [Google Scholar] [CrossRef]
- Owens, M.K.; Lyons, R.K.; Alejandro, C.L. Rainfall partitioning within semiarid juniper communities: Effects of event size and canopy cover. Hydrol. Process. 2006, 20, 3179–3189. [Google Scholar] [CrossRef]
- Calder, I.R. Dependence of rainfall interception on drop size.1. Development of the two-layer stochastic model. J. Hydrol. 1996, 185, 363–378. [Google Scholar] [CrossRef]
- Attarod, P.; Sadeghi, S.M.M.; Pypker, T.G.; Bagheri, H.; Bagheri, M.; Bayramzadeh, V. Needle-leaved trees impacts on rainfall interception and canopy storage capacity in an arid environment. New For. 2015, 46, 339–355. [Google Scholar] [CrossRef]
- Staelens, J.; De Schrijver, A.; Verheyen, K.; Verhoest, N.E.C. Rainfall partitioning into throughfall, stemflow, and interception within a single beech (Fagus sylvatica L.) canopy: Influence of foliation, rain event characteristics, and meteorology. Hydrol. Process. 2008, 22, 33–45. [Google Scholar] [CrossRef]
- Zheng, J.; Fan, J.; Zhang, F.; Yan, S.; Xiang, Y. Rainfall partitioning into throughfall, stemflow and interception loss by maize canopy on the semi-arid Loess Plateau of China. Agric. Water Manag. 2018, 195, 25–36. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tree Species | Study Period | Pg (mm) | TF (mm) | SF (mm) | I (mm) |
|---|---|---|---|---|---|
| R. pseudoacacia | Growing season | 1002.1 ± 1.6 | 801.7 ± 1.5 (80.0%) | 14.0 ± 0.03 (1.4%) a | 186.4 ± 0.1 (18.6%) |
| Dormant season | 240.1 ± 1.0 | 214.4 ± 0.9 (89.3%) | 3.8 ± 0.02 (1.6%) a | 21.8 ± 0.1 (9.1%) | |
| Annual | 1242.2 ± 1.1 | 1016.1 ± 1.0 (81.8%) | 17.8 ± 0.02 (1.4%) a | 208.2 ± 0.1 (16.8%) | |
| P. tabuliformis | Growing season | 1002.1 ± 1.6 | 752.6 ± 1.3 (75.1%) | 7.2 ± 0.01 (0.7%) b | 242.3 ± 0.2 (24.2%) |
| Dormant season | 240.1 ± 1.0 | 180.8 ± 0.8 (75.3%) | 1.8 ± 0.01 (0.7%) b | 57.6 ± 0.2 (24.0%) | |
| Annual | 1242.2 ± 1.1 | 933.4 ± 0.9 (75.1%) | 9.0 ± 0.01(0.7%) b | 299.9 ± 0.2 (24.1%) |
| Rainfall Partition Components | Growing Season (May–October) | Dormant Season (November–April) | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ta (°C) | RH (%) | WS (m s−1) | LAI | Ta (°C) | RH (%) | WS (m s−1) | PAI | ||||||||||
| r | p | r | p | r | p | r | p | r | p | r | p | r | p | r | p | ||
| TF (mm) | R. pseudoacacia | 0.1 | 0.46 | 0.39 | 0.09 | 0.05 | 0.69 | 0.61 | 0.04 | 0.19 | 0.22 | −0.2 | 0.21 | 0.09 | 0.56 | 0.32 | 0.23 |
| P. tabuliformis | 0.26 | 0.11 | 0.23 | 0.15 | 0.05 | 0.75 | 0.15 | 0.32 | 0.18 | 0.21 | −0.19 | 0.23 | 0.1 | 0.53 | 0.18 | 0.42 | |
| TF (%) | R. pseudoacacia | 0.27 | 0.14 | 0.55 | 0.12 | 0.09 | 0.52 | 0.58 | 0.03 | −0.03 | 0.81 | 0.02 | 0.91 | 0.04 | 0.81 | 0.04 | 0.33 |
| P. tabuliformis | −0.02 | 0.89 | 0.3 | 0.06 | 0.02 | 0.93 | 0.29 | 0.34 | −0.05 | 0.73 | −0.22 | 0.15 | 0.29 | 0.06 | 0.08 | 0.51 | |
| SF (mm) | R. pseudoacacia | 0.19 | 0.16 | 0.3 | 0.13 | 0 | 0.99 | 0.3 | 0.18 | 0.2 | 0.33 | 0.11 | 0.6 | −0.1 | 0.62 | 0.29 | 0.23 |
| P. tabuliformis | 0.33 | 0.11 | 0.09 | 0.55 | 0.03 | 0.86 | 0.26 | 0.42 | 0.33 | 0.17 | −0.04 | 0.88 | −0.02 | 0.93 | 0.21 | 0.62 | |
| SF (%) | R. pseudoacacia | 0.35 | 0.08 | 0.09 | 0.5 | −0.02 | 0.88 | 0.03 | 0.31 | 0.11 | 0.61 | 0.17 | 0.43 | −0.15 | 0.47 | 0.11 | 0.41 |
| P. tabuliformis | 0.45 | 0.07 | −0.16 | 0.33 | 0 | 0.99 | 0.19 | 0.42 | 0.35 | 0.14 | −0.28 | 0.25 | 0.13 | 0.59 | 0.17 | 0.52 | |
| I (mm) | R. pseudoacacia | 0.27 | 0.06 | 0.25 | 0.07 | 0.08 | 0.58 | 0.54 | 0.02 | 0.14 | 0.36 | −0.09 | 0.59 | 0.05 | 0.75 | 0.31 | 0.41 |
| P. tabuliformis | 0.36 | 0.22 | 0.18 | 0.26 | 0.12 | 0.47 | 0.33 | 0.18 | 0.13 | 0.42 | −0.1 | 0.51 | −0.05 | 0.98 | 0.29 | 0.14 | |
| I (%) | R. pseudoacacia | 0.24 | 0.07 | 0.55 | 0.08 | 0.08 | 0.54 | 0.55 | 0.04 | 0.12 | 0.46 | 0.31 | 0.06 | −0.07 | 0.68 | 0.18 | 0.35 |
| P. tabuliformis | −0.08 | 0.96 | −0.29 | 0.07 | −0.01 | 0.93 | 0.55 | 0.23 | −0.24 | 0.13 | 0.21 | 0.18 | −0.33 | 0.06 | 0.08 | 0.29 | |
| Tree Species | Variable | Fitted Equation | p | PPg | PRD | PRI | PLAI | R2 |
|---|---|---|---|---|---|---|---|---|
| R. pseudoacacia | TF (mm) | y = −0.38 + 0.94Pg − 0.03RD − 0.14RI − 0.26LAI | <0.001 | 0.000 | 0.02 | 0.00 | 0.01 | 0.99 |
| TF (%) | y = 78.08 + 0.31Pg − 0.02 RD + 0.10RI − 0.08LAI | <0.001 | 0.002 | 0.89 | 0.84 | 0.05 | 0.53 | |
| SF (mm) | y = −0.09 + 0.02Pg − 0.01RD − 0.003RI | <0.001 | 0.000 | 0.34 | 0.17 | - | 0.94 | |
| SF (%) | y = 0.75 + 0.01Pg + 0.002RD − 0.01RI | <0.001 | 0.000 | 0.61 | 0.30 | - | 0.67 | |
| I (mm) | y = 0.42 + 0.04Pg + 0.03RD + 0.13RI + 0.05LAI | <0.001 | 0.000 | 0.01 | 0.05 | 0.03 | 0.77 | |
| I (%) | y = 23.87 − 0.34Pg − 0.05RD − 0.34RI + 0.41LAI | <0.001 | 0.000 | 0.59 | 0.41 | 0.02 | 0.47 | |
| P. tabuliformis | TF (mm) | y = −0.41 + 0.85Pg − 0.02RD − 0.05RI | <0.001 | 0.000 | 0.04 | 0.26 | - | 0.99 |
| TF (%) | y = 70.25 + 0.26Pg + 0.02RD + 0.36RI | <0.001 | 0.004 | 0.82 | 0.42 | - | 0.55 | |
| SF (mm) | y = −0.03 + 0.01Pg − 0.01RD + 0.001RI | <0.001 | 0.000 | 0.16 | 0.84 | - | 0.96 | |
| SF (%) | y = 0.23 + 0.01Pg − 0.01RD +0.01RI | <0.001 | 0.000 | 0.67 | 0.13 | - | 0.59 | |
| I (mm) | y = 0.39 + 0.14Pg + 0.02RD +0.05RI | <0.001 | 0.000 | 0.02 | 0.21 | - | 0.93 | |
| I (%) | y = 29.17 − 0.27Pg − 0.03RD − 0.41RI | <0.001 | 0.004 | 0.77 | 0.37 | - | 0.29 |
| Tree Species | Variable | Fitted Equation | p | PPg | PRD | PRI | R2 |
|---|---|---|---|---|---|---|---|
| R. pseudoacacia | TF (mm) | y = −0.03 + 0.92Pg − 0.02RD + 0.02RI | <0.001 | 0.000 | 0.25 | 0.43 | 0.99 |
| TF (%) | y = 90.48 + 0.01Pg + 0.02 RD + 0.55RI | 0.015 | 0.912 | 0.59 | 0.28 | 0.23 | |
| SF (mm) | y = −0.07 + 0.02Pg + 0.01RD + 0.008RI | <0.001 | 0.000 | 0.38 | 0.67 | 0.97 | |
| SF (%) | y = 0.53 + 0.05Pg + 0.02RD − 0.17RI | 0.005 | 0.191 | 0.50 | 0.61 | 0.38 | |
| I (mm) | y = 0.16 + 0.06Pg − 0.01RD + 0.10RI | <0.001 | 0.000 | 0.18 | 0.47 | 0.81 | |
| I (%) | y = 8.77 − 0.11Pg + 0.01RD + 0.08RI | 0.003 | 0.151 | 0.93 | 0.92 | 0.25 | |
| P. tabuliformis | TF (mm) | y = −0.14 + 0.78Pg − 0.01RD + 0.27RI | <0.001 | 0.000 | 0.39 | 0.10 | 0.99 |
| TF (%) | y = 75.17 + 0.55Pg − 0.21RD − 2.32RI | 0.010 | 0.012 | 0.11 | 0.24 | 0.31 | |
| SF (mm) | y = −0.04 + 0.01Pg + 0.001RD + 0.01RI | <0.001 | 0.000 | 0.62 | 0.09 | 0.99 | |
| SF (%) | y = 0.11 + 0.01Pg − 0.002RD + 0.05RI | 0.010 | 0.294 | 0.79 | 0.59 | 0.58 | |
| I (mm) | y = 0.58 + 0.21Pg − 0.04RD + 0.01RI | <0.001 | 0.000 | 0.23 | 0.98 | 0.75 | |
| I (%) | y = 24.67 − 0.16Pg + 0.11RD − 1.88RI | <0.001 | 0.42 | 0.39 | 0.33 | 0.32 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, C.; Yao, Q.; Meng, H.; Zhou, B.; Wang, Q.; Luo, Y. Rainfall Partitioning by Evergreen and Deciduous Broad-Leaved Xerophytic Tree Species: Influence of Rainfall, Canopy Characteristics, and Meteorology. Water 2022, 14, 3723. https://doi.org/10.3390/w14223723
Ma C, Yao Q, Meng H, Zhou B, Wang Q, Luo Y. Rainfall Partitioning by Evergreen and Deciduous Broad-Leaved Xerophytic Tree Species: Influence of Rainfall, Canopy Characteristics, and Meteorology. Water. 2022; 14(22):3723. https://doi.org/10.3390/w14223723
Chicago/Turabian StyleMa, Changkun, Qian Yao, Haobo Meng, Beibei Zhou, Quanjiu Wang, and Yi Luo. 2022. "Rainfall Partitioning by Evergreen and Deciduous Broad-Leaved Xerophytic Tree Species: Influence of Rainfall, Canopy Characteristics, and Meteorology" Water 14, no. 22: 3723. https://doi.org/10.3390/w14223723
APA StyleMa, C., Yao, Q., Meng, H., Zhou, B., Wang, Q., & Luo, Y. (2022). Rainfall Partitioning by Evergreen and Deciduous Broad-Leaved Xerophytic Tree Species: Influence of Rainfall, Canopy Characteristics, and Meteorology. Water, 14(22), 3723. https://doi.org/10.3390/w14223723