
Temperature-Dependent Mixotrophy in Natural Populations of the Toxic Dinoflagellate Karenia brevis

Abstract
:1. Introduction
2. Materials and Methods
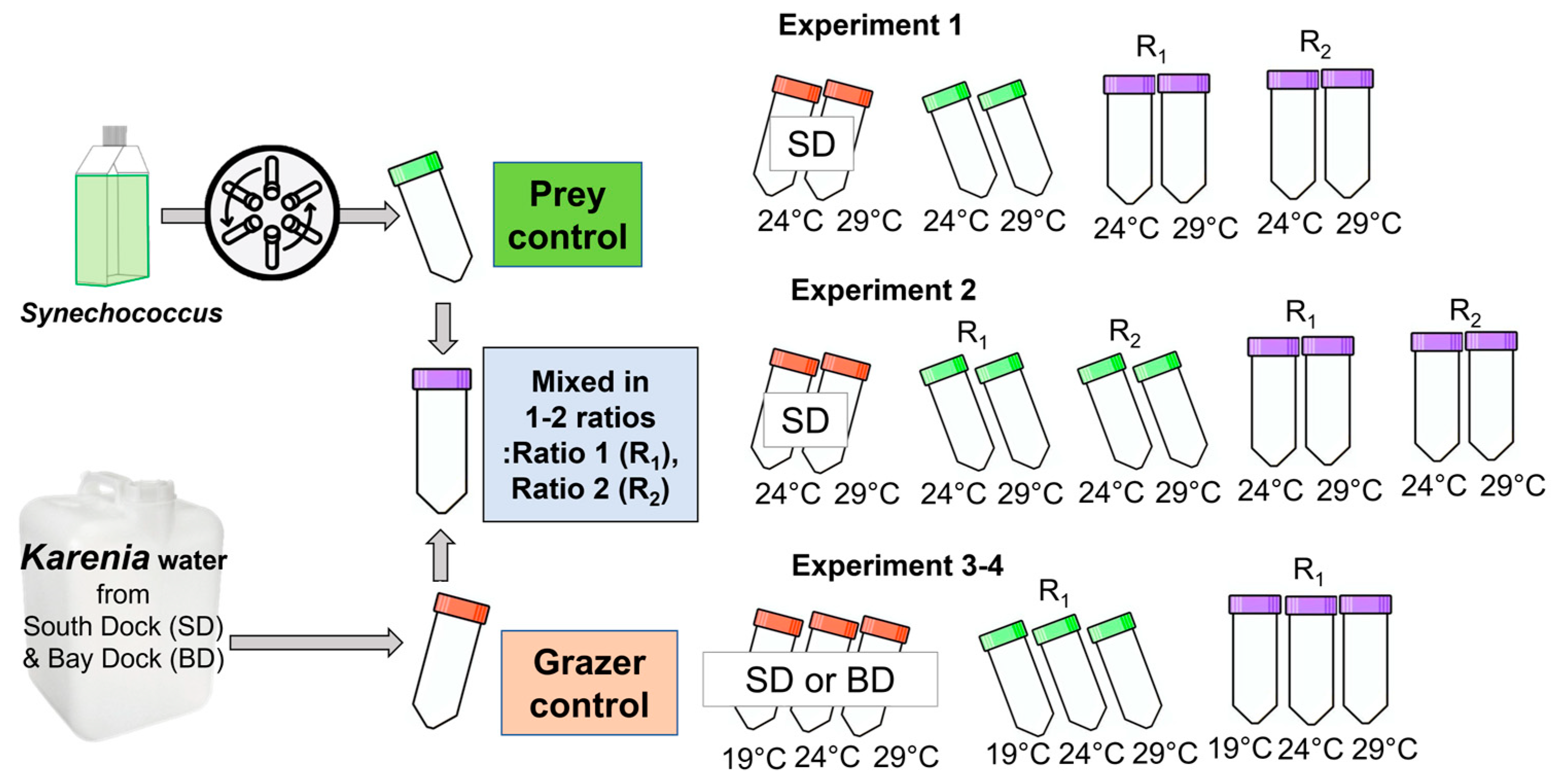
2.1. Field Sampling
2.2. Prey Culture
2.3. Short-Term Ingestion Experiments
2.4. Enumeration of Grazer and Prey and Rate Calculations
2.5. Pulse-Amplitude-Modulated (PAM) Fluorometry
2.6. Statistical Analysis
3. Results
3.1. Environmental Conditions
3.2. Synechococcus Cell Concentration Changes
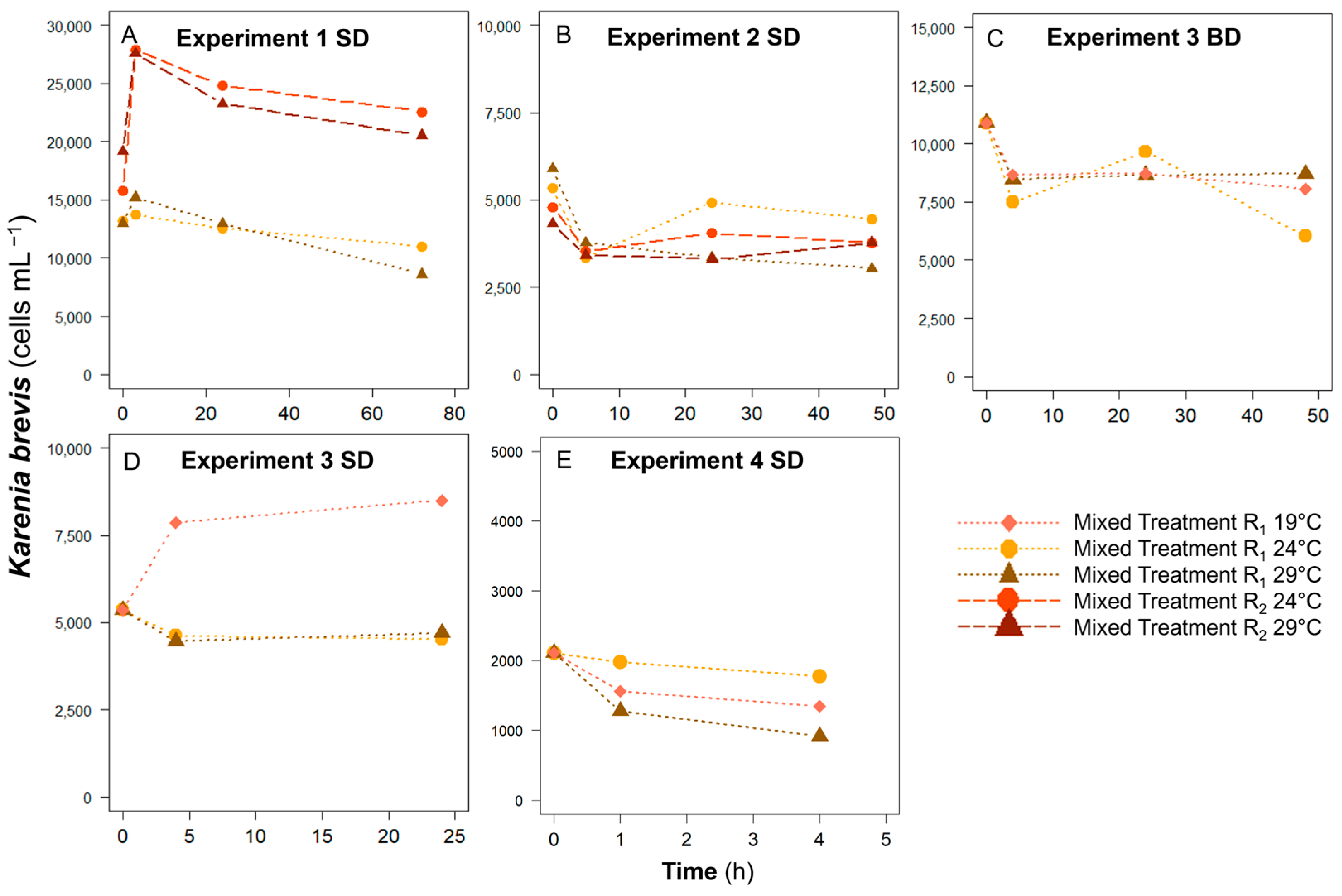
3.3. Karenia brevis Cell Concentration Changes
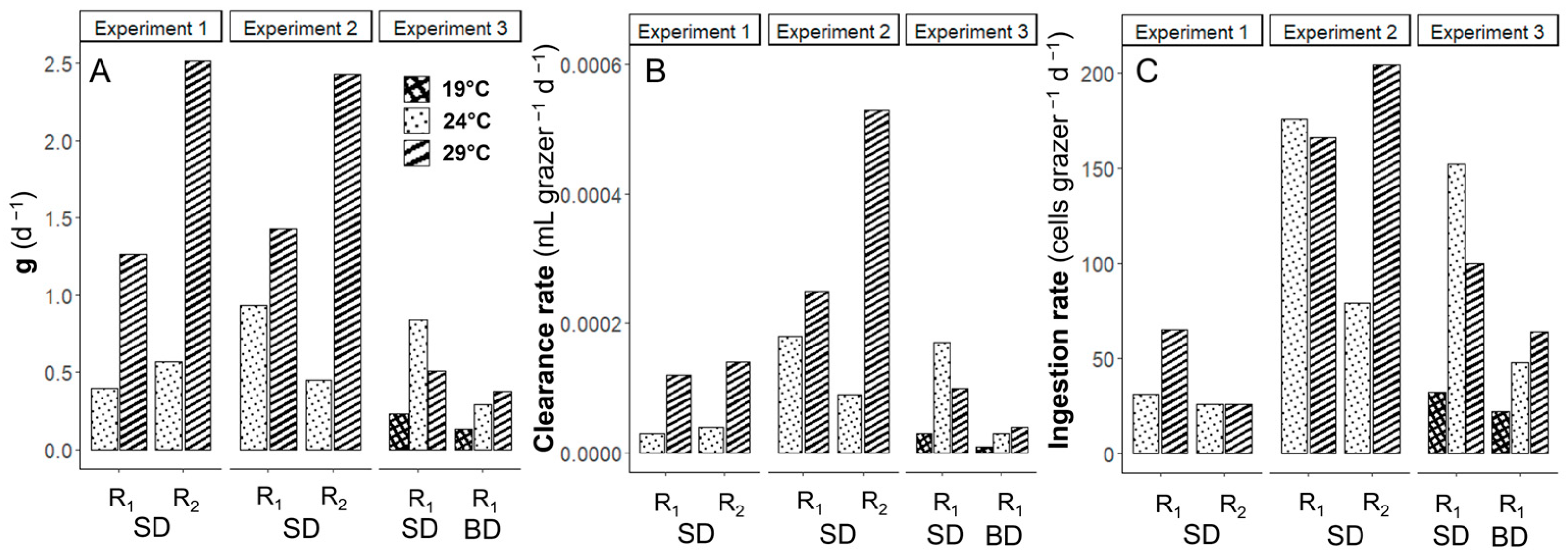
3.4. Temperature Effects on Grazing of Karenia brevis on Synechococcus
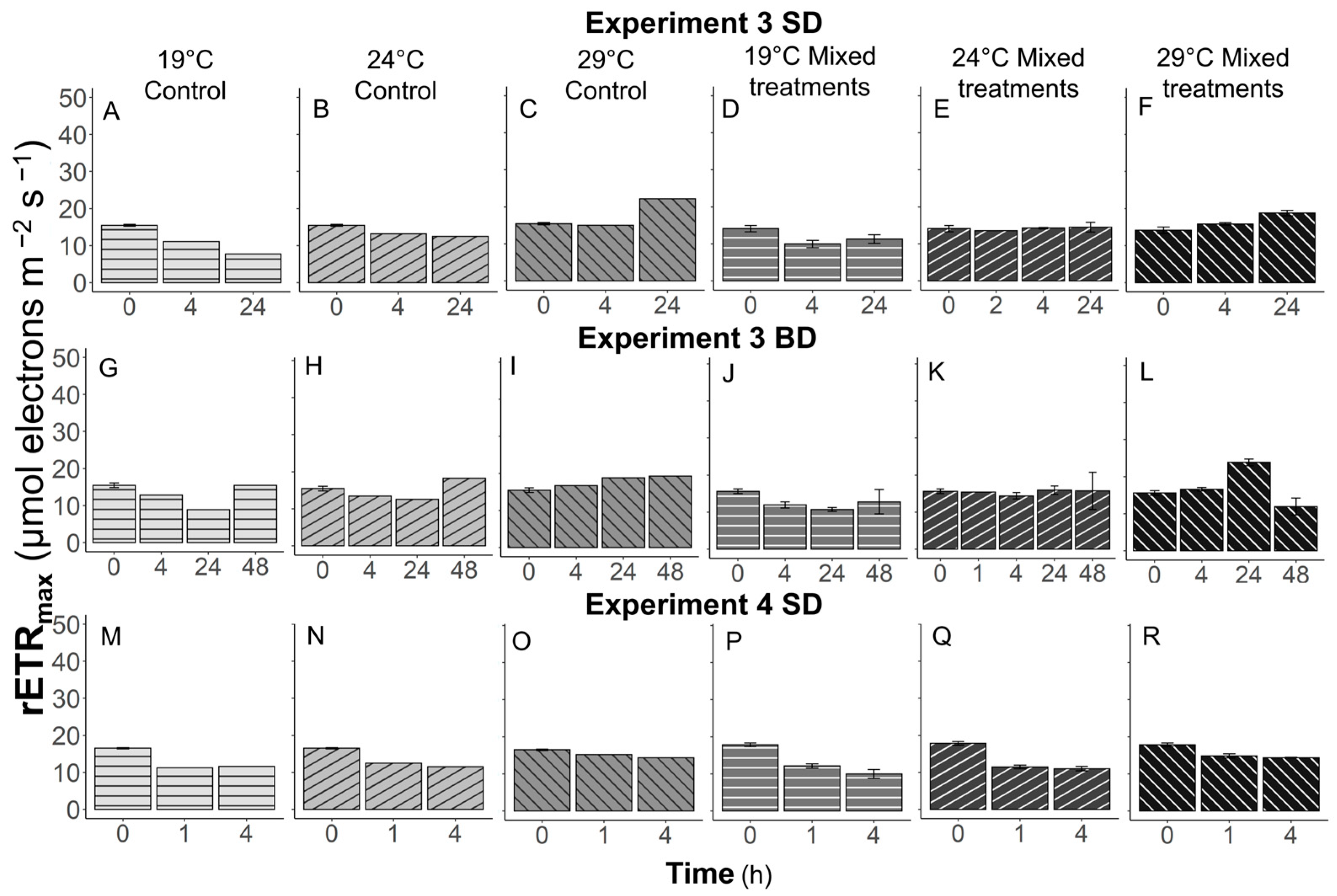
3.5. Temperature Effects on Photosynthetic Performance
4. Discussion
4.1. Grazing on Synechococcus by Natural Karenia brevis Population
4.2. Warming Shifts Karenia brevis towards More Phago-Heterotrophic in a Short-Term
4.3. The Potential Role of Karenia brevis Grazing in the Surface Layer on the West Florida Shelf
5. Conclusions and Summary
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brown, J.H.; Gillooly, J.F.; Allen, A.P.; Savage, V.M.; West, G.B. Toward a metabolic theory of ecology. Ecology 2004, 85, 1771–1789. [Google Scholar] [CrossRef]
- Allen, A.P.; Gillooly, J.F.; Brown, J.H. Linking the global carbon cycle to individual metabolism. Funct. Ecol. 2005, 19, 202–213. [Google Scholar] [CrossRef]
- Wilken, S.; Huisman, J.; Naus-Wiezer, S.; van Donk, E. Mixotrophic organisms become more heterotrophic with rising temperature. Ecol. Lett. 2013, 16, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Ng, H.Y.-T.; Zhang, S.; Liu, H. Effects of temperature on a mixotrophic dinoflagellate (Lepidodinium sp.) under different nutritional strategies. Mar. Ecol. Prog. Ser. 2021, 678, 37–49. [Google Scholar] [CrossRef]
- You, J.H.; Jeong, H.J.; Lim, A.S.; Ok, J.H.; Kang, H.C. Effects of irradiance and temperature on the growth and feeding of the obligate mixotrophic dinoflagellate Gymnodinium smaydae. Mar. Biol. 2020, 167, 64. [Google Scholar] [CrossRef]
- Glibert, P.M.; Mitra, A. From webs, loops, shunts, and pumps to microbial multitasking: Evolving concepts of marine microbial ecology, the mixoplankton paradigm, and implications for a future ocean. Limnol. Oceanogr. 2022, 67, 585–597. [Google Scholar] [CrossRef]
- Brand, L.E.; Campbell, L.; Bresnan, E. Karenia: The biology and ecology of toxic genus. Harmful Algae 2012, 14, 156–178. [Google Scholar] [CrossRef] [PubMed]
- Brand, L.E.; Compton, A. Long-term increase in Karenia brevis abundance along the Southwest Florida coast. Harmful Algae 2007, 6, 232–252. [Google Scholar] [CrossRef] [PubMed]
- Heil, C.A.; Amin, S.A.; Glibert, P.M.; Hubbard, K.A.; Li, M.; Martínez, J.M.; Weisberg, R.; Liu, Y.; Sun, Y. Termination patterns of Karenia brevis blooms in the eastern Gulf of Mexico. In Proceedings of the 19th International Conference on Harmful Algae, La Paz, Mexico, 10–15 October 2022. [Google Scholar] [CrossRef]
- Heil, C.A.; Bronk, D.A.; Mulholland, M.R.; O’Neil, J.M.; Bernhardt, P.W.; Mursako, S.; Havens, J.A.; Vargo, G.A. Influence of daylight surface aggregation behavior on nutrient cycling during a Karenia brevis (Davis) G. Hansen & Ø. Moestrup bloom: Migration to the surface as a nutrient acquisition strategy. Harmful Algae 2014, 38, 86–94. [Google Scholar] [CrossRef]
- Weisberg, R.H.; Zheng, L.; Liu, Y.; Lembke, C.; Lenes, J.M.; Walsh, J.J. Why no red tide was observed on the West Florida Continental Shelf in 2010. Harmful Algae 2014, 28, 119–126. [Google Scholar] [CrossRef]
- Jeong, H.J.; Park, J.Y.; Nho, J.H.; Park, M.O.; Ha, J.H.; Seong, K.A.; Jeng, C.; Seong, C.N.; Lee, K.Y.; Yih, W.H. Feeding by red-tide dinoflagellates on the cyanobacterium Synechococcus. Aquat. Microb. Ecol. 2005, 41, 131–143. [Google Scholar] [CrossRef]
- Glibert, P.M.; Burkholder, J.M.; Kana, T.M.; Alexander, J.; Skelton, H.; Shilling, C. Grazing by Karenia brevis on Synechococcus enhances its growth rate and may help to sustain blooms. Aquat. Microb. Ecol. 2009, 55, 17–30. [Google Scholar] [CrossRef]
- Ahn, S.H.; Mayali, X.; Weber, P.K.; Glibert, P.M. Grazing by the toxigenic dinoflagellate Karenia brevis on Synechococcus and Rhodomonas: Effects of food quantity and quality and photophysiological responses of predator and prey. Limnol. Oceanogr. 2024; to be submitted. [Google Scholar]
- Heil, C.A.; Revilla, M.; Glibert, P.M.; Murasko, S. Nutrient quality drives differential phytoplankton community composition on the southwest Florida Shelf. Limnol. Oceanogr. 2007, 52, 1067–1078. [Google Scholar] [CrossRef]
- Sipler, R.E.; Bronk, D.A.; Seitzinger, S.P.; Lauck, R.; McGuinness, L.R.; Kirkpatrick, G.J.; Heil, C.A.; Kerkhof, L.J.; Schofield, O.M. Trichodesmium-derived dissolved organic matter is a source of nitrogen capable of supporting the growth of toxic red tide Karenia brevis. Mar. Ecol. Prog. Ser. 2013, 483, 31–45. [Google Scholar] [CrossRef]
- Zhang, Q.; Yu, R.; Song, J.; Yan, T.; Wang, Y.; Zhou, M. Will harmful dinoflagellate Karenia mikimotoi grow phagotrophically? Chin. J. Oceanol. Limnol. 2011, 29, 849–859. [Google Scholar] [CrossRef]
- Zhang, Q.; Song, J.; Yu, R.; Yan, T.; Wang, Y.; Kong, F.; Zhou, M. Roles of mixotrophy in blooms of different dinoflagellates: Implications from the growth experiment. Harmful Algae 2013, 30, 10–26. [Google Scholar] [CrossRef]
- Ok, J.H.; Jeong, H.J.; Lim, A.S.; Kang, H.C.; You, J.H.; Park, S.A.; Eom, S.H. Lack of mixotrophy in three Karenia species and the prey spectrum of Karenia mikimotoi (Gymnodiniales, Dinophyceae). Algae 2023, 38, 39–55. [Google Scholar] [CrossRef]
- Guillard, R.R.L.; Hargraves, P.E. Stichochrysis immobilis is a diatom, not a chrysophyte. Phycologia 1993, 32, 234–236. [Google Scholar] [CrossRef]
- Heinbokel, J.F. Studies on the functional role of Tintinnids in the Southern California Bight. I. Grazing and growth rates in laboratory cultures. Mar. Biol. 1978, 47, 177–189. [Google Scholar] [CrossRef]
- Frost, B.W. Effects of size and concentration of food particles on the feeding behavior of the marine planktonic copepod. Limnol. Oceanogr. 1972, 17, 805–815. [Google Scholar] [CrossRef]
- Platt, T.; Gallegos, C.L.; Harrison, W.G. Photoinhiition of photosynthesis in natural assemblages of marine phytoplankton. J. Mar. Res. 1980, 38, 687–701. [Google Scholar]
- Pécseli, H.L.; Truslen, J.K.; Fiksen, Ø. Predator-prey encounter and capture rates in turbulent environments. Limnol. Oceanogr. 2014, 4, 85–105. [Google Scholar] [CrossRef]
- Liu, K.; Ng, H.Y.-T.; Gao, Z.; Liu, H. Selective feeding of a mixotrophic dinoflagellate (Lepidodinium sp.) in response to experimental warming and inorganic nutrient imbalance. Front. Microbiol. 2022, 13, 805306. [Google Scholar] [CrossRef] [PubMed]
- Lim, A.S.; Jeong, H.J.; Ok, J.H.; You, J.H.; Kang, H.C.; Kim, S.J. Effects of light intensity and temperature on growth and ingestion rates of the mixotrophic dinoflagellate Alexandrium pohangense. Mar. Biol. 2019, 166, 98. [Google Scholar] [CrossRef]
- Lepori-Bui, M.; Paight, C.; Eberhard, E.; Mertz, C.M.; Moeller, H.V. Evidence for evolutionary adaptation of mixotrophic nanoflagellates to warmer temperatures. Glob. Chang. Biol. 2022, 28, 7094–7107. [Google Scholar] [CrossRef] [PubMed]
- Paul, J.H.; Alfreider, A.; Kang, J.B.; Stokes, R.A.; Griffin, D.; Campbell, L.; Ornolfsdottir, E. Form IA rbcL transcript associated with a low salinity/high chlorophyll plume (‘Green River’) in the eastern Gulf of Mexico. Mar. Ecol. Prog. Ser. 2000, 198, 1–8. [Google Scholar] [CrossRef]
- Wawrik, B.; Callaghan, A.V.; Bronk, D.A. Use of inorganic and organic nitrogen by Synechococcus spp. and diatoms on the West Florida Shelf as measured using stable isotope probing. Appl. Environ. Microb. 2009, 75, 6662–6670. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| South Dock | Bay Dock | South Dock | |||
|---|---|---|---|---|---|
| Experiment 1 | Experiment 2 | Experiment 3 | Experiment 3 | Experiment 4 | |
| 28 November | 29 November | 30 November | 30 November | 2 December | |
| Temperature (°C) | 24.0 | 22.5 | 23.5 | 23.9 | 22.0 |
| Nitrate + Nitrite (µM) | 0.53 | 0.25 | <0.05 | 0.19 | 0.18 |
| Ammonium (µM) | 3.77 | 0.59 | 2.41 | 0.53 | <0.04 |
| Phosphate (µM) | 0.21 | 0.12 | 0.10 | 0.25 | 0.10 |
| Exp | C0:P0 (vol:vol) | Temp | C0 | P0 | C0:P0 (Cell:Cell) | Hourly g | Hourly CR | Hourly IR | Daily g | Daily CR | Daily IR |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 3:1 | 24 °C | 1.2 | 13.2 | 90:1 | 0.10 | 0.000007 | 7 | 0.40 | 0.00003 | 31 |
| 29 °C | 1.4 | 13.0 | 106:1 | 0.04 | 0.000003 | 3 | 1.26 | 0.00012 | 65 | ||
| 5:4 | 24 °C | 0.9 | 15.8 | 63:1 | 0.11 | 0.000010 | 4 | 0.57 | 0.00004 | 26 | |
| 29 °C | 0.9 | 19.1 | 48:1 | 0 | 0 | 0 | 2.51 | 0.00014 | 26 | ||
| 2 | 2:3 | 24 °C | 1.3 | 5.3 | 243:1 | 0 | 0 | 0 | 0.93 | 0.00018 | 176 |
| 29 °C | 1.2 | 5.9 | 209:1 | 0.05 | 0.000010 | 12 | 1.43 | 0.00025 | 166 | ||
| 5:8 | 24 °C | 1.0 | 4.8 | 216:1 | 0 | 0 | 0 | 0.45 | 0.00009 | 79 | |
| 29 °C | 1.0 | 4.3 | 242:1 | 0 | 0 | 0 | 2.43 | 0.00053 | 204 | ||
| 3 | 1:4 | 19 °C | 1.1 | 5.4 | 208:1 | 0.06 | 0.000010 | 8 | 0.23 | 0.00003 | 32 |
| 24 °C | 1.1 | 5.4 | 208:1 | 0.11 | 0.000020 | 18 | 0.84 | 0.00017 | 152 | ||
| 29 °C | 1.1 | 5.4 | 208:1 | 0.01 | 0.000001 | 1 | 0.51 | 0.00010 | 100 | ||
| 1:1 * | 19 °C | 1.5 | 10.9 | 141:1 | 0 | 0 | 0 | 0.13 | 0.00001 | 22 | |
| 24 °C | 1.5 | 10.9 | 141:1 | 0.01 | 0.000001 | 2 | 0.29 | 0.00003 | 48 | ||
| 29 °C | 1.5 | 10.9 | 141:1 | 0 | 0 | 0 | 0.38 | 0.00004 | 64 | ||
| 4 | 1:4 | 19 °C | 0.9 | 2.1 | 440:1 | 0.01 | 0.000003 | 3 | |||
| 24 °C | 0.9 | 2.1 | 440:1 | 0.05 | 0.000028 | 26 | |||||
| 29 °C | 0.9 | 2.1 | 440:1 | 0.12 | 0.000073 | 67 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahn, S.H.; Glibert, P.M. Temperature-Dependent Mixotrophy in Natural Populations of the Toxic Dinoflagellate Karenia brevis. Water 2024, 16, 1555. https://doi.org/10.3390/w16111555
Ahn SH, Glibert PM. Temperature-Dependent Mixotrophy in Natural Populations of the Toxic Dinoflagellate Karenia brevis. Water. 2024; 16(11):1555. https://doi.org/10.3390/w16111555
Chicago/Turabian StyleAhn, So Hyun, and Patricia M. Glibert. 2024. "Temperature-Dependent Mixotrophy in Natural Populations of the Toxic Dinoflagellate Karenia brevis" Water 16, no. 11: 1555. https://doi.org/10.3390/w16111555