Abstract
The biodiversity of stream fishes is critically threatened globally, and a major factor leading to the loss of biodiversity is anthropogenic land use in stream catchments, which act as stressors to stream fishes. Declines in the biodiversity of stream fish are often identified by a loss of species or fewer individuals comprising assemblages, but biological degradation can also occur with increases in non-native species and/or the spread of fish tolerant to anthropogenic land use, suggesting the importance of accounting for the distinctness of assemblages along with richness and diversity to best characterize the response of stream fish assemblages to anthropogenic landscape stressors. We summarized stream fish assemblages from 10,522 locations through multiple biodiversity indices and then quantified index responsiveness to natural landscape variables and anthropogenic land use in stream network catchments across five freshwater ecoregions in the temperate mesic portion of the United States. Indices included species richness, Shannon’s diversity, Pielou’s evenness, beta diversity, taxonomic diversity, and taxonomic distinctness. First, we tested for correlations among indices across freshwater ecoregions and found that while species richness and Shannon’s diversity were always highly correlated, taxonomic distinctiveness was not highly correlated with other biodiversity indices measured except taxonomic diversity. Then, we used multiple linear regression to predict biodiversity indices in each of the five freshwater ecoregions to identify significant landscape variables from natural landscape and anthropogenic land uses. Most indices were consistently predicted by catchment area, and many were predicted by elevation, except for beta diversity, emphasizing the importance of these natural landscape variables on biodiversity. In contrast, taxonomic distinctness was often predicted by the amount of urban land use in the catchment, but the direction of the relationship varied. The proportion of agriculture land use in the network catchment was a more consistent predictor of species richness, beta diversity, and Shannon’s diversity. Our analyses show that taxonomic distinctness in freshwater fishes characterize a unique element of biodiversity in relationships with anthropogenic land uses in a streams network catchment. Taxonomic distinctness may also be an effective metric for the bioassessment of stream fishes along with richness and diversity indices to help preserve biodiversity in regard to current and future anthropogenic land uses.
1. Introduction
Habitat degradation and loss caused by anthropogenic landscape stressors are global threats to the biological diversity of stream fishes [1,2,3,4,5,6]. Reduced stream fish biodiversity may be characterized by lost species or fewer individuals but increases in non-native species and the spread of fishes tolerant of anthropogenic stressors may also be indicators of biological degradation [7,8,9,10,11,12]. For example, anthropogenic stressors can encourage the extirpations of endemic fishes intolerant to resulting habitat alterations, while other species more tolerant of stressors may thrive under modified habitat conditions or due to the suppression of predators or competitors that do not persist in altered habitats [2,9,10]. Such changes may be reflected by increased faunal similarity across regions subjected to anthropogenic stressors [11,12,13]. Because of this, the measures of species richness and diversity alone may not be fully effective at accounting for changes in biodiversity resulting from anthropogenic stressors [14].
Commonly used indices representing biological diversity in community assemblages based solely on the numbers of species and the abundances of individuals have limitations, but, when used in concert with phylogenic measures characterizing specific types of faunal changes in diversity or distinctness, can inform conservation strategies to help preserve phylogenic diversity [15]. In many studies of stream habitats, species richness has been shown to decline with greater types and intensities of anthropogenic stressors in stream catchments [16,17]. However, in some systems, anthropogenic stressors leading to increases in stream temperature and/or increases in nutrients and greater system productivity also contribute to increased richness [18,19]. Species richness can also vary widely across regions due to biogeographic history, stream size, and other natural landscape-scale variables that may influence the natural potential of stream habitats to support various species [12,20,21,22,23]. Species richness summaries for individual sites also do not account for differences in rare species versus common species, giving as much weight to common, abundant species as to rare species with few individuals. Increasingly, species richness is suggested to be used in concert with other measures of biodiversity or community structure that account for the replacement of the proportional abundances of individual species, and this may provide insight into the influences of environmental variation on communities [15,24,25,26,27]. For example, measures of evenness account for abundances, with similarity in measures for two assemblages indicating more equal numbers of individuals per species in the assemblages. In contrast, an assemblage that includes few species with high abundances compared to multiple others with low abundances has low evenness [15]. As a second example, Shannon’s diversity index weights all species proportionately to their abundances; high values for Shannon’s diversity indicate more equitable distributions of individuals present in an assemblage, while low values may indicate assemblage dominance by a few species [15]. Together, species richness, Pielou’s evenness, and Shannon’s diversity can effectively indicate variation in diversity driven by the loss of species or individuals, yet not necessarily the gains of non-natives or species tolerant of anthropogenic stressors [28]. Beta-diversity is a commonly used index representing the variation in species composition among sample sites [25,27]. Local area contributions of beta-diversity can be driven by natural landscape variables and influenced by anthropogenic land use in the network catchment. A better understanding of the influence of natural and anthropogenic variables affecting beta-diversity is warranted to inform relevant conservation strategies for protecting biodiversity across large region.
Taxonomic distinctness measures can indicate changes in biodiversity that do not necessarily result in changes to species richness, evenness, or Shannon’s diversity [14,29]. Taxonomic diversity and taxonomic distinctness indices depend on the composition of site-specific assemblages found throughout an entire study region. They include a hierarchical measure of relatedness based on taxonomy between fish species within a single sample to the pool of species found throughout the region [30,31]. In minimally disturbed streams, fish assemblages may include more species that occupy a wider breadth in taxonomic diversity and distinctness [32,33]. These can include endemic habitat specialists, species with specialized feeding preferences, or species with unique life history traits. Under anthropogenic stressors, a loss of endemic specialist species and/or a spread of tolerant species may result in a smaller number of taxonomic groups and a decline in fish assemblage distinctness. This loss of distinctness can happen through the loss of endemic specialist species, unique genera distinctness, or entire intolerant family groups. The loss of assemblage distinctness can also occur through the expansion of similarity at the genera or family group level via emigration. Because of this, taxonomic distinctness measures can indicate unique taxonomic change in assemblages resulting from anthropogenic land uses and urbanization.
Given the strength of multiple indices at showing different features of assemblages, the comparison of how multiple biodiversity indices may be associated with natural landscape variables and anthropogenic landscape stressors could improve the understanding of how well biodiversity indices reflect habitat degradation and loss resulting from stressors. Further, such a study conducted over large regions that accounts for anthropogenic land use as a source of stress on stream conditions and evaluates the response of multiple indices could yield new insights into the broad patterns of biodiversity associated with large-scale influences, such as anthropogenic land uses. To help meet these needs, the goal of this study is to characterize patterns in the multiple biodiversity indices of stream fish assemblages across five biogeographic regions of the temperate mesic portion of the United States and assess the influences of anthropogenic landscape variables on the indices while also accounting for the influences of natural factors known to be important to stream organisms and their habitats. Indices to be evaluated include species richness, Shannon’s diversity, Pielou’s evenness, beta diversity, taxonomic diversity, and taxonomic distinctness. We have three objectives. First, we characterize major patterns in multiple fish biodiversity indices across five study freshwater ecoregions, including interrelationships among the various indices within and across freshwater ecoregions. Second, we identify dominant natural landscape variables and anthropogenic stressors important in explaining variation in each of the indices. Finally, we quantify how the influences of natural landscape variables and anthropogenic land uses on biodiversity indices vary across five freshwater ecoregions. Addressing these objectives offers new insights into the influences of natural landscape variables and anthropogenic land uses on stream fish assemblages across large regions and provides opportunities to inform conservation efforts to protect stream fishes and to preserve biodiversity from current and future stressors.
2. Materials and Methods
2.1. Study Area
This study was conducted within five freshwater ecoregions in the temperate mesic portion of the United States, covering an area greater than 2,000,000 km2 and using data characterizing stream fish assemblages from 10,522 locations across all ecoregions. Freshwater ecoregions were delineated by the World Wildlife Fund, who have boundaries that follow large river basins with similar biogeographic history in fish faunas [34,35,36,37]. The similarity of fish fauna biogeographic history within freshwater ecoregions defines ecologically relevant units for the grouping of fishes based on evolutionary patterns across large landscapes [34,35,36,38,39]. Freshwater ecoregions were used in this study to help control for natural variation and to evaluate different biodiversity indices in showing index responsiveness to anthropogenic landscape variables, which also vary in type and intensity across study freshwater ecoregions. Ecoregions included the Appalachian Piedmont, Chesapeake Bay, Laurentian Great Lakes, Middle Missouri, and the Upper Mississippi [37], see Figure 1.
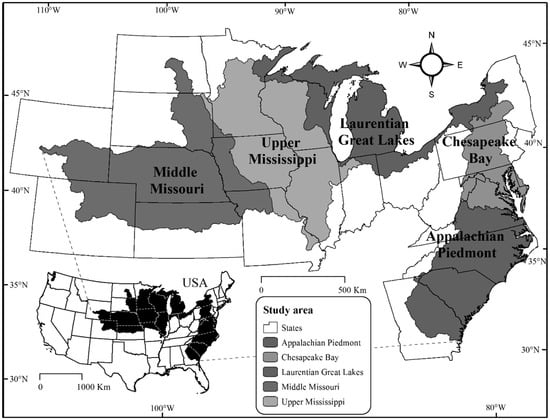
Figure 1.
Five freshwater ecoregions of the United States that comprised the study area [37].
Natural landscape variables within the study area are highly diverse, with contrasting surficial geology from freshwater ecoregions including previously glaciated landscapes in the Chesapeake Bay and Laurentian Great Lakes as well as areas that have not been glaciated, including the coastal plains and highlands of the Appalachian Piedmont freshwater ecoregion. The precipitation gradient increases from west to east, and the air temperature gradient increases from north to south. In the eastern and central portion of the study area, the Middle Missouri and Upper Mississippi freshwater ecoregions include landscapes with intensive agriculture, while high percentages of grasslands also occur within the western Middle Missouri freshwater ecoregion. High percentages of forested areas are located within the Laurentian Great Lakes, and the Appalachian Piedmont and the Chesapeake Bay freshwater ecoregions include areas of high urban land use. Hereafter, freshwater ecoregions will be referred to as ecoregions.
2.2. Stream Layer
The 1:100,000-scale National Hydrography Dataset Plus Version 1 (NHD) national streams layer was used for the geographic representation of stream reaches and their associated local catchments [40]. We defined a stream reach as described in Wang et al., 2011 [41]. Catchments summarizing information over two spatial extents were used in this analysis based on the NHD. Local catchments include all land that drains directly into an individual stream reach without being transported via other fluvial pathways represented in the NHD, and network catchments encompass all land upstream of and draining into a given reach via fluvial pathways and including the local catchment.
2.3. Natural Landscape Variables
Six natural landscape variables were evaluated in this study in different spatial extents (Table 1). The average elevation of local catchments was developed from the U.S. Geological Survey (USGS) National Elevation Data (NED) [42]. Mean annual air temperature and mean annual precipitation were summarized from 1981 to 2010 by the PRISM Climate Group [43,44]. Air temperature was summarized in local catchments for analysis, and precipitation, also summarized in local catchments, was aggregated throughout entire upstream network catchments for analysis, following [45]. Surficial lithology was available continuously across the conterminous United States [46,47], and we summarized categories into a single variable. “Fine surficial lithology” grouped major categories of lithology based on relatively low hydraulic conductivities, and this variable, attributed to local catchments and aggregated within network catchments, is an indicator of streams with more variable vs. more stable flow regimes. The baseflow index (modeled by USGS) [48], is a measure characterizing the percentage of groundwater contribution to stream flows, and we attributed this factor to local catchments and aggregated it within network catchments for analysis. Last, network catchment areas for all stream reaches in the study ecoregions were summarized based on the aggregated areas of local catchments provided in the NHD.

Table 1.
Landscape variable descriptions and descriptive statistics of landscape variables for the five freshwater ecoregions (i.e., mean, minimum, maximum, and the 10th and 90th percentile values), with n = number of sample sites by ecoregion.
2.4. Anthropogenic Land Uses
Four anthropogenic land use variables were evaluated across the five study ecoregions. Agricultural land use, urban land use, and dam density in network catchments have been shown to influence stream fish assemblages throughout the study region [49,50,51,52,53]. Additionally, stream road crossings may influence fishes, since many crossings can increase stream habitat fragmentation throughout a catchment and can be reflective of barriers to fish movement due to the structures associated with crossings, such as culverts [54,55]. Urban and agriculture summaries were developed from the 2001 National Land Cover Dataset (NLCD) [56,57]. Urban land use included low, medium, and high intensity developed land along with open space developed land cover classes. Agricultural land use included pasture/hay and cultivated crop land cover classes. The density of dams was developed from the National Anthropogenic Barrier Dataset (NABD) [58], and the density of stream-road crossings was developed from the U.S. Census Bureau TIGER/line files data [59]. All anthropogenic land use variables were summarized based on local catchments and then aggregated to network catchments following methods described in [45].
2.5. Fish Data
Data characterizing stream fish assemblages were gathered and referenced to stream reaches in the NHD. Assemblages were sampled with electrofishing by state and federal programs using methods determined to be comparable for whole fish assemblages for creeks, small rivers, and medium rivers ≤10,000 km2 in drainage areas between years 1990 to 2010 [60,61]. Sampling years included in analysis were chosen to appropriately match natural landscape variables and anthropogenic land use information included in study. Fish used in analysis were identified to species, and all scientific naming was standardized to the Integrated Taxonomic Information System [62].
2.6. Biodiversity Indices
Six measures of fish assemblage biodiversity were calculated for this study. Three of these indices were derived from site-specific data characterizing the number of species and their abundances. These indices included species richness (S), species diversity based on Shannon’s diversity index (H’), and Pielou’s evenness (J’) [63,64]. Shannon’s index and Pielou’s evenness indices were calculated using the function “diversity” from the package “vegan” in R [65,66]. The index for total beta diversity (herein beta diversity) was calculated for the regional species pool across all sites in the study region using percentage difference dissimilarity index from fish abundance data, following [26,27]. The local contribution to beta diversity was calculated for each study site [25,27]. Total beta diversity metrics were calculated using the function “beta.div.comp” and computed for the local contribution of beta diversity for each site using the function “LCBD.comp” from the package “adespatial” in R [25,26,27,66,67]. The last two indices were calculated describing taxonomic diversity (Δ) and taxonomic distinctness (Δ*) [31]. These indices were derived from fish abundance data and were based on relationships in the composition of stream fish assemblages at individual study sites evaluated against taxonomic relatedness from the regional species pool across all sites for the entire study area [30,31]. This contrasts with species richness, Shannon’s diversity, and Pielou’s evenness, which are calculated from the assemblages of a single study site. Taxonomic relatedness is derived in part from the cladistics classification of all organisms that account for taxonomic levels at which species are related (species, genus, family, order, and class). A numeric weighting of the branch lengths of the hierarchical cladistics classification tree accounts for interrelationships among all species in the regional species pool—see Table S1. For a hypothetical cladogram, see [30], which highlights differences in relatedness weighting among individuals. Five taxonomic levels were used, namely species, genus, family, order, and class, to calculate the distinctness weight from the master fish species list for the entire study area—see Table S1. The distinctness weight was calculated using the function “taxa2dist,” and taxonomic diversity and taxonomic distinctness indices were calculated using the function “taxondive” from the package “vegan” in R [65,66]. Before analysis, variables and metrics were transformed or standardized to better meet the assumptions of linearity, homoscedasticity, and comparability across biodiversity indices.
2.7. Data Analysis
2.7.1. Comparing Landscape Features across Regions
Descriptive statistics were calculated using the “stats” package in R to investigate ranges in natural landscape variables and anthropogenic land uses across study ecoregions, including means, minimums, maximums, and 10th and 90th percentiles [66]. These statistics describe the relative differences in the dispersions and potential thresholds of natural landscape variables and anthropogenic land uses across the study ecoregions.
2.7.2. Pearson’s Correlation among Fish Biodiversity Indices
Pearson’s correlation analysis was used to compare co-variability among fish biodiversity indices and to characterize how these interrelationships varied across ecoregions. Species richness was square root transformed, and all other indices were standardized by calculating z-scores using the function “scale” from the package “base” in R [66]. Correlations were judged to be high when r values were ≥0.65, moderate at 0.35 to <0.65, and low when values were <0.35.
2.7.3. Regression Analysis
Multiple linear regression models were developed to identify the most important natural landscape variables and anthropogenic land uses predicting each of the six fish biodiversity indices and to evaluate how models varied across the five study ecoregions. The best linear regression models were developed utilizing an exhaustive search with a branch-and-bound algorithm for the best subset of five or fewer natural landscape variables and/or anthropogenic land use variables (from a total of 10 possible predictors, Table 1) predicting each of six fish biodiversity indices in all five study ecoregions [68,69,70]. The best models were selected using the Bayesian information criterion (BIC). A standardized regression coefficient (β) value was calculated to evaluate the importance of variables selected for inclusion in the models predicting biodiversity indices. This dimensionless coefficient (which has a value between 0 and 1) is used to identify important variables and any irrational variable in a regression function. A value near 1 indicates an important predictor to the dependent variable, and a value near 0 indicates an unimportant predictor [71,72]. To identify collinearity among explanatory variables, a variance inflation factor (VIF) was calculated. Multiple linear regression models were calculated using the function “regsubsets” from the package “leaps” in R [66,70]. The standardized regression coefficient (β) was calculated using the function “lm.beta” from the package “QuantPsyc” in R [66,73].
3. Results
3.1. Study Area
Natural landscape variables across the study ecoregions are highly diverse (Table 1). The mean network catchment area of study sites ranged from 155 km2 in the Appalachian Piedmont ecoregion to 764 km2 in the Middle Missouri ecoregion. Fine lithology in the network catchment was lowest in the Upper Mississippi (mean of 9%) and highest in the Laurentian Great Lakes (mean of 42%). Mean local elevation was lowest at study sites in the Appalachian Piedmont at 157 m and ranged from 20 m to 1166 m across all study sites (Table 1). Mean local elevation was highest at study sites in the Middle Missouri at 591 m. Climate varied across sites, with the lowest mean annual air temperatures in the Upper Mississippi (8 °C) and highest mean annual air temperature and precipitation in the Appalachian Piedmont (15 °C, 1215 mm, respectively Table 1). The lowest mean annual precipitation was in the Middle Missouri ecoregion at 709 mm.
Anthropogenic land use also varied widely across ecoregions. The mean percentage of urban land use in the network catchments of study sites was highest in the Chesapeake Bay at 32% and lowest at study sites in the Middle Missouri at 6% (Table 1). The mean percentage of agriculture land use in the network catchment also varied across ecoregions; it was highest in the Upper Mississippi at 60%, with 10th and 90th percentiles of 16% and 90%, respectively. The mean network catchment percent agriculture was lowest in Appalachian Piedmont with a value of 21%, and with 10th and 90th percentiles of 4% and 41%, respectively. The density of stream-road crossings in the network catchments of the study sites was highest in the Chesapeake Bay at 54/100 km2 and was lowest in the network catchment for study sites in Middle Missouri at 36/100 km2. The density of dams in the network catchment of study sites was highest in the Appalachian Piedmont at 3.91/100 km2 and lowest in the network catchment of study sites in the Upper Mississippi at 0.71/100 km2.
3.2. Regional Patterns in Biodiversity Indices
The study area supported 312 species of fish, comprising 98 genera, 34 families, 21 orders, and 2 classes; see Table S1. Patterns in biodiversity indices varied across ecoregions (Table 2). Species richness across all sites ranged from a low of one in every ecoregion to a maximum of 48 in the Upper Mississippi. While mean species richness was highest in the Appalachian Piedmont (15 species), mean species richness ranged to a low of 10 in both the Chesapeake Bay and Middle Missouri. Differences across ecoregions were also shown when considering the 10th percentile value for species richness, which highlight regions with larger fish species pools that have, on average, relatively more species present even at lower quality stream sites. In the Chesapeake Bay ecoregion, the 10th percentile value for species richness was two, while in the Appalachian Piedmont, it was eight, indicating a substantial number of sites with low species richness in the Chesapeake Bay ecoregion (Table 2). Shannon’s diversity and Pielou’s evenness values were also variable. Similar to species richness, the mean Shannon’s diversity was highest in the Appalachian Piedmont (1.89, Table 2), while the mean evenness was highest in the Chesapeake Bay (0.61, Table 2). This likely reflects the fact that while sites in the Chesapeake Bay support fewer species on average compared to other ecoregions, the species comprising assemblages are more even in numbers. Mean diversity and mean evenness across sites were lowest in the Middle Missouri. Finally, the highest value for Shannon’s diversity was found in the Upper Mississippi at 3.11 (Table 2). Patterns in the local contributions of beta diversity were similar across ecoregions. Local contributions to beta diversity at the study sites were lowest in the Laurentian Great Lakes (0.0000759, Table 2) and highest in the Laurentian Great Lakes and Upper Mississippi (0.0001091, Table 2). Beta diversity across sites in the Appalachian Piedmont had a higher 10th and 90th percentile value of 0.000097 and 0.0001056, respectively, which is reflective of higher mean biodiversity across all study sites in the Appalachian Piedmont than the average for other study sites across other study ecoregions (Table 2). Patterns in taxonomic diversity varied compared to patterns expressed by species richness and Shannon’s diversity. Mean taxonomic diversity and distinctness were highest in the Appalachian Piedmont (54.05 and 69.98, respectively, Table 2), following the highest values for species richness and Shannon’s diversity in the Appalachian Piedmont ecoregion. In contrast, the second highest mean taxonomic diversity and distinctness value were in the Middle Missouri ecoregion (41.89 and 66.40, respectively). While mean species richness and Shannon’s diversity were low here, this result emphasizes that even in ecoregions with lower species richness or Shannon’s diversity, sites may still support assemblages with a high degree of taxonomic diversity and taxonomic distinctness.
Table 2.
Descriptive statistics (i.e., mean, range, and 10th and 90th percentiles) of fish biodiversity indices in five freshwater ecoregions.
3.3. Regional Patterns in the Correlation of Biodiversity Indices
Patterns in correlation among biological indices were similar across study ecoregions (Table 3). Species richness and Shannon’s diversity were highly correlated in each of the five study ecoregions and ranged in strength from a high of r = 0.91 in the Chesapeake Bay to a low of r = 0.74 in the Middle Missouri. Shannon’s diversity also had moderate to high correlation with taxonomic diversity across all ecoregions; the highest value was r = 0.75 in the Upper Mississippi and the lowest value was r = 0.54 in the Appalachian Piedmont (Table 3). Taxonomic diversity was highly correlated with taxonomic distinctness across four ecoregions and moderately correlated in the Middle Missouri (highest value of r = 0.79 in the Chesapeake Bay to a lowest value of r = 0.62 in the Middle Missouri ecoregion, Table 3). Interestingly, while taxonomic diversity was moderately correlated with species richness and moderately to highly correlated with Shannon’s diversity, taxonomic distinctness was never highly correlated with these factors (highest value of r = 0.44 in the Upper Mississippi), emphasizing that this metric is likely capturing a different biodiversity component than the other evaluated indices (Table 3). Beta diversity was moderately negatively correlated with species richness and Shannon’s diversity in three of the five regions. Beta diversity was moderately positive correlated with taxonomic distinctness in the Chesapeake Bay and Laurentian Great Lakes ecoregions (range of 0.48 to −0.54, Table 3). Evenness was highly negatively correlated with species richness in the Chesapeake Bay and moderately negatively correlated with species richness in the four other study ecoregions and Shannon’s diversity in the Chesapeake Bay ecoregion (range of 0.28 to −0.65, Table 3).
Table 3.
Pearson’s correlation (r) between the six biodiversity indices for the five study freshwater ecoregions.
3.4. Predicting Biodiversity Indices from Landscape Variables with Linear Regression Models
All biodiversity indices were significantly predicted by at least two landscape variables in linear regression models. Adjusted R2 ranged from 0.52 for species richness in the Upper Mississippi ecoregions to 0.04 for Pielou’s evenness in the Appalachian Piedmont (Table 4). All natural landscape variables and anthropogenic land use variables selected as predictor for inclusion into models were significant (p ≤ 0.01), and their variance inflation factors (VIF) were low (i.e., maximum VIF = 6.4) for all the best models across all five study ecoregions. The low VIF values equal to or below 6.4 for all selected predictor variables indicated that there was low collinearity between variables selected in the best models predicting biodiversity indices [74]. Biodiversity indices were variably predicted across ecoregions described by the variance in adjusted R2 values from linear regression models (Table 4). Species richness was the best predicted biodiversity index in four out of five ecoregions with beta diversity being better predicted in the Appalachian Piedmont ecoregion (Table 4). Taxonomic diversity and distinctness biodiversity indices were the best predicted in the Chesapeake Bay ecoregion with adjusted R2 = 0.37 (Table 4). The best predicted index on average across all study ecoregions was species richness with a mean adjusted R2 = 0.37 (Figure 2). The second best predicted index on average across the study ecoregions was a taxonomic diversity with a mean adjusted R2 = 0.27 (Figure 2), while the least predicted biodiversity index on average was Pielou’s evenness across study ecoregions with a mean adjusted R2 = 0.14 (Figure 2).
Table 4.
Results of multiple linear regression models for the relationships of the biodiversity indices to natural landscape variables and anthropogenic land use variables, with degrees of freedom (d. f.), adjusted R2 (Adj R2), and standardized regression coefficient (β) describing the variable strength in the model and direction of influence on biodiversity indices in relation to variables. Predictor variables are as follows: Area = area; Elev = elevation; Fine = percent fine lithology; Baseflow = percent groundwater contribution to baseflow; AirTemp = mean annual air temperature; Precip = mean annual precipitation; Urban = percent urban land use; Ag = percent agri-cultural land use; Rd = density of stream-road crossings; Dam = density of dams; and c = local catchment summary and nc = network catchment summary. All variables selected for inclusion in best models as predictors for biodiversity indices were significant at p-value ≤ 0.01.
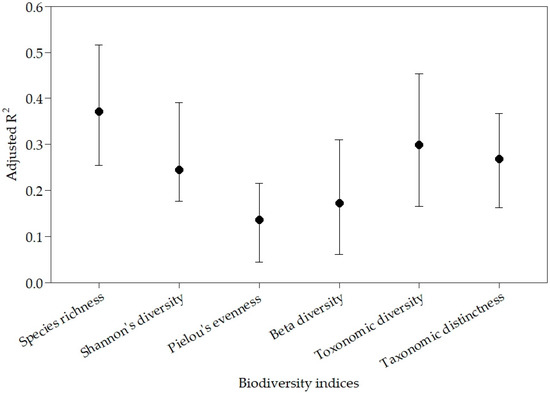
Figure 2.
Range in adjusted R2 values in biodiversity indices across five study freshwater ecoregions from the multiple linear regressions. The dots are mean values with the whiskers representing maximum and minimum values.
The important natural landscape variables and anthropogenic land use predictors of biodiversity indices were variable across study ecoregions and by biodiversity indices (Table 4). The most important predictor of biodiversity indices was network catchment area. The catchment area was the strongest or second strongest predictor in 24 of the 30 best models predicting all biodiversity indices across the five study ecoregions (Table 4). Catchment area was a positive predictor of species richness, Shannon’s diversity, taxonomic diversity, taxonomic distinctness, and a negative predictor of Pielou’s evenness for all ecoregions, but it was not selected in the best models as a predictor of beta diversity in the Chesapeake Bay or Laurentian Great Lakes and had a negative prediction of beta diversity in the Upper Mississippi ecoregion (Table 4). Local catchment elevation was also important for predicting beta diversity, taxonomic diversity, and taxonomic distinctness indices in four of the five ecoregions (Table 4). Local catchment elevation was a negative predictor of biodiversity indices, except for species richness in the Laurentian Great Lakes and Shannon’s diversity in the Upper Mississippi. Catchment agriculture was an important anthropogenic land use predicting species richness, Shannon’s diversity, and beta diversity in three of the five ecoregions, and it also significantly predicted species richness in Middle Missouri, taxonomic diversity in the Chesapeake Bay and Upper Mississippi, and taxonomic distinctness in Laurentian Great Lakes (Table 4). The positive or negative influence of catchment agriculture to predict biodiversity indices varied by ecoregion and index. Catchment urban land use was a consistent predictor of taxonomic distinctness in four of the five ecoregions, but the direction of that influence varied by ecoregions (Table 4). Both local air temperature and catchment precipitation were important predictors of indices across all ecoregions, yet no consistent patterns could be discerned in their ability to positively or negatively predict specific indices or in regional trends.
3.5. Regional Differences in Predicting Biodiversity Indices from Landscape Variables
Regression models showed regional differences in the influences of network catchment urban and agricultural land use on biodiversity indices. Network catchment urban was a good predictor for taxonomic distinctness in four of the ecoregions except the Chesapeake Bay ecoregion, and the direction of the influence varied across ecoregions. Network catchment urban had a positive influence on taxonomic distinctness in the Appalachian Piedmont ecoregion and a negative influence on taxonomic distinctness in the Laurentian Great Lakes, Middle Missouri, and Upper Mississippi ecoregions. Although they had comparatively less predictive capacity than other metrics, the density of stream road crossings and dams in the network catchment were significant predictors of species richness and Shannon’s diversity in the Chesapeake Bay ecoregions. The density of stream-road crossings in the network catchment was a significant positive predictor of species richness and Shannon’s diversity in the Chesapeake Bay and a positive predictor in the Middle Missouri. The density of stream-road crossings was a significant negative predictor beta diversity in Laurentian Great Lakes and Upper Mississippi ecoregions. The density of dams in the network catchment was also a significant positive predictor of species richness and Shannon’s diversity in the Chesapeake Bay and taxonomic diversity in the Laurentian Great Lakes ecoregions (Table 4). Biodiversity indices were significant predictors, but weakly influenced by network catchment fine lithology, baseflow index, network catchment precipitation, and local catchment air temperature with regional patterns and the direction of influence variable by specific index and across ecoregions.
4. Discussion
We characterized patterns in six biodiversity indices for stream fish assemblages across five freshwater ecoregions of the eastern United States. Our results showed that interrelationships among these biodiversity indices were consistent in their correlations across ecoregions with variability in their values. Species richness and Shannon’s diversity, for example, were always highly correlated in each of the five study ecoregions, yet taxonomic distinctiveness was never highly correlated with either of these two variables, emphasizing the independence of distinctness from richness and Shannon’s diversity. Then, we assessed the major influences of both natural landscape variables and anthropogenic stressors on biodiversity indices. The results of multiple linear regression models suggest that the percentage of agriculture and urban land use in the network catchments is the most important anthropogenic landscape predictor of biodiversity and network catchment area, and local catchment elevation explained the most variance in most biodiversity indices across study ecoregions. Network catchment agriculture had a fairly consistent positive association with species richness and Shannon’s diversity. In contrast, network catchment urban land use most consistently predicted taxonomic distinctness. The differences characterized in the response of taxonomic distinctness to anthropogenic land use in stream network catchments across ecoregions may be contributing to differing proportions of assemblage membership from specialist versus generalist species or to the extensiveness of differing regional levels of anthropogenic land use. Together, the results of our study suggest that multiple measures of fish biodiversity could be used to better characterize the response of stream fish assemblages more thoroughly to landscape influences, not only because they capture the unique attributes of the fish assemblage but also because indices are being differently influenced by various land uses.
4.1. Correlation among Biodiversity Indices
The measures of biodiversity were variably intercorrelated but showed similar trends in correlations across ecoregions. Species richness and Shannon’s diversity indices were always highly correlated across all study ecoregions, as were Shannon’s diversity and taxonomic diversity. Taxonomic diversity and taxonomic distinctness were moderately to highly correlated in each of the five study ecoregions (Table 3). In contrast, taxonomic distinctness was never strongly correlated with species richness, Shannon’s diversity, Pielou’s evenness or beta diversity. Similar relationships between biodiversity indices were seen in macroinvertebrate communities from stream sites in western and central Finland, and they found that taxonomic distinctness of macroinvertebrate assemblages was not correlated with Shannon’s diversity or Pielou’s evenness [75]. A similar study found that fish species richness and taxonomic distinctness were moderately to weakly correlated for lake fish assemblages in northeastern Finland [76]. The results of these studies, along with ours, emphasize that these biodiversity indices represent the unique attributes of stream fish assemblages, and that commonly used metrics such as species richness, Shannon’s diversity, and Pielou’s evenness may not be fully effective for capturing differences in the regional distinctiveness of freshwater assemblages, including differences resulting from anthropogenic landscape variables.
4.2. Natural Landscape Variables Predicting Biodiversity Indices
Of the major natural and anthropogenic gradients analyzed, biodiversity indices were most strongly influenced by network catchment area—they were significant predictors of all biodiversity indices for all study ecoregions, except beta diversity in the Middle Missouri and Laurentian Great Lakes. This result follows from species–area relationships for fish fauna [23,77,78,79]. With increasing stream size, there was an increase in stream productivity, habitat heterogeneity, and habitat area, promoting fish assemblage diversity [23,80,81,82]. Local elevation was also an important natural landscape predictor of beta diversity, taxonomic diversity, and taxonomic distinctness indices, with negative effects in four of the five ecoregions (Table 4). This may be related to stream size and due to an increase in assemblage taxonomic complexity at lower elevations where increased stability in flow, temperature regimes and spatial connectivity in fluvial systems, and the complexity of habitats contribute to increased taxonomic diversity with the additions of unique genera and family groups [83,84]. Together, these findings underscore the role of natural landscape variables on biodiversity indices and the sensitivity of indices to catchment area and elevation as a control of habitat heterogeneity and influence on biodiversity in the fish assemblages of fluvial ecosystems.
Network catchment fine lithology was also an important predictor of biodiversity indices for the Laurentian Great Lakes ecoregion, positively influencing species richness and Shannon’s diversity, and negatively influencing Pielou’s evenness and beta diversity in the ecoregions. The percentage of fine lithology in network catchments has been shown to be important for fish assemblage structure due to its influence on thermal and hydrologic conditions within streams [85,86]. Higher proportions of fine lithology in a catchment often suggests reduced groundwater influence on streams, with an increase in contribution to discharge from overland flow, warmer stream temperatures, and an increased variability in stream flow. This can lead to greater stream productivity from nutrient additions from overland flow and warmer stream temperatures.
Both mean annual air temperature in the local catchment and mean annual precipitation in the network catchment were important predictors of biodiversity indices across all study ecoregions, yet no consistent patterns could be interpreted in the ability of these variables to predict specific indices or in regional trends. For example, species richness and Shannon’s diversity were positively associated with air temperature in the Chesapeake Bay and Laurentian Great Lakes ecoregions but were negatively associated with air temperature in the Appalachian Piedmont. In more northern regions, air temperature may play a more important role influencing in-stream temperatures and in part positively affecting species richness and Shannon’s diversity than in more southern regions with warmer climates and air temperature. Additionally, network catchment precipitation had a positive influence on species richness, Shannon’s diversity and both taxonomic indices, with a negative influence on beta diversity in the Upper Mississippi, yet network catchment precipitation was not a good predictor for any biodiversity indices in the Appalachian Piedmont ecoregion, except a positive influence on beta diversity. Precipitation may have weak influences on the biodiversity of stream fish assemblages in areas such as the Appalachian Piedmont ecoregion with higher mean annual precipitation rates but may have a stronger influence in more arid ecoregions, such as the Upper Mississippi, among our study ecoregions. Despite the lack of consistent trends across indices or ecoregions, the variable associations between air temperature and precipitation on biodiversity indices do suggest their importance in controlling stream fish assemblages. Further, the influence of air temperature and precipitation underscores the fact that stream fish biodiversity has the potential to be influenced by climate change [1,87,88] and that a better understanding of the regional influences of climate on stream fish assemblages is warranted.
4.3. Anthropogenic Land Uses Predicting Biodiversity Indices
The percent of network catchment agriculture and urban land use were important predictors of biodiversity indices across all study ecoregions; however, the association of these land uses with indices were variable. Network catchment agriculture was shown to generally be positively associated with fish species richness and Shannon’s diversity across study ecoregions. An exception was a negative association with species richness in the Middle Missouri Ecoregions. It also had negative associations with beta diversity in Chesapeake Bay, Middle Missouri, and Upper Mississippi. A study of stream fishes from 47 sites in southeastern Wisconsin, United States, also found that species richness and diversity were positively associated with network catchment agriculture [16]. Their study sites also had a wide range in anthropogenic land use in their network catchments, including some with high levels of urban and agricultural lands. They summarized that, if impervious surfaces associated with urban land uses were low in network catchments, sites with high levels of agriculture land use still had the potential for high species richness and diversity. However, a different study showed that with high levels of agriculture (>50% in network catchments), fish index of biotic integrity (IBI) scores from streams decreased across study sites that had moderate levels of agriculture land use in their network catchments [89]. This suggests that agriculture land use may have negative effects on stream fish assemblages that are not necessarily reflected in changes in species richness and diversity. Such changes could include losses of intolerant fishes and co-occurring increases in tolerant fish or nonnatives. It also emphasizes the importance of using multiple metrics to best characterize changes in biodiversity that may result in streams from anthropogenic land uses. Our findings, along with those of the studies referenced above, could also result from differences in types of agriculture occurring in the landscape and/or specific mechanisms by which agriculture could influence streams; therefore, investigations that accounted for specific changes in stream habitats that occurred through agriculture (e.g., increased nutrient loadings and/or changes in sediment, thermal, or flow regimes) would likely yield more specific insights into how fish respond to agriculture land use in the network catchment.
Network catchment urban land use was a consistent predictor of taxonomic distinctness in four of the five ecoregions, although its influence on taxonomic distinctness was positive in the Appalachian Piedmont and negative in the Laurentian Great Lakes, Middle Missouri, and Upper Mississippi ecoregions These varying influences on taxonomic. distinctness may be due in part to regional influences related to the specific proportion or intensity of urban land use in the ecoregions or the interaction between natural landscape factors, such as lithology and topography, and their control on the delivery of runoff and other materials to stream systems [90]. High amounts of network catchment urban land use could shift fish assemblage composition from endemic faunas to more tolerant fish fauna, promoting fish assemblages with more regional similarity [91,92]. This change in taxonomic distinctness may also be due to a reduction in intolerant or specialist fish species and a decline in regional family, genus, and species assemblage specialization because of the degradation or loss of specific habitats or niches [93]. More significantly, these results suggest that high amounts of network catchment urban land use are negatively affecting taxonomic distinctness and may reflect the loss of endemic or sensitive fish taxa and increases in more tolerant taxa, leading to the homogenization of assemblages across study ecoregions [2,10,11,12]. These results emphasize how taxonomic distinctness measures can indicate unique changes occurring in stream fish assemblages resulting from anthropogenic land uses and that taxonomic distinctness may also be an effective metric for the bioassessment of streams along with richness and diversity [14,94].
The densities of road crossings and dams in catchments were also significant predictors of species richness, Shannon’s diversity, and beta diversity in some study ecoregions. Studies have emphasized how these landscape variables may have the potential to influence stream fishes, e.g., [51,52,55,95], and our results support this. However, research also suggests that greater numbers of dams and/or stream road crossings may potentially make stream habitats more favorable for more species than make them unfavorable to those species that could be negatively affected by barriers. A study of dam influences on stream fishes in Michigan and Wisconsin showed increases in IBI scores with more upstream dams [41]. They attributed their findings to potential modifications in habitat that benefited species including reduced sediment loads and potentially more stable stream flow regimes in streams downstream of dams. Ref. [51] also showed that lentic adapted species increase with dams, while intolerant, lithophilic, and rheophilic species declined across the conterminous USA.
4.4. Spatial Extent of Study and Scope of Natural and Anthropogenic Environmental Gradients Captured
The spatial extent over which this study was conducted offers important insights into the response of stream fish assemblages to natural landscape variables and anthropogenic land uses. Our study area included five ecoregions encompassing an area greater than two million square km, and we tested associations between stream fishes and landscape variables using data from 10,522 study sites on streams and rivers occurring within twenty-two states. This provided a unique opportunity for conducting regional comparisons based on extensive variability in fish assemblages and the characterizations of species compositions that were vital in summarizing patterns in biodiversity. Additionally, this large data set allowed us the ability to capture equally broad ranges in environmental characteristics to associate with biodiversity indices. While the amount of variance predicted in the biodiversity indices were moderately low, given the spatial extent of the study, species richness and taxonomic distinctness indices were relatively well predicted across ecoregions (i.e., mean adjusted R2 = 0.37 and 0.25, respectively, Figure 2). For Shannon’s diversity, Pielou’s evenness, beta diversity, and taxonomic diversity, we were likely unable to capture regionally specific mechanisms that may be most strongly influencing these indices.
5. Conclusions
This study emphasized that, across ecoregions, biodiversity indices for stream fish assemblages are significantly influenced by both natural landscape variables and anthropogenic land uses. Catchment area was a strong predictor of all biodiversity indices across study ecoregions and should be controlled for when utilizing biodiversity indices as response metrics in the assessments of stream integrity. Taxonomic distinctiveness was found to represent a unique attribute of stream fish assemblages compared to diversity and richness. Taxonomic distinctiveness was also predicted by local elevation, which should similarly be accounted for when using this index for assessment.
This study also showed the importance of anthropogenic land uses in predicting biodiversity indices, underscoring their value in identifying broad changes in stream fish assemblages resulting from anthropogenic stressors. Taxonomic distinctness was consistently predicted by network catchment urban land use, while network catchment agriculture was a better predictor of species richness and Shannon’s diversity across ecoregions. These findings reflect how unique the attributes of fish assemblages are influenced by anthropogenic land uses differently and that those changes are likely due in part to shifting species membership and not always due to the loss of species or individuals [8,10]. Based on our results, we recommend using multiple biodiversity measures to capture the unique attributes of stream fish assemblages when testing for the effects of anthropogenic stressors. This has important implications for the development of the multimetric indices of biotic integrity to assess streams and emphasizes the importance of identifying species-specific targets when implementing actions to conserve or protect biodiversity in stream systems.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/w15081591/s1, Table S1: Fish taxon list for study region.
Author Contributions
Conceptualization, D.T. and D.I.; methodology, D.T. and Y.T.; formal analysis, D.T. and Y.T.; resources, D.T. and D.I.; data curation, D.T.; writing—original draft preparation, D.T. and D.I.; writing—review and editing, D.T. and D.I.; visualization, D.T.; project administration, D.I.; funding acquisition, D.I. and D.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was conducted with support from the U.S. Geological Survey Aquatic GAP Project, U.S. Fish and Wildlife Service, National Fish Habitat Partnership, USDA’s Forest Service, grants from Michigan State University, a scholarship from the Austrian Marshall Plan Foundation, and a fellowship from the Red Cedar Fly Fishers.
Data Availability Statement
The data in this study are unavailable due to privacy restrictions.
Acknowledgments
We thank graduate committee members for providing feedback that helped to improve an early version of the manuscript and the anonymous reviewers that substantially helped to improve this manuscript. We are grateful for the discussion and insights provided by Arthur Cooper during the manuscript development and revisions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.I.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.H.; Soto, D.; Stiassny, M.L.J.; et al. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. Camb. Philos. Soc. 2006, 81, 163–182. [Google Scholar] [CrossRef] [PubMed]
- Helfman, G.S. Fish Conservation: A Guide to Understanding and Restoring Global Aquatic Biodiversity and Fishery Resources; Island Press: Washington, DC, USA, 2007. [Google Scholar]
- Jelks, H.L.; Walsh, S.J.; Burkhead, N.M.; Contreras-Balderas, S.; Díaz-Pardo, E.; Hendrickson, D.A.; Lyons, J.; Mandrak, N.E.; McCormick, F.; Nelson, J.S.; et al. Conservation status of imperiled North American freshwater and diadromous fishes. Fisheries 2008, 33, 372–407. [Google Scholar] [CrossRef]
- Vörösmarty, C.J.; McIntyre, P.B.; Gessner, M.O.; Dudgeon, D.; Prusevich, A.; Green, P.; Glidden, S.; Bunn, S.E.; Sullivan, C.A.; Liermann, C.R.; et al. Global threats to human water security and river biodiversity. Nature 2010, 467, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Burkhead, N.M. Extinction rates in North American freshwater fishes 1900–2010. BioScience 2012, 62, 798–808. [Google Scholar] [CrossRef]
- Dudgeon, D. Threats to freshwater biodiversity in a changing world. In Global Environmental Change; Springer: Berlin/Heidelberg, Germany, 2014; pp. 243–253. [Google Scholar]
- Rahel, F.J. Homogenization of fish faunas across the United States. Science 2000, 288, 854–856. [Google Scholar] [CrossRef] [PubMed]
- Scott, M.C.; Helfman, G.S. Native invasions, homogenization, and the mismeasure of integrity of fish assemblages. Fisheries 2001, 26, 6–15. [Google Scholar] [CrossRef]
- Olden, J.D.; Poff, N.L. Ecological processes driving biotic homogenization: Testing a mechanistic model using fish fauna. Ecology 2004, 85, 1867–1875. [Google Scholar] [CrossRef]
- Scott, M.C. Winners and losers among stream fishes in relation to land use legacies and urban development in the southeastern US. Biol. Conserv. 2006, 127, 301–309. [Google Scholar] [CrossRef]
- Villéger, S.; Blanchet, S.; Beauchard, O.; Oberdorff, T.; Brosse, S. Homogenization patterns of the world’s freshwater fish faunas. Proc. Natl. Acad. Sci. USA 2011, 108, 18003–18008. [Google Scholar] [CrossRef]
- Marr, S.M.; Olden, J.D.; Leprieur, F.; Arismendi, I.; Ćaleta, M.; Morgan, D.L.; Nocita, A.; Šanda, R.; Tarkan, A.S.; García-Berthou, E. A global assessment of freshwater fish introductions in Mediterranean-climate regions. Hydrobiologia 2013, 719, 317–329. [Google Scholar] [CrossRef]
- Toussaint, A.; Beauchard, O.; Oberdorff, T.; Brosse, S.; Villéger, S. Historical assemblage distinctiveness and the introduction of widespread non-native species explain worldwide changes in freshwater fish taxonomic dissimilarity. Glob. Ecol. Biogeogr. 2014, 23, 574–584. [Google Scholar] [CrossRef]
- Schweiger, O.; Klotz, S.; Durka, W.; Kühn, I. A comparative test of phylogenetic diversity indices. Oecologia 2008, 157, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Magurran, A.E. Measuring Biological Diversity, 2nd ed.; Blackwell: Oxford, UK, 2004. [Google Scholar]
- Wang, L.; Lyons, J.; Kanehl, P. Impacts of urbanization on stream habitat and fish across multiple spatial scales. Environ. Manag. 2001, 28, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Esselman, P.C.; Infante, D.M.; Wang, L.; Wu, D.; Cooper, A.R.; Taylor, W.W. An index of cumulative disturbance to river fish habitats of the conterminous United States from landscape anthropogenic activities. Ecol. Restor. 2011, 29, 133–151. [Google Scholar] [CrossRef]
- Hughes, R.M.; Kaufmann, P.R.; Herlihy, A.T.; Kincaid, T.M.; Reynolds, L.; Larsen, D.P. A process for developing and evaluating indices of fish assemblage integrity. Can. J. Fish. Aquat. Sci. 1998, 55, 1618–1631. [Google Scholar] [CrossRef]
- Pont, D.; Hugueny, B.; Beier, U.; Goffaux, D.; Melcher, A.; Noble, R.; Rogers, C.; Roset, N.; Schmutz, S. Assessing river biotic condition at the continental scale: A European approach using functional metrics and fish assemblages. J. Appl. Ecol. 2006, 43, 70–80. [Google Scholar] [CrossRef]
- Hocutt, C.H.; Wiley, E.O. (Eds.) The Zoogeography of North American Freshwater Fishes; John Wiley & Sons: New York, NY, USA, 1986. [Google Scholar]
- Mayden, R.L. Vicariance biogeography, parsimony, and evolution in North American freshwater fishes. Syst. Zool. 1988, 37, 329–355. [Google Scholar] [CrossRef]
- Schlosser, I.J. Stream fish ecology: A landscape perspective. BioScience 1991, 41, 704–712. [Google Scholar] [CrossRef]
- Goldstein, R.M.; Meador, M.R. Comparisons of fish species traits from small streams to large rivers. Tran. Am. Fish. Soc. 2004, 133, 971–983. [Google Scholar] [CrossRef]
- Stirling, G.; Wilsey, B. Empirical relationships between species richness, evenness, and proportional diversity. Am. Nat. 2001, 158, 286–299. [Google Scholar] [CrossRef]
- Legendre, P.; De Cáceres, M. Beta diversity as the variance of community data: Dissimilarity coefficients and partitioning. Ecol. Lett. 2013, 16, 951–963. [Google Scholar] [CrossRef] [PubMed]
- Podani, J.; Ricotta, C.; Schmera, D. A general framework for analyzing beta diversity, nestedness and related community-level phenomena based on abundance data. Ecol. Complex. 2013, 15, 52–61. [Google Scholar] [CrossRef]
- Legendre, P. Interpreting the replacement and richness difference components of beta diversity. Glob. Ecol. Biogeogr. 2014, 23, 1324–1334. [Google Scholar] [CrossRef]
- Magurran, A.E.; Phillip, D.A.T. Implications of species loss in freshwater fish assemblages. Ecography 2001, 24, 645–650. [Google Scholar] [CrossRef]
- Warwick, R.M.; Clarke, K.R. New ‘biodiversity’ measures reveal a decrease in taxonomic distinctness with increasing stress. Mar. Ecol. Prog. Ser. 1995, 129, 301–305. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. A taxonomic distinctness index and its statistical properties. J. Appl. Ecol. 1998, 35, 523–531. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. A further biodiversity index applicable to species lists: Variation in taxonomic distinctness. Mar. Ecol. Prog. Ser. 2001, 21, 265–278. [Google Scholar] [CrossRef]
- Hughes, R.M.; Whittier, C.M.R.; Larsen, D.P. A regional framework for establishing recovery criteria. Environ. Manag. 1990, 14, 673–683. [Google Scholar] [CrossRef]
- Angermeier, P.L.; Winston, M.R. Characterizing fish community diversity across Virginia landscapes: Prerequisite for conservation. Ecol. Appl. 1999, 9, 335–349. [Google Scholar] [CrossRef]
- Maxwell, J.R.; Edwards, C.J.; Jensen, M.E.; Paustian, S.J.; Parrott, H.; Hill, D.M. A Hierarchical Framework of Aquatic Ecological Units in North America (Nearctic Zone). In General Technical Report NC–176; USDA Forest Service, North Central Forest Experiment Station: St. Paul, MN, USA, 1995. [Google Scholar]
- Matthews, W.J. Patterns in Freshwater Fish Ecology; Chapman and Hall: New York, NY, USA, 1998. [Google Scholar]
- Abell, R.; Olsen, D.M.; Dinerstein, E.; Hurley, P.; Diggs, J.T.; Eichbaum, W.; Walters, S.; Wettengel, W.; Allnutt, T.; Loucks, C.J.; et al. Freshwater Ecoregions of North America: A Conservation Assessment; Island Press: Washington, DC, USA, 2000. [Google Scholar]
- Abell, R.; Thieme, M.L.; Revenga, C.; Bryer, M.; Kottelat, M.; Bogutskaya, N.; Coad, B.; Mandrak, N.; Contreras Balderas, S.; Bussing, W.; et al. Freshwater ecoregions of the world: A new map of biogeographic units for freshwater biodiversity conservation. BioScience 2008, 58, 403–414. [Google Scholar] [CrossRef]
- Deweber, J.T.; Sleezer, L.; Frimpong, E.A. A new regionalization framework to quantify how physiography mediates the effect of land use on stream fishes 2019. In Advances in Understanding Landscape Influences on Freshwater Habitats and Biological Assemblages; Hughes, R.M., Infante, D.M., Wang, L., Chen, K., Terra, B.F., Eds.; American Fisheries Society Symposium: Bethesda, MD, USA, 2019; pp. 321–350. [Google Scholar]
- Herlihy, A.T.; Sifneos, J.C.; Hughes, R.M.; Peck, D.V.; Mitchell, R.M. Lotic fish assemblage clusters across the conterminous United States and their associations with environmental variables 2019. In Advances in Understanding Landscape Influences on Freshwater Habitats and Biological Assemblages; Hughes, R.M., Infante, D.M., Wang, L., Chen, K., Terra, B.F., Eds.; American Fisheries Society Symposium: Bethesda, MD, USA, 2019; pp. 385–408. [Google Scholar]
- U.S. Environmental Protection Agency. U.S. Geological Survey (USEPA & USGS) National Hydrography Dataset Plus, NHDPlusV1. 2005. Available online: www.horizon-systems.com/nhdplus/ (accessed on 12 November 2014).
- Wang, L.; Infante, D.; Esselman, P.; Cooper, A.; Wu, D.; Taylor, W.; Beard, D.; Whelan, G.; Ostroff, A. A hierarchical spatial framework and database for the national river fish habitat condition assessment. Fisheries 2011, 36, 436–449. [Google Scholar] [CrossRef]
- Gesch, D.B. The National Elevation Dataset. In Digital Elevation Model Technologies and Applications: The DEM User’s Manual, 2nd ed.; Maune, D., Ed.; ASPRS: Bethesda, MD, USA, 2007; pp. 99–118. [Google Scholar]
- Daly, C.; Halbleib, M.; Smith, J.I.; Gibson, W.P.; Doggett, M.K.; Taylor, G.H.; Curtis, J.; Pasteris, P.P. Physiographically sensitive mapping of climatological temperature and precipitation across the conterminous United States. Int. J. Climatol. 2008, 28, 2031–2064. [Google Scholar] [CrossRef]
- PRISM Climate Group. Oregon State University. 2013. Available online: http://www.prism.oregonstate.edu/ (accessed on 14 March 2023).
- Tsang, Y.; Wieferich, D.; Fung, K.; Infante, D.M.; Cooper, A.R. An approach for aggregating upstream catchment information to support research and management of fluvial systems across large landscapes. SpringerpPlus 2014, 23, 589. [Google Scholar] [CrossRef] [PubMed]
- Soller, D.R.; Reheis, M.C. Surficial Materials in the Conterminous United States: U.S. Geological Survey 2004. Open–File Report 03–275, Scale 1:5,000,000. Available online: http://pubs.usgs.gov/of/2003/of03–275/ (accessed on 12 November 2014).
- Cress, J.; Soller, D.; Sayre, R.; Comer, P.; Warner, H. Terrestrial Ecosystems, Surficial Lithology of the Conterminous United States: U.S. Geological Survey Scientific Investigations Map 3126, scale 1:5,000,000, 1 Sheet. 2010. Available online: http://pubs.usgs.gov/sim/3126 (accessed on 12 November 2014).
- Wolock, D.M. Base-Flow Index Grid for the Conterminous United States: U.S. Geological Survey Open-File Report 03–263, Digital Data Set. 2003. Available online: http://water.usgs.gov/lookup/getspatial?bfi48grd (accessed on 12 November 2014).
- Perkin, J.S.; Troia, M.J.; Shaw, D.C.R.; Gerken, J.E.; Gido, K.B. Multiple watershed alterations influence fish community structure in Great Plains prairie streams. Ecol. Freshw. Fish 2016, 25, 141–155. [Google Scholar] [CrossRef]
- Cheng, S.T.; Herricks, E.E.; Tsai, W.P.; Chang, F.J. Assessing the natural and anthropogenic influences on basin-wide fish species richness. Sci. Total Environ. 2016, 572, 825–836. [Google Scholar] [CrossRef]
- Cooper, A.R.; Infante, D.M.; Wehrly, K.E.; Wang, L.; Brenden, T.O. Identifying indicators and quantifying large-scale effects of dams on fishes. Ecol. Indic. 2016, 61, 646–657. [Google Scholar] [CrossRef]
- Cooper, A.R.; Infante, D.M.; Daniel, W.M.; Wehrly, K.E.; Wang, L.; Brenden, T.O. Assessment of dam effects on streams and fish assemblages of the conterminous USA. Sci. Total Environ. 2017, 586, 879–889. [Google Scholar] [CrossRef]
- Thornbrugh, D.J.; Infante, D.M. Landscape effects on steam fishes: Broad-scale responses to anthropogenic land use across temperate mesic regions of the United States. In Advances in Understanding Landscape Influences on Freshwater Habitats and Biological Assemblages; Hughes, R.M., Infante, D.M., Wang, L., Chen, K., Terra, B.F., Eds.; American Fisheries Society Symposium: Bethesda, MD, USA, 2019; p. 90. [Google Scholar]
- Bouska, W.W.; Paukert, C.P. Road crossing designs and their impact on fish assemblages of Great Plains streams. Trans. Am. Fish. Soc. 2010, 139, 214–222. [Google Scholar] [CrossRef]
- Januchowski–Hartley, S.R.; McIntyre, P.B.; Diebel, M.; Doran, P.J.; Infante, D.M.; Joseph, C.; Allan, J.D. Restoring aquatic ecosystem connectivity requires expanding barrier inventories. Fron. Ecol. Environ. 2013, 11, 211–217. [Google Scholar] [CrossRef]
- Homer, C.; Huang, C.; Yang, L.; Wylie, B.; Coan, M. Development of a 2001 National Landcover Database for the United States. PERS 2004, 70, 829–840. [Google Scholar]
- U.S. Environmental Protection Agency (USEPA) United States Environmental Protection Agency Multi-Resolution Land Characteristics Consortium. 2008. Available online: www.epa.gov/mrlc/ (accessed on 12 October 2021).
- U.S. Geological Survey (USGS) National Anthropogenic Barrier Dataset (NABD). 2012. Available online: https://www.sciencebase.gov/catalog/item/512cf142e4b0855fde669828 (accessed on 12 November 2014).
- U.S. Census Bureau, Redistricting Census 2000 TIGER/ Line Files [Machine-Readable Data Files]. Washington, DC: U.S. Census Bureau. 2002. Available online: www.census.gov/geo/maps-data/data/tiger.html (accessed on 12 November 2014).
- Esselman, P.C.; Infante, D.M.; Wang, L.; Cooper, A.R.; Wieferich, D.; Tsang, Y.; Thornbrugh, D.J.; Taylor, W.W. Regional fish community indicators of landscape disturbance to catchments of conterminous United States. Ecol. Indic. 2013, 26, 163–173. [Google Scholar] [CrossRef]
- Daniel, W.M.; Infante, D.M.; Hughes, R.M.; Tsang, Y.; Esselman, P.C.; Wieferich, D.; Herreman, K.; Cooper, A.R.; Wang, L.; Taylor, W.W. Characterizing coal and mineral mines as a regional source of stress to stream fish assemblages. Ecol. Indic. 2015, 50, 50–61. [Google Scholar] [CrossRef]
- ITIS Retrieved [04, 26, 2010], from the Integrated Taxonomic Information System. 2010. Available online: http://www.itis.gov (accessed on 12 October 2021).
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Pielou, E.C. Ecological Diversity; Wiley and Sons: New York, NY, USA, 1975. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. ‘Vegan’: Community Ecology Package, R Package Version 2.0–9. 2020. Available online: http://CRAN.R-project.org/package=vegan (accessed on 12 January 2023).
- R Core Team R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. 2022. Available online: http://www.R-project.org/ (accessed on 12 January 2023).
- Dray, S.; Bauman, D.; Blanchet, G.; Borcard, D.; Clappe, S.; Guenard, G.; Jombart, T.; Larocque, G.; Legendre, P.; Madi, N.; et al. Adespatial: Multivariate Multiscale Spatial Analysis, R Package Version 0.3-16. 2022. Available online: https://CRAN.R-project.org/package=adespatial (accessed on 12 January 2023).
- Little, J.D.C.; Murty, K.G.; Sweeney, D.W.; Karel, C. An algorithm for the traveling salesman problem. Oper. Res. 1963, 11, 972–989. [Google Scholar] [CrossRef]
- Miller, A.J. Subset Selection in Regression, 2nd ed.; Chapman and Hall: New York, NY, USA, 2002. [Google Scholar]
- Lumley, T. Based on Fortran Code by Alan Miller. Leaps: Regression Subset Selection, R Package Version 3.1; 2020. Available online: https://CRAN.R-project.org/package=leaps (accessed on 12 January 2023).
- McCuen, R.H.; Snyder, W.M. Hydrologic Modelling: Statistical Method and Applications; Prentice-Hall, A Division of Simon & Schuster, Inc.: Englewood Cliffs, NJ, USA, 1985. [Google Scholar]
- Tsang, Y.; Felton, G.K.; Moglen, G.E.; Paul, M. Region of influence method improves macroinvertebrate predictive models in Maryland. Ecol. Modell. 2011, 222, 3473–3485. [Google Scholar] [CrossRef]
- Fletcher, T.D. QuantPsyc: Quantitative Psychology Tools, R Package Version 1.6. 2022. Available online: https://CRAN.R-project.org/package=QuantPsyc (accessed on 12 January 2023).
- Legendre, P.; Legendre, L. Numerical Ecology, 3rd English ed.; Elsevier Science BV: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Heino, J.; Mykrä, H.; Hämäläinen, H.; Aroviita, J.; Muotka, T. Responses of taxonomic distinctness and species diversity indices to anthropogenic impacts and natural environmental gradients in stream macroinvertebates. Freshw. Biol. 2007, 52, 1846–1861. [Google Scholar] [CrossRef]
- Heino, J.; Soininen, J.; Lappalainen, J.; Virtanen, R. The relationship between species richness and taxonomic distinctness in freshwater organisms. Limnol. Oceanogr. 2005, 5, 978–986. [Google Scholar] [CrossRef]
- Eadie, J.M.; Hurly, T.A.; Montgomerie, R.D.; Teather, K.L. Lakes and rivers as islands: Species—Area relationships in the fish faunas of Ontario. Environ. Biol. Fishes 1986, 15, 81–89. [Google Scholar] [CrossRef]
- Angermeier, P.L.; Schlosser, I.J. Species-area relationship for stream fishes. Ecology 1989, 70, 1450–1462. [Google Scholar] [CrossRef]
- Thornbrugh, D.J.; Gido, K.B. Influence of spatial positioning within stream networks on fish assemblage structure in the Kansas River basin, USA. Can. J. Fish. Aquat. Sci. 2010, 67, 143–156. [Google Scholar] [CrossRef]
- Sheldon, A.L. Species diversity and longitudinal succession in stream fishes. Ecology 1968, 49, 193–198. [Google Scholar] [CrossRef]
- Vannote, R.L.; Minshall, G.W.; Cummins, K.W.; Sedell, J.R.; Cushing, C.E. The river continuum concept. Can. J. Fish. Aquat. Sci. 1980, 37, 130–137. [Google Scholar] [CrossRef]
- Marsh-Matthews, E.; Matthews, W.J. Geographic, terrestrial and aquatic factors: Which most influence the structure of stream fish assemblages in the Midwestern United States? Ecol. Freshw. Fish 2000, 9, 9–21. [Google Scholar] [CrossRef]
- Angermeier, P.L.; Winston, M.R. Local vs. regional influences on local diversity in stream fish communities of Virginia. Ecology 1998, 79, 911–927. [Google Scholar] [CrossRef]
- Jackson, D.A.; Peres-Neto, P.R.; Olden, J.D. What controls who is where in freshwater fish communities—The roles of biotic, abiotic, and spatial factors. Can. J. Fish. Aquat. Sci. 2011, 58, 157–170. [Google Scholar]
- Zorn, T.G.; Seelbach, P.W.; Wiley, M.J. Distributions of stream fishes and their relationship to stream size and hydrology in Michigan’s Lower Peninsula. Trans. Am. Fish. Soc. 2002, 131, 70–85. [Google Scholar] [CrossRef]
- Wehrly, K.E.; Wiley, M.J.; Seelbach, P.W. Classifying regional variation in thermal regime based on stream fish community patterns. Trans. Am. Fish. Soc. 2003, 132, 18–38. [Google Scholar] [CrossRef]
- Buisson, L.; Thuiller, W.; Lek, S.; Limp, P.; Grenouillet, G. Climate change hastens the turnover of stream fish assemblages. Glob. Chang. Biol. 2008, 14, 2232–2248. [Google Scholar] [CrossRef]
- Olden, J.D.; Kennard, M.J.; Leprieur, F.; Tedesco, P.A.; Winemiller, K.O.; García-Berthou, E. Conservation biogeography of freshwater fishes: Recent progress and future challenges. Divers. Distrib. 2010, 16, 496–513. [Google Scholar] [CrossRef]
- Wang, L.; Lyons, J.; Kanehl, P.; Gatti, R. Influences of watershed land use on habitat quality and biotic integrity in Wisconsin streams. Fisheries 1997, 22, 6–12. [Google Scholar] [CrossRef]
- Utz, R.M.; Hilderbrand, R.M.; Raesly, R.L. Regional differences in patterns of fish species loss with changing land use. Biol. Conserv. 2010, 143, 688–699. [Google Scholar] [CrossRef]
- McKinney, M.L.; Lockwood, J.L. Biotic homogenization: A few winners replacing many losers in the next mass extinction. Trends Ecol. Evol. 1999, 14, 450–453. [Google Scholar] [CrossRef] [PubMed]
- Rahel, F.J. Homogenization of freshwater faunas. Annu. Rev. Ecol. Syst. 2002, 33, 291–315. [Google Scholar] [CrossRef]
- Villéger, S.; Miranda, J.R.; Hernández, D.F.; Mouillot, D. Contrasting changes in taxonomic vs. functional diversity of tropical fish communities after habitat degradation. Ecol. Appl. 2010, 20, 1512–1522. [Google Scholar] [CrossRef] [PubMed]
- Stoddard, J.L.; Herlihy, A.T.; Peck, D.V.; Hughes, R.M.; Whittier, T.R.; Tarquinio, E. A process for creating multimetric indices for large-scale aquatic surveys. J. N. Am. Benthol. Soc. 2008, 27, 878–891. [Google Scholar] [CrossRef]
- Perkin, J.S.; Gido, K.B. Fragmentation alters stream fish community structure in dendritic ecological networks. Ecol. Appl. 2012, 22, 2176–2187. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).