




In Vitro Assessment of Salinity Stress Impact on Early Growth in Ten Certified Palestinian Barley Cultivars (Hordeum vulgare L.) Potentially Suitable for Cultivation on Former Quarry Substrates


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Area of Study and Soil Sampling
2.3. Methods
2.3.1. Plant Seed Disinfection and Germination
2.3.2. Electrical Conductivity (EC) and Salinity Contents of the Soil
2.3.3. Statistical Analysis
3. Results
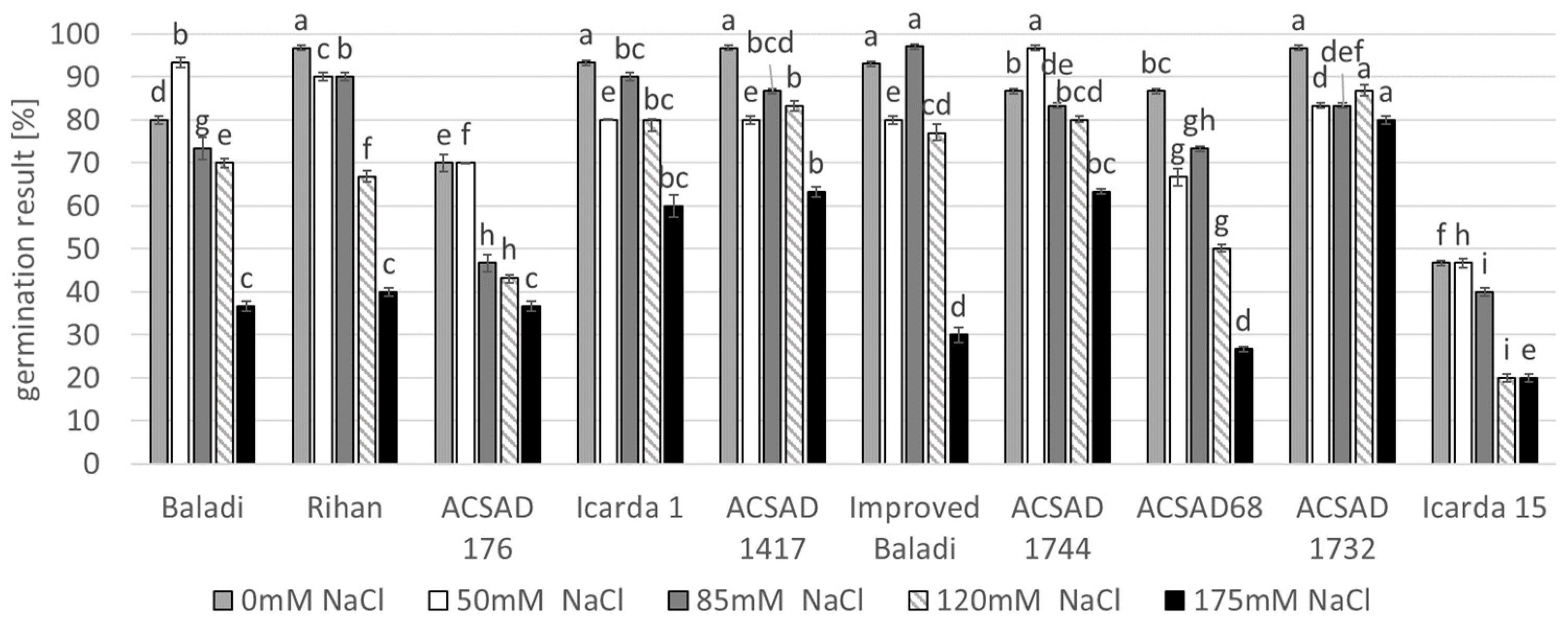
3.1. Effect of Salinity Stress on Seed Germination of Barley
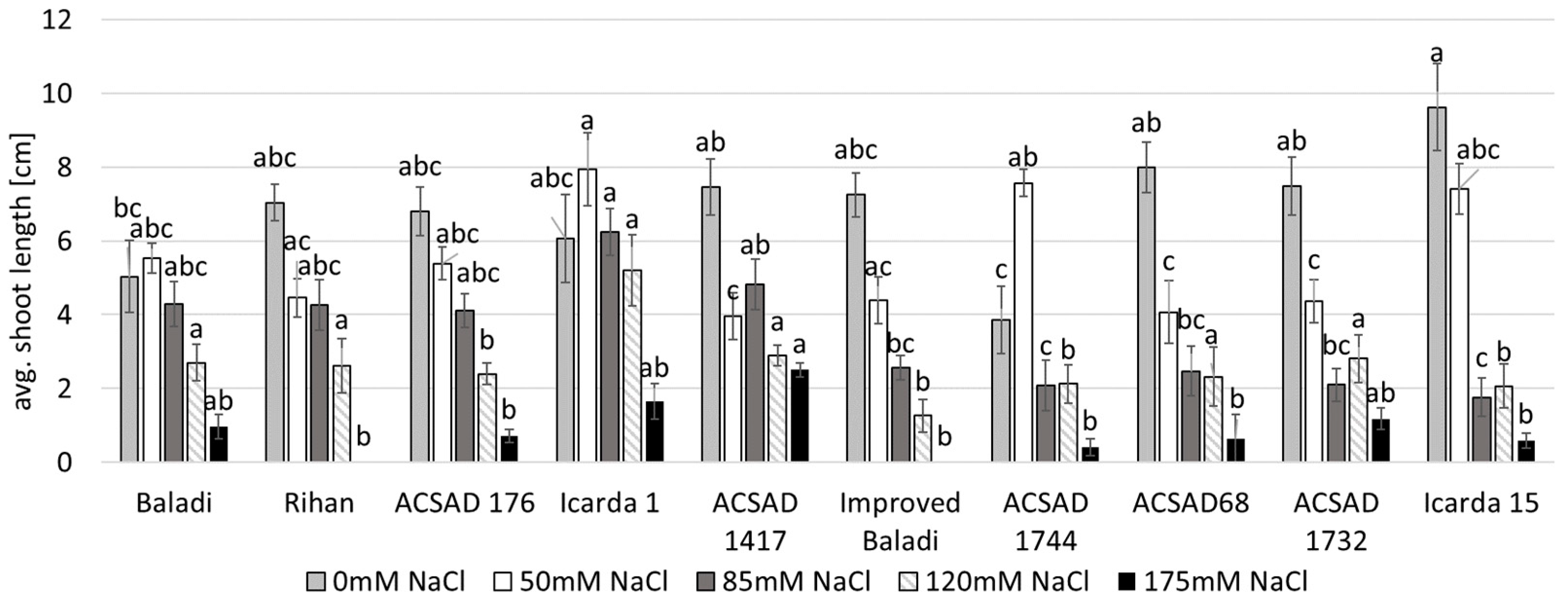
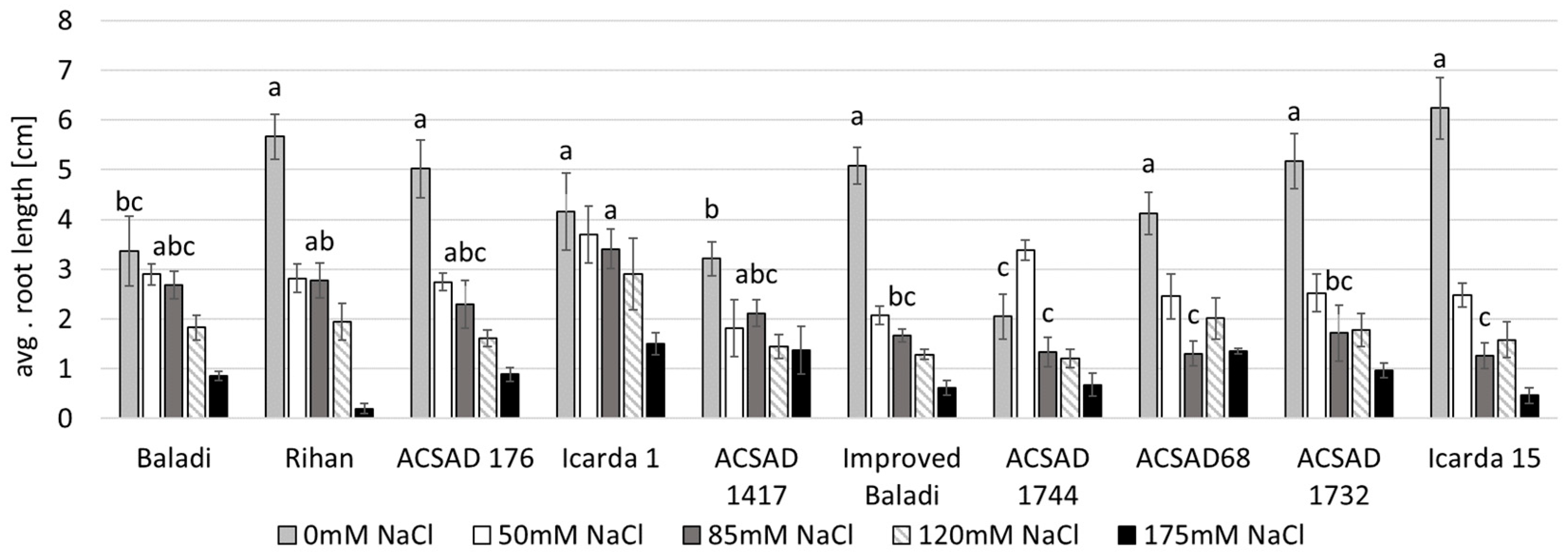
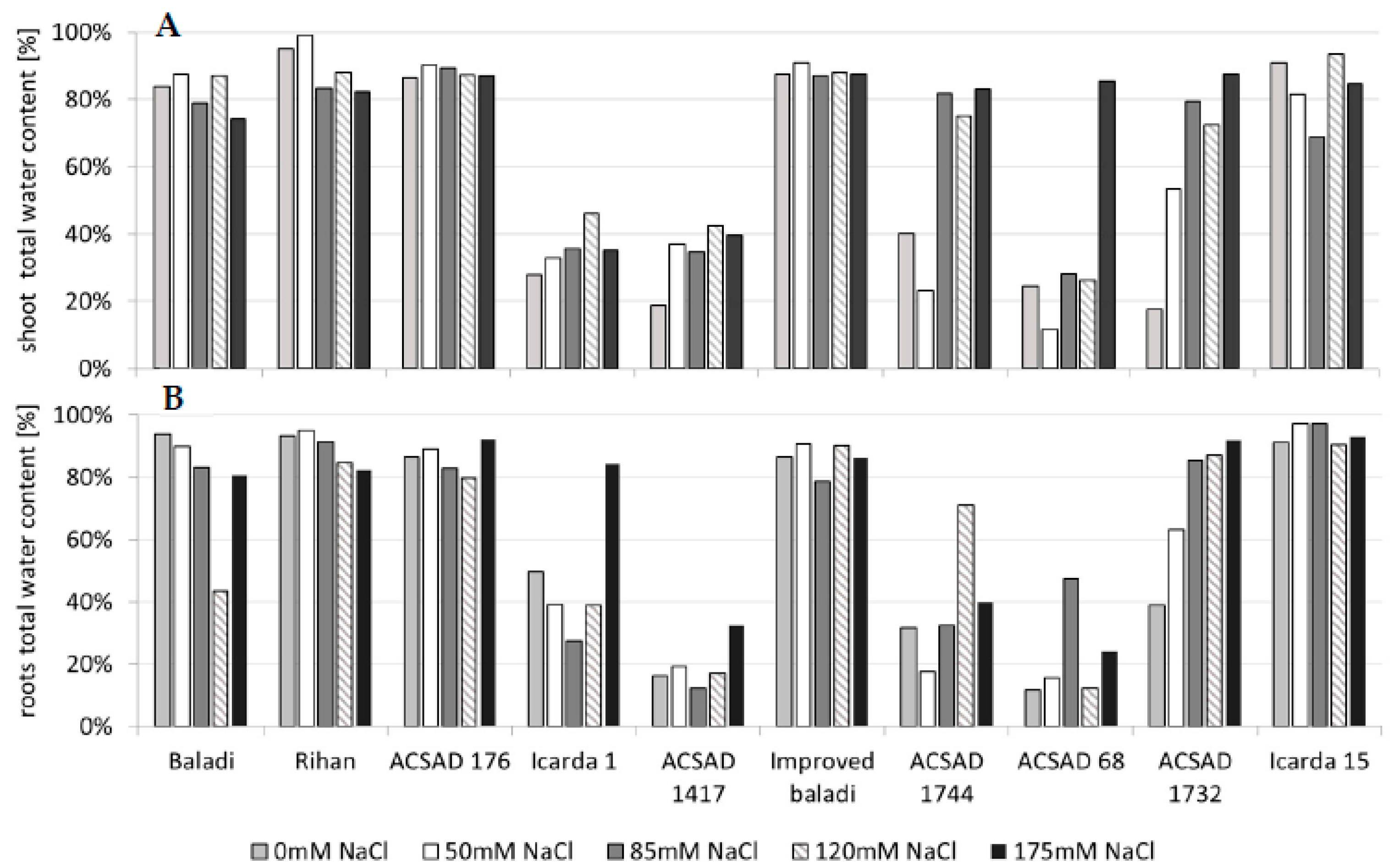
3.2. Effect of Salinity Stress on Water Content and Biomass
3.3. Electrical Conductivity for Quarry Substrate
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ali, B.; Saleem, M.H.; Ali, S.; Shahid, M.; Sagir, M.; Tahir, M.B.; Qureshi, K.A.; Jaremko, M.; Selim, S.; Hussain, A.; et al. Mitigation of Salinity Stress in Barley Genotypes with Variable Salt Tolerance by Application of Zinc Oxide Nanoparticles. Front. Plant Sci. 2022, 13, 2850. [Google Scholar] [CrossRef]
- Raza, A.; Tabassum, J.; Fakhar, A.Z.; Sharif, R.; Chen, H.; Zhang, C.; Ju, L.; Fotopoulos, V.; Siddique, K.H.M.; Singh, R.K.; et al. Smart Reprograming of Plants against Salinity Stress Using Modern Biotechnological Tools. Crit. Rev. Biotechnol. 2022, 1–28. [Google Scholar] [CrossRef]
- Cope, J.E.; Norton, G.J.; George, T.S.; Newton, A.C. Evaluating Variation in Germination and Growth of Landraces of Barley (Hordeum vulgare L.) Under Salinity Stress. Front. Plant Sci. 2022, 13, 2093. [Google Scholar] [CrossRef]
- McFarlane, D.J.; George, R.J.; Barrett-Lennard, E.G.; Gilfedder, M. Salinity in Dryland Agricultural Systems: Challenges and Opportunities. In Innovations in Dryland Agriculture; Springer: Berlin/Heidelberg, Germany, 2017; pp. 521–547. [Google Scholar] [CrossRef]
- Podmore, C. Dryland Salinity—Causes and Impacts. Prime Facts J. 2009, 936, 1–6. [Google Scholar]
- Mbarki, S.; Sytar, O.; Cerda, A.; Zivcak, M.; Rastogi, A.; He, X.; Zoghlami, A.; Abdelly, C.; Brestic, M. Strategies to Mitigate the Salt Stress Effects on Photosynthetic Apparatus and Productivity of Crop Plants. In Salinity Responses and Tolerance in Plants, Volume 1: Targeting Sensory, Transport and Signaling Mechanisms; Springer: Berlin/Heidelberg, Germany, 2018; pp. 85–136. [Google Scholar] [CrossRef]
- Artiola, J.F.; Walworth, J.L.; Musil, S.A.; Crimmins, M.A. Soil and Land Pollution. In Environmental and Pollution Science; Elsevier: Amsterdam, The Netherlands, 2019; pp. 219–235. [Google Scholar] [CrossRef]
- Gwisai, R.D.; Areola, O.O.; Segosebe, E.M. Physico-Chemical Analysis in Surface Waters around the Closed Gaborone Sanitary Landfill in Botswana. Environ. Ecol. Res. 2019, 7, 220–238. [Google Scholar] [CrossRef] [Green Version]
- Warrence, N.J.; Bauder, J.W.; Pearson, K.E. Basics of Salinity and Sodicity Effects on Soil Physical Properties; Department of Land Resources and Environmental Sciences: Montana State University: Bozeman, MT, USA, 2002; p. 129. [Google Scholar]
- Bado, S.; Forster, B.; Ghanim, A.; Jankowicz-Cieslak, J. Protocols for Pre-Field Screening of Mutants for Salt Tolerance in Rice, Wheat and Barley; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Najjar, A.A.; Kuhn, A.J.; Al-Tardeh, S.M.; Kuchendorf, C.M. Microalgae and Biochar Agro-Fertilization of the Palestinian Rehan Barley Cultivar under Salinity Stress. Agronomy 2021, 11, 2309. [Google Scholar] [CrossRef]
- McKenzie, R.C.; Sprout, C.H.; Clark, N.F. The Relationship of The Yield of Irrigated Barley to Soil Salinity as Measured by Several Methods. Can. J. Soil Sci. 2011, 63, 519–528. [Google Scholar] [CrossRef]
- Isayenkov, S.V. Genetic Sources for the Development of Salt Tolerance in Crops. Plant Growth Regul. 2019, 89, 1–17. [Google Scholar] [CrossRef]
- Isayenkov, S.V.; Maathuis, F.J.M. Plant Salinity Stress: Many Unanswered Questions Remain. Front. Plant Sci. 2019, 10, 80. [Google Scholar] [CrossRef] [Green Version]
- Abido, W.A.E.; Zsombik, L.; Abido, W.A.E.; Zsombik, L. Effect of Salinity on Germination Characters and Seedlings Parameters of Egyptian Flax Cultivars Growing in Nyiregyhaza; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar] [CrossRef]
- Ali, B.; Hafeez, A.; Ahmad, S.; Javed, M.A.; Afridi, M.S.; Dawoud, T.M.; Almaary, K.S.; Muresan, C.C.; Marc, R.A.; Alkhalifah, D.H.M.; et al. Bacillus Thuringiensis PM25 Ameliorates Oxidative Damage of Salinity Stress in Maize via Regulating Growth, Leaf Pigments, Antioxidant Defense System, and Stress Responsive Gene Expression. Front. Plant Sci. 2022, 13, 2568. [Google Scholar] [CrossRef]
- Ali, B.; Wang, X.; Saleem, M.H.; Hafeez, A.; Afridi, M.S.; Khan, S.; Ullah, I.; Amaral Júnior, A.T.D.; Alatawi, A.; Ali, S. PGPR-Mediated Salt Tolerance in Maize by Modulating Plant Physiology, Antioxidant Defense, Compatible Solutes Accumulation and Bio-Surfactant Producing Genes. Plants 2022, 11, 345. [Google Scholar] [CrossRef] [PubMed]
- Serrano, R.; Mulet, J.M.; Rios, G.; Marquez, J.A.; de Larrinoa, I.F.; Leube, M.P.; Mendizabal, I.; Pascual-Ahuir, A.; Proft, M.; Ros, R.; et al. A Glimpse of the Mechanisms of Ion Homeostasis during Salt Stress. J. Exp. Bot. 1999, 50, 1023–1036. [Google Scholar] [CrossRef]
- al Tardeh, S.; Iraki, N. Morphological and Anatomical Responses of Two Palestinian Tomato (Solanum lycopersicon L.) Cultivars to Salinity during Seed Germination and Early Growth. Acad. J. 2013, 12, 4788–4797. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Irving, L.J.; McGill, C.; Matthew, C.; Zhou, D.; Kemp, P. The Effects of Salinity and Osmotic Stress on Barley Germination Rate: Sodium as an Osmotic Regulator. Ann. Bot. 2010, 106, 1027–1035. [Google Scholar] [CrossRef] [Green Version]
- Sarraf, M.; Vishwakarma, K.; Kumar, V.; Arif, N.; Das, S.; Johnson, R.; Janeeshma, E.; Puthur, J.T.; Aliniaeifard, S.; Chauhan, D.K.; et al. Metal/Metalloid-Based Nanomaterials for Plant Abiotic Stress Tolerance: An Overview of the Mechanisms. Plants 2022, 11, 316. [Google Scholar] [CrossRef]
- Horie, T.; Karahara, I.; Katsuhara, M. Salinity Tolerance Mechanisms in Glycophytes: An Overview with the Central Focus on Rice Plants. Rice 2012, 5, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, S.; Zhang, Q.; Liu, M.; Zhou, H.; Ma, C.; Wang, P. Regulation of Plant Responses to Salt Stress. Int. J. Mol. Sci. 2021, 22, 4609. [Google Scholar] [CrossRef] [PubMed]
- Kheloufi, A.; Mansouri, L.M. Anatomical Changes Induced by Salinity Stress in Root and Stem of Two Acacia Species (A. karroo and A. saligna). Agric. For. 2019, 65, 137–150. [Google Scholar] [CrossRef]
- Lungoci, C.; Motrescu, I.; Filipov, F.; Rimbu, C.M.; Jitareanu, C.D.; Ghitau, C.S.; Puiu, I.; Robu, T. Salinity Stress Influences the Main Biochemical Parameters of Nepeta Racemosa Lam. Plants 2023, 12, 583. [Google Scholar] [CrossRef]
- Azeem, M.; Pirjan, K.; Qasim, M.; Mahmood, A.; Javed, T.; Muhammad, H.; Yang, S.; Dong, R.; Ali, B.; Rahimi, M. Salinity Stress Improves Antioxidant Potential by Modulating Physio-Biochemical Responses in Moringa Oleifera Lam. Sci. Rep. 2023, 13, 2895. [Google Scholar] [CrossRef]
- Hussain, M.I.; Al-Dakheel, A.J.; Chaudhry, U.K.; Khan, M.I.; ALHaithloul, H.A.S.; Alghanem, S.M.; Alaklabi, A. Morpho-Physiological Response of Barley to Assess Genotypic Differences of Salinity Tolerance under Hyper Arid Climate. Agric. Water Manag. 2022, 272, 107832. [Google Scholar] [CrossRef]
- Arshad, K. Importance of Barley for Human Beings–AgriHunt—A Hunt for Agricultural Knowledge. Available online: https://agrihunt.com/articles/major-crops/importance-of-barley-for-human-beings/ (accessed on 8 January 2023).
- Langridge, P. Economic and Academic Importance of Barley; Springer: Berlin/Heidelberg, Germany, 2018; pp. 1–10. [Google Scholar] [CrossRef]
- Ganeshan, S.; Chibbar, R.N.; Dahleen, L.S.; Tranberg, J.; Lemaux, P.G. Barley. Compend. Transgenic Crop Plants 2008, 101–138. [Google Scholar] [CrossRef]
- Editors of Encyclopedia Barley-Cereal. Encyclopedia Britannica 2023. Available online: https://www.britannica.com/plant/barley-cereal (accessed on 6 March 2023).
- Palestinian Central Bureau of Statistics Cumulative Amounts of Rainfall from the Beginning of the Current Rainy Season 2022/2021 till 14 March 2022. Available online: https://pcbs.gov.ps/post.aspx?lang=en&ItemID=4199 (accessed on 6 March 2023).
- Al-Tardeh, S.; Sawidis, T.; Diannelidis, B.-E.; Delivopoulos, S. Water Content and Reserve Allocation Patterns within the Bulb of the Perennial Geophyte Red Squill (Liliaceae) in Relation to the Mediterranean Climate. Botany 2008, 86, 291–299. [Google Scholar] [CrossRef]
- Cappelletti, C. Water Content in Plants and Equations Used to Determine It. Ann. Bot. 1954, 24, 408–430. [Google Scholar]
- Clause, S.D.; Sasse, J.M. BRASSINOSTEROIDS: Essential Regulators of Plant Growth and Development. Annu. Rev. Plant Biol. 2003, 49, 427–451. [Google Scholar] [CrossRef] [Green Version]
- Oh, M.H.; Romanow, W.G.; Smith, R.C.; Zamski, E.; Sasse, J.; Clouse, S.D. Soybean BRU1 Encodes a Functional Xyloglucan Endotransglycosylase That Is Highly Expressed in Inner Epicotyl Tissues during Brassinosteroid-Promoted Elongation. Plant Cell Physiol. 1998, 39, 124–130. [Google Scholar] [CrossRef] [Green Version]
- Silva, N.C.Q.; de Souza, G.A.; Pimenta, T.M.; Brito, F.A.L.; Picoli, E.A.T.; Zsögön, A.; Ribeiro, D.M. Salt Stress Inhibits Germination of Stylosanthes Humilis Seeds through Abscisic Acid Accumulation and Associated Changes in Ethylene Production. Plant Physiol. Biochem. 2018, 130, 399–407. [Google Scholar] [CrossRef]
- Kumar, V.; Joshi, S.; Pant, N.C.; Sangwan, P.; Yadav, A.N.; Saxena, A.; Singh, D. Molecular Approaches for Combating Multiple Abiotic Stresses in Crops of Arid and Semi-Arid Region. In Energy, Environment, and Sustainability; Springer: Berlin/Heidelberg, Germany, 2019; pp. 149–170. [Google Scholar] [CrossRef]
- Wu, G.Q.; Jiao, Q.; Shui, Q.Z. Effect of Salinity on Seed Germination, Seedling Growth, and Inorganic and Organic Solutes Accumulation in Sunflower (Helianthus annuus L.). Plant Soil Environ. 2015, 61, 220–226. [Google Scholar] [CrossRef] [Green Version]
- Kaveh, H.; Nemati, H.; Farsi, M.; Vatandoost Jartoodeh, S. How Salinity Affect Germination and Emergence of Tomato Lines. J. Biol. Environ. Sci. 2011, 5, 159–163. [Google Scholar]
- Jadidi, O.; Etminan, A.; Azizi-Nezhad, R.; Ebrahimi, A.; Pour-Aboughadareh, A. Physiological and Molecular Responses of Barley Genotypes to Salinity Stress. Genes 2022, 13, 2040. [Google Scholar] [CrossRef]
- Hu, Y.; Schmidhalter, U. Drought and Salinity: A Comparison of Their Effects on Mineral Nutrition of Plants. J. Plant Nutr. Soil Sci. 2005, 168, 541–549. [Google Scholar] [CrossRef]
- Wu, H.; Jiang, H.; Liu, C.; Deng, Y. Growth, Pigment Composition, Chlorophyll Fluorescence and Antioxidant Defenses in the Red Alga Gracilaria Lemaneiformis (Gracilariales, Rhodophyta) under Light Stress. South Afr. J. Bot. 2015, 100, 27–32. [Google Scholar] [CrossRef]
- Vysotskaya, L.; Hedley, P.E.; Sharipova, G.; Veselov, D.; Kudoyarova, G.; Morris, J.; Jones, H.G. Effect of Salinity on Water Relations of Wild Barley Plants Differing in Salt Tolerance. AoB Plants 2010, 2010, plq006. [Google Scholar] [CrossRef] [PubMed]
- Maggio, A.; Hasegawa, P.M.; Bressan, R.A.; Federica Consiglio, M.; Joly, R.J. Review: Unravelling the Functional Relationship between Root Anatomy and Stress Tolerance. Funct. Plant Biol. 2001, 28, 999–1004. [Google Scholar] [CrossRef]
- Zhao, C.; Zhang, H.; Song, C.; Zhu, J.K.; Shabala, S. Mechanisms of Plant Responses and Adaptation to Soil Salinity. Innovation 2020, 1, 100017. [Google Scholar] [CrossRef] [PubMed]
- Grigore, M.N.; Toma, C. A Proposal for a New Halophytes Classification, Based on Integrative Anatomy Observations. Muz. Olteniei Craiova. Studii şi Comunicări, Ştiinţele Naturii 2010, 26, 45–50. [Google Scholar]
- Turkan, I. Plant Responses to Drought and Salinity Stress: Developments in a Post-Genomic Era; Academic Press: Cambridge, MA, USA, 2011; ISBN 0123876826. [Google Scholar]
- Weber, D.J.; Gul, B.; Khan, M.A. Halophytic Characteristics and Potential Uses of Allenrolfea Occidentalis. Prospect. Saline Agric. 2002, 37, 333–352. [Google Scholar] [CrossRef]
- Pan, J.; Peng, F.; Tedeschi, A.; Xue, X.; Wang, T.; Liao, J.; Zhang, W.; Huang, C. Do Halophytes and Glycophytes Differ in Their Interactions with Arbuscular Mycorrhizal Fungi under Salt Stress? A Meta-Analysis. Bot. Stud. 2020, 61, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Cheeseman, J.M. The Evolution of Halophytes, Glycophytes and Crops, and Its Implications for Food Security under Saline Conditions. New Phytol. 2015, 206, 557–570. [Google Scholar] [CrossRef]
- Al-Joulani, N. Soil Contamination in Hebron District Due to Stone Cutting Industry. Jordan J. Appl.Sci.—Nat. Sci. 2008, 10, 37–50. [Google Scholar]
- van Iersel, M. EC and PH: What Is It and Why It Matters. Available online: https://hortphys.uga.edu/hortphys/files/2020/03/EC-and-pH.pdf (accessed on 6 March 2023).
- Ule, O.; Ogbonna, D.N.; Okparanma, R.N.; Nrior, R.R. Myco-Enhanced Bioremediation in Open Field Crude Oil Contaminated Soil Using Mucor Racemosus and Aspergillus Niger. Curr. J. Appl. Sci. Technol. 2021, 40, 119–141. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Baladi | Rihan | ACSAD 176 | Icarda 1 | ACSAD 1417 | Improved Baladi | ACSAD 1744 | ACSAD 68 | ACSAD 1732 | Icarda 15 | |
|---|---|---|---|---|---|---|---|---|---|---|
| 0 mM NaCl | 3.0 ± 0.11 | 1.1 ±0.11 | 1.9 ± 0.02 | 1.5 ± 0.03 | 1.5 ± 0 | 0.2 ± 0.72 | 1.5 ± 0 | 1.8 ± 0 | 1.3 ± 0 | N/A |
| 50 mM NaCl | 2.3 ± 0.08 | 1.2 ± 0.05 | 2.3 ± 0.13 | 1.6 ± 0.09 | 1.8 ± 0.03 | 0.2 | 1.1 ± 0 | 4.1 ± 0.13 | 1.5 ± 0.12 | N/A |
| 85 mM NaCl | 3.4 ± 0.31 | 1.3 ± 0.07 | N/A | 1.6 ± 0.05 | 1.6 ± 0.03 | 0.3 | 1.6 ± 0.01 | 2.2 ± 0.13 | 1.8 ± 0 | N/A |
| 120 mM NaCl | 1.6 ± 0.49 | 2.6 ± 0.06 | N/A | 1.8 ± 0.08 | 1.8 ± 0.03 | 2.2 | 2.0 ± 0.06 | 4.4 | 1.2 ± 0.01 | N/A |
| 175 mM NaCl | N/A | N/A | N/A | 2.1 | 4.4 ± 0.83 | N/A | 3.9 ± 0.54 | N/A | 1.8 ± 0.05 | N/A |
| Soil Sample | Region in Hebron Governorate | Measured Temperature (°C) | Measured Conductivity of Soil/Water Mix (5:1) [µS/cm] | ECt Calculated for Soil [µS/cm] | EC25 Calculated for 25 °C [dS m−1] | TSS–Total Soluble Salts (ppm) |
|---|---|---|---|---|---|---|
| 1. Red soil, middle of stones, north-west Hebron | Bani Naim 31°30′57.1″ N 35°10′58.5″ E | 23.1 | 702 | 667 | 6.923 × 10−4 | 0.443 |
| 2. Bani Naim Ashour 1 | Bani Naim | 24 | 344 | 309 | 3.152 × 10−4 | 0.097 |
| 3. Bani Naim 1, center of quarry | Bani Naim | 23.6 | 417 | 382 | 3.927 × 10−4 | 0.150 |
| 4. Bani Naim Ashour 2 | Bani Naim | 22.8 | 493 | 458 | 4.782 × 10−4 | 0.219 |
| 5. Bani Naim sample 3 | Bani Naim | 23.8 | 550 | 515 | 5.274 × 10−4 | 0.272 |
| 6. Side of quarry, 2 cm depth | AlShyoukh | 24 | 348 | 313 | 3.193 × 10−4 | 0.099 |
| 7. Light soil, 15 cm depth | AlShyoukh | 25.8 | 219 | 184 | 1.81 × 10−4 | 0.0333 |
| 8. Red stones, middle of quarry | AlShyoukh | 26.4 | 546 | 511 | 4.967 × 10−4 | 0.254 |
| 9. Light soil under the cutting machine | AlShyoukh | 24.8 | 403 | 368 | 3.695 × 10−4 | 0.140 |
| 10. Abu Bassam, white stone | Seir 30°11′10.2″ N 35°19′05.7″ E | 26.2 | 356 | 330 | 3.221 × 10−4 | 0.106 |
| 11. Seir Jamal Nassar 220 cm depth | Seir | 23.4 | 914 | 879 | 9.071 × 10−4 | 0.797 |
| 12. White, soft road of quarry | Seir | 22.6 | 278 | 243 | 2.547 × 10−4 | 0.0619 |
| 13. Light red, Jamal Nassar, 15 cm above stone | Seir | 26.1 | 543 | 508 | 4.968 × 10−4 | 0.252 |
| 14. Red stone, Jamal Nassar, from surface of quarry | Seir | 25.9 | 839 | 804 | 7.895 × 10−4 | 0.635 |
| 15. Quarry road, Taffouh | Taffouh 31°32′20.7″ N 35°03′09.5″ E | 30 | 461 | 448.6 | 4.037 × 10−4 | 0.181 |
| 16. Quarry, Taffouh | Taffouh | 30.8 | 295 | 282.6 | 2.498 × 10−4 | 0.071 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Tardeh, S.M.; Alqam, H.N.; Kuhn, A.J.; Kuchendorf, C.M. In Vitro Assessment of Salinity Stress Impact on Early Growth in Ten Certified Palestinian Barley Cultivars (Hordeum vulgare L.) Potentially Suitable for Cultivation on Former Quarry Substrates. Water 2023, 15, 1065. https://doi.org/10.3390/w15061065
Al-Tardeh SM, Alqam HN, Kuhn AJ, Kuchendorf CM. In Vitro Assessment of Salinity Stress Impact on Early Growth in Ten Certified Palestinian Barley Cultivars (Hordeum vulgare L.) Potentially Suitable for Cultivation on Former Quarry Substrates. Water. 2023; 15(6):1065. https://doi.org/10.3390/w15061065
Chicago/Turabian StyleAl-Tardeh, Sharaf M., Hala N. Alqam, Arnd J. Kuhn, and Christina M. Kuchendorf. 2023. "In Vitro Assessment of Salinity Stress Impact on Early Growth in Ten Certified Palestinian Barley Cultivars (Hordeum vulgare L.) Potentially Suitable for Cultivation on Former Quarry Substrates" Water 15, no. 6: 1065. https://doi.org/10.3390/w15061065
APA StyleAl-Tardeh, S. M., Alqam, H. N., Kuhn, A. J., & Kuchendorf, C. M. (2023). In Vitro Assessment of Salinity Stress Impact on Early Growth in Ten Certified Palestinian Barley Cultivars (Hordeum vulgare L.) Potentially Suitable for Cultivation on Former Quarry Substrates. Water, 15(6), 1065. https://doi.org/10.3390/w15061065