

Small Organisms in a Large River: What Provides the High Diversity of Scaled Chrysophytes in the Ob River?
 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Field and Laboratory Methods
3. Results
3.1. Water Parameters
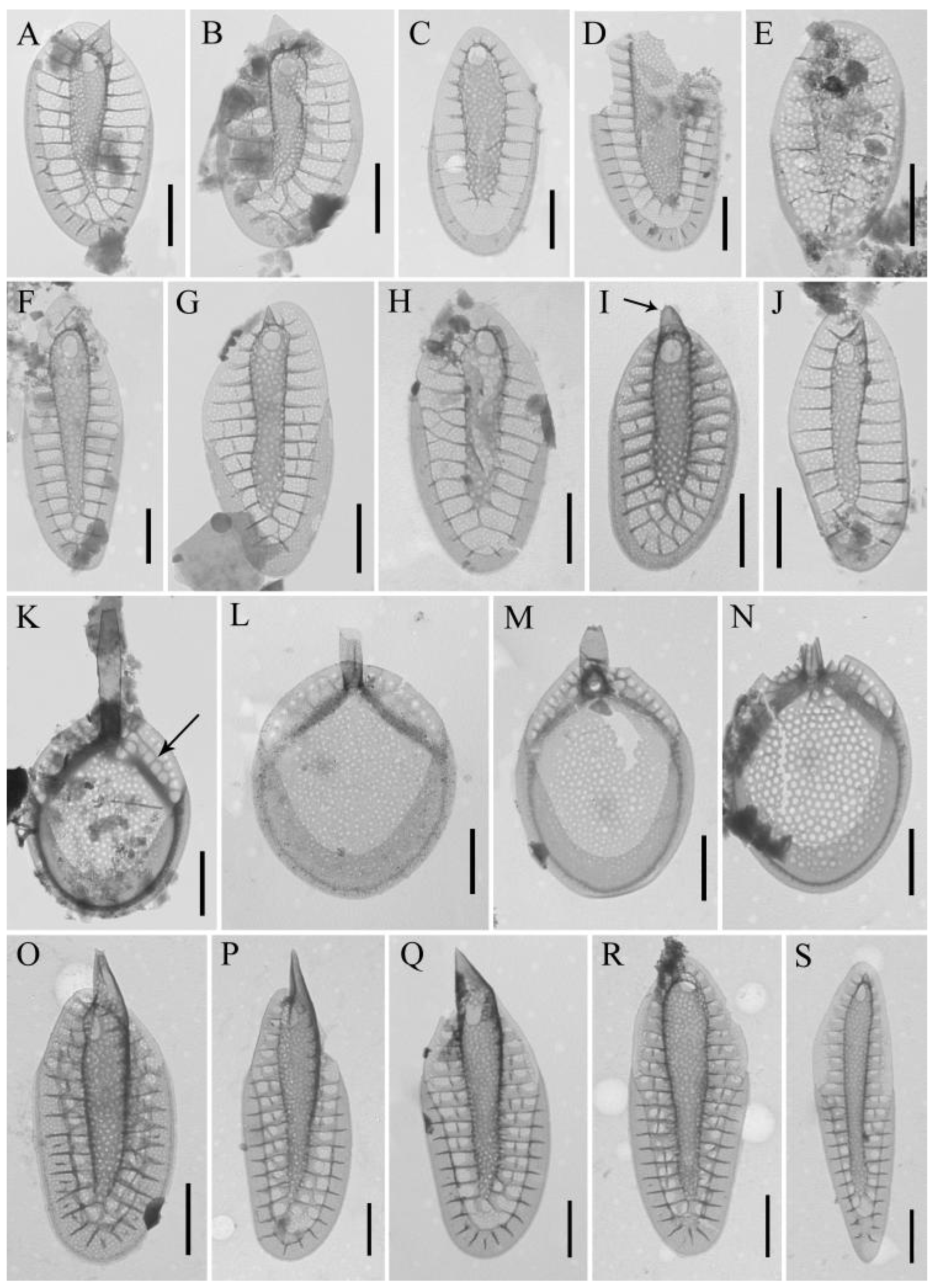
3.2. Diversity and Distribution of Chrysophytes
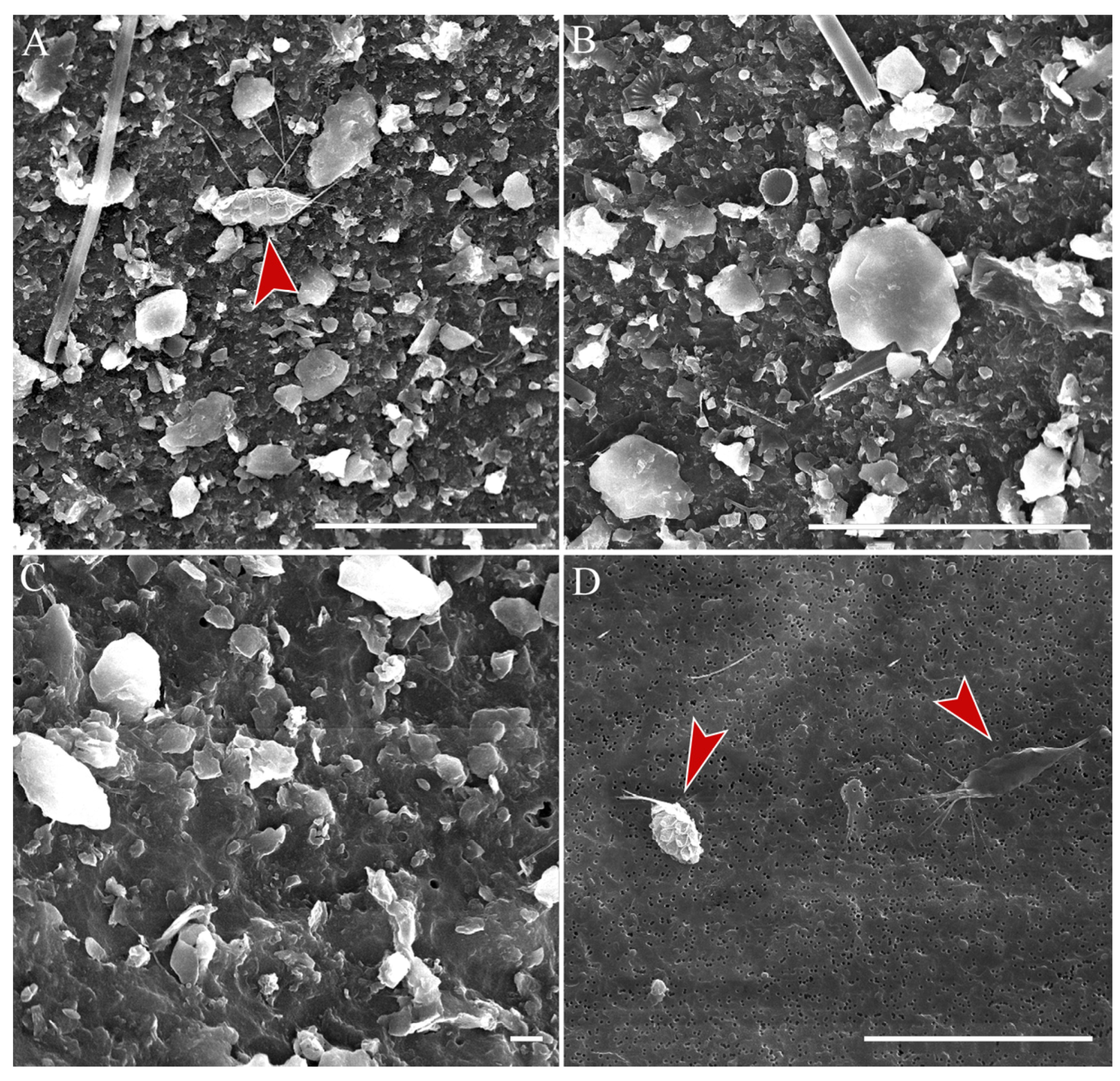
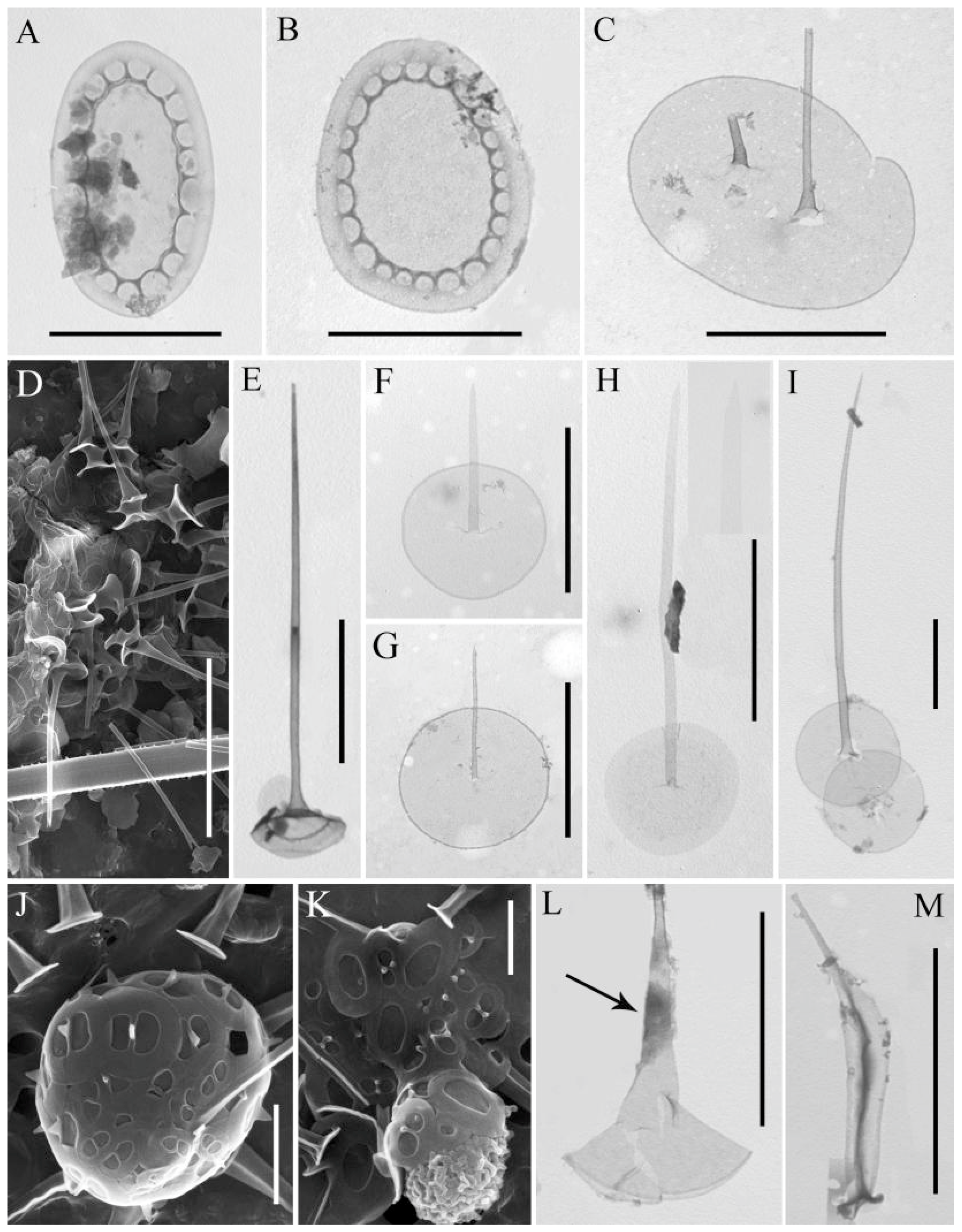
3.3. Rare Species and Species with Specific Morphology
3.4. Undetermined Species
3.5. Stomatocysts
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Finlay, B.J.; Clarke, K.J. Ubiquitous dispersal of microbial species. Nature 1999, 400, 828. [Google Scholar] [CrossRef]
- Finlay, B.J. Global dispersal of free-living microbial eukaryote species. Science 2002, 296, 1061–1063. [Google Scholar] [CrossRef] [PubMed]
- O’Malley, M.A. ‘Everything is everywhere: But the environment selects’: Ubiquitous distribution and ecological determinism in microbial biogeography. Stud. Hist. Philos. Biol. Biomed. Sci. 2008, 39, 314–325. [Google Scholar] [CrossRef]
- Field, C.B.; Behrenfeld, M.J.; Randerson, J.T.; Falkowski, P.G. Primary production of the biosphere: Integrating terrestrial and oceanic components. Science 1998, 281, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Benoiston, A.-S.; Ibarbalz, F.M.; Bittner, L.; Guidi, L.; Jahn, O.; Dutkiewicz, S.; Bowler, C. The evolution of diatoms and their biogeochemical functions. Phil. Trans. R. Soc. B 2017, 372, 20160397. [Google Scholar] [CrossRef] [PubMed]
- Strelnikova, N.I.; Gladenkov, A.Y. Diatoms and their application in stratigraphic and paleogeographic studies. Issues Mod. Algol. 2019, 2, 1–38. [Google Scholar] [CrossRef]
- Duff, K.E.; Zeeb, B.A.; Smol, J.P. Chrysophyte cyst biogeographical and ecological distributions: A synthesis. J. Biogeogr. 1997, 24, 791–812. [Google Scholar] [CrossRef]
- Kristiansen, J. Dispersal and biogeography of silica-scaled chrysophytes. Biodiv. Conserv. 2008, 17, 419–426. [Google Scholar] [CrossRef]
- Harwood, D.M.; Gersonde, R. Lower Cretaceous diatoms from ODP Leg 113 Site 693 (Wendell Sea). Part 2: Resting spores, Chrysophycean cysts, and endoskeletal dinoflagellate, and notes on the origin of diatoms. In Proceedings of the Ocean Drilling Program, Science Result; Barker, P.F., Kennett, J.P., Eds.; Ocean Drilling Program: College Station, TX, USA, 1990; pp. 403–425. [Google Scholar]
- Boo, S.M.; Kim, H.S.; Shin, W.; Boo, G.H.; Cho, S.M.; Jo, B.Y.; Kim, J.H.; Kim, J.H.; Yang, E.C.; Siver, P.A.; et al. Complex phylogeographic patterns in the freshwater alga Synura provide new insights into ubiquity vs. endemism in microbial eukaryotes. Mol. Ecol. 2010, 19, 4328–4338. [Google Scholar] [CrossRef]
- Siver, P.A.; Wolfe, A.P. Scaled chrysophytes in middle Eocene lake sediments from northwestern Canada, including descrip-tions of six new species. Nova Hedwig. Beih. 2005, 128, 295–308. [Google Scholar]
- Siver, P.A.; Wolfe, A.P. Eocene scaled chrysophytes with pronounced modern affinities. Int. J. Plant Sci. 2005, 166, 533–536. [Google Scholar] [CrossRef]
- Siver, P.A.; Lott, A.M.; Wolfe, A.P. A summary of Synura taxa in early Cenozoic deposits from northern Canada. Nova Hedwig. Beih. 2013, 142, 181–190. [Google Scholar]
- Siver, P.A.; Jo, B.Y.; Kim, J.I.; Shin, W.; Lott, A.M.; Wolfe, A.P. Assessing the evolutionary history of the class Synurophyceae (Heterokonta) using molecular, morphometric, and paleobiological approaches. Am. J. Bot. 2015, 102, 921–941. [Google Scholar] [CrossRef] [PubMed]
- Siver, P.A. Synurophyte algae. In Freshwater Algae of North America: Ecology and Classification, 2nd ed.; Academic Press: Boston, MA, USA, 2015; pp. 607–651. [Google Scholar]
- Siver, P.A.; Vigna, M.S. The distribution of scaled chrysophytes in the delta region of the Parana River, Argentina. Nova Hedwig. 1997, 64, 421–453. [Google Scholar] [CrossRef]
- Bessudova, A.; Sorokovikova, L.M.; Tomberg, I.V.; Likhoshway, Y.V. Silica-scaled chrysophytes in large tributaries of Lake Baikal. Cryptogam. Algol. 2018, 39, 145–165. [Google Scholar] [CrossRef]
- Bessudova, A.; Gabyshev, V.A.; Firsova, A.D.; Gabysheva, O.I.; Bukin, Y.U.; Likhoshway, Y.V. Diversity of silica-scaled chrysophytes and physicochemical parameters of their environment in the estuaries of rivers in the Arctic watershed of Yakutia, Russia. Sustainability 2021, 13, 13768. [Google Scholar] [CrossRef]
- Sandgren, C.D.; Smol, J.P.; Kristiansen, J. Chrysophyte Algae: Ecology, Phylogeny and Development; Cambridge University Press: New York, NY, USA, 1995; 399p. [Google Scholar]
- Scoble, J.M.; Cavalier-Smith, T. Scale evolution in Paraphysomonadida (Chrysophyceae): Sequence phylogeny and revised taxonomy of Paraphysomonas, new genus Clathromonas, and 25 new species. Europ. J. Protistol. 2014, 50, 551–592. [Google Scholar] [CrossRef]
- Bessudova, A.; Firsova, A.; Bukin, Y.; Kopyrina, L.; Zakharova, Y.; Likhoshway, Y. Under-ice development of silica-scaled chrysophytes with different trophic mode in two ultraoligotrophic lakes of Yakutia. Diversity 2023, 15, 326. [Google Scholar] [CrossRef]
- Siver, P.A. The distribution of scaled chrysophytes along a pH gradient. Can. J. Bot. 1989, 67, 2120–2130. [Google Scholar] [CrossRef]
- Paterson, A.M.; Cumming, B.F.; Smol, J.P.; Hall, R.I. Scaled chrysophytes as indicators of water quality changes since preindustrial times in the Muskoka-Haliburton region, Ontario, Canada. Can. J. Fish. Aquat. Sci. 2001, 58, 2468–2481. [Google Scholar] [CrossRef]
- Kristiansen, J. Cosmopolitan chrysophytes. Syst. Geog. Plants 2000, 70, 291–300. [Google Scholar] [CrossRef]
- Škaloud, P.; Škaloudová, M.; Pichrtová, M.; Němcová, Y.; Kreidlová, J.; Pusztai, M. www.chrysophytes.eu—A database on distribution and ecology of silica-scaled chrysophytes in Europe. Nova Hedwig. Beih. 2013, 142, 141–146. [Google Scholar]
- Siver, P.A. The distribution and variation of Synura species (Chrysophyceae) in Connecticut, USA. Nord. J. Bot. 1987, 7, 107–116. [Google Scholar] [CrossRef]
- Schlichting, H.E. The role of waterfowl in the dispersal of algae. Trans. Am. Microsc. Soc. 1960, 79, 160–166. [Google Scholar] [CrossRef]
- Siver, P.A.; Lott, A.M. Biogeographic patterns in scaled chrysophytes from east coast of North America. Freshw. Biol. 2012, 57, 451–466. [Google Scholar] [CrossRef]
- Bessudova, A.; Bukin, Y.; Likhoshway, Y.V. Dispersal of silica-scaled chrysophytes in northern water bodies. Diversity 2021, 13, 284. [Google Scholar] [CrossRef]
- Bulavina, A.S. Modelling of the Ob River runoff under climate change. Tr. KSC RAS 2021, 3, 18–28. [Google Scholar] [CrossRef]
- Bulavina, A.S. Spatio-temporal analysis of climatic factors of the Ob River runoff formation. Tr. KSC RAS 2022, 4, 15–26. [Google Scholar] [CrossRef]
- Aleshina, N.I.; Gefke, I.V. Features of hydrological regime of the Upper Ob for the possibility of water management. Intern. J. Hum. Nat. Sci. 2019, 11–12, 57–60. [Google Scholar] [CrossRef]
- Broecker, W.S. Thermohaline circulation, the Achilles heel of our climate system: Will man-made CO2 upset the current balance. Science 1997, 278, 1582–1588. [Google Scholar] [CrossRef]
- Kulakov, M.Y. A new approach for modelling of water circulation in Arctic Seas. Probl. Arct. Antarct. 2012, 2, 55–62. [Google Scholar]
- Soromotin, A.; Moskovchenko, D.; Khoroshavin, V.; Prikhodko, N.; Puzanov, A.; Kirillov, V.; Koveshnikov, M.; Krylova, E.N.; Krasnenko, A.; Pechkin, A. Major, trace and rare earth element distribution in water, suspended particulate matter and stream sediments of the Ob River mouth. Water 2022, 14, 2442. [Google Scholar] [CrossRef]
- Wetzel, R.G. Limnological Analysis, 3rd ed.; Springer: New York, NY, USA, 2000; 430p. [Google Scholar]
- Hillard, D.K.; Asmund, B. Studies on chrysophyceae from some ponds and lakes in Alaska. II. Notes on the genera Dinobryon, Hyalobryon and Epipyxis with descriptions of new species. Hydrobiologia 1963, 22, 331–397. [Google Scholar] [CrossRef]
- Asmund, B. Studies on Chrysophyceae from some ponds and lakes in Alaska. VI. Occurrence of Synura species. Hydrobiologia 1968, 31, 497–515. [Google Scholar] [CrossRef]
- Balonov, I.M. Genus Synura Ehr. (Chrysophyta): Biology, ecology and systematic. In Biology, Morphology and Systematics of the Aquatic Organisms; Science: Leningrad, Russia, 1976; pp. 61–81. [Google Scholar]
- Bessudova, A.; Bukin, Y.S.; Sorokovikova, L.M.; Firsova, A.D.; Tomberg, I.V. Silica-scaled chrysophytes in small lakes of the lower Yenisei basin, the Arctic. Nova Hedwig. 2018, 107, 315–336. [Google Scholar] [CrossRef]
- Kristiansen, J.; Preisig, H.R. Chrysophyta and Haptophyta algae, 2nd part: Synurophyceae. In Süsswasser Flora Von Mitteleuropa (Freshwater Flora of Central Europe); Springer: Berlin, Germany, 2007; pp. 1–252. [Google Scholar]
- Nicholls, K.N. Spiniferomonas (Chrysophyceae) in Ontario lakes including a revision and descriptions of two new species. Can. J. Bot. 1981, 59, 107–117. [Google Scholar] [CrossRef]
- Nicholls, K.N. Spiniferomonas septispina and S. enigmata, two new algal species confusing the distinction between Spiniferomonas and Chrysosphaerella (Chrysophyceae). Pl. Syst. Evol. 1984, 148, 103–117. [Google Scholar] [CrossRef]
- Kristiansen, J.; Tong, D. Chrysosphaerella annulata n. sp., a new scale-bearing chrysophyte. Nord. J. Bot. 1989, 9, 329–332. [Google Scholar] [CrossRef]
- Duff, K.E.; Zeeb, B.A.; Smol, J.P. Altas of Chrysophycean Cysts; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1995; pp. 1–189. [Google Scholar] [CrossRef]
- Wilkinson, A.N.; Zeeb, B.A.; Smol, J.P. Atlas of Chrysophycean Cysts; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2001; Volume II, pp. 1–169. [Google Scholar] [CrossRef]
- Piątek, J.; Piątek, M. Chrysophyte stomatocysts of the sulphuric salt marsh in the Owczary Reserve (Central Poland). Pol. Bot. J. 2005, 50, 97–106. [Google Scholar]
- Piątek, J. Chrysophyte stomatocysts from sediments in a manmade water reservoir in central Poland. Ann. Bot. Fennici. 2007, 44, 186–193. [Google Scholar]
- Pang, W.; Wang, Y.; Wang, Q. Ten new chrysophycean stomatocysts ornamented with spines from bogs near Da’erbin Lake, China. Nova Hedwig. 2012, 94, 193–207. [Google Scholar] [CrossRef]
- Pang, W.; Wang, Q. Chrysophycean stomatocysts from Xinjiang Province, China. Phytotaxa 2016, 288, 41–50. [Google Scholar] [CrossRef]
- Safronova, T.V. Golden Algae (Chrysophyceae, Synurophyceae) of Nature Reserves of Leningrad Oblast and Sankt-Petersburg. Ph.D. Thesis, BIN RAN, Sankt-Petersburg, Russia, 2018; pp. 1–204. [Google Scholar]
- Firsova, A.D.; Bessudova, A.Y.; Likhoshway, Y.V. Chrysophycean stomatosysts in tributaries of northern limit of Lake Baikal. Acta Biol. Sib. 2018, 4, 25–44. [Google Scholar]
- Firsova, A.D.; Bessudova, A.Y.; Sorokovikova, L.M.; Zhychenko, N.A.; Istomina, N.A.; Sezko, N.P.; Likhoshway, Y.V. Stomatocyst diversity in the first years of the plankton species structure formation in Reservoir of Hydropower Plants (Boguchany Reservoir, Russia). Phytotaxa 2019, 424, 18–32. [Google Scholar] [CrossRef]
- Firsova, A.D.; Bessudova, A.Y.; Kopyrina, L.I.; Likhoshway, Y.V. Chrysophycean stomatocysts from two unique lakes of Yakutia (Russia). Phytotaxa 2020, 474, 197–217. [Google Scholar] [CrossRef]
- Ignatenko, M.; Yatsenko-Stepanova, T.; Kapustin, D. Morphological variability of stomatocyst 131 Pang & Wang (Chrysophyceae) from a freshwater shallow lake in South Urals, Russia. Phytotaxa 2022, 542, 100–104. [Google Scholar] [CrossRef]
- Bessudova, A.Y.; Sorokovikova, L.M.; Sinyukovich, V.N.; Firsova, A.D.; Tomberg, I.V.; Likhoshway, Y.V. Effects of water levels on species diversity of silica-scaled chrysophytes in large tributaries of Lake Baikal. Acta Biol. Sib. 2020, 55, 7–22. [Google Scholar] [CrossRef]
- Siver, P.A.; Voloshko, L.N.; Gavrilova, O.V.; Getsen, M.V. The scaled chrysophyte flora of the Bolshezemelskaya tundra. Nova Hedwig. 2005, 128, 125–150. [Google Scholar]
- Bessudova, A.; Gabyshev, V.; Bukin, Y.; Gabysheva, O.; Likhoshway, Y.V. Species richness of scaled chrysophytes in arctic waters in the Tiksi Region (Yakutia, Russia). Acta Biol. Sib. 2022, 8, 431–459. [Google Scholar]
- Bessudova, A.; Domysheva, V.M.; Firsova, A.D.; Likhoshway, Y.V. Silica-scaled chrysophytes of Lake Baikal. Acta Biol. Sib. 2017, 3, 47–56. [Google Scholar] [CrossRef]
- Bessudova, A.; Tomberg, I.V.; Firsova, A.D.; Kopyrina, L.I.; Likhoshway, Y.V. Silica-scaled chrysophytes in lakes Labynkyr and Vorota of the Sakha (Yakutia) Republic, Russia. Nova Hedwig. Beih. 2019, 148, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Firsova, A.D.; Bessudova, A.; Sorokovikova, L.M.; Tomberg, I.V.; Likhoshway, Y.V. The diversity of chrysophycean algae in an arctic zone of river and sea water mixing, Russia. Am. J. Plant Sci. 2015, 6, 2439–2452. [Google Scholar] [CrossRef]
- Bessudova, A.; Firsova, A.D.; Tomberg, I.V.; Sorokovikova, L.M.; Likhoshway, Y.V. Biodiversity of silica-scaled chrysophytes in tributaries ofnorthern limit of Lake Baikal. Acta Biol. Sib. 2018, 4, 75–84. [Google Scholar] [CrossRef]
- Nĕmcová, Y.; Pusztai, M.; Škaloudová, M.; Neustupa, J. Silica-scaled chrysophytes (Stramenopiles, Ochrophyta) along a salinity gradient: A case study from the Gulf of Bothnia western shore (northern Europe). Hydrobiologia 2015, 50, 187–197. [Google Scholar] [CrossRef]
- Siver, P.A. The distribution and ecology of Spiniferomonas (Chrysophyceae) in Connecticut (U.S.A). Nord. J. Bot. 1988, 8, 205–212. [Google Scholar] [CrossRef]
- Firsova, A.D.; Kuzmina, A.E.; Tomberg, I.V.; Potemkina, T.G.; Likhochway, Y.V. Seasonal dynamics of Chrysophyte stomatocyst formation in the plankton of Southern Baikal. Izv. Akad. Nauk. SSSR Seriya Biol. 2008, 5, 589–596. [Google Scholar] [CrossRef]
- Firsova, A.D.; Bessudova, A.; Likhoshway, Y.V. New data of chrysophycean stomatocysts from Lake Baikal. Acta Biol. Sib. 2017, 3, 113. [Google Scholar] [CrossRef]
- Cronberg, G. Changes in phytoplankton of Lake Trummen induced by restoration. Hydrobiologia 1982, 86, 185–193. [Google Scholar] [CrossRef]
- Sandgren, C.D. The ecology of chrysophyte flagellates: Their growth and perennation strategies as freshwater phytoplankton. In Growth and Reproductive Strategies of Freshwater Phytoplankton; Sandgren, C.D., Ed.; Cambridge university Press: Cambridge, UK, 1988; pp. 9–104. [Google Scholar]
- Sandgren, C.D. Chrysophyte reproduction and resting cysts: A paleolimnologist’s primer. J. Paleol. 1991, 5, 1–9. [Google Scholar] [CrossRef]
- Korkonen, S. Chrysophyte Stomatocysts and Their Biogeography in Finland; Academic Dissertation: Helsinki, Finland, 2020; pp. 1–65. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| River Transect | St. No. | Station Position in the Riverbed | Coordinates N/E | Date of Sampling dd.mm.yy | Time of Sampling hh:mm | Hmax,m | h, m | T, °C | pH | S, m | V, m·s−1 | O2 mg·L | General View of the Sample in the SEM, Magnification ×1500–1600 | Diatoms Abundance, 103 Cells·L−1/ Biomass, g·m−3 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Kazym- Mys | K1 | Left bank | 64.6555° 65.6333° | 8 June 2022 | 08:35 | 18.8 | 3.8 | 13.1 | 7.31 | 0.54 | 0.12 | 9.5 |  | 90.4/1.27 |
| K2 | The center of the riverbed | 64.6532° 65.6333° | 8 June 2022 | 10:00 | 15.8 | 3.0 | 13.1 | 7.30 | 0.45 | 0.22 | 9.58 |  | 57.1/1.08 | |
| K3 | Right bank | 64.6655° 65.6391° | 8 June 2022 | 11:00 | 7.0 | 1.4 | 13.1 | 7.20 | 0.55 | 0.19 | 9.35 |  | 91.2/0.86 | |
| K4 | Shallow bay | 64.6666° 65.6166° | 7 June 2022 | 11:30 | 0.5 | 0.1 | 14.0 | - | to the bottom | 0.0 | - |  | 21.6/0.03 | |
| Azovy | A1 | Left bank | 64.8666° 65.1833° | 9 June 2022 | 10:30 | 10.1 | 2.0 | 14.33 | 7.52 | 0.50 | 0.03 | 9.23 |  | 48.5/0.78 |
| A2 | The center of the riverbed | 64.8769° 65.1906° | 9 June 2022 | 15:00 | 22.7 | 4.5 | 14.27 | 7.53 | 0.50 | 0.15 | 9.50 |  | 45.6/0.81 | |
| A3 | Right bank | 64.8666° 65.1910° | 9 June 2022 | 16:30 | 22.0 | 4.4 | 14.33 | 7.48 | 0.45 | 0.35 | 9.18 |  | 92.0/0.62 | |
| A4 | Right bank | 64.8789° 65.1956° | 9 June 2022 | 09:30 | 0.5 | 0.0 | 14.33 | - | to the bottom | −0.1 reverse flow | 8.81 |  | 50.4/0.58 | |
| Salemal | S1 | Left bank, tributaries | 66.7932° 68.8884° | 12 June 2022 | 16:30 | 1.9 | 0.1 | 13.0 | - | 0.35 | 0–0.15 | 9.85 |  | 208.5/2.48 |
| S2 | Left bank | 66.7831° 68.9657° | 12 June 2022 | 24:00 | 16.0 | 3.2 | 12.9 | 7.87 | 0.50 | 0.20 | 10.18 |  | 92.4/1.17 | |
| S3 | To the left of the riverbed center | 66.7804° 68.9658° | 12 June 2022 | 22:30 | 25.0 | 5.0 | 14.4 | 7.82 | 0.54 | 0.26 | 9.37 |  | 61.0/0.90 | |
| S4 | The center of the riverbed | 66.7769° 68.9660° | 12 June 2022 | 19:10 | 27.0 | 5.4 | 14.4 | 7.78 | 0.57 | 0.38 | 9.71 |  | 62.7/0.80 | |
| S5 | The right of the center of the riverbed | 66.7741° 68.9663° | 12 June 2022 | 16:30 | 25.0 | 5.0 | 14.33 | 7.77 | 0.60 | 0.27 | 9.83 |  | 70.4/0.80 | |
| S6 | Right bank | 66.7715° 68.9665° | 12 June 2022 | 13:00 | 18.0 | 3.8 | 14.15 | 7.77 | 0.60 | 0.30 | - |  | 38.5/0.52 |
| Site Species | K1 | K2 | K3 | K4 | A1 | A2 | A3 | A4 | S2 | S3 | S4 | S5 | S6 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1. | Chrysosphaerella coronacircumspina Wujek and Kristiansen in Wujek, Gretz and Wujek | + | ||||||||||||
| 2. | Chrysosphaerella sp. | + | + | + | ||||||||||
| 3. | Paraphysomonas gladiata Preisig and Hibberd | + | + | + | ||||||||||
| 4. | P. cf. vulgaris vulgaris Scoble andCavalier-Smith | + | + | |||||||||||
| 5. | P. uniformis uniformis Scoble and Cavalier-Smith | + | + | + | + | |||||||||
| 6. | Paraphysomonas sp. 1 | + | ||||||||||||
| 7. | Paraphysomonas sp. 2 | + | ||||||||||||
| 8. | Spiniferomonas abrupta Nielsen | + | ||||||||||||
| 9. | S. bourrellyi Takahashi | + | + | + | + | + | + | |||||||
| 10. | S. cornuta Balonov | + | ||||||||||||
| 11. | S. minuta Nicholls | + | + | |||||||||||
| 12. | S. serrata Nicholls | + | + | |||||||||||
| 13. | S. silverensis Nicholls | + | ||||||||||||
| 14. | S. takahashii Nicholls | + | ||||||||||||
| 15. | S. triangularis Siver | + | ||||||||||||
| 16. | S. trioralis Takahashi | + | + | + | + | + | + | + | ||||||
| 17. | Spiniferomonas sp. | + | ||||||||||||
| 18. | Mallomonas acaroides Perty | + | + | + | + | + | + | + | + | + | ||||
| 19. | M. actinoloma Asmund and Takahashi | + | + | |||||||||||
| 20. | M. actinoloma var. maramuresensis Asmund and Takahashi | + | + | + | ||||||||||
| 21. | M. annulata Harris | + | + | + | ||||||||||
| 22. | M. akrokomos Ruttner | + | + | + | + | |||||||||
| 23. | M. alpina Pascher and Ruttner | + | + | + | + | |||||||||
| 24. | M. alata Asmund | + | + | + | ||||||||||
| 25. | M. alata f. hualvensis Asmund, Cronberg and Dürrschmidt | + | + | + | ||||||||||
| 26. | M. elongata Reverdin | + | + | |||||||||||
| 27. | M. eoa Takahashi in Asmund and Takahashi | + | + | |||||||||||
| 28. | M. caudata Iwanoff | + | + | + | + | |||||||||
| 29. | M. calceolus Bradley | + | ||||||||||||
| 30. | M. crassisquama (Asmund) Fott | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 31. | M. crassisquama var. papillosa Siver and Skogstad | + | + | + | + | |||||||||
| 32. | M. costata Dürrschmidt | + | + | |||||||||||
| 33. | M. heterospina Lund | + | + | + | ||||||||||
| 34. | M. insignis Pénard | + | ||||||||||||
| 35. | M. lychenensis Conrad | + | + | + | ||||||||||
| 36. | M. mangofera var. mangofera Harris and Bradley | + | + | |||||||||||
| 37. | M. multiunca Asmund | + | + | |||||||||||
| 38. | M. munda (Asmund, Cronberg and Dürrschmidt) Nemcova | + | ||||||||||||
| 39. | M. papillosa Harris and Bradley | + | + | + | ||||||||||
| 40. | M. punctifera Korshikov | + | + | + | + | + | + | + | + | + | + | |||
| 41. | M. kalinae Řezáčová | + | + | |||||||||||
| 42. | M. kuzminii Gusev and Kulikovskiy | + | ||||||||||||
| 43. | M. scalaris Dürrschmidt | + | ||||||||||||
| 44. | M. scrobiculata Nicholls | + | ||||||||||||
| 45. | M. striata var. serrata Harris and Bradley | + | + | + | ||||||||||
| 46. | M. tonsurata Teiling | + | + | + | + | + | + | + | + | |||||
| 47. | M. torquata var. torquata Asmund and Cronberg | + | ||||||||||||
| 48. | M. trummensis Cronberg | + | + | + | ||||||||||
| 49. | Mallomonas sp. | + | ||||||||||||
| 50. | Synura americana Škaloud, Kynčlová, Benada, Kofroňová, Škaloudová | + | + | + | + | + | + | + | + | + | + | + | ||
| 51. | S. cf. asmundiae (Cronberg and Kristiansen) Škaloud, Kristiansen and Škaloudová | + | ||||||||||||
| 52. | S. conopea Škaloudand Kynclová | + | ||||||||||||
| 53. | S. glabra (Korshikov) Škaloud and Kynclová | + | + | + | + | + | + | + | ||||||
| 54. | S. echinulata Korshikov | + | + | + | ||||||||||
| 55. | S. biseriata Balonov | + | + | + | ||||||||||
| 56. | S. borealis Škaloud and Škaloudová | + | + | + | + | + | ||||||||
| 57. | S. cf. hibernica Škaloud and Škaloudová | + | + | |||||||||||
| 58. | S. petersenii (Korshikov) Škaloud and Kynclová | + | + | + | + | + | + | + | + | + | ||||
| 59. | S. praefracta (Asmund) Škaloud and Škaloudová | + | ||||||||||||
| 60. | S. punctulosa Balonov | + | + | + | + | + | ||||||||
| 61. | S. multidentata Péterfi and Momeu | + | + | + | ||||||||||
| 62. | S. spinosa Korshikov | + | + | + | + | |||||||||
| 63. | S. spinosa f. longispina Petersen and Hansen | + | ||||||||||||
| 64. | S. splendida Korshikov | + | ||||||||||||
| 65. | S. uvella Ehrenberg | + | + | + | + | |||||||||
| 66. | Synura sp. 1 | + | + | + | ||||||||||
| 67. | Synura sp. 2 | + | + | |||||||||||
| TOTAL | 8 | 7 | 19 | 54 | 22 | 9 | 13 | 31 | 11 | 5 | 11 | 6 | 13 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bessudova, A.; Likhoshway, Y.; Firsova, A.; Mitrofanova, E.; Koveshnikov, M.; Soromotin, A.; Khoroshavin, V.; Kirillov, V. Small Organisms in a Large River: What Provides the High Diversity of Scaled Chrysophytes in the Ob River? Water 2023, 15, 3054. https://doi.org/10.3390/w15173054
Bessudova A, Likhoshway Y, Firsova A, Mitrofanova E, Koveshnikov M, Soromotin A, Khoroshavin V, Kirillov V. Small Organisms in a Large River: What Provides the High Diversity of Scaled Chrysophytes in the Ob River? Water. 2023; 15(17):3054. https://doi.org/10.3390/w15173054
Chicago/Turabian StyleBessudova, Anna, Yelena Likhoshway, Alena Firsova, Elena Mitrofanova, Mikhail Koveshnikov, Andrey Soromotin, Vitaly Khoroshavin, and Vladimir Kirillov. 2023. "Small Organisms in a Large River: What Provides the High Diversity of Scaled Chrysophytes in the Ob River?" Water 15, no. 17: 3054. https://doi.org/10.3390/w15173054
APA StyleBessudova, A., Likhoshway, Y., Firsova, A., Mitrofanova, E., Koveshnikov, M., Soromotin, A., Khoroshavin, V., & Kirillov, V. (2023). Small Organisms in a Large River: What Provides the High Diversity of Scaled Chrysophytes in the Ob River? Water, 15(17), 3054. https://doi.org/10.3390/w15173054