Abstract
A novel heterotrophic aerobic denitrifying Pseudomonas hunanensis strain DC-2 was screened from the sediments of Lake Dianchi and identified with high nitrification/denitrification ability. Within 30 h, the removal efficiency of ammonium-N and nitrate-N could reach 98.8% and 88.4%, respectively. The results of the single-factor experiments indicated that strain DC-2 exhibited excellent denitrification ability under the conditions of using sodium citrate as the nitrogen source, with an initial pH of 7, a C/N ratio of 10, and a temperature of 30 °C. Nitrogen balance experiments suggested that this strain removed N mainly via assimilation. Moreover, the N removal pathway was explored by genome and enzymatic assays, and a complex nitrogen metabolism pathway was established, including heterotrophic nitrification-aerobic denitrification (HN-AD), assimilatory reduction of nitrate (ANRA), and ammonia assimilation. Additionally, strain DC-2 was immobilized into particles for denitrification, demonstrating excellent efficacy in continuous total nitrogen removal (84.8% for TN). Hence, strain DC-2 demonstrated significant potential in treating real aquaculture wastewater.
1. Introduction
The rapidly developed aquaculture sector has played a crucial role in addressing worldwide food security challenges [1,2]. However, the excessive nitrogen produced by aquaculture operations (e.g., feces, residual feeds, and bacterial cell fragments) has a negative impact on the aquatic ecosystems. For example, the direct discharge of N-rich aquaculture wastewater into adjacent water bodies accelerates eutrophication, posing significant environmental, ecological, and economic risks [3]. Therefore, there is an urgent need to remove N from aquaculture system effectively and economically.
Microbial technology is increasingly employed in the treatment of wastewater from aquaculture. This approach is favored for its low operational costs, high efficiency in pollutant removal, and environmentally friendly attributes [4,5]. However, traditional nitrification and denitrification processes, which required separate operations, increase both the expenses incurred and the area required for wastewater treatment [6]. Recently, an increasing number of studies have focused on heterotrophic nitrification–aerobic denitrification (HN-AD) bacteria [7,8,9]. Compared to conventional denitrification technologies, the HN-AD technique utilized organic carbon, withstood high levels of dissolved oxygen (DO), and allowed a single heterotrophic microorganism to simultaneously perform nitrification and denitrification [10,11]. Previous studies have isolated several HN-AD bacteria from inland aquatic ecosystems and marine surroundings, like Pseudomonas sp. DM02 [7], Pseudomonas sp. N31942 [8], Pseudomonas sp. G16 [9], Halomonas venusta SND-01 [11], etc., and have been successfully applied in the denitrification of aquaculture wastewater. However, the majority of the research concentrated on isolating bacteria and assessing their nitrogen removal capabilities, with limited research on nitrogen metabolism pathways and functional gene expression during the HN-AD process.
HN-AD bacteria primarily remove nitrogen via a complete denitrification pathway involving NH4+-N → NH2OH → NO2−-N → NO3−-N → NO2−-N → NO → N2O/N2 [11]. However, low enzyme activity or genetic deletions can result in an incomplete denitrification pathway (NH4+-N → NH2OH → NO → N2O/N2) [12]. In addition, recent reports have indicated that a single strain can exhibit various nitrogen metabolic routes. For instance, researchers [13] identified the presence of three distinct nitrogen metabolism routes in strain Y-9 isolated from a long-term flooded paddy soil, including Nitrate assimilation, DNRA, and denitrification. These findings indicate a more varied network of pathways for nitrogen removal than previously recognized. Therefore, it is essential to investigate the denitrification properties of individual HN-AD strains and analyze their N metabolism pathways to deepen the theoretical understanding of the metabolic mechanisms of HN-AD strains.
In this investigation, a novel aerobic denitrifying strain was screened from the surface sediment of Lake Dianchi (Yunnan, China) and identified as Pseudomonas hunanensis DC-2. The nitrogen removal capacity of the strain was examined across both singular (NH4+-N, NO3−-N, NO2−-N) and mixed N sources. Multiple factors (carbon source, C/N ratio, pH, and temperature) were adjusted to identify the optimal nitrogen removal efficiency. Furthermore, the main focus was on the nitrogen removal mechanism, determined through nitrogen mass balance, genome, and enzyme activity analyses. Finally, the strain was immobilized onto bacterial particles to evaluate its performance and continuous suitability for treating actual aquaculture wastewater using the HN-AD bacteria. This study can not only clarify the denitrification mechanism of heterotrophic aerobic denitrifying Pseudomonas hunanensis strain DC-2 but also provide a new microbial resource to enhance the aquatic environment in freshwater aquaculture.
2. Materials and Methods
2.1. Isolation and Identification
High-efficiency denitrifying microorganisms were filtered from the sediment of Dianchi Lake (24°83′ N, 102°71′ E) under aerobic heterotrophic conditions. Surface sediments (0–10 cm) were collected from the lakebed using a Peterson grab sampler, quickly transported to the laboratory, and sieved through a 100-mesh screen to remove large gravel and debris. After settling, the sediments were stored in a temperature-controlled room (approximately 25 °C) for future use. The physicochemical properties of the sediments before the experiment were as follows: total nitrogen (TN) was 1.76 ± 0.16 mg/g, and total carbon (TC) was 97.36 ± 15.66 mg/g. The methods for isolation and purification were carried out as detailed by Qing et al. [6]. The shape of the bacterial cells was examined by scanning electron microscopy (SEM, S-3400N, Hitachi, Tokyo, Japan). The 16S rRNA gene of the strain was magnified utilizing the general primers 27F (AGAGTTTGATCCTGGCTCAG) and 1492R (GGTTACCTTGTTACGACTT). The amplification products underwent sequencing at Wuhan Aikangjian Biotechnology Co., Ltd. (Wuhan, China). The obtained nucleotide sequences were compared using the BLAST tool from NCBI (National Center for Biotechnology and Information) [11]. The phylogenetic tree of the novel strain was created through the Mega 6.0 software and the neighbor connection method.
2.2. The HN-AD Characterization of Strain DC-2
The bacterial suspension was inoculated at a 1% (v/v) ratio into heterotrophic nitrification medium (HM), nitrate-mediated aerobic denitrification medium (NAM), nitrite-mediated aerobic denitrification medium (NIM) and mixed nitrogen source medium (MM), respectively. The uninoculated media served as a control. The medium compositions are shown in the Supplementary Materials (see Table S1 in Supplementary Materials). The above media underwent sterilization at 121 °C for a duration of 30 min and sealed with a breathable membrane. Aerobic cultivation was carried out by continuous shaking at 160 r/min at 30 °C for 30 h. Every 6 h, samples were collected to measure the bacterial cell density (OD600), together with the concentrations of NH4+-N, NO3−-N, and NO2−-N. The measurements were performed three times to ensure accuracy and reliability.
2.3. Optimization of Factors Affecting NH4+-N Removal Ability
The effects of carbon (C) source, C/N ratio, pH, and temperature on the ammonia nitrogen metabolic ability of strain DC-2 were examined via single-factor tests. The C source options included sodium succinate hexahydrate, glucose, sodium citrate dihydrate, sodium acetate trihydrate, sucrose, and sodium tartrate. Different C/N ratios of 2, 5, 10, 15, 20, and 25 were tested. Various initial pH values of 4, 5, 6, 7, 8, 9, 10, and 11 were examined. Additionally, temperatures of 15 °C, 20 °C, 25 °C, 30 °C, 35 °C, and 40 °C were assessed. The bacterial suspension was introduced into ammonia nitrogen medium (HM) at a 1% (v/v) ratio. The concentrations of OD600, NO3−-N, NO2−-N, and NH4+-N were measured every 6 h. The experiments were replicated three times, with uninoculated media serving as the control.
2.4. Nitrogen Mass Balance
In order to determine the N mass balance of the strain, the bacterial suspension was inoculated at a 1% (v/v) ratio into three separate 500 mL sealed serum bottles. Each bottle contained a distinct medium of 100 mL (HM, NAM, NIM). Subsequently, the bottles underwent sequential oxygenation for ten minutes to purge gases from the medium and ensure the aerobic states in bottles. The serum bottles were subjected to incubation of 24 h at 30 °C, 160 r/min. The experiment involved collecting samples at the commencement and conclusion stages, followed by the determination of TN, DTN (dissolved total N), NH4+-N, NO3−-N, and NO2−-N concentrations. The biological-N content was determined by subtracting the unfiltered TN from DTN. The experiments were performed three times under sterile conditions. The uninoculated media were used as the controls.
2.5. The Whole Genome
To identify functional genes connected to N metabolism pathways, DNA extracted from the strain was dispatched to Bioyi Biotechnology Co., Ltd. Wuhan, China for whole-genome sequencing. Single-molecule sequencing of the DNA was performed using the Oxford Nanopore Technology sequencer PromethION. Functional prediction and gene annotation were executed using databases such as COG, KEGG, RefSeq, Pfam, SwissProt, GO, and TIGRFAMs. Subsequently, the fully assembled genome of the strain was submitted to the NCBI database.
2.6. The Activity of Nitrogen-Metabolizing Enzymes
A 1% (v/v) ratio of the bacterial suspension was added to 100 mL of AM and cultivated at 160 r/min, 30 °C for a duration of 24 h. Crude enzyme extracts devoid of cells were prepared through ultrasonication (300 W, 3 min) for subsequent enzyme activity analysis [14]. Enzyme-linked immunosorbent assay (ELISA) was utilized to determine the activities of Hydroxylamine oxidase (HAO) and Ammonia monooxygenase (AMO), whereas the methods outlined in previous studies were employed to measure the activities of nitrate reductase (NR) and nitrite reductase (NIR) [15].
2.7. Application in Aquaculture Wastewater
Strain DC-2 was fixed through the following procedure: Initially, strain DC-2 was cultured in HM medium for 24 h. Then, a solution was prepared by dissolving 10 g of polyvinyl alcohol (PVA) and 0.5 g of sodium alginate (SA) in 90 mL of pure water. The solution was stirred thoroughly until entirely soluble, and then sterilized at 121 °C for 30 min in a high-pressure autoclave. After cooling, bacterial solution of strain DC-2 (10 mL, OD600 ≈ 1.80) was added and stirred thoroughly. This mixture was slowly dropped into a 2% CaCl2 solution to create the immobilized bacteria particles (Diameter: 2–3 mm) and then cross-linked with three freeze–thaw cycles between 4 °C and −20 °C to produce elastic hydrogel biocarriers [16]. Wastewater from a recirculating aquaculture system in Wuhan was gathered and passed through a 0.45 µm membrane for filtration before the use. Then 20% (w/v) immobilized bacterial beads were introduced to 100 mL of real aquaculture wastewater (adjust the value of C/N ratio to 10). The wastewater without bacterial beads was designated as the blank group. Each treatment was replicated three times and cultured at 30 °C and 160 r/min, with each 24 h period constituting a cycle. The experiment continued for three consecutive cycles. After each cycle, the wastewater in the Erlenmeyer flasks was entirely exchanged for new aquaculture wastewater and an equivalent amount of carbon source was added, and the immobilized spheres were subjected to three deionized water washes. After each cycle, measurements were taken for the concentrations of NH4+-N, NO2−-N, NO3−-N, and TN.
2.8. Analytical Method and Statistical Analysis
NO3−-N concentrations were analyzed by ion chromatography (ICS-5000, Thermos Fisher Scientific, Waltham, MA, USA), NO2−-N concentrations were analyzed by N-(1-naphthyl)-ethylene diamine photometry [17], and NH4+-N concentrations were determined using Nessler reagent photometric method [18]. The strain’s growth was indicated by determining OD600 using the UV spectrophotometer. The calculation method for the removal efficiency of ammonia nitrogen, nitrate nitrogen, and nitrite nitrogen refers to the article by Shun et al. [19]. The results are represented by the mean value ± standard deviation. Origin 2018 was used for date analysis and graph construction. Excel software was utilized for statistical analyses.
3. Results and Discussion
3.1. Isolation and Identification
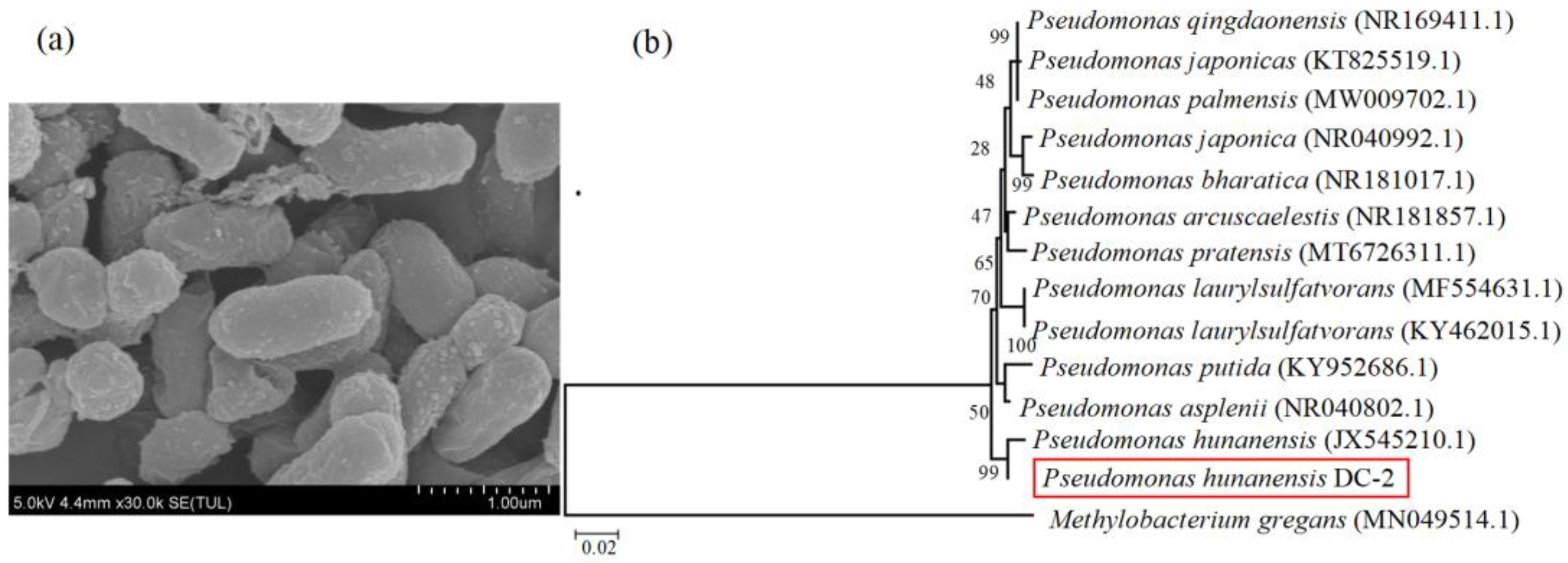
Scanning electron microscopy images (SEM) revealed that the strain exhibits a morphology of short rods with a length of less than 1 μm (Figure 1a). The 16S rRNA gene sequence (1391 bp) extracted from the acquired strain was sent for inclusion in the GenBank database (accession number: PP528522). Following the analysis of the partial sequence of the 16S rRNA gene from the strain, a BLAST homology search revealed that strain DC-2 exhibited the highest similarity (99%) with Pseudomonas hunanensis (JX545210.1). Consequently, the strain was designated as Pseudomonas hunanensis DC-2 (hereafter referred to as strain DC-2) (Figure 1b).
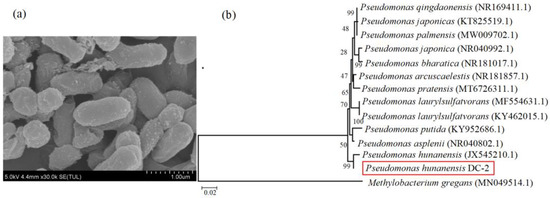
Figure 1.
The characteristics of strain DC-2: (a) scanning electron microscopy image, (b) the phylogenetic tree based on 16S rRNA gene sequences.
3.2. HN-AD Characteristics of Strain DC-2
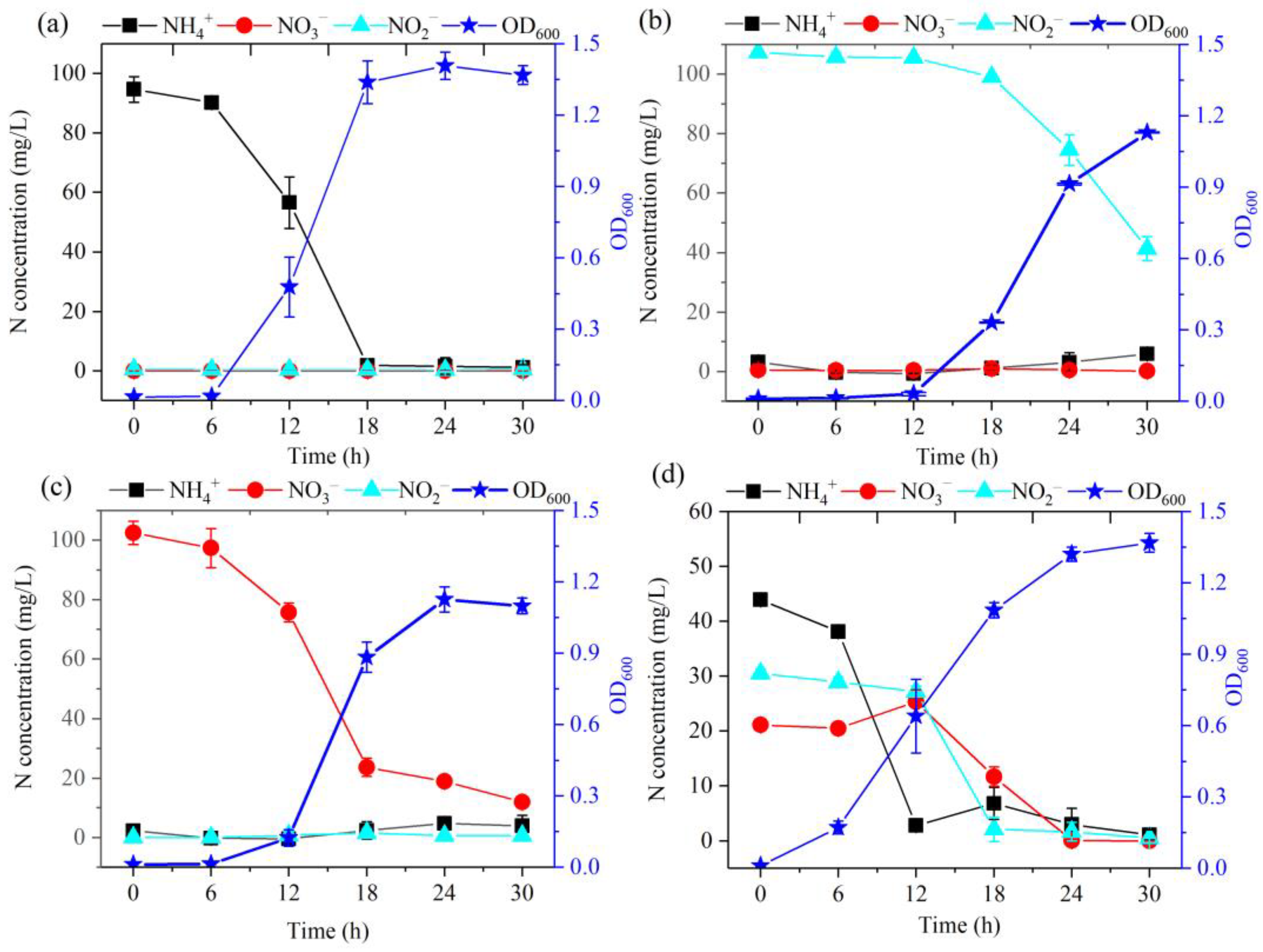
Strain DC-2 demonstrated the capability to remove ammonium-N within aerobic conditions (Figure 2a). The results demonstrated robust growth of strain DC-2 with ammonium-N serving as the unique N source. During the initial 16 h, the development of strain DC-2 was slow, with the OD600 gradually increasing from 0.01 to 0.02. The average removal rate of NH4+-N stood at 5.46 mg/L/h, slightly higher than those previously reported for HN-AD bacteria like strain TJPU04 (4.69 mg/L/h) [20]. At 30 h, the OD600 reached to 1.37, while the NH4+-N concentration dropped to 1.1 mg/L, achieving the removal efficiency of 98.8%. The inverse correlation between the concentration of ammonium-N and OD600 suggested that the degradation of ammonium-N is intimately linked to the growth of strain DC-2. This is in comparison to the use of nitrate-N and nitrite-N as the N sources (Figure 2b,c); strain DC-2 exhibited superior growth performance and the highest biomass, indicating that NH4+-N was a high-quality N source for strain DC-2. Throughout the process of removing ammonia-N, there was no observed amassment of nitrate-N or nitrite-N. This phenomenon may have been attributed to the direct assimilation of a portion of ammonia into biomass, a process characterized by the absence of energy consumption. Concurrently, a fraction of ammonia underwent incomplete denitrification reactions, resulting in its conversion into gaseous forms (NH4+ → NH2OH → NO → N2O/N2) [12], akin to observations in strain G16 [9]. Integrating subsequent genomic analysis findings, the annotation of nirBD genes within the genome of strain DC-2 not only catalyzed the reduction of nitrate nitrogen but also potentially regulated the conversion of nitrite nitrogen to nitric oxide [13]. The results indicate that strain DC-2 can effectively utilize ammonium.
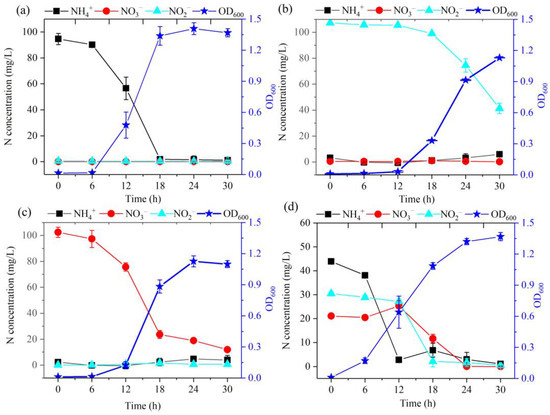
Figure 2.
HN-AD characteristics of strain DC-2 in different N sources: (a) NH4+-N (100 mg/L); (b) NO2−-N (100 mg/L); (c) NO3−-N (100 mg/L). (d) Mixed nitrogen sources: NH4+-N (50 mg/L), NO3−-N (30 mg/L), NO2−-N (20 mg/L).
When nitrite-N was employed as the initial N source, strain DC-2 had an adaptation time of 12 h (Figure 2b), which extended beyond when ammonia-N was used as the initial N source (less than 6 h). The extended adaptation period was attributed to the toxic effects of high concentrations of nitrite on microorganisms [21], necessitating a longer adaptation time for the strain to acclimate to the adverse conditions [17]. After 12 h, strain DC-2 began its exponential growth stage, reaching its maximum biomass (OD600 = 1.13) at 30 h. Meanwhile, the concentration of NO2−-N reduced from the initial 107.25 mg/L to 41.32 mg/L, achieving the removal rate of 61.5% within 30 h. With the degradation of nitrite, 5.96 mg/L of ammonia-N was accumulated at 30 h, which may have been caused by assimilation/dissimilation reduction. The process was similar to that of Rhodococcus erythropolis strain Y10 [17]. Hence, some of the degraded nitrites were assimilated into biomass through assimilation, while another portion was transformed into gaseous nitrogen through denitrification.
Strain DC-2 demonstrated its effectiveness in nitrate removal under aerobic conditions (Figure 2c). During the first 6 h, there was a gradual reduction in the level of nitrate nitrogen, declining from 102.52 mg/L to 97.41 mg/L. This decline might be attributed to the constrained utilization of biomass (OD600 < 0.01). Following a 12 h period of incubation, strain DC-2 transitioned into the exponential growth stage, achieving its peak OD600 value (1.13) while exhibiting a maximal N elimination rate of 8.68 mg/L/h spanning the period from the 12th to 18th hour and surpassing the rate reported for Klebsiella pneumonia CF-S9, which had a maximum removal rate about 6 mg/L/h [22]. The nitrate nitrogen level declined from 102.50 mg/L to 11.90 mg/L at 30 h, with a removal efficiency of 88.4%. At 30 h, ammonium was detected following the resolution of nitrate. Therefore, strain DC-2 demonstrates the capability of nitrate nitrogen reduction to ammonia nitrogen in aerobic conditions.
The use of the mixed N sources by strain DC-2 was depicted in Figure 2d. In the initial 6 h, the degradation rates for NH4+-N, NO3−-N, and NO2−-N were generally slow. After that, there was an acceleration in the reduction rate of NH4+-N, resulting in the removal of 93.2% of NH4+-N within 12 h. However, during hours 6–12, there was an observed increase in the concentration of nitrate from 20.50 mg/L to 25.40 mg/L, likely resulting from ammonium oxidation. After 12 h, there was a notable decrease in the concentrations of NO3−-N and NO2−-N, while the concentration of NH4+-N slightly increased in 18 h. The TN removal rate achieved 95.1% in 24 h. These results demonstrated that the aerobic denitrifying strain DC-2 preferentially consumed ammonium nitrogen, then used nitrate and nitrite for aerobic denitrification. The sequence of utilization of three forms of nitrogen by strain DC-2 was NH4+-N, NO3−-N, and NO2−-N. The results indicated that strain DC-2 possessed the ability to perform simultaneous nitrification and denitrification.
3.3. Effects of Environmental Factors on Nitrogen Removal Characteristic of Strain DC-2
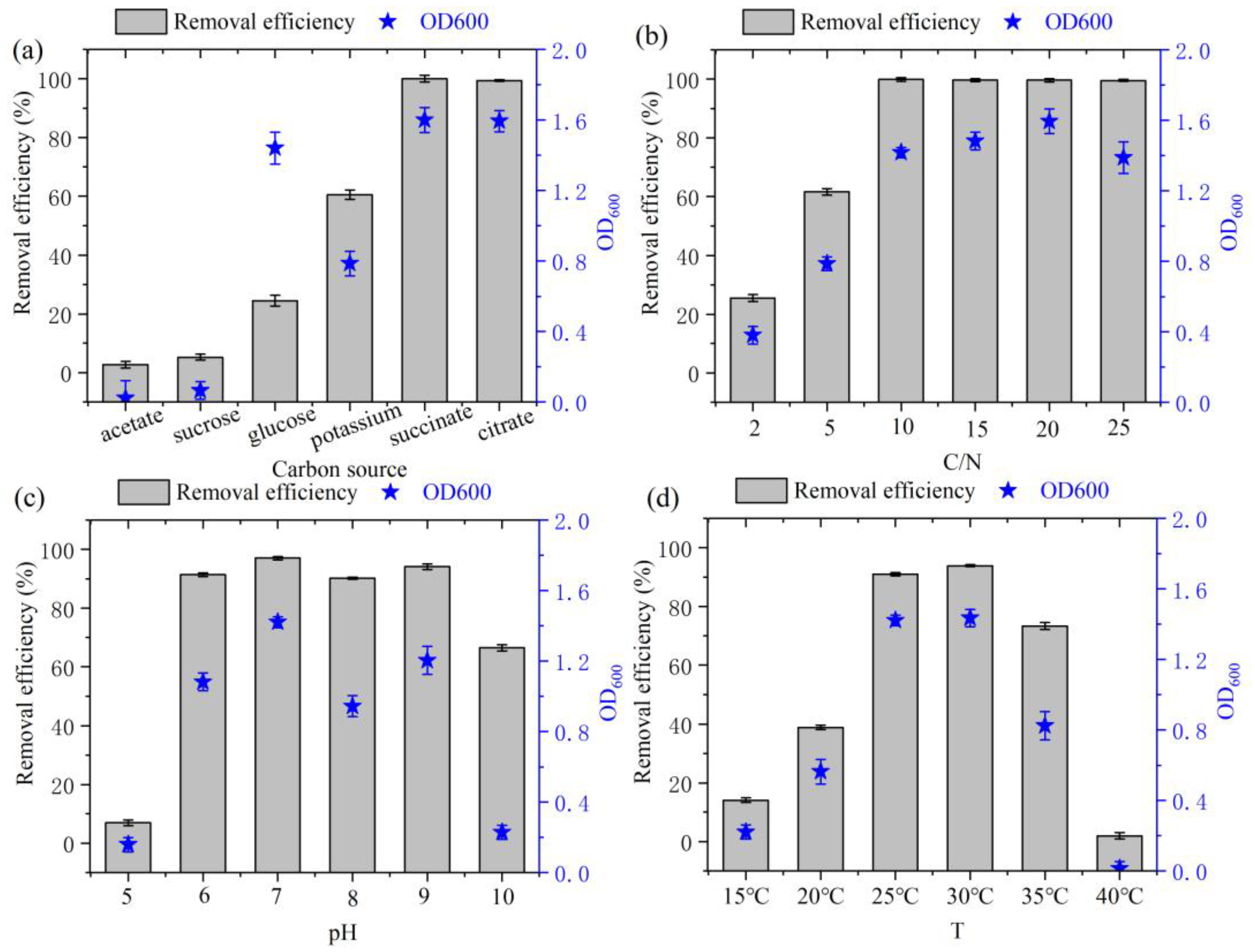
Organic carbon is often utilized as a nutrient source for bacterial propagation [23]. Media were prepared using glucose, sucrose, sodium acetate, sodium citrate, sodium succinate, and potassium sodium tartrate as unique C sources. As shown in Figure 3a, strain DC-2 was capable of utilizing these five carbon sources. Among these, the growth of the strain was notably better when sodium succinate and sodium citrate were used as the C source, resulting in a maximum nitrogen removal rate (>99%) and highest OD600 (1.60 and 1.59, respectively). The optimal carbon source identified in this study aligns with that for the Marinobacter strain NNA5 [24]. This suggested that HN-AD bacteria typically metabolized carboxylates like sodium citrate and sodium succinate due to their simpler chemical structures and lower molecular weights, facilitating absorption through the tricarboxylic acid cycle [9].
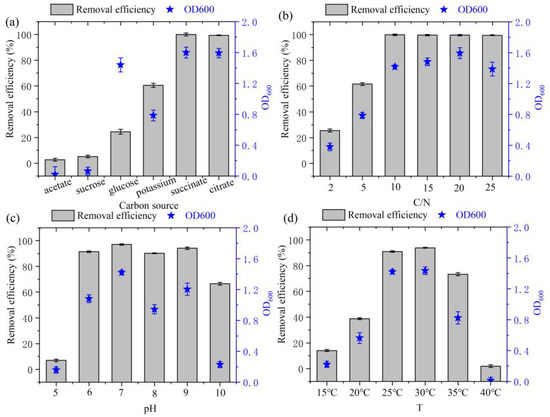
Figure 3.
Effects of various environmental conditions on the growth and N removal of strain DC-2: (a) C source; (b) C/N ratio; (c) initial pH; (d) temperature.
The HN-AD performance of heterotrophic bacteria is significantly affected by C/N ratio [25]. Both OD600 and the rate of ammonia nitrogen removal increased with lower C/N ratio (Figure 3b), indicating better denitrification performance when the C/N ratio exceeded 10 (with removal rates reaching 99%) and peaking at a C/N ratio of 20 with an OD600 of 1.59. However, despite the C/N ratio reaching 25, there was no improvement observed in the removal rate of ammonia nitrogen. Hence, taking cost into account, the most favorable C/N ratio was targeted to be 10. Several reports also indicated that strain Hydrocarbonoclasticus RAD-2 exhibited an optimal C/N ratio ranging from 5 to 15 [26], while for strain Halophilic Vibrio diabolicus SF16, the most favorable C/N ratio was reported to be 10 [27].
The growth and HN-AD performance of strain DC-2 was significantly affected by initial pH value (Figure 3c). Overall, the strain exhibited varying degrees of growth across the pH range from 5 to 10. The growth of strain DC-2 was sluggish when the pH < 5 or >10. However, it exhibited a rapid growth rate within the pH range of 6 to 9, suggesting its enhanced adaptability to moderately acidic and alkaline conditions. Presence of excessive H+ and OH− ions under acidic and alkaline pH circumstances may impact the enzymatic activity related to the denitrification process, sequentially affecting the rate of ammonia nitrogen removal [9]. Additionally, at a pH of 7.0, strain DC-2 demonstrated its peak biomass (OD600 = 1.42) and the highest NH4+-N removal rate (97.0%), mirroring the findings observed for strain Spongiibacter taiwanensis sp. nov [28].
Temperature is a primary limiting factor affecting the denitrification capabilities of most HN-AD strains [20]. The ammonia nitrogen removal rates of strain DC-2 varied significantly at different temperatures (p < 0.05, Figure 3d). At lower than 20 °C, the denitrification ability of strain DC-2 was inhibited, which was similar to the strain N31942 [8]. This is attributed to low temperatures inhibiting cell proliferation enzyme activity [29]. At temperatures between 25 °C and 30 °C, the NH4+-N removal efficiency of strain DC-2 exceeded 90%, underscoring its robust ammonia removal capacity under mesophilic conditions. Within 24 h, the NH4+-N removal efficiency attained the highest point (94.9%) under the optimal temperature conditions (30 °C).
Overall, strain DC-2 exhibited excellent denitrification performance under a broad range of conditions that included C/N ratio (10–20), carbon sources (such as sodium succinate and sodium citrate), pH (6–9), and temperatures (25–30 °C), demonstrating its high adaptability to different environmental factors.
3.4. Denitrification Pathway Analysis
3.4.1. Nitrogen Mass Balance Analysis
To further identify the pathways of N loss, N balance analysis was conducted using media with various initial N sources (Table 1). In the scenario where NH4+-N exclusively served as the N source, a substantial portion, representing 63.2% of the initial N, was converted into biomass N, while 30.4% transitioned into gaseous N. Moreover, during this process, no discernible accumulation of NO3−-N or NO2−-N was observed. Because of the fact that assimilation does not lead to N loss, ammonium-N may have been converted into gaseous N via dissimilation. Hence, the utilization of ammonium-N by strain DC-2 included assimilation and dissimilation pathways, with assimilation being predominant. This phenomenon agrees with strain Pseudomonas putida NP5 [30], while there is a discrepancy with strain Acinetobacter sp. ND7 [31], primarily attributed to the denitrification of bacteria. When nitrite-N was the unique N source, 42.1% of the nitrite underwent transformation into biomass N, and 9.2% was transformed into gaseous N, with a slight accumulation of ammonium-N and 34.4% of the nitrite-N remaining. Hence, it is inferred that strain DC-2 converts nitrite into ammonium, which is subsequently assimilated into biological N at a rapid rate. The remaining nitrite could be converted into gaseous N through the denitrification pathway. When nitrate-N was the sole N source, 55.3% of the nitrate-N was converted into biological N, and 28.6% ultimately transformed into gaseous N, with an accumulation of 13.8% ammonium-N in medium. Hence, this indicated that strain DC-2 primarily degraded nitrate through assimilation, and the accumulation of ammonium-N suggested that nitrate might have been converted to ammonium through either assimilation or dissimilation. When ammonia-N, nitrate-N, and nitrite-N served as N sources, strain DC-2 predominantly assimilated N (41.5–63.2%), while the remaining N was converted into gaseous forms via aerobic denitrification (9.2–30.4%). This behavior was similarly observed in Paracoccus strain AC-3 and strain Pseudomonas chengduensis BF6 [32,33].

Table 1.
Nitrogen(N) mass balance of strain DC-2 in different N sources in 24 h.
3.4.2. Whole Genome Analysis
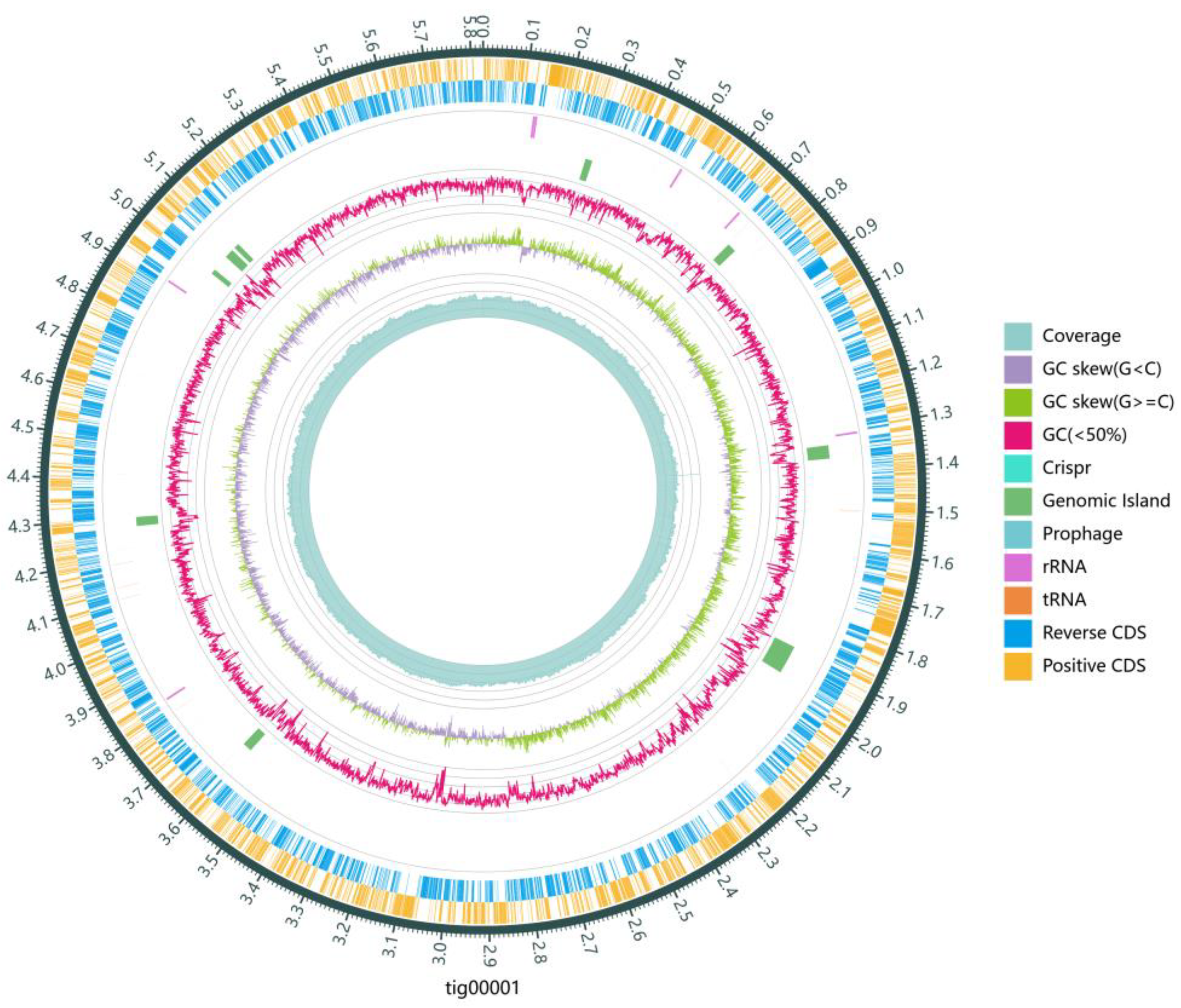
Further investigation into the N removal pathway involved sequencing, assembling, and annotating the genome of strain DC-2. The draft genome of strain DC-2 contained one circular chromosome (6,328,663 bp), as shown in Figure 4. This genome sequence length was comparable to that of most HNAD strains, including strain Achromobacter sp. HNDS-1 and Enterobacter sp. HNDS-6 [34]. Analysis of functional elements revealed the prediction of 11 genomic islands (GIs), one prophage, and two potential plasmid sequences within the DC-2 genome, though no CRISPR sequence structures were identified. The genomic data were submitted to GenBank (Accession number: PRJNA1092366).
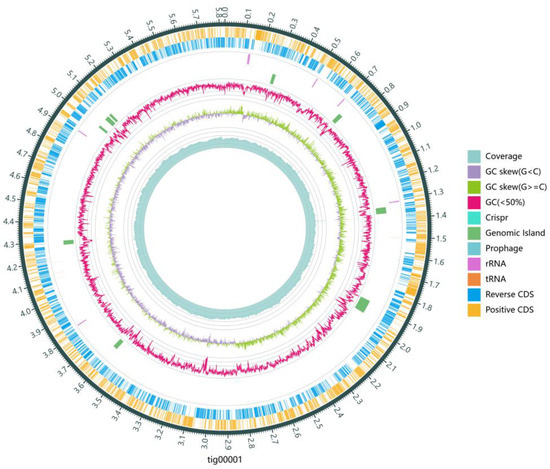
Figure 4.
Circular map of strain DC-2 genome. From outside to inside, there are Reverse CDS (marigold), Positive CDS (steel blue), tRNA (orange) and rRNA (purple), prphage (cadet blue), Genomic island (sea green), Crispr (light sea green), GC ratio (carmine means GC% < 50%), GC-SKEW (olive drab means G% ≥ C%, grayish purple means G% < C%) and coverage (cadet blue).
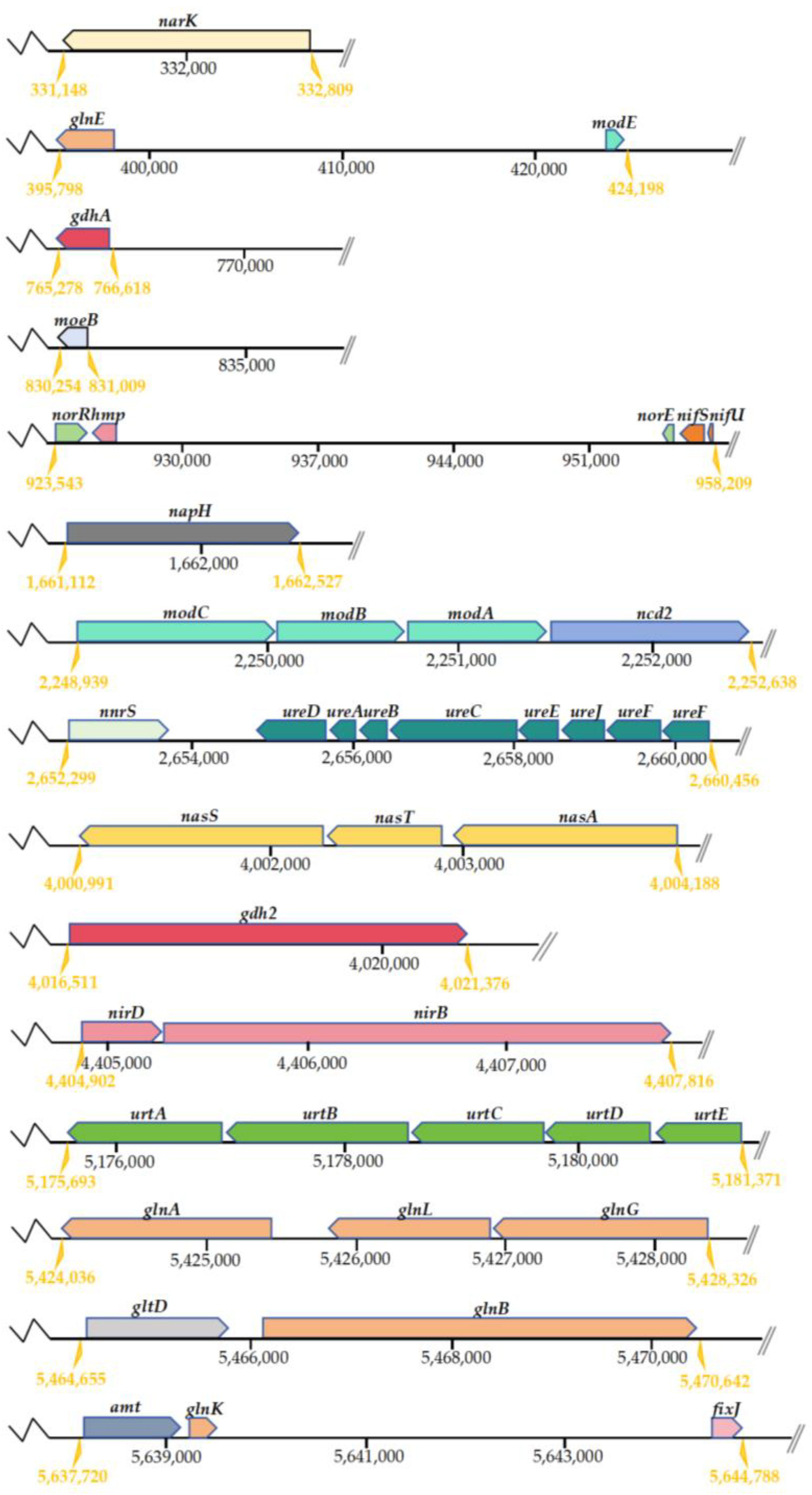
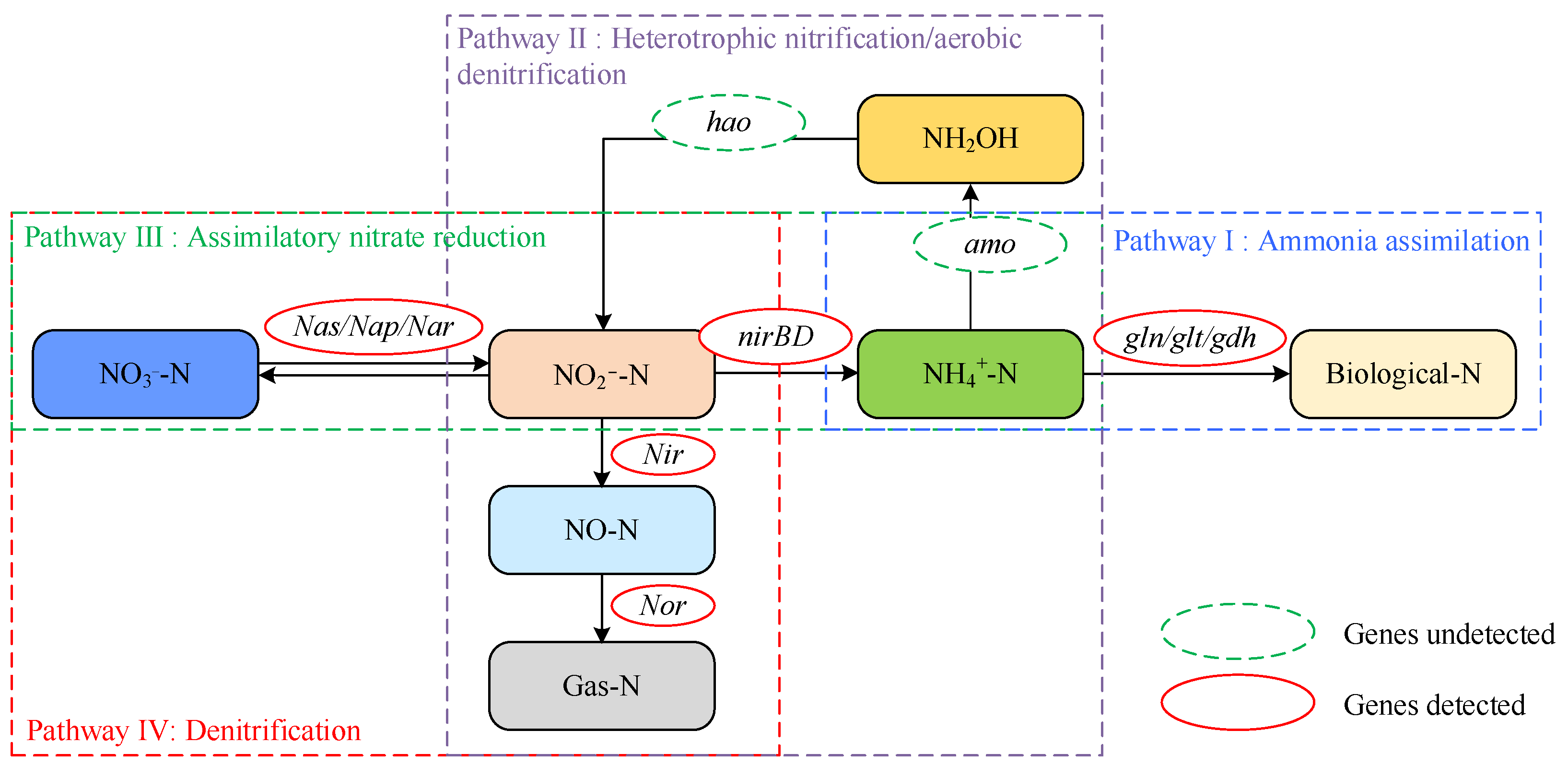
The investigation of genes related to N metabolism pathways was conducted using the KEGG Automatic Annotation Server (KAAS). Based on the predictions made by KAAS, the N metabolism pathways in strain DC-2 included ammonium assimilation, assimilatory nitrate reduction (ANRA), and heterotrophic nitrification–aerobic denitrification (HN-AD), with 64 relevant genes involved in the N metabolism pathways of strain DC-2 (Table S2 in Supplementary Materials). The positions of the key functional genes involved in nitrogen metabolism in strain DC-2 are shown in Figure 5. Among these, proteins encoded by narK, nrtC, and amt are responsible for transporting nitrate/nitrite, nitrite, and ammonium, respectively. These proteins facilitate the cellular transportation of inorganic N in strain DC-2.
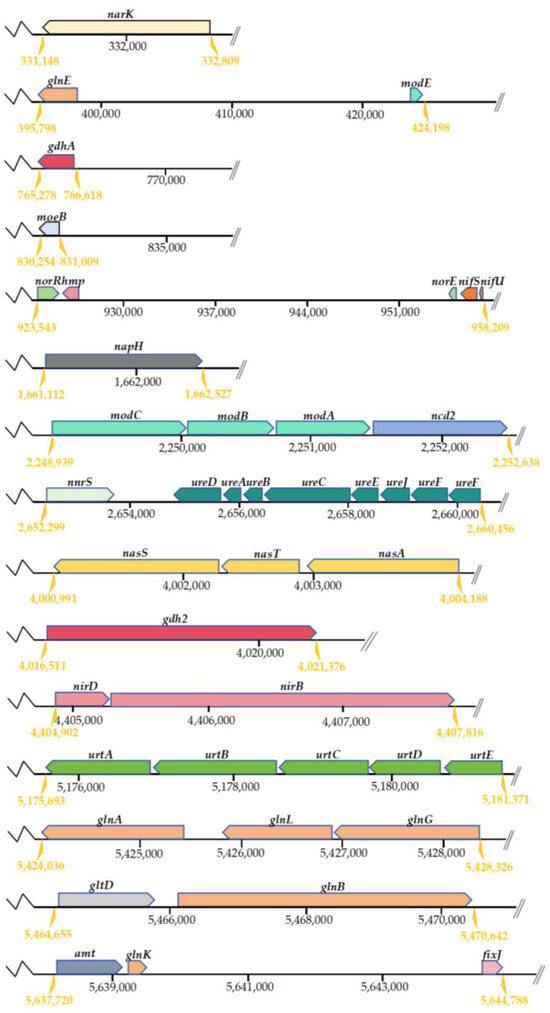
Figure 5.
Sequential arrangements of putative genes for nitrogen metabolic pathway on the genome of strain DC-2.
In the pathway of ammonium assimilation, we have identified genes associated with both the glutamate dehydrogenase pathway (gdh2, gdhA) and the glutamine synthetase–glutamate synthase pathway (gltBD and glnAEGKL). These genes are pivotal in facilitating the intracellular assimilation of ammonium [35]. Additionally, genes related to nitrogen fixation (nifU, nifS, vnfA) were annotated in strain DC-2. The annotation of the urtABCDE cluster and the ureABCDEFGJ cluster indicated the potential for strain DC-2 to utilize urea, revealing its potential for biological nitrogen fixation [36].
Approximately 35 genes were associated with the denitrification process. Among them, the gene napH encoded a periplasmic nitrate reductase, responsible for the transformation process of NO3−-N into NO2−-N. The NorR and norE genes participated in the transformation of NO to N2O. However, the genome of strain DC-2 lacked the key genes for Nir (nitric oxide formation) and Nos (nitrogen gas formation), resulting in an incomplete denitrification pathway being annotated. In the conventional denitrification pathway, the critical step of reducing NO2−-N to NO was mainly regulated by the genes encoding nitrite reductase nirK/nirS. Recent studies, however, have suggested that the operon nirBD could also be responsible for the conversion from NO2−-N into NO [37]. Thus, the nirBD genes in the genome of strain DC-2 not only significantly contribute to catalyzing the reduction of NO3−-N but may also regulate the conversion from NO2−-N to NO [13]. The denitrification reaction required the participation of the metal ion molybdenum, and genes related to molybdenum cofactor synthesis (moaBE, mobA, mocA, modABCE, moeAB) were annotated in strain DC-2. The moa and mob gene clusters perform a vital function in the generation of narJ. The gene hmp, encoding nitric oxide dioxygenase, which can convert NO to NO3−-N, might be a reason for the accumulation of NO3−-N. The nasA gene (encoding assimilatory nitrate reductase) and nirBD (encoding assimilatory nitrite oxidoreductase) formed the nas gene cluster, responsible for the nitrate assimilation/dissimilation process and aiding in aerobic denitrification [38]. Genes involved in denitrification regulation (anr, nnr, nnrS, rpoN) and in the expression of denitrification electrons (azu, cycA) were also annotated, further indicating the HN-AD capability of strain DC-2. Although KAAS did not map key genes involved in nitrification (amo, hao), the N balance mass analysis suggested that when ammonium nitrogen served as the N source, 30.42% of the N was converted into gaseous nitrogen. Assimilation typically does not cause nitrogen loss, thereby inferring nitrification occurred in the system. HN-AD strains commonly exhibit the absence of specific critical genes associated with N transformation pathways. For instance, critical pathway genes engaged in nitrification (HAO, AMO) and denitrification (Nos, Nir) were lacking in strain KSND [39]. HN-AD strains may possess novel N metabolism pathways and enzyme systems, but their mechanisms remain to be elucidated.
In general, different functional genes associated with diverse N metabolism pathways were identified in strain DC-2. According to the genes with functional roles, several N metabolism pathways were proposed, embracing ammonium assimilation, ANRA, and HN-AD.
3.4.3. Analysis of Enzyme Activity of N Metabolism
The enzymatic catalysis was essential for the biological removal of nitrogen compounds [40]. To further verify the proposed nitrogen metabolism pathways, the activities of four crucial enzymes associated with nitrogen removal (AMO, HAO, NR, and NIR) were measured (Table 2). Among these, AMO and HAO were key enzymes in the nitrification process [41], while NR and NIR were crucial in the denitrification process [42]. The results indicated that activities were detected for AMO, HAO, NR, and NIR, demonstrating the expression of these enzymes and indicating that strain DC-2 possessed a complete nitrification/denitrification pathway. In the whole genome of strain DC-2, genes related to amo were not found, yet the expression of AMO suggested that strain DC-2 might carry a novel type of AMO composed of subunits different from those of traditional AMO, responsible for the conversion of ammonia nitrogen to hydroxylamine [43]. The activity of HAO demonstrated the presence of an unknown enzyme capable of degrading hydroxylamine in this strain [44]. Based on these findings, the potential nitrogen removal pathway could be as follows: ammonium nitrogen might be catalyzed by AMO to convert into NH2OH, which was further oxidized to NO2−-N under the catalysis of HAO and eventually transformed into gaseous nitrogen through the denitrification process.
Table 2.
Key enzymes activities in HN-AD process.
3.4.4. Nitrogen Metabolic Pathway
Based on the research findings of N mass balance, genomic analysis, and enzyme activity analysis, a novel pathway for nitrogen metabolic was proposed, as shown in Figure 6. The KAAS localization results suggested that strain DC-2 can assimilate ammonium nitrogen into biomass (Pathway I: NH4+-N → Biomass Nitrogen). Nitrogen balance analysis indicated that when ammonium served as the initial N source, there was a N loss of 30.4 ± 4.1%. Combined with the activities of the AMO and HAO enzymes, this suggested another pathway for ammonium removal via HN-AD into gaseous N (Pathway II), with assimilation (59.1%) playing a main role, similar to strain SND-01 (65.3%) [11]. NO3−-N and NO2−-N could be reduced to ammonium via assimilatory reduction (Pathway III) or transformed into gaseous N through denitrification (Pathway IV), with assimilation also playing an important role in this pathway. Overall, multiple nitrogen metabolism pathways may be present in strain DC-2, which could be advantageous for treating wastewater with varying compositions of inorganic nitrogen.
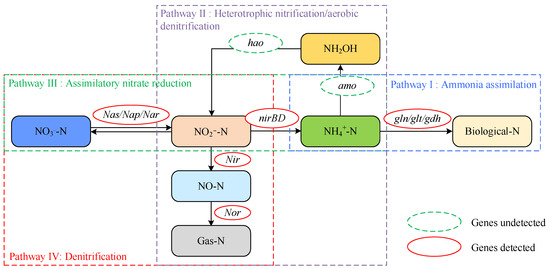
Figure 6.
The integrated N metabolic pathways of strain DC-2.
3.5. Application in Aquaculture Wastewater
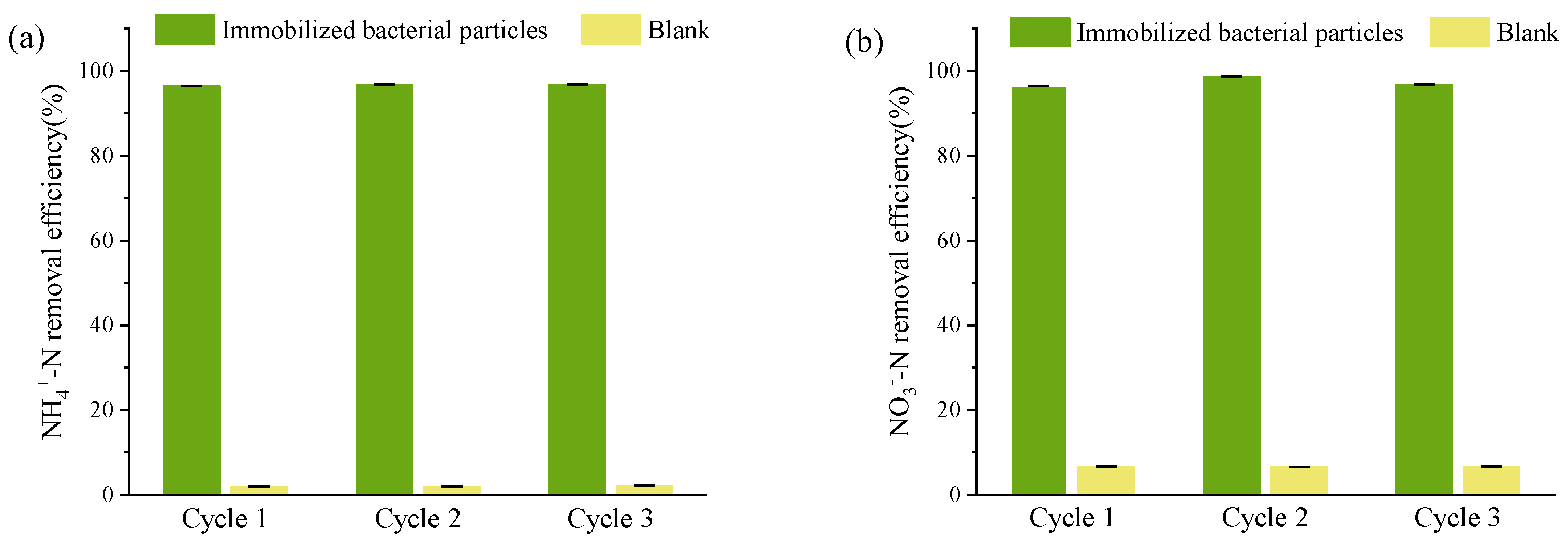
It was crucial to ensure strain DC-2’s sustainability and its ability to continuously treat wastewater [45]. Immobilized bacterial particles were utilized to evaluate the stability of strain DC-2 in treating real aquaculture wastewater. The aquaculture wastewater employed in the experimental setup featured NH4+-N and NO3−-N as the primary N pollutants, with concentrations of 2.90 ± 0.08 mg/L and 9.51 ± 0.52 mg/L, respectively. These constituted 19.1% and 62.6% of the total inorganic nitrogen content. Strain DC-2 was immobilized into bacterial particles, which exhibited excellent NH4+-N and NO3−-N removal rates (Figure 7); TN concentration decreased from the initial 15.23 ± 0.03 mg/L to 2.32 ± 0.44 mg/L, indicating a nitrogen elimination rate of 84.8 ± 2.9% in the system, similar to strain RAD-17 (84.2%) [46] and strain DM02 (88.2%) [7]. Across three cycles, there was no notable decline observed in the removal rates for NH4+-N (> 96%) and NO3−-N (>96%) (p > 0.05). There was an absence of NO2−-N accumulation throughout the duration of the study (with effluent nitrite nitrogen content averaging 0.10 ± 0.18 mg/L). Therefore, the application of strain DC-2 in treating real aquaculture wastewater holds significant research value.
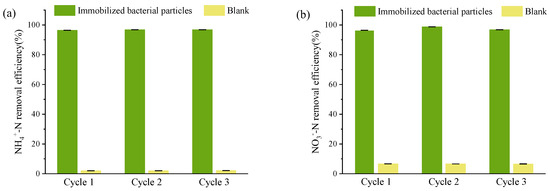
Figure 7.
Study on the continuous treatment of ammonium nitrogen and nitrate nitrogen in real aquaculture wastewater by bioaugmented immobilized strain DC-2: (a) NH4+-N; (b) NO3−-N.
4. Conclusions
A heterotrophic aerobic denitrifying Pseudomonas hunanensis strain DC-2 was isolated from the surface sediments of Lake Dianchi with substantial denitrification capability. It is capable of concurrently removing various nitrogen compounds without the accumulation of NO2−-N. The results of the single-factor experiments indicated that strain DC-2 exhibited excellent denitrification ability under the conditions of using sodium citrate as the nitrogen source, with an initial pH of 7, a C/N ratio of 10, and a temperature of 30 °C. According to N mass balance, whole-genome analysis, and key enzyme activity assessments, various metabolic pathways were hypothesized for strain DC-2, encompassing ammonium assimilation, ANRA, and HN-AD. Furthermore, strain DC-2 was immobilized into particles for denitrification, demonstrating effective N removal capabilities in real aquaculture wastewater, thus affirming its significant practical utility in aquaculture wastewater treatment.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/w16111625/s1, Table S1: Medium composition; Table S2: Putative genes related to nitrogen metabolism in Pseudomonas hunanensis DC-2 genome.
Author Contributions
Conceptualization, X.S. and C.T.; methodology, X.S., X.W., B.X., C.W. and C.T.; software, X.S. and C.T.; validation, C.T., X.W. and B.X.; formal analysis, X.S. and C.T.; investigation, X.S.; resources, X.W.; data curation, X.S., C.T. and X.W.; writing—original draft preparation, X.S.; writing—review and editing, C.T., X.W. and C.W.; visualization, X.S. and C.T.; supervision, B.X., X.W. and C.T.; project administration, C.T.; funding acquisition, X.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the China’s National Key R&D Programmes (grant number: 2022YFC3203601), the Key R&D plan of Hubei Province (grant number: 2022BCA068), and the Yunnan Province-Kunming City Major Science and Technology Project (grant number: 202202AH210006).
Data Availability Statement
The corresponding author holds the datasets produced and analyzed during the present study, which are accessible upon request.
Acknowledgments
We appreciate the assistance with experimental instruments provided by the Analytical and Testing Center of the Institute of Hydrobiology, Chinese Academy of Sciences.
Conflicts of Interest
The authors affirm that they do not possess any known conflicting financial concerns or personal associations that could have potentially influenced the reported work in this paper.
References
- Rajeev, R.; Adithya, K.K.; Kiran, G.S.; Selvin, J. Healthy microbiome: A key to successful and sustainable shrimp aquaculture. Rev. Aquac. 2021, 13, 238–258. [Google Scholar] [CrossRef]
- Xie, Y.; Tian, X.; He, Y.; Dong, S.; Zhao, K. Nitrogen removal capability and mechanism of a novel heterotrophic nitrification-aerobic denitrification bacterium Halomonas sp. DN3. Bioresour. Technol. 2023, 387, 129596. [Google Scholar] [CrossRef]
- Lu, Z.; Cheng, X.; Xie, J.; Li, Z.; Li, X.; Jiang, X.; Zhu, D. Iron-based multi-carbon composite and Pseudomonas furukawaii ZS1 co-affect nitrogen removal, microbial community dynamics and metabolism pathways in low-temperature aquaculture wastewater. J. Environ. Manag. 2024, 349, 119471. [Google Scholar] [CrossRef]
- Shao, Y.; Zhong, H.; Mao, X.; Zhang, H. Biochar-immobilized Sphingomonas sp. and Acinetobacter sp. isolates to enhance nutrient removal: Potential application in crab aquaculture. Aquac. Environ. Interact. 2020, 12, 251–262. [Google Scholar] [CrossRef]
- Liu, T.; Wang, B.; Liu, M.; Jiang, K.; Wang, L. Stutzerimonas frequens strain TF18 with superior heterotrophic nitrification-aerobic denitrification ability for the treatment of aquaculture effluent. Process Biochem. 2023, 130, 156–165. [Google Scholar] [CrossRef]
- Qing, H.; Donde, O.O.; Tian, C.; Wang, C.; Wu, X.; Feng, S.; Liu, Y.; Xiao, B. Novel heterotrophic nitrogen removal and assimilation characteristic of the newly isolated bacterium Pseudomonas stutzeri AD-1. J. Biosci. Bioeng. 2018, 126, 339–345. [Google Scholar] [CrossRef]
- Deng, M.; Zhao, X.; Senbati, Y.; Song, K.; He, X. Nitrogen removal by heterotrophic nitrifying and aerobic denitrifying bacterium Pseudomonas sp. DM02: Removal performance, mechanism and immobilized application for real aquaculture wastewater treatment. Bioresour. Technol. 2021, 322, 124555. [Google Scholar] [CrossRef]
- Liu, S.; Liu, Q.; Wu, H.; Jiang, W.; Kahaer, A.; Tang, Q.; Hu, Z.; Hong, C.; Liu, D. Integrative chemical and omics analysis of the ammonia nitrogen removal characteristics and mechanism of a novel oligotrophic heterotrophic nitrification-aerobic denitrification bacterium. Sci. Total Environ. 2022, 852, 158519. [Google Scholar] [CrossRef]
- Gao, Y.; Zhu, J.; Wang, K.; Ma, Y.; Fang, J.; Liu, G. Discovery of a heterotrophic aerobic denitrification Pseudomonas sp. G16 and its unconventional nitrogen metabolic pathway. Bioresour. Technol. 2023, 387, 129670. [Google Scholar] [CrossRef] [PubMed]
- Arts, P.A.; Robertson, L.A.; Gijs Kuenen, J. Nitrification and denitrification by Thiosphaera pantotropha in aerobic chemostat cultures. FEMS Microbiol. Ecol. 1995, 18, 305–315. [Google Scholar] [CrossRef]
- Huang, M.-Q.; Cui, Y.-W.; Yang, H.-J.; Xu, M.-J.; Cui, Y.; Chen, Z. A halophilic aerobic-heterotrophic strain SND-01: Nitrogen removal by ammonium assimilation and heterotrophic nitrification-aerobic denitrification. Bioresour. Technol. 2023, 374, 128758. [Google Scholar] [CrossRef]
- Huang, M.-Q.; Cui, Y.-W.; Huang, J.-L.; Sun, F.-L.; Chen, S. A novel Pseudomonas aeruginosa strain performs simultaneous heterotrophic nitrification-aerobic denitrification and aerobic phosphate removal. Water Res. 2022, 221, 118823. [Google Scholar] [CrossRef]
- Huang, X.J.; Weisener, C.G.; Ni, J.P.; He, B.H.; Xie, D.T.; Li, Z.L. Nitrate assimilation, dissimilatory nitrate reduction to ammonium, and denitrification coexist in Pseudomonas putida Y-9 under aerobic conditions. Bioresour. Technol. 2020, 312, 123597. [Google Scholar] [CrossRef]
- Hu, J.; Yan, J.; Wu, L.; Bao, Y.; Yu, D.; Li, J. Simultaneous nitrification and denitrification of hypersaline wastewater by a robust bacterium Halomonas salifodinae from a repeated-batch acclimation. Bioresour. Technol. 2021, 341, 125818. [Google Scholar] [CrossRef]
- Zhang, M.; He, T.; Chen, M.; Wu, Q. Ammonium and hydroxylamine can be preferentially removed during simultaneous nitrification and denitrification by Pseudomonas taiwanensis EN-F2. Bioresour. Technol. 2022, 350, 126912. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Lei, Z.; He, X.; Zhang, Z.; Yang, Y.; Sugiura, N. Nitrate removal by Thiobacillus denitrificans immobilized on poly (vinyl alcohol) carriers. J. Hazard. Mater. 2009, 163, 1090–1095. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Huang, S.; Tian, Y.; Lu, X. Heterotrophic ammonium assimilation: An important driving force for aerobic denitrification of Rhodococcus erythropolis strain Y10. Chemosphere 2022, 291, 132910. [Google Scholar] [CrossRef] [PubMed]
- Rice, E.W.; Bridgewater, L.; American Public Health Association (Eds.) Standard Methods for the Examination of Water and Wastewater; American Public Health Association: Washington, DC, USA, 2012. [Google Scholar]
- Shu, H.; Sun, H.M.; Huang, W.; Zhao, Y.; Ma, Y.H.; Chen, W.; Sun, Y.P.; Chen, X.Y.; Zhong, P.; Yang, H.R.; et al. Nitrogen removal characteristics and potential application of the heterotrophic nitrifying-aerobic denitrifying bacteria Pseudomonas mendocina S16 and Enterobacter cloacae DS’5 isolated from aquaculture wastewater ponds. Bioresour. Technol. 2022, 345, 126541. [Google Scholar] [CrossRef]
- He, X.; Sun, Q.; Xu, T.; Dai, M.; Wei, D. Removal of nitrogen by heterotrophic nitrification–aerobic denitrification of a novel halotolerant bacterium Pseudomonas mendocina TJPU04. Bioprocess Biosyst. Eng. 2019, 42, 853–866. [Google Scholar] [CrossRef]
- Wan, C.L.; Yang, X.; Lee, D.J.; Du, M.A.; Wan, F.; Chen, C. Aerobic denitrification by novel isolated strain using NO2−-N as nitrogen source. Bioresour. Technol. 2011, 102, 7244–7248. [Google Scholar] [CrossRef]
- Padhi, S.K.; Tripathy, S.; Sen, R.; Mahapatra, A.S.; Mohanty, S.; Maiti, N.K. Characterisation of heterotrophic nitrifying and aerobic denitrifying Klebsiella pneumoniae CF-S9 strain for bioremediation of wastewater. Int. Biodeterior. Biodegrad. 2013, 78, 67–73. [Google Scholar] [CrossRef]
- Gu, X.; Leng, J.; Zhu, J.; Zhang, K.; Zhao, J.; Wu, P.; Xing, Q.; Tang, K.; Li, X.; Hu, B. Influence mechanism of C/N ratio on heterotrophic nitrification- aerobic denitrification process. Bioresour. Technol. 2022, 343, 126116. [Google Scholar] [CrossRef]
- Liu, Y.; Ai, G.-M.; Miao, L.-L.; Liu, Z.-P. Marinobacter strain NNA5, a newly isolated and highly efficient aerobic denitrifier with zero N2O emission. Bioresour. Technol. 2016, 206, 9–15. [Google Scholar] [CrossRef]
- Zhang, M.; Lu, H.; Cai, L.; Sun, P.; Ma, B.; Li, J.; Chen, G.; Ruan, Y. C/N ratios inform sustainable aerobic denitrification for nitrogen pollution control: Insights into the key parameter from a view of metabolic division. J. Clean. Prod. 2023, 414, 137565. [Google Scholar] [CrossRef]
- Kong, D.; Li, W.; Deng, Y.; Ruan, Y.; Chen, G.; Yu, J.; Lin, F. Denitrification-potential evaluation and nitrate-removal-pathway analysis of aerobic denitrifier strain Marinobacter hydrocarbonoclasticus RAD-2. Water 2018, 10, 1298. [Google Scholar] [CrossRef]
- Duan, J.; Fang, H.; Su, B.; Chen, J.; Lin, J. Characterization of a halophilic heterotrophic nitrification–aerobic denitrification bacterium and its application on treatment of saline wastewater. Bioresour. Technol. 2015, 179, 421–428. [Google Scholar] [CrossRef]
- Jean, W.D.; Yeh, Y.T.; Huang, S.P.; Chen, J.S.; Shieh, W.Y. Spongiibacter taiwanensis sp. nov., a marine bacterium isolated from aged seawater. Int. J. Syst. Evol. Microbiol. 2016, 66, 4094–4098. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.-G.; Zhang, X.-J.; Wang, Z.-S. Temperature effect on aerobic denitrification and nitrification. J. Environ. Sci. 2003, 15, 669–673. [Google Scholar]
- Yang, L.; Wang, X.H.; Cui, S.; Ren, Y.X.; Yu, J.; Chen, N.; Xiao, Q.; Guo, L.K.; Wang, R.H. Simultaneous removal of nitrogen and phosphorous by heterotrophic nitrification-aerobic denitrification of a metal resistant bacterium Pseudomonas putida strain NP5. Bioresour. Technol. 2019, 285, 121360. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; Li, X.; Fan, W.; Wang, J. Heterotrophic nitrification and aerobic denitrification by a novel Acinetobacter sp. ND7 isolated from municipal activated sludge. Bioresour. Technol. 2020, 301, 122749. [Google Scholar] [CrossRef]
- Zheng, L.; Dong, Y.; Li, B.; Yin, T.; Liu, C.; Lin, H. Simultaneous removal of high concentrations of ammonia nitrogen and calcium by the novel strain Paracoccus denitrificans AC-3 with good environmental adaptability. Bioresour. Technol. 2022, 359, 127457. [Google Scholar] [CrossRef] [PubMed]
- Yi, M.; Wang, H.; Ma, X.; Wang, C.; Wang, M.; Liu, Z.; Lu, M.; Cao, J.; Ke, X. Efficient nitrogen removal of a novel Pseudomonas chengduensis strain BF6 mainly through assimilation in the recirculating aquaculture systems. Bioresour. Technol. 2023, 379, 129036. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, Q.; Yang, X.; Wu, D.; Li, Y.; Di, H. Isolation and characteristics of two heterotrophic nitrifying and aerobic denitrifying bacteria, Achromobacter sp. strain HNDS-1 and Enterobacter sp. strain HNDS-6. Environ. Res. 2023, 220, 115240. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, L.; Sun, H.; Wang, W.; Yang, Y.; Qi, Z.; Liu, X. Ammonia assimilation: A double-edged sword influencing denitrification of Rhodobacter azotoformans and for nitrogen removal of aquaculture wastewater. Bioresour. Technol. 2022, 345, 126495. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Tang, Z.; Xia, S.; Jiang, Y.; Li, M.; Wang, B. The intrinsic relevance of nitrogen removal pathway to varying nitrate loading rate in a polycaprolactone-supported denitrification system. Biodegradation 2022, 33, 317–331. [Google Scholar] [CrossRef]
- Li, Y.; Li, C.-X.; Lin, W.; Wang, S.-S.; Zhang, W.-X.; Jiang, Y.-M.; Zhang, Y.; Han, Y.-H. Full evaluation of assimilatory and dissimilatory nitrate reduction in a new denitrifying bacterium Leclercia adecarboxylata strain AS3-1: Characterization and functional gene analysis. Environ. Technol. Innov. 2021, 23, 101731. [Google Scholar] [CrossRef]
- Zhang, M.; Li, A.; Yao, Q.; Wu, Q.; Zhu, H. Nitrogen removal characteristics of a versatile heterotrophic nitrifyingaerobic denitrifying bacterium, Pseudomonas bauzanensis DN13-1, isolatedfrom deep-sea sediment. Bioresour. Technol. 2020, 305, 122626. [Google Scholar] [CrossRef] [PubMed]
- Jin, P.; Chen, Y.; Yao, R.; Zheng, Z.; Du, Q. New insight into the nitrogen metabolism of simultaneous heterotrophic nitrification-aerobic denitrification bacterium in mRNA expression. J. Hazard. Mater. 2019, 371, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Hu, Y.; Qiu, G.; Liang, D.; Li, Y.; Cheng, J.; Chen, Y.; Wang, G.; Xie, J.; Zhu, X. Genomics and metabolic characteristics of simultaneous heterotrophic nitrification aerobic denitrification and aerobic phosphorus removal by Acinetobacter indicus CZH-5. Bioresour. Technol. 2024, 395, 130322. [Google Scholar] [CrossRef]
- Rout, P.R.; Bhunia, P.; Dash, R.R. Simultaneous removal of nitrogen and phosphorous from domestic wastewater using Bacillus cereus GS-5 strain exhibiting heterotrophic nitrification, aerobic denitrification and denitrifying phosphorous removal. Bioresour. Technol. 2017, 244, 484–495. [Google Scholar] [CrossRef]
- Yang, J.X.; Feng, L.; Pi, S.S.; Cui, D.; Ma, F.; Zhao, H.P.; Li, A. A critical review of aerobic denitrification: Insights into the intracellular electron transfer. Sci. Total Environ. 2020, 731, 139080. [Google Scholar] [CrossRef]
- Song, T.; Zhang, X.L.; Li, J.; Wu, X.Y.; Feng, H.X.; Dong, W.Y. A review of research progress of heterotrophic nitrification and aerobic denitrification microorganisms (HNADMs). Sci. Total Environ. 2021, 801, 149319. [Google Scholar] [CrossRef]
- Wu, S.Q.; Lv, N.; Zhou, Y.; Li, X.F. Simultaneous nitrogen removal via heterotrophic nitrification and aerobic denitrification by a novel Lysinibacillus fusiformis B301. Water Environ. Res. 2023, 95, e10850. [Google Scholar] [CrossRef]
- Zhang, W.; Shen, J.; Zhang, H.; Zheng, C.; Wei, R.; Gao, Y.; Yang, L. Efficient nitrate removal by Pseudomonas mendocina GL6 immobilized on biochar. Bioresour. Technol. 2021, 320, 124324. [Google Scholar] [CrossRef]
- Ruan, Y.J.; Taherzadeh, M.J.; Kong, D.D.; Lu, H.F.; Zhao, H.P.; Xu, X.Y.; Liu, Y.; Cai, L. Nitrogen removal performance and metabolic pathways analysis of a novel aerobic denitrifying halotolerant Pseudomonas balearica strain RAD-17. Microorganisms 2020, 8, 72. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).