
Microorganisms Directly Affected Sediment Carbon–Nitrogen Coupling in Two Constructed Wetlands


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Area Selection and Sample Collection
2.2. Determination of Physicochemical Properties of Sediment and Plant
2.3. Prediction of Microbial Function
2.4. Data Analysis
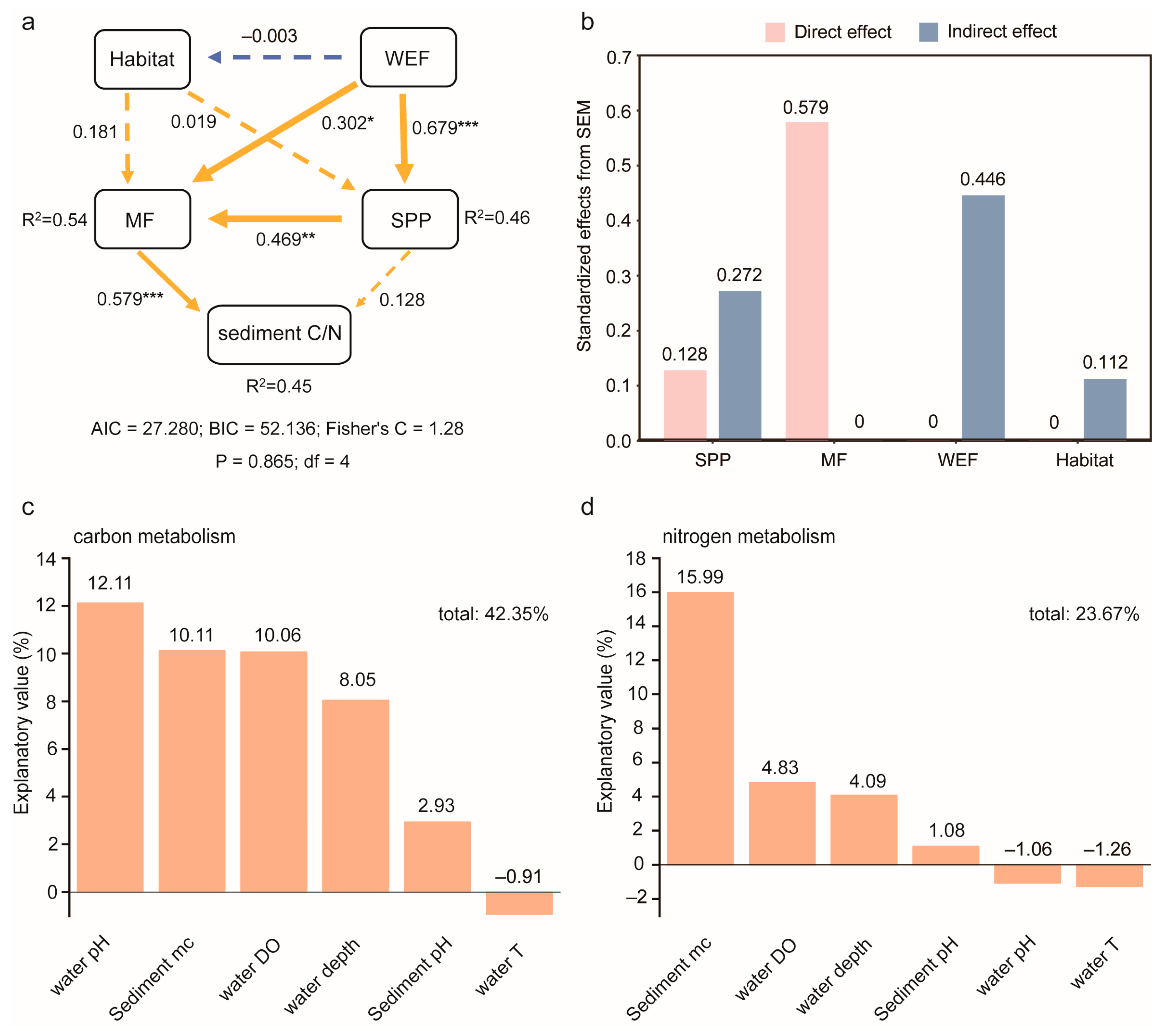
3. Results
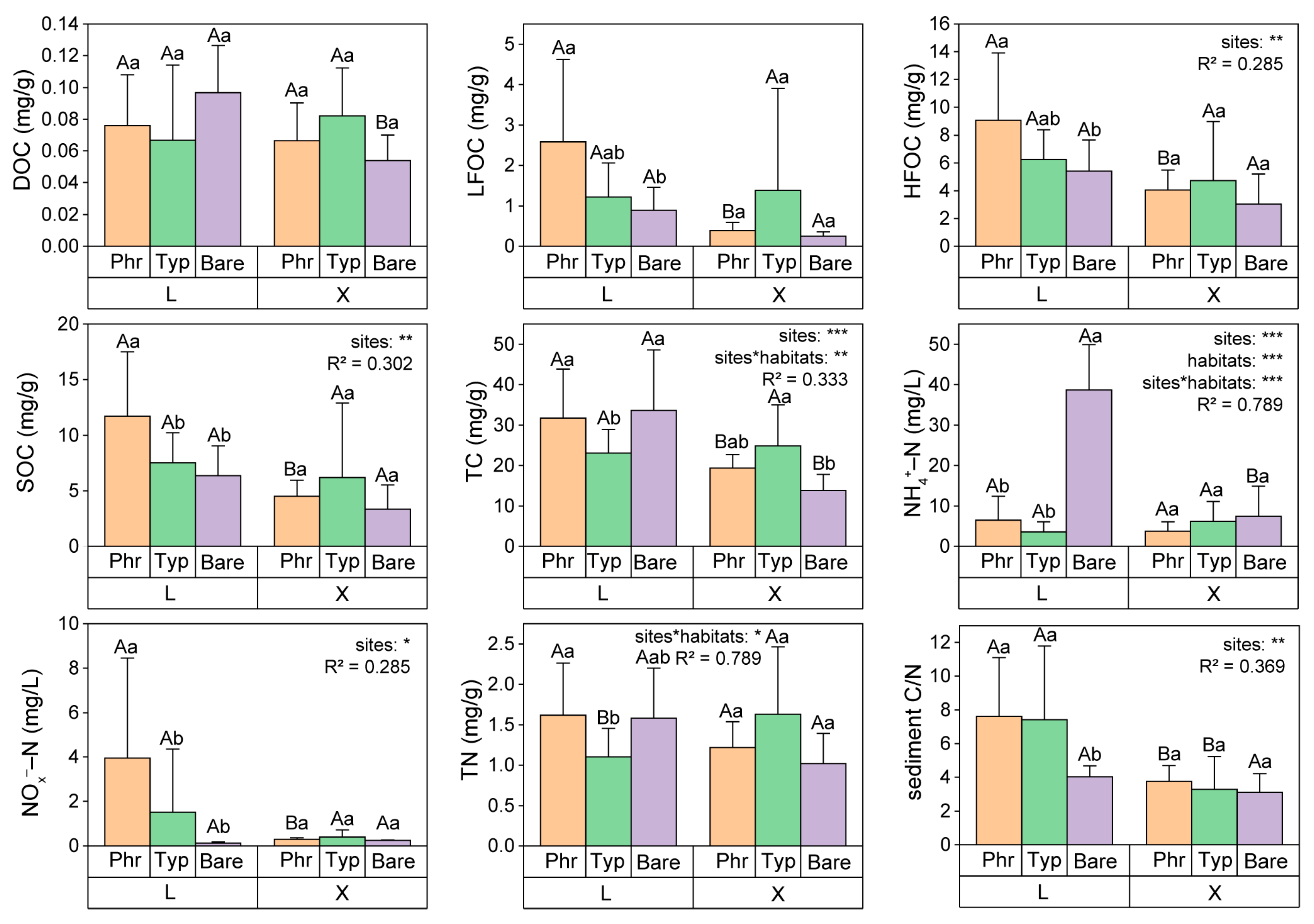
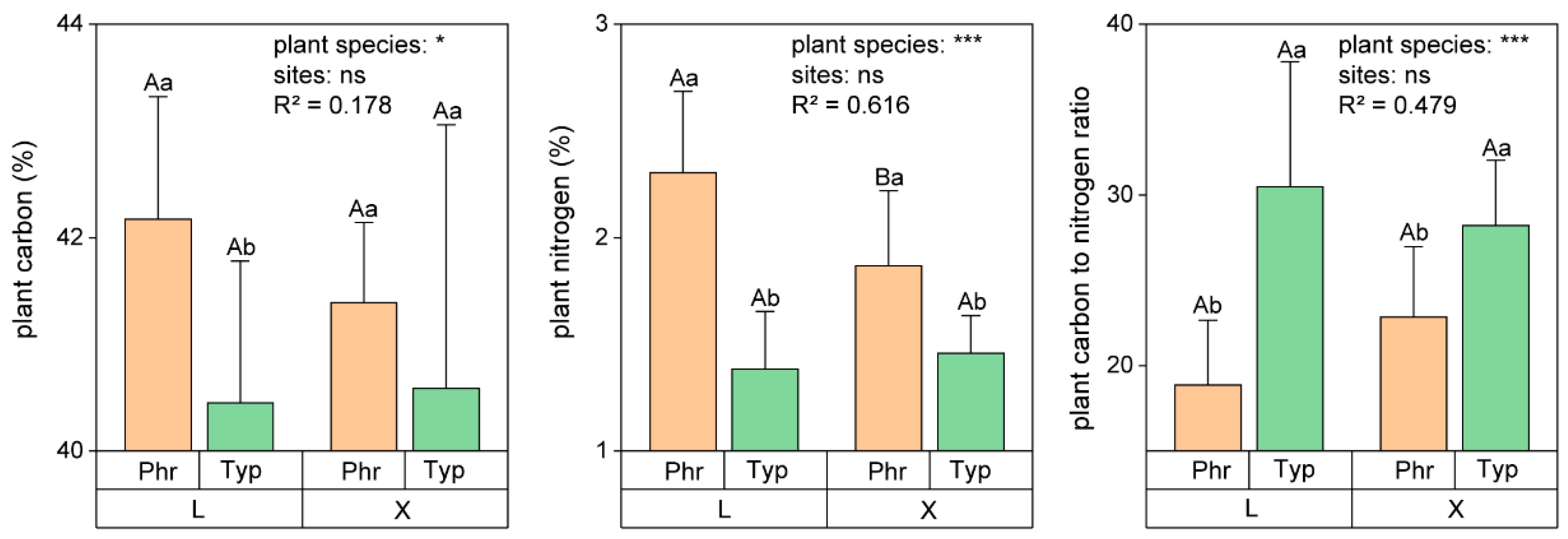
3.1. Differences of Carbon and Nitrogen Components in Sediments and Plants in the Two Constructed Wetlands
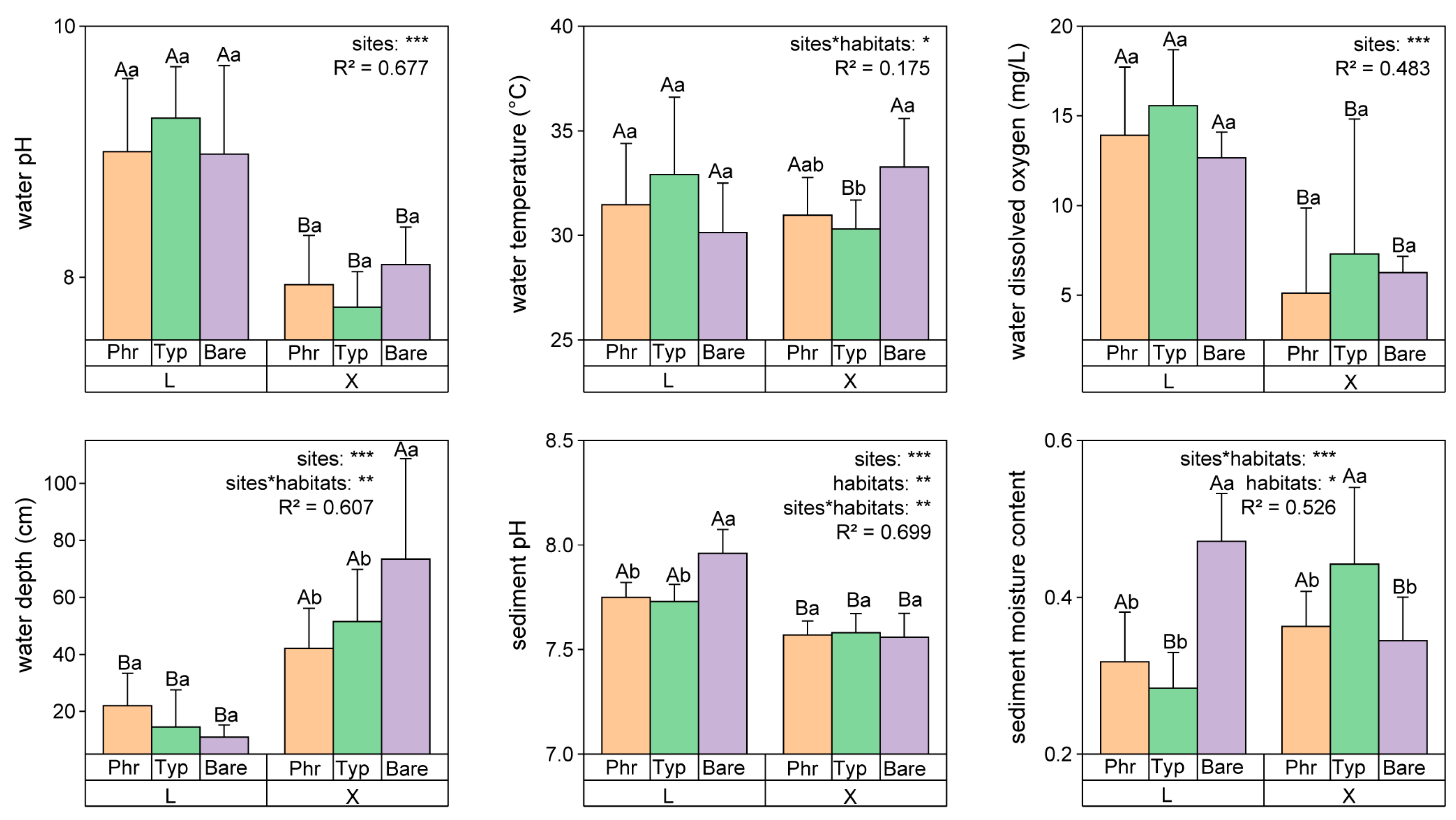
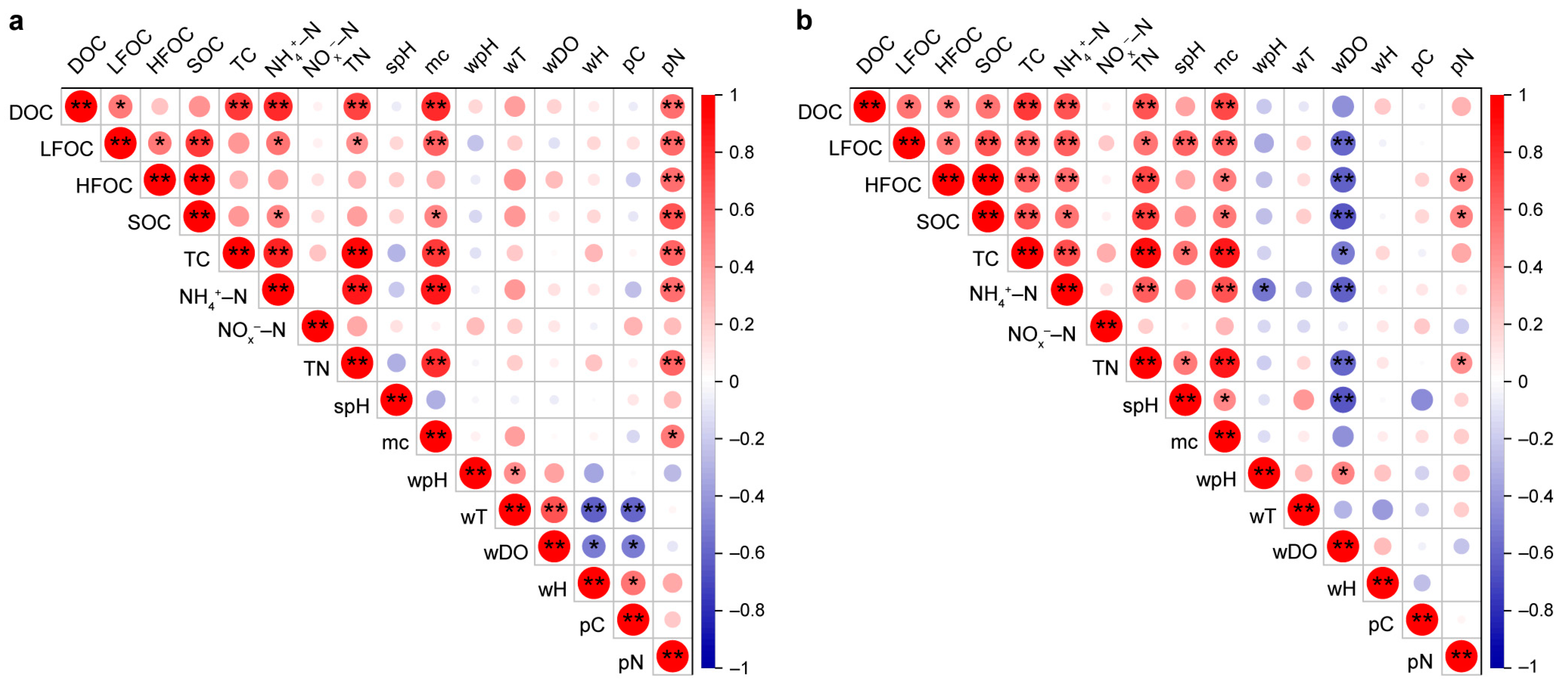
3.2. Differences in Environmental Factors and Their Correlation with Carbon and Nitrogen Fractions of Sediments and Plants
3.3. Microbial Richness and Community Composition in Different Habitats and Factors Affecting Sediment Carbon to Nitrogen Ratio
4. Discussion
4.1. Carbon–Nitrogen Coupling in Constructed Wetland Plants
4.2. Carbon–Nitrogen Coupling in Constructed Wetland Sediments
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Merot, P.; Squividant, H.; Aurousseau, P.; Hefting, M.; Burt, T.; Maitre, V.; Kruk, M.; Butturini, A.; Thenail, C.; Viaud, V. Testing a Climato-Topographic Index for Predicting Wetlands Distribution along an European Climate Gradient. Ecol. Model. 2003, 163, 51–71. [Google Scholar] [CrossRef]
- Tiner, R.W. Assessing Cumulative Loss of Wetland Functions in the Nanticoke River Watershed Using Enhanced National Wetlands Inventory Data. Wetlands 2005, 25, 405–419. [Google Scholar] [CrossRef]
- Moomaw, W.R.; Chmura, G.L.; Davies, G.T.; Finlayson, C.M.; Middleton, B.A.; Natali, S.M.; Perry, J.E.; Roulet, N.; Sutton-Grier, A.E. Wetlands in a Changing Climate: Science, Policy and Management. Wetlands 2018, 38, 183–205. [Google Scholar] [CrossRef]
- Nahlik, A.M.; Fennessy, M.S. Carbon Storage in US Wetlands. Nat. Commun. 2016, 7, 13835. [Google Scholar] [CrossRef] [PubMed]
- Roe, S.; Streck, C.; Obersteiner, M.; Frank, S.; Griscom, B.; Drouet, L.; Fricko, O.; Gusti, M.; Harris, N.; Hasegawa, T.; et al. Contribution of the Land Sector to a 1.5 °C World. Nat. Clim. Chang. 2019, 9, 817–828. [Google Scholar] [CrossRef]
- Hu, S.; Niu, Z.; Chen, Y.; Li, L.; Zhang, H. Global Wetlands: Potential Distribution, Wetland Loss, and Status. Sci. Total Environ. 2017, 586, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Davidson, N.C. How Much Wetland Has the World Lost? Long-Term and Recent Trends in Global Wetland Area. Mar. Freshw. Res. 2014, 65, 934–941. [Google Scholar] [CrossRef]
- Vymazal, J. Constructed Wetlands for Treatment of Industrial Wastewaters: A Review. Ecol. Eng. 2014, 73, 724–751. [Google Scholar] [CrossRef]
- Li, Y.; Zhu, G.; Ng, W.J.; Tan, S.K. A Review on Removing Pharmaceutical Contaminants from Wastewater by Constructed Wetlands: Design, Performance and Mechanism. Sci. Total Environ. 2014, 468, 908–932. [Google Scholar] [CrossRef]
- Knicker, H. Soil Organic N—An under-Rated Player for C Sequestration in Soils? Soil Biol. Biochem. 2011, 43, 1118–1129. [Google Scholar] [CrossRef]
- Zhao, X.; Chen, J.; Guo, M.; Li, C.; Hou, N.; Bai, S. Constructed Wetlands Treating Synthetic Wastewater in Response to Day-Night Alterations: Performance and Mechanisms. Chem. Eng. J. 2022, 446, 137460. [Google Scholar] [CrossRef]
- Wu, H.; Wang, R.; Yan, P.; Wu, S.; Chen, Z.; Zhao, Y.; Cheng, C.; Hu, Z.; Zhuang, L.; Guo, Z.; et al. Constructed Wetlands for Pollution Control. Nat. Rev. Earth Environ. 2023, 4, 218–234. [Google Scholar] [CrossRef]
- Zaehle, S.; Dalmonech, D. Carbon-Nitrogen Interactions on Land at Global Scales: Current Understanding in Modelling Climate Biosphere Feedbacks. Curr. Opin. Environ. Sustain. 2011, 3, 311–320. [Google Scholar] [CrossRef]
- Niu, S.; Song, L.; Wang, J.; Luo, Y.; Yu, G. Dynamic Carbon-Nitrogen Coupling under Global Change. Sci. China Life Sci. 2023, 66, 771–782. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.Y.; Huang, N.; Zhao, C.M.; Li, J.H. Responses of Carbon Cycling and Soil Organic Carbon Content to Nitrogen Addition in Grasslands Globally. Soil Biol. Biochem. 2023, 186, 109164. [Google Scholar] [CrossRef]
- Cao, Q.; Wang, H.; Zhang, Y.; Lal, R.; Wang, R.; Ge, X.; Liu, J. Factors Affecting Distribution Patterns of Organic Carbon in Sediments at Regional and National Scales in China. Sci. Rep. 2017, 7, 5497. [Google Scholar] [CrossRef] [PubMed]
- Qi, Q.; Zhang, D.; Zhang, M.; Tong, S.; Wang, W.; An, Y. Spatial Distribution of Soil Organic Carbon and Total Nitrogen in Disturbed Carex Tussock Wetland. Ecol. Indic. 2021, 120, 106930. [Google Scholar] [CrossRef]
- Wang, J.; Chen, G.; Fu, Z.; Qiao, H.; Liu, F. Assessing Wetland Nitrogen Removal and Reed (Phragmites Australis) Nutrient Responses for the Selection of Optimal Harvest Time. J. Environ. Manag. 2021, 280, 111783. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Leng, Z.; Li, J.; Yan, C.; Wang, X.; Jia, H.; Chen, L.; Zhang, S.; Zheng, X.; Du, D. Sulfur Mediated Heavy Metal Biogeochemical Cycles in Coastal Wetlands: From Sediments, Rhizosphere to Vegetation. Front. Environ. Sci. Eng. 2022, 16, 102. [Google Scholar] [CrossRef]
- Chen, L.; Fang, K.; Wei, B.; Qin, S.; Feng, X.; Hu, T.; Ji, C.; Yang, Y. Soil Carbon Persistence Governed by Plant Input and Mineral Protection at Regional and Global Scales. Ecol. Lett. 2021, 24, 1018–1028. [Google Scholar] [CrossRef]
- Wang, L.; Mei, W.; Yin, Q.; Guan, Y.; Le, Y.; Fu, X. The Variability in CO2 Fluxes at Different Time Scales in Natural and Reclaimed Wetlands in the Yangtze River Estuary and Their Key Influencing Factors. Sci. Total Environ. 2021, 799, 149441. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.-M.; Lu, Y.-T.; Qiu, Q.-L.; Fan, D.-M.; Wang, X.-C.; Zheng, X.-Q. Influence of Different Nitrogen Sources on Carbon and Nitrogen Metabolism and Gene Expression in Tea Plants (Camellia sinensis L.). Plant Physiol. Biochem. 2021, 167, 561–566. [Google Scholar] [CrossRef] [PubMed]
- Moreau, D.; Bardgett, R.D.; Finlay, R.D.; Jones, D.L.; Philippot, L. A Plant Perspective on Nitrogen Cycling in the Rhizosphere. Funct. Ecol. 2019, 33, 540–552. [Google Scholar] [CrossRef]
- Averill, C.; Turner, B.L.; Finzi, A.C. Mycorrhiza-Mediated Competition between Plants and Decomposers Drives Soil Carbon Storage. Nature 2014, 505, 543–545. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Li, B.; Wu, J.; Hu, S. Invasive Plants Differentially Affect Soil Biota through Litter and Rhizosphere Pathways: A Meta-Analysis. Ecol. Lett. 2019, 22, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Witzgall, K.; Vidal, A.; Schubert, D.I.; Höschen, C.; Schweizer, S.A.; Buegger, F.; Pouteau, V.; Chenu, C.; Mueller, C.W. Particulate Organic Matter as a Functional Soil Component for Persistent Soil Organic Carbon. Nat. Commun. 2021, 12, 4115. [Google Scholar] [CrossRef] [PubMed]
- Perkowski, E.A.; Waring, E.F.; Smith, N.G. Root Mass Carbon Costs to Acquire Nitrogen Are Determined by Nitrogen and Light Availability in Two Species with Different Nitrogen Acquisition Strategies. J. Exp. Bot. 2021, 72, 5766–5776. [Google Scholar] [CrossRef] [PubMed]
- Hoek, T.A.; Axelrod, K.; Biancalani, T.; Yurtsev, E.A.; Liu, J.; Gore, J. Resource Availability Modulates the Cooperative and Competitive Nature of a Microbial Cross-Feeding Mutualism. PLoS Biol. 2016, 14, e1002606. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Chen, M.; Yang, Z.; Cong, M.; Zhu, X.; Jia, H. Soil Microbial Community Response to Nitrogen Application on a Swamp Meadow in the Arid Region of Central Asia. Front. Microbiol. 2022, 12, 797306. [Google Scholar] [CrossRef]
- Montesinos-Navarro, A. Nitrogen Transfer between Plant Species with Different Temporal N-Demand. Ecol. Lett. 2023, 26, 1676–1686. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Soong, J.L.; Horton, A.J.; Campbell, E.E.; Haddix, M.L.; Wall, D.H.; Parton, W.J. Formation of Soil Organic Matter via Biochemical and Physical Pathways of Litter Mass Loss. Nat. Geosci. 2015, 8, 776–779. [Google Scholar] [CrossRef]
- Xia, S.; Song, Z.; Singh, B.P.; Guo, L.; Bolan, N.; Wang, W.; Lin, G.; Fang, Y.; Wen, X.; Wang, J.; et al. Contrasting Patterns and Controls of Soil Carbon and Nitrogen Isotope Compositions in Coastal Wetlands of China. Plant Soil 2023, 489, 483–505. [Google Scholar] [CrossRef]
- Lin, Y.; Tang, K.W.; Ye, G.; Yang, P.; Hu, H.-W.; Tong, C.; Zheng, Y.; Feng, M.; Deng, M.; He, Z.-Y.; et al. Community Assembly of Comammox Nitrospira in Coastal Wetlands across Southeastern China. Appl. Environ. Microbiol. 2023, 89, e0080723. [Google Scholar] [CrossRef] [PubMed]
- Nissan, A.; Alcolombri, U.; Peleg, N.; Galili, N.; Jimenez-Martinez, J.; Molnar, P.; Holzner, M. Global Warming Accelerates Soil Heterotrophic Respiration. Nat. Commun. 2023, 14, 3452. [Google Scholar] [CrossRef]
- Domeignoz-Horta, L.A.; Pold, G.; Liu, X.J.A.; Frey, S.D.; Melillo, J.M.; DeAngelis, K.M. Microbial Diversity Drives Carbon Use Efficiency in a Model Soil. Nat. Commun. 2020, 11, 3684. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Zhang, J.; Muneer, M.A.; Xue, K.; Niu, H.; Ji, B. Plant Community and Soil Available Nutrients Drive Arbuscular Mycorrhizal Fungal Community Shifts during Alpine Meadow Degradation. Fungal Ecol. 2023, 62, 101211. [Google Scholar] [CrossRef]
- Huang, X.; Li, Y.; Lin, H.; Wen, X.; Liu, J.; Yuan, Z.; Fu, C.; Zheng, B.; Gong, L.; Zhan, H.; et al. Flooding Dominates Soil Microbial Carbon and Phosphorus Limitations in Poyang Lake Wetland, China. Catena 2023, 232, 107468. [Google Scholar] [CrossRef]
- Li, J.; Jiang, M.; Pei, J.; Fang, C.; Li, B.; Nie, M.; Scott, T. Convergence of Carbon Sink Magnitude and Water Table Depth in Global Wetlands. Ecol. Lett. 2023, 26, 797–804. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, F.; Li, X.; Li, C.; Zhao, Y.; Gao, Y.; Liu, J. Effects of Plants and Soil Microorganisms on Organic Carbon and the Relationship between Carbon and Nitrogen in Constructed Wetlands. Environ. Sci. Pollut. Res. 2023, 30, 62249–62261. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, B.; Jing, Y.; Kang, X.; Zhang, C. Design of Constructed Wetland in Xinxue River Estuary into Nansi Lake in Eastern Line of South-to-North Water Transfer Project. China Water Wastewater 2008, 24, 49–51. [Google Scholar]
- Cao, Q.; Wang, R.; Zhang, H.; Ge, X.; Liu, J. Distribution of Organic Carbon in the Sediments of Xinxue River and the Xinxue River Constructed Wetland, China. PLoS ONE 2015, 10, e134713. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Zhang, X.; Li, Y.; Jiao, Y.; Fan, L.; Jiang, Y.; Qu, C.; Filimonenko, E.; Jiang, Y.; Tian, X.; et al. Nitrogen Fertilizer Builds Soil Organic Carbon under Straw Return Mainly via Microbial Necromass Formation. Soil Biol. Biochem. 2024, 188, 109223. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian Classifier for Rapid Assignment of RRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- Louca, S.; Parfrey, L.W.; Doebeli, M. Decoupling Function and Taxonomy in the Global Ocean Microbiome. Science 2016, 353, 1272–1277. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Cao, B.; Chen, Z.; Xu, K. Grafting Enhances the Photosynthesis and Nitrogen Absorption of Tomato Plants Under Low-Nitrogen Stress. J. Plant Growth Regul. 2022, 41, 1714–1725. [Google Scholar] [CrossRef]
- Hu, X.; Wang, X.; Abbas, T.; Fang, T.; Miao, D.; Li, Y.; Chang, S.X.; Li, Y. Higher Ammonium-to-Nitrate Ratio Shapes Distinct Soil Nitrifying Community and Favors the Growth of Moso Bamboo in Contrast to Broadleaf Tree Species. Biol. Fertil. Soils 2021, 57, 1171–1182. [Google Scholar] [CrossRef]
- Pontiggia, D.; Benedetti, M.; Costantini, S.; De Lorenzo, G.; Cervone, F. Dampening the DAMPs: How Plants Maintain the Homeostasis of Cell Wall Molecular Patterns and Avoid Hyper-Immunity. Front. Plant Sci. 2020, 11, 613259. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Jin, X.; Jiang, M.; Lü, X.; Lou, Y. Helophytes Adapt to Water and N-Enrichment Stresses by Adjusting and Coordinating Stoichiometry Characteristics in Main Organs. Sci. Total Environ. 2023, 898, 165538. [Google Scholar] [CrossRef]
- Fang, J.; Zhao, R.; Cao, Q.; Quan, Q.; Sun, R.; Liu, J. Effects of Emergent Aquatic Plants on Nitrogen Transformation Processes and Related Microorganisms in a Constructed Wetland in Northern China. Plant Soil 2019, 443, 473–492. [Google Scholar] [CrossRef]
- Liu, Y.; Shen, J.; Zhang, C.; Chen, Z. Impact of Rubber-Based Land Use Changes on Soil Properties and Carbon Pools: A Meta-Analysis. Catena 2023, 227, 107121. [Google Scholar] [CrossRef]
- Knops, J.M.H.; Bradley, K.L.; Wedin, D.A. Mechanisms of Plant Species Impacts on Ecosystem Nitrogen Cycling. Ecol. Lett. 2002, 5, 454–466. [Google Scholar] [CrossRef]
- Vymazal, J. Plants in Constructed, Restored and Created Wetlands. Ecol. Eng. 2013, 61, 501–504. [Google Scholar] [CrossRef]
- Gutiérrez-Girón, A.; Rubio, A.; Gavilán, R.G. Temporal Variation in Microbial and Plant Biomass during Summer in a Mediterranean High-Mountain Dry Grassland. Plant Soil 2014, 374, 803–813. [Google Scholar] [CrossRef]
- Fierer, N.; Walsh, C.M. Can We Manipulate the Soil Microbiome to Promote Carbon Sequestration in Croplands? PLoS Biol. 2023, 21, e3002207. [Google Scholar] [CrossRef]
- Lehmann, J.; Kleber, M. The Contentious Nature of Soil Organic Matter. Nature 2015, 528, 60–68. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Fang, J.; Li, X.; Li, C.; Zhao, Y.; Liu, J. Microorganisms Directly Affected Sediment Carbon–Nitrogen Coupling in Two Constructed Wetlands. Water 2024, 16, 1550. https://doi.org/10.3390/w16111550
Wang Y, Fang J, Li X, Li C, Zhao Y, Liu J. Microorganisms Directly Affected Sediment Carbon–Nitrogen Coupling in Two Constructed Wetlands. Water. 2024; 16(11):1550. https://doi.org/10.3390/w16111550
Chicago/Turabian StyleWang, Yan, Jiaohui Fang, Xin Li, Changchao Li, Yongkang Zhao, and Jian Liu. 2024. "Microorganisms Directly Affected Sediment Carbon–Nitrogen Coupling in Two Constructed Wetlands" Water 16, no. 11: 1550. https://doi.org/10.3390/w16111550