Abstract
In order to investigate the effects of selenium (Se) against cadmium (Cd) toxicity, 180 healthy grass carp were separated into three groups and fed diets containing 0.147 (control group), 0.562, and 1.044 mg/kg of selenium Yeast throughout 60 days. In grass carp livers, malondialdehyde (MDA) content, antioxidant enzyme activities, and apoptosis-related gene expression were examined. As a result of acute exposure to cadmium, MDA content decreased significantly. With time, catalase (CAT), total superoxide dismutase (T-SOD), and glutathione peroxidase (GSH-Px) activities changed. The relative transcript levels of heavy metal scavenging genes abcc2 and mt2 were significantly reduced. The relative levels of expression of jnk, bax, caspase-3, caspase-8, and caspase-9 in apoptosis-associated factors were significantly elevated after cadmium exposure. Selenium-supplementation downregulated the expression of apoptosis-related factors. As compared to the control group, liver cells supplemented with selenium had a significantly lower apoptotic index. Comprehensive analysis showed that dietary selenium supplementation significantly attenuated cadmium-induced peroxidative damage and apoptosis in liver by increasing GSH-Px activity, and that cadmium toxicity could be alleviated by the addition of yeast selenium.
1. Introduction
Cadmium (Cd) is an unnecessary heavy metal element that is mainly used in industrial production such as electroplating and metallurgy [1,2]. According to studies, both acute and chronic exposure to cadmium can cause organic damage, including damage to the kidneys, liver, and bones [3]. Due to the illegal discharge of industrial wastewater and the infiltration of groundwater, the physiological functions and survival of aquatic animals contaminated by cadmium are threated. Reactive oxygen species (ROS) are induced to be produced in cells when aquatic organisms are exposed to cadmium, while large amounts of ROS trigger peroxidation reactions in the organism and subsequently trigger DNA damage [4,5]. It has been demonstrated in several studies that cadmium can cause apoptosis by increasing the activity of caspase-3 and caspase-8 in rainbow trout (Oncorhynchus mykiss) and carp cells [6,7]. Chan et al. [8] found that cadmium causes aberrant development in zebrafish embryos and induces apoptosis in embryonic cells by up-regulating the gene expression of caspase-7. A previous study has found that cadmium induced mitophagy in Hepa1-6 cells [9]. Meanwhile, in recent research, cadmium exposure will cause tubular and glomerular damage in mice [10].
Selenium (Se) is an essential trace element for maintaining the physiological activities of living organisms, and yeast selenium is widely used in aquaculture because of its adequate absorption and great muscle deposition rate [11]. Most of the biological activities of selenium are realized through its conversion into active selenoproteins, ranging from antioxidant and anti-inflammatory effects to active thyroid hormone production [12]. In particular, selenium is an important component of the GSH-PX enzyme system, which acts to convert GSH to GSSG and reduces toxic peroxides to non-toxic hydroxyl compounds, thereby protecting the structure and function of cell membranes [13]. The mitochondrial respiratory chain is a major source of ROS, and ROS production increases when mitochondria are damaged. Selenium has been shown to significantly reduce mitochondrial membrane potential, restore mitochondrial function, and reduce ROS production, thereby protecting against oxidative damage [14]. It has been proved that selenium-supplementation can reduce the absorption of cadmium by the organism, enhance the antioxidant function of the organism, scavenge cadmium-induced free radicals, and thus protect the organism against cadmium toxicity [15]. Selenium supplementation also greatly reduces lipid peroxidation damage in fish liver due to cadmium exposure [16]. Selenium deficiency reduces the activity of antioxidant enzymes in carp, leading to excessive accumulation of reactive oxygen species, causing oxidative stress and leading to apoptosis [17]. In addition, selenium supplementation not only improved the survival rate of TM3 cells in mice, but also improved cadmium-induced apoptosis by inhibiting caspase-3 activity [18].
In order to investigate the role of dietary selenium-supplementation on cadmium acute stress in grass carp liver. In this study, grass carp feed was supplemented with selenium yeast, and cadmium exposure experiments were conducted after 60 days. In the liver of grass carp, antioxidant indexes, apoptosis status, apoptosis pathway JNK detection, and transcript levels of caspase-related genes were assessed. To establish a theoretical foundation for the protective effects of aquatic animals exposed to heavy metal pollution, various dietary selenium concentrations were tested for their protective benefits against cadmium stress in the water.
2. Materials and Methods
2.1. Experimental Compound Feed
The formulation of the feed supplementation was set based on published studies on the nutritional requirements of grass carp with the particular elements shown in Table 1 [19,20,21]. Three doses of selenium supplementation, 0, 0.6, and 1.2 mg/kg, were established in this experiment based on previously published selenium Yeast (SY) concentration in grass carp diets [19,20]. Selenium yeast (Se content 2000 mg/kg) was purchased from Hubei Anqi Yeast Co.

Table 1.
Composition and nutrients content of the basal diet.
The feed ingredients were processed, pulverized, sieved through a 40-mesh sieve using a laboratory-type expanded feed pelletizer (Model 21204547), and mixed thoroughly according to the formula proportions using the progressive expansion method. The feed was then made into soft pellets with a particle size of 2.0 mm, dried in a 50 °C oven, and kept in a refrigerator at 4 °C as a backup. Se content of the feeds was determined using Inductively Coupled Plasma Optical Emission Spectrometry (ICP-OES). The Se content in the three groups of basic feeds was calculated to be 0.147 (control group), 0.562, and 1.044 mg/kg.
2.2. Experimental Fish Management and Experimental Design
Fish used in this experiment were handled humanely according to Huazhong Agricultural University’s Animal Management and Ethics Committee regulations (Approval code: HZAUFI-2019-040).
Juvenile grass carp were purchased from Bairong Fishery (Jingzhou, China) from the same batch of breeding, healthy, and disease-free grass carp. The fish were temporarily raised in a recirculating water culture system at Huazhong Agricultural University’s College of Aquatic Sciences. Before the start of the formal experiment, the grass carp were fed with basal feed for 2 weeks for domestication. A total of 180 healthy fish of uniform size were randomly divided into 9 breeding buckets (500 L) after 2 weeks, and every 3 buckets were used as one treatment, which were divided into 3 treatments and fed with three concentration gradients, with 3 replicates of each treatment.
The experimental period was conducted at 9:00 and 16:00 twice a day, and 3% of the total fish mass was fed, with the intake estimated after half an hour after feeding. Every two weeks, the grass carp were weighed and the daily feeding rate was adjusted according to the total mass. The average water temperature was maintained at 26.5 ± 1.0 °C, the average pH was maintained at 7.8 ± 0.2, the average dissolved oxygen > 5.5 mg/L, and the selenium concentration in the water <0.06 mg/kg. The experimental period was 60 days, and no mortality occurred during the culture.
After the feed selenium feeding experiment, the cadmium attack experiment was conducted after 48 h of fasting for grass carp. Seven concentration gradients of 0, 30, 60, 90, 120, 150, and 180 mg/L CdCl2·2.5H2O were set to determine the semi-lethal concentration (LC50) of cadmium to grass carp, and the LC50 of CdCl2·2.5H2O to grass carp was determined to be 90 mg/L. One tenth of the LC50 (9 mg/L) was used as the exposure concentration. Samples were taken after 0 h, 12 h, 24 h, 72 h, and 96 h of CdCl2·2.5H2O exposure. Three grass carp were randomly selected from each parallel group, and the fish were rapidly and deeply anesthetized using 200 mg/L MS-222 (Sigma, St.Louis, MO, USA). We dissected and separated the livers, weighed them, then froze them in liquid nitrogen and stored them at −80 °C in order to extract the RNA and determine the biochemical index. Hepatocytes were also fixed in paraformaldehyde and examined histopathologically.
2.3. Analysis of Malondialdehyde and Antioxidative Enzyme Activity
Liver tissues were weighed, cut into small pieces, homogenized in an ice-water bath, centrifuged to remove the supernatant, and the homogenized supernatant was taken for assay (protein concentration was determined by BCA (Beyotime, Shanghai, China) kit). Malondialdehyde (MDA), catalase (CAT), glutathione peroxidase (GSH-Px), and superoxide dismutase (T-SOD) were measured following the instructions, and the kits were purchased from Nanjing Jiancheng Bioengineering Institute(Nanjing, China)
2.4. Apoptosis-Related Gene Expression Levels Analysis
Electrophoresis on a 1% agarose gel was performed after TRIzol-extracted liver RNA samples were examined for integrity. A NanoDrop 2000 nucleic acid analyzer (Thermo Scientific, Waltham, MA, USA) was used to assess RNA concentrations. The Hifair® II 1st strand cDNA synthesis Super mix kit (YEASEN, Shanghai, China) was used to reverse-transcribe the cDNA, which was then kept at −20 °C.
To determine the transcript levels of genes involved in heavy metal-induced apoptosis, we created primers based on core sequence fragments obtained from GenBank(Tianyi Huiyuan Biotechnology Co., Ltd., Wuhan, China). The primer sequences can be found in Table 2. A Quant Studio 6 Flex Real-Time PCR Detection System (Applied Biosystems, Foster City, California, USA) was used to perform the qPCR reactions using Hieff® qPCR SYBR Green Master Mix (No Rox Plus)(Shanghai, YEASEN). The reaction system was: 10 µL Hieff™ qPCR SYBR® Green Master Mix (No Rox Plus)(Shanghai, YEASEN), 0.4 µL each of upstream and downstream primers, 7.2 µL of double-distilled water, and 2 µL of cDNA template. In the reaction procedure, 95 °C predenaturation was followed by denaturation for 10 s, then 58 °C annealing, 72 °C extension, and 40 cycles of 95 °C. The relative gene expression was quantified using the 2−ΔΔCt method and β-actin was chose to normalize the data.
Table 2.
Primers of q-PCR.
2.5. TUNEL Method to Detect Apoptosis in Liver Tissues
Apoptosis detection was performed on liver tissues using DAPI and TUNEL staining, in which positive cell nuclei fluoresced in green. A fluorescence microscope was used to examine and photograph grass carp liver tissue sections. With Image Pro-Plus 6.0 image analysis software, we counted the number of positive cells and total number of cells under three fields of view and calculated the apoptosis index by dividing the number of positive cells by the number of total cells.
2.6. Statistical Analysis
All data in the experiment were first processed initially using Excel and experimental data were represented as mean ± standard deviation (SD). SPSS 19.0 software was used to analyze the experimental results. For determining significance, one-way analysis of variance (one-way ANOVA) was conducted once the normal distribution and chi-square values of the experimental data had been verified. For the study of multiple comparisons, Duncan’s test was employed. A difference was deemed significant if p < 0.05.
3. Results
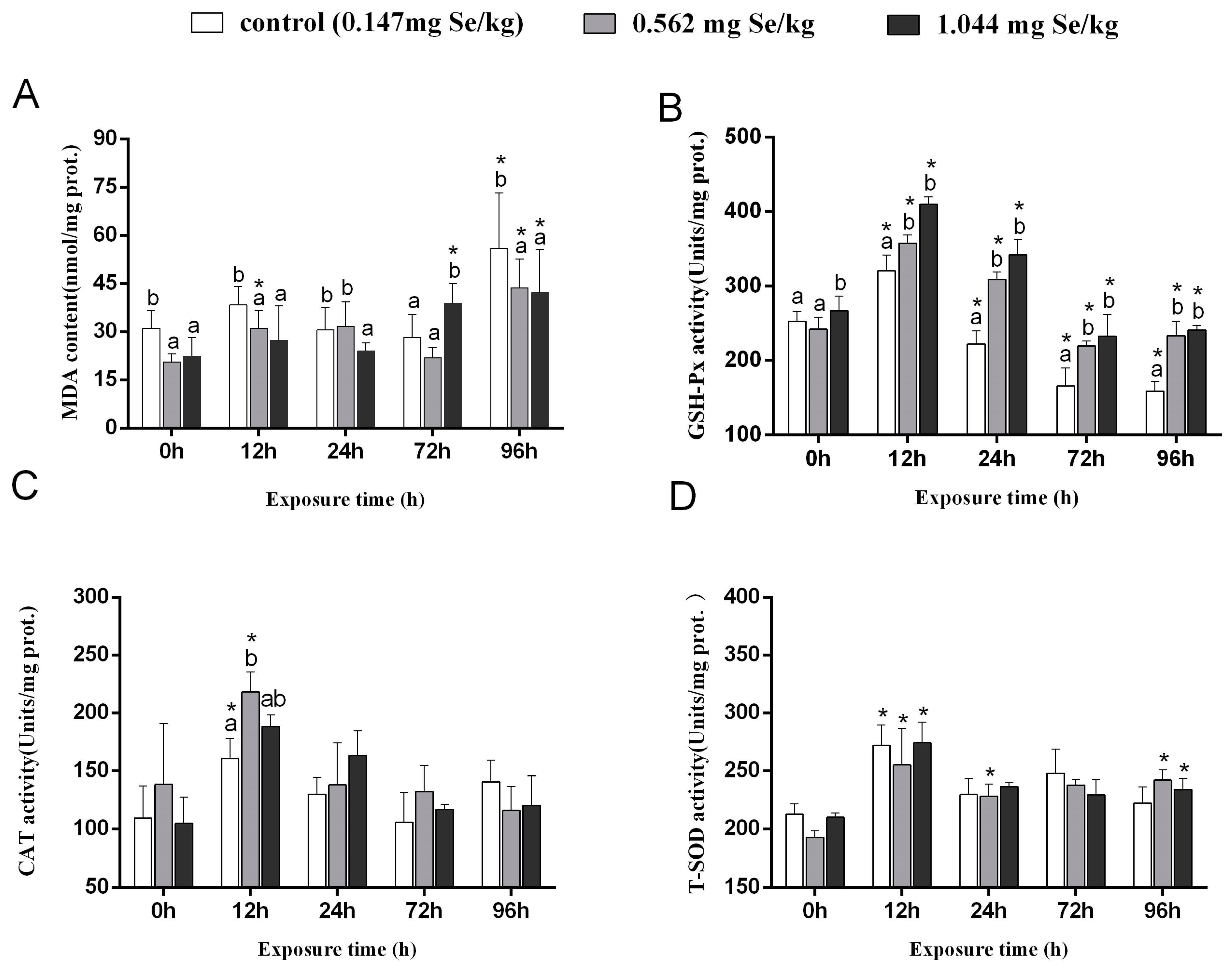
3.1. MDA Content and Antioxidant Enzyme Activity
MDA levels in grass carp livers in the control group increased considerably post-cadmium stress (Figure 1A). After 24 h of cadmium stress, MDA content was significantly increased in the 0.147 mg/kg and 0.562 mg/kg groups compared to the 1.044 mg/kg group. The content of MDA of grass carp exposed to cadmium for 96 h was significantly higher than it was before the stress (0 h), with the 0.147 mg/kg group having the highest MDA level compared to the two selenium-supplemented groups.
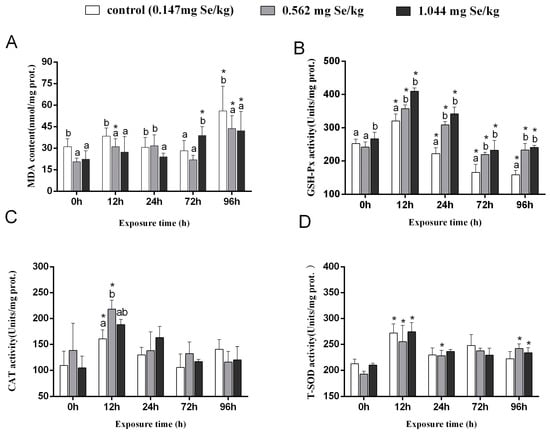
Figure 1.
Effects of dietary selenium on MDA (A), GSH-Px (B), CAT (C), and T-SOD (D) levels in liver of grass carp under cadmium stress (Note: Different letters indicate significant differences between different treatment groups at the same time point (p < 0.05), and asterisks indicate significant differences between the same treatment group and the 0 h group at different time periods; N = 9; ).
In grass carp, GSH-Px activity first increased and then decreased after cadmium stress. In both selenium-supplemented groups, GSH-Px activity was markedly higher than in the control group (Figure 1B). As a result of exposure to cadmium stress for 12 h, the 0.562 mg/kg group showed markedly higher CAT activity than the control group. There were no discernible variations in the three groups’ CAT activities after exposure to cadmium for 24 h, 72 h, or 96 h (Figure 1C). The T-SOD activity of cadmium exposure showed an increasing and decreasing trend, among which the highest activity was observed at 12 h. A higher level of T-SOD activity was observed after 96 h in the selenium supplemented group compared to the 0 h (Figure 1D).
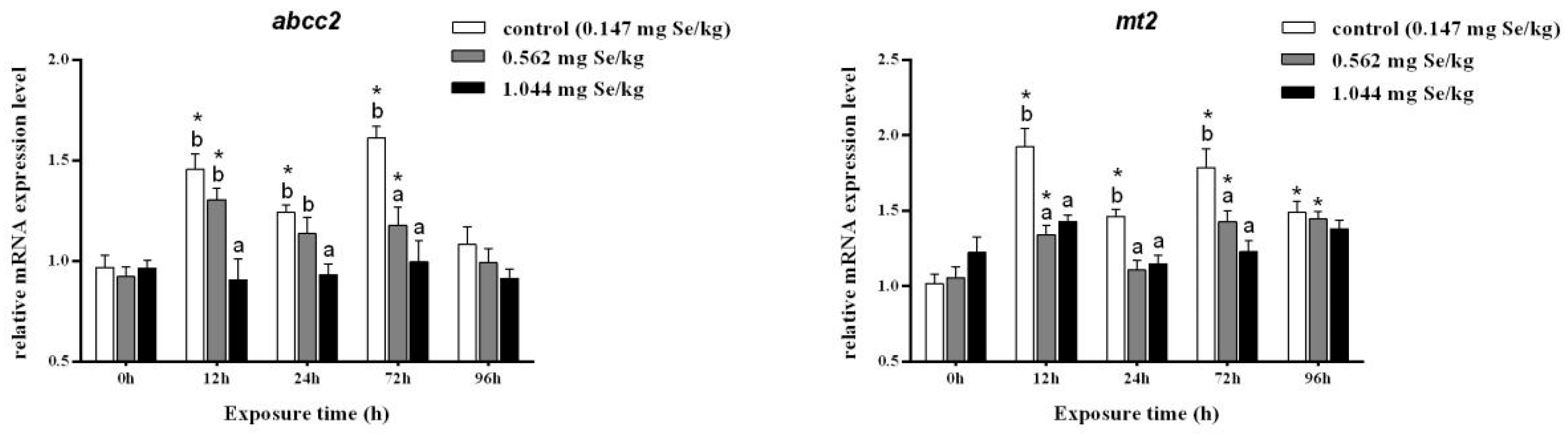
3.2. Expression Levels of Heavy Metal Scavenging Genes
Cadmium exposure at 12, 24, and 72 h resulted in significantly higher relative abcc2 expression levels in the 0.147 mg/kg group. Among them, the relative expression of abcc2 in the selenium-supplemented group decreased significantly after 12 h of cadmium exposure. A 96 h dose of cadmium stress decreased RNA levels of abcc2, and no significant differences were observed among treatment groups.
Cadmium stress significantly increased the relative expression level of mt2 in the liver of grass carp. After selenium supplementation, the relative expression level of mt2 was significantly reduced, and there was no significant difference between the three groups after 96 h of cadmium exposure (Figure 2).
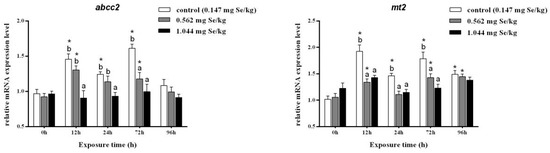
Figure 2.
Effects of dietary selenium on the relative transcription level of abcc2 and mt2 in liver of grass carp under cadmium stress (Note: Different letters indicate significant differences between different treatment groups at the same time point (p < 0.05), and asterisks indicate significant differences between the same treatment group and the 0 h group at different time periods; N = 9; ).
3.3. Expression Levels of Apoptosis Gene
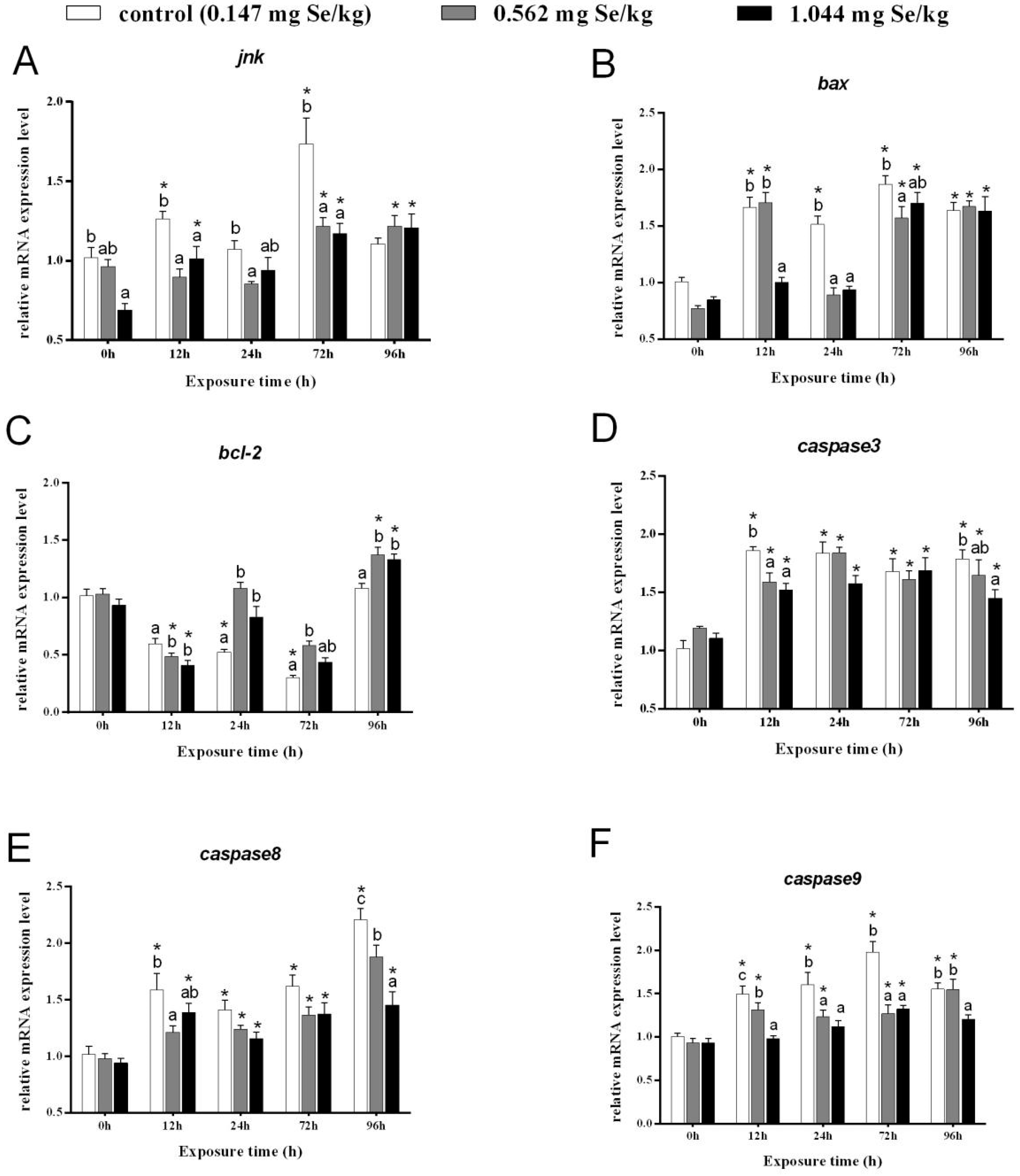
After 12 h of cadmium exposure, the relative transcript levels of jnk in the control group were obviously elevated and were notably higher than those in the selenium-supplemented group. After 72 h of cadmium exposure, a significant difference was found between the jnk of the 0.147 mg/kg group and that of the selenium-supplemented group. However, the relative expression of jnk in the 0.562 mg/kg group and 1.044 mg/kg group was significantly higher than that before cadmium exposure (Figure 3A).
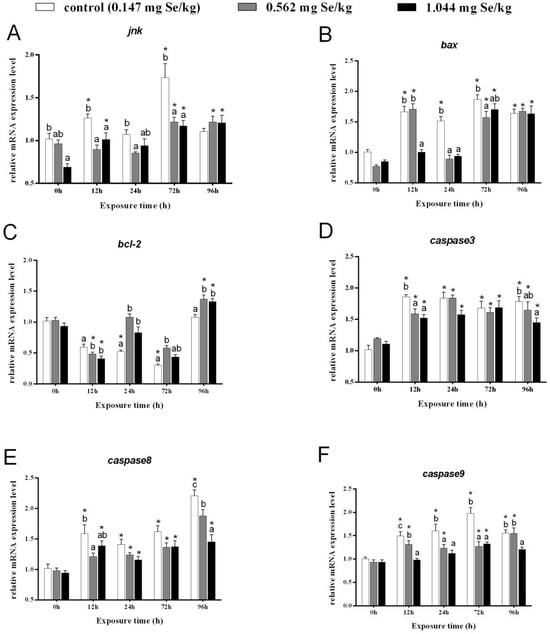
Figure 3.
Effects of dietary selenium on the relative transcription level of jnk (A), bax (B), bcl-2 (C), caspase-3 (D), caspase-8 (E), and caspase-9 (F) in liver of grass carp under cadmium stress (Note: Different letters indicate significant differences between different treatment groups at the same time point (p < 0.05), and asterisks indicate significant differences between the same treatment group and the 0h group at different time periods; N = 9; ).
In the control group, the levels of bax transcripts were much higher than before exposure, while expression of bax in the 1.044 mg/kg group was markedly reduced after 12 and 24 h of cadmium exposure. A significant increase in bax expression was observed after 72 h of exposure to cadmium. All the groups were notably higher than before stress after 96 h, although differences between groups were not significant (Figure 3B).
At 12 h, grass carp with selenium supplementation had notably lower levels of bcl-2. A significant increase in bcl-2 expression after selenium supplementation was evident after 24 and 72 h after cadmium exposure, but a significant decrease was observed in the control group before cadmium exposure (Figure 3C).
After 12 h, the control and selenium-supplementation groups showed evidently higher expression levels of caspase-3. Neither group showed significant differences after 24 or 72 h of cadmium exposure (Figure 3D). However, at 96 h, caspase-3 expression showed a decreasing trend with increasing selenium concentration.
After 12 h of cadmium stress, relatively higher expression of caspase-8 was found in the control and 1.044 mg/kg groups than before stress, which decreased after selenium-supplementation. The relative expression of caspase-8 in the livers of grass carp from all three groups was significantly elevated at 24 h and 72 h, but no significant difference was observed between the groups. After 96 h of cadmium stress, caspase-8 expression increased markedly in all groups and decreased markedly with increasing selenium-supplementation concentration (Figure 3E).
Under heavy metal cadmium stress, relatively higher caspase9 expression was observed in both the control and 0.562 mg/kg groups. Following 12 h, 24 h, and 72 h, selenium-supplementation reduced caspase-9 expression clearly compared to control groups. Notably, caspase-9 expression was significantly different between selenium-supplemented and control groups after 12 h and 96 h (Figure 3F).
3.4. Effect of Dietary Selenium on Apoptosis of Grass Carp Liver Cells under Cadmium Stress
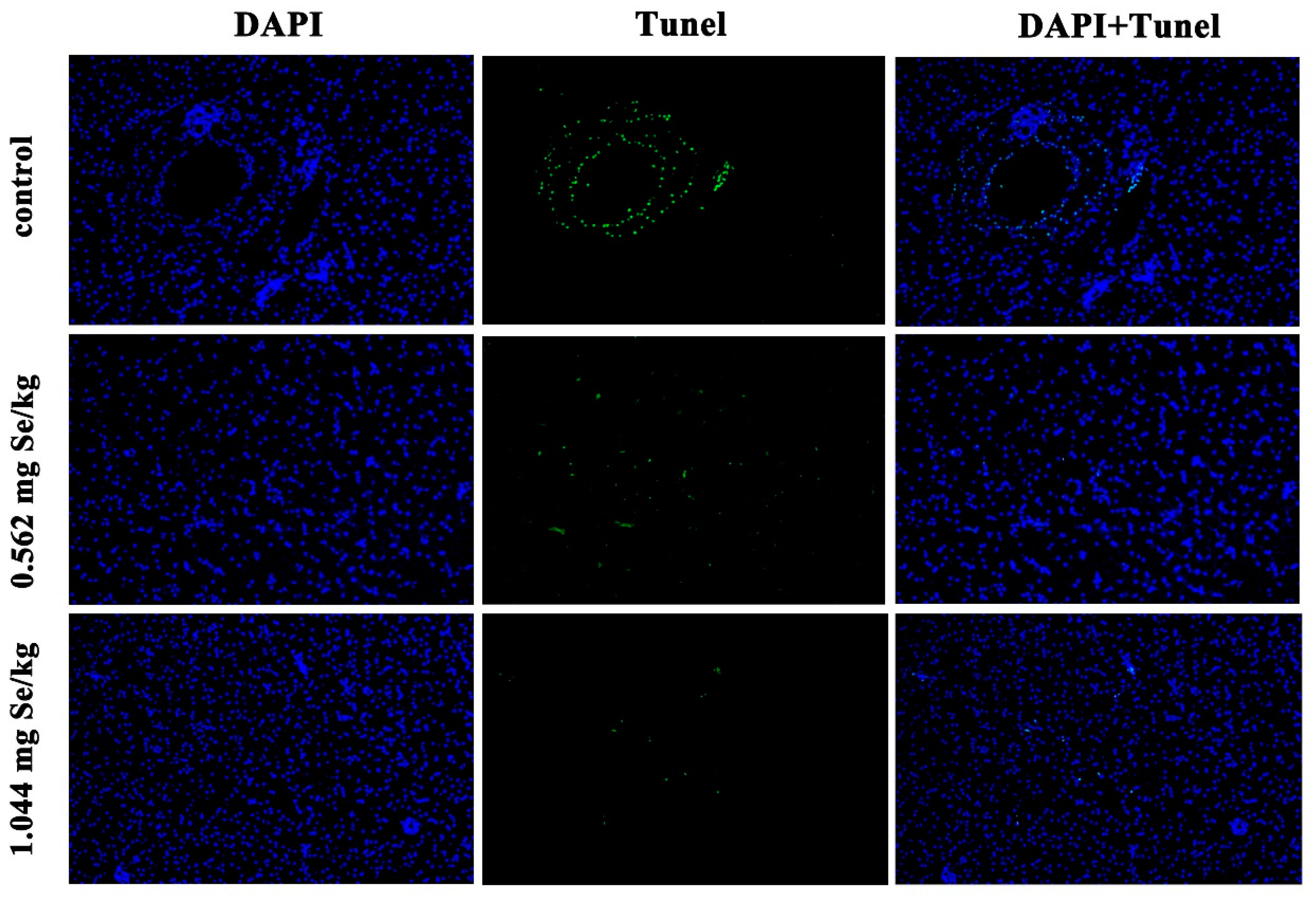
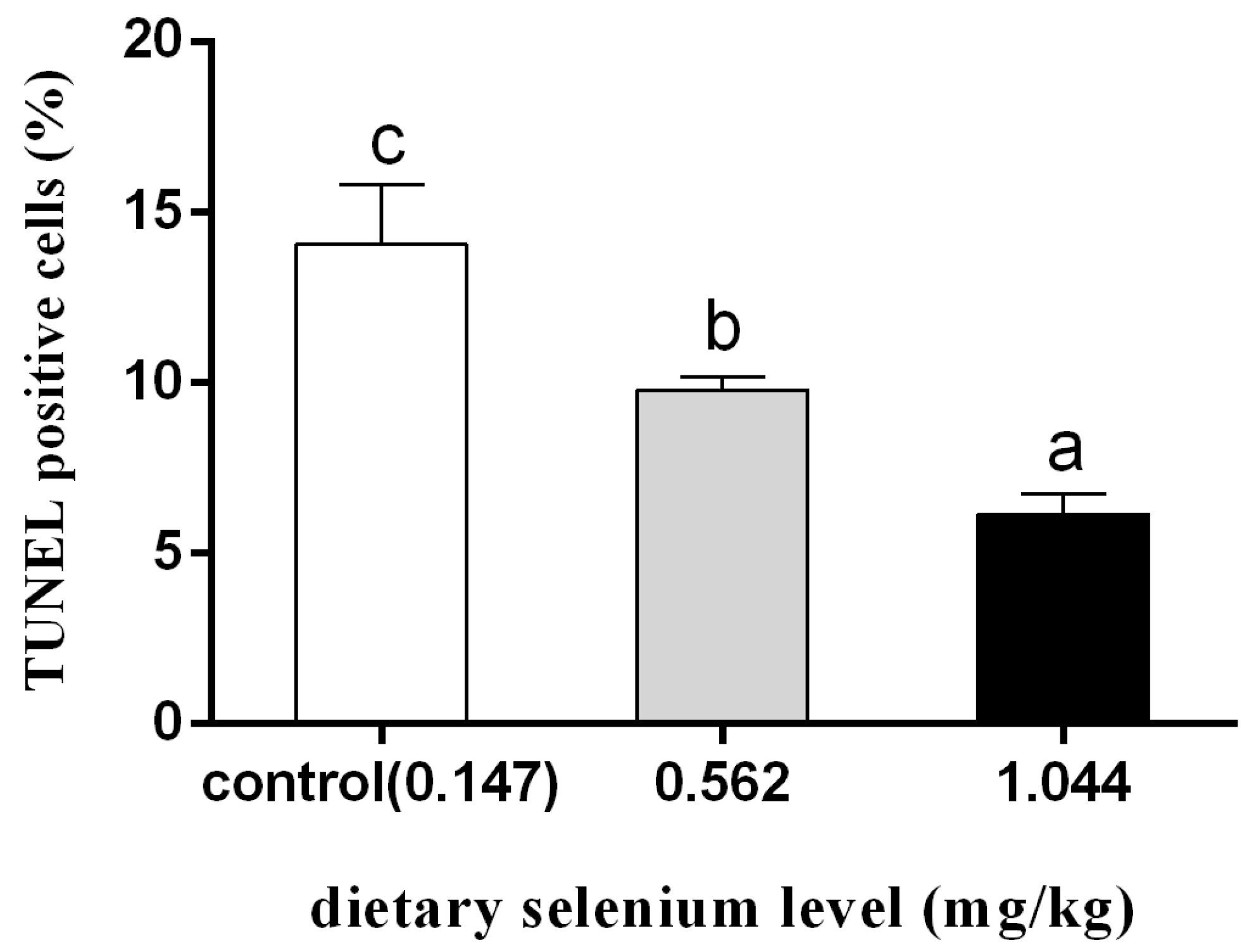
According to Figure 4, after 12 h of cadmium exposure, the selenium-supplemented group showed significantly reduced green fluorescence compared to the control group. Quantitative analysis showed that the apoptotic index of selenium-supplementation was much lower than that of the control group, and the higher the selenium-supplementation, the lower the apoptotic index (Figure 5).
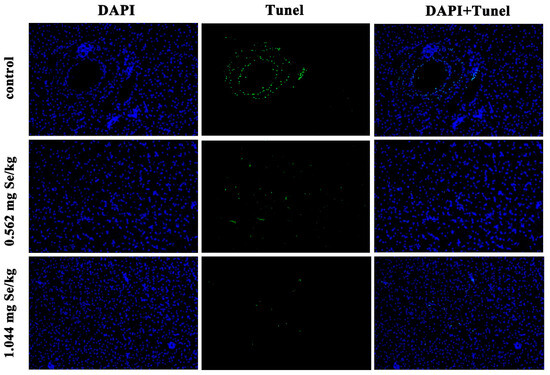
Figure 4.
Effect of dietary selenium on apoptosis of liver cells in grass carp under cadmium stress. Green color indicates TUNEL positive cells.
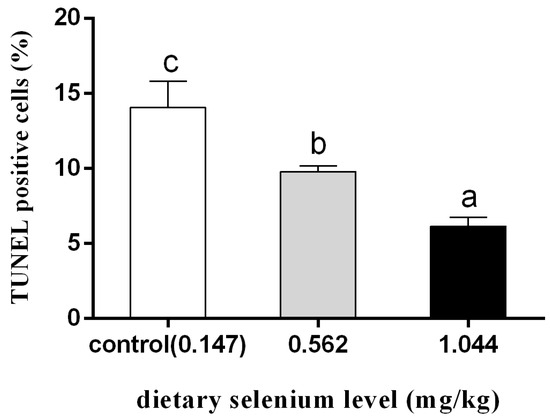
Figure 5.
Quantitative analysis of percentage of apoptotic cells (different letters indicate significant differences between treatment groups at the same time point; N = 9; ).
4. Discussion
Aquatic organisms are more susceptible to cadmium contamination than terrestrial animals, and aquatic animals accumulate cadmium [22]. In aquatic ecosystems, cadmium accumulates in aquatic organisms and harms them. Animals’ livers are found to be affected by cadmium because it increases reactive oxygen species, which attack membranes and lead to lipid peroxidation [23,24]. Assessment of molecular damage in fish organisms can be accomplished by measuring levels of MDA, which results from lipid peroxidation reactions [16]. According to Jia et al. [25], grass carp livers exposed to Cd water for seven days contain significantly more MDA than those exposed to control water. Exposure to cadmium may cause oxidative damage to grass carp’s liver.
The trace element selenium is essential for animal development. Studies have shown that selenium has an antagonistic effect on the toxicity of cadmium [16,26,27]. A study in grass carp livers found that MDA levels in the selenium supplemented group were significantly lower than those in the control group. In grass carp lymphocytes, MDA content increased significantly after acute exposure to cadmium, but decreased significantly after co-exposure to cadmium and selenium [28]. Liu et al. [27] and Li et al. [29] found that chickens fed a mixture of cadmium chloride (CdCl2) and sodium selenite (Na2SeO3) had considerably lower MDA levels in liver and kidney than chickens fed cadmium alone. Selenoprotein T has been linked to problems with lipid and glucose metabolism as well as cancer in recent research [30].
By generating a large amount of ROS, oxidative stress activates the antioxidant defense system, which maintains the organism’s normal function [26]; GSH-Px, CAT, and SOD are three enzymes that have been detected in the present study as indicators of the antioxidant system of the organism [31]. Among them, there was a significant increase in GSH-Px, CAT, and SOD activities after 12 h of cadmium stress, which may be a compensatory response by the organism to oxidative stress [32]. However, when the duration of cadmium stress increased, the activities of these three enzymes began to decline again. This is due to the fact that with the prolongation of stress, excessive ROS were produced in the cells, and the large amount of ROS counteracted the detoxification or antioxidant mechanisms. Due to this situation, when oxidative damage is severe, compensatory mechanisms fail, and antioxidant enzymes are suppressed, ultimately leading to the death of cells [32,33]. Pillet et al. [34] found that GSH-Px activity in carp liver was reduced considerably after 7 days of exposure to Cd-contaminated water, which is in agreement with what we observed in grass carp. A significant increase in GSH-Px activity was observed in groups supplemented with selenium under cadmium stress compared to control groups. Talas et al. [16] made a similar finding in salmonid liver, where they found that selenium-supplemented groups had significantly higher GSH-Px activity than control groups. Our study found no obvious differences in CAT or T-SOD activities between selenium-supplemented and control groups, suggesting that selenium increases GSH-Px activity as its main mechanism of action. The antioxidant capacity of grass carp may be improved by dietary selenium supplementation, since selenium is an important component of GSH-Px.
MT is an important metal detoxification protein that helps cells resist heavy metal toxicity and has a great affinity for metal ions [35]. MT is widely found in microorganisms, plants, and animals. It plays important functions such as removing non-essential heavy metals, regulating the concentration of essential heavy metals and antioxidant. Cadmium stress will activate the body’s endogenous protection mechanism and increase the content of MT protein [36]. Thus, MT gene expression can be used to assess heavy metal damage suffered by the body [37]. Tan et al. [38] found that cadmium could induce the relative transcription level of mt2 gene in grass carp liver, and with the extension of exposure time, the transcription level showed a trend of increasing first and decreasing, which was consistent with the results observed in the control group. This indicated that mt2 gene was induced to increase significantly to resist the toxicity of heavy metal cadmium under cadmium stress, and this induction effect had a time effect, which was greater in the early stage of exposure. In contrast, at the late stage of cadmium induction, the rise of cadmium accumulation in most tissues was slowed down, and MT2 mRNA expression was reduced. This may be due to the “indication of saturation” or “acclimation processes of the burdens” induced by metal accumulation [38]. After entering the body, cadmium is initially absorbed by the liver, combined with GSH in the liver and discharged into the bile, and can also be stored after binding to MT [39]. A significant difference was found between the selenium supplementation group and the control group in the relative transcription level of the mt2 gene, indicating that cadmium symptoms were mild and the mt2 gene was not induced [40].
MRPs (multidrug resistance proteins) are a group of crucial elements in the removal of heavy metals. The MRP family has two members, ABCC2 and MRP2 [41,42]. Numerous organic anions, including endogenous substances like bilirubin and foreign substances like heavy metals, are secreted and eliminated by the liver under the control of ABCC2. Payen et al. [43] according to studies, ABCC2 is crucial in assisting fish to handle the stress caused by heavy metals. In zebrafish embryos exposed to cadmium stress, the abcc2 gene expression increased significantly [44]. Cadmium exposure increases abcc2 gene expression in grass carp liver and kidney first, then decreases [38]. This experiment also found that with the increase of heavy metal cadmium exposure time, abcc2’s relative expression level first increased, then decreased, and reached the peak at 12 h, indicating that cadmium induced abcc2 mainly in the early stage. In this study, the relative transcription level of abcc2 gene in 1.044 mg/kg of selenium group was not affected by cadmium stress, indicating that dietary selenium supplementation effectively alleviated the heavy metal toxicity of cadmium.
Heavy metals have been shown to cause apoptosis in animal cells in numerous studies [45,46]. The regulation of apoptotic pathway mainly depends on the changes in apoptosis related molecules (e.g., JNK, Bax). JNK regulates downstream pro-apoptotic (bax) and anti-apoptotic genes (bcl-2) by phosphorylating or activating specific transcription factors [47,48]. Under cadmium stress in water, the transcript levels of the jnk and bax genes were significantly upregulated, suggesting grass carp livers undergo apoptosis. Bcl-2 is an anti-apoptotic protein that prevents oligomerization by forming a heterodimer with Bax [49]. After cadmium exposure, bcl-2 gene relative expression levels increased and then decreased, suggesting that bcl-2 is activated in the early stage of cadmium stress. Under short-term cadmium stress, grass carp can produce a large amount of BCL-2 protein to remove excessive ROS and inhibit apoptosis by up-regulating bcl-2 gene [50]. With the increase of cadmium exposure time, ROS could not be scavenged in time, resulting in severe oxidative damage to cells. Therefore, we can only inhibit bcl-2 expression and up-regulate bax gene to cause cell apoptosis to achieve the purpose of maintaining homeostasis [51]. Duan et al. [52] found that the relative transcript levels of bcl-2 genes in the liver of grass carp were significantly increased after 8 days of cadmium stress, and significantly decreased after 16 days of stress. In fish, gene expression levels of the Caspase family can be used as a marker of early apoptosis [6]. Cadmium poisoning activatescCaspase-3A activity of the purse red common carp (Cyprinus carpio) liver [6]. Cadmium stress also caused significant elevation of caspase-9 gene of grass carp liver [52]. After selenium supplementation, the relative expression of anti-apoptotic genes increased significantly, and the pro-apoptotic genes decreased significantly. Selenium is considered to be the most effective substance for the prevention of cadmium damage [53]. Recent studies have shown that the addition of selenium to the diet can reduce the expression of lead-induced pro-apoptotic genes in chicken kidney cells, thereby inhibiting apoptosis [54]. Zhang et al. [28] study showed that cadmium-induced apoptosis could be reduced after pre-incubation of selenium in carp lymphocytes. Similarly, selenium has been shown to reduce cadmium-induced apoptosis in grass carp liver cells [55]. Therefore, the heavy metal cadmium can cause apoptosis by activating the caspase pathway and is antagonistic to selenium.
TUNEL analysis of liver cells from the purse red common carp revealed that cadmium stress increased apoptosis significantly [6]. The TUNEL analysis in the study revealed that all liver cells of grass carp were apoptotic to varying degrees after cadmium stress, but it was obvious that the selenium-supplementation group played a protective role against cadmium-induced apoptosis. Selenium also antagonized heavy metal cadmium-induced apoptosis in chicken liver cells [56]. In mouse Leydig TM3 cells, selenium inhibited cadmium-induced apoptosis by inhibiting JNK–Caspase-3 signaling [18]. In the same way, jnk and caspase series gene expression in the selenium-supplemented group was significantly decreased. Thus, selenium may ameliorate cadmium-induced apoptosis by inhibiting the transcript levels of JNK–Caspase pathway-related genes, as well as up-regulating bcl-2 and down-regulating bax.
5. Conclusions
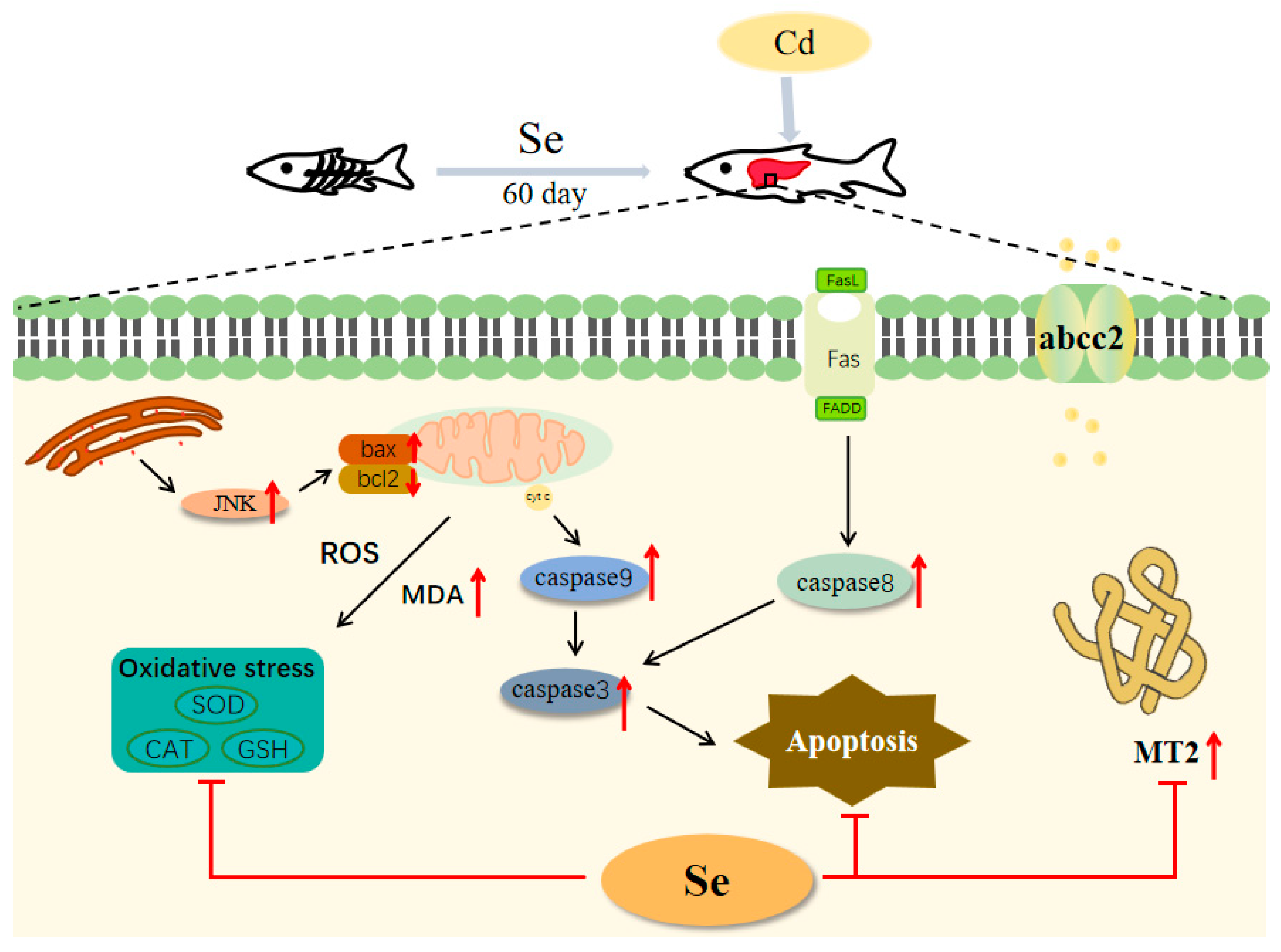
As shown in Figure 6, selenium significantly enhanced GSH-Px activity. Cadmium stress led to an increase in the expression of heavy metal detoxification genes abcc2 and mt, which could be significantly reduced by the addition of selenium. Meanwhile, the addition of selenium to the feed could inhibit heavy metal cadmium-induced apoptosis. However, according to the findings of this study, the difference in the amount of yeast selenium added does not seem to have a great difference on the expression of antioxidant enzymes, apoptosis-related genes, and heavy metal detoxification genes in fish. Therefore, in practical production, the suggested concentration of added yeast selenium is 0.562 mg/kg. On the basis of this study, we have a deeper comprehension of the physiological toxicity of cadmium to fish and lay a theoretical foundation for the management of cadmium poisoning in grass carp.
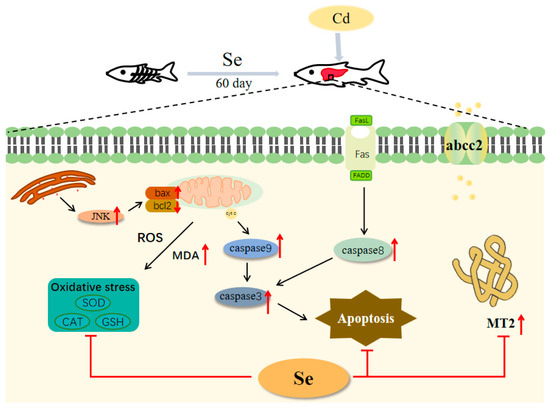
Figure 6.
Schematic representation of the protective effect of selenium on cadmium-induced liver apoptosis in grass carp (Ctenopharyngodon idellus).
Author Contributions
Conceptualization, Y.-L.Z., P.M. and R.T.; Data curation, Y.-L.Z. and P.M.; Formal analysis, Y.-L.Z. and P.M.; Funding acquisition, R.T. and D.-P.L.; Investigation, Y.-L.Z. and P.M.; Methodology, Y.-L.Z., P.M. and M.L.; Supervision, R.T. and D.-P.L.; Visualization, Y.-L.Z., P.M. and M.L.; Writing—original draft, Y.-L.Z. and P.M.; Writing—review and editing, Y.-L.Z., P.M., M.L., R.T. and D.-P.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Key Research and Development Program of China (2023YFD2400503), the earmarked fund for CARS (CARS-45), and the National Natural Science Foundation (No. 31502140).
Data Availability Statement
The data supporting the conclusions of this article will be made available by the authors on request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Lucia, M.; Andre, J.M.; Gonzalez, P.; Baudrimont, M.; Gontier, K.; Maury-Brachet, R.; Davail, S. Impact of cadmium on aquatic bird Cairina moschata. Biometals 2009, 22, 843–853. [Google Scholar] [CrossRef] [PubMed]
- Roccheri, M.C.; Agnello, M.; Bonaventura, R.; Matranga, V. Cadmium induces the expression of specific stress proteins in sea urchin embryos. Biochem. Biophys. Res. Commun. 2004, 321, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Hagedoorn, I.J.M.; Gant, C.M.; v Huizen, S.; Maatman, R.; Navis, G.; Bakker, S.J.L.; Laverman, G.D. Lifestyle-Related Exposure to Cadmium and Lead is Associated with Diabetic Kidney Disease. J. Clin. Med. 2020, 9, e2432. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Chen, M.G.; Ma, Q. Activation of Nrf2 in defense against cadmium-induced oxidative stress. Chem. Res. Toxicol. 2008, 21, 1375–1383. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lo, L.; Chan, W. Temporal expression and T3 induction of thyroid hormone receptors alpha1 and beta1 during early embryonic and larval development in zebrafish (Danio rerio). Mol. Cell. Endocrinol. 2000, 159, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.; Xu, Z.; Qiao, P.P.; Liu, S.; Zhang, L.; He, P.H.; Zhang, X.Y.; Wang, Y.N.; Min, W.P. Cadmium induces liver cell apoptosis through caspase-3A activation in purse red common carp (Cyprinus carpio). PLoS ONE 2013, 8, e83423. [Google Scholar] [CrossRef]
- Krumschnabel, G.; Ebner, H.L.; Hess, M.W.; Villunger, A. Apoptosis and necroptosis are induced in rainbow trout cell lines exposed to cadmium. Aquat. Toxicol. 2010, 99, 73–85. [Google Scholar] [CrossRef]
- Chan, P.K.; Cheng, S.H. Cadmium-induced ectopic apoptosis in zebrafish embryos. Arch. Toxicol. 2003, 77, 69–79. [Google Scholar] [CrossRef]
- Bai, H.; Yang, F.; Jiang, W.J.; Hu, A.M.; Chang, H.F.; Zhang, Y.L.; Jiang, L.; Lin, S.X.; Lu, Z.T.; Zhang, C.Y.; et al. Molybdenum and cadmium co-induce mitophagy and mitochondrial dysfunction via ROS-mediated PINK1/Parkin pathway in Hepa1-6 cells. Ecotoxicol. Environ. Saf. 2021, 224, e112618. [Google Scholar] [CrossRef]
- Ouyang, C.; Ke, H.Y.; Zhou, J.; Zhao, J.L.; Ma, X.; Liu, Y.M.; Li, X.H.; Li, W.W. Renal injury induced by cadmium chloride and the protective effect of vitamin C in mice. J. Hyg. Res. 2022, 51, 791–807. [Google Scholar]
- Tan, C.; Gao, Z.Y.; Wang, X.; Zhou, H.H.; Mai, K.S.; He, G. Nutritional effect of yeast selenium and its application in aquaculture. Hebei Fish. 2018, 7, 52–54+60. [Google Scholar]
- Zheng, R.L.; Chen, D.Y.; Su, J.Y.; Lai, J.; Wang, C.Y.; Chen, H.T.; Ning, Z.H.; Liu, X.; Tian, X.G.; Li, Y.H.; et al. Inhibition of HAdV-14 induced apoptosis by selenocystine through ROS-mediated PARP and p53 signaling pathways. J. Trace Elem. Med. Biol. 2023, 79, 127213. [Google Scholar] [CrossRef]
- Zheng, S.F.; Jin, X.; Chen, M.H.; Shi, Q.X.; Zhang, H.F.; Xu, S.W. Hydrogen sulfide exposure induces jejunum injury via CYP450s/ROS pathway in broilers. Chemosphere 2019, 214, 25–34. [Google Scholar] [CrossRef]
- Rayman, M.P. Selenium and human health. Lancet 2012, 379, 1256–1268. [Google Scholar] [CrossRef]
- Cui, J.; Liu, T.; Li, Y.; Li, F. Selenium reduces cadmium uptake into rice suspension cells by regulating the expression of lignin synthesis and cadmium-related genes. Sci. Total Environ. 2018, 644, 602–610. [Google Scholar] [CrossRef]
- Talas, Z.S.; Orun, I.; Ozdemir, I.; Erdogan, K.; Alkan, A.; Yilmaz, I. Antioxidative role of selenium against the toxic effect of heavy metals (Cd+2, Cr+3) on liver of rainbow trout (Oncorhynchus mykiss). Fish Physiol. Biochem. 2008, 34, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Cao, J.W.; Xu, T.C.; Liu, T.J.; Zhu, M.R.; Guo, M.R. Selenium deficiency induced inflammation and apoptosis via NF-κB and MAPKs pathways in muscle of common carp (Cyprinus carpio L.). Fish Shellfish Immunol. 2023, 138, 108847. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.M.; Wang, S.S.; Zhang, C.Q.; Hu, X.D.; Zhou, L.; Li, Y.H.; Xu, L.C. Selenium ameliorates cadmium-induced mouse leydig TM3 cell apoptosis via inhibiting the ROS/JNK/c-jun signaling pathway. Ecotoxicol. Environ. Saf. 2020, 192, e110266. [Google Scholar] [CrossRef]
- Zheng, L.; Feng, L.; Jiang, W.D.; Wu, P.; Tang, L.; Kuang, S.Y.; Zeng, Y.Y.; Zhou, X.Q.; Liu, Y. Selenium deficiency impaired immune function of the immune organs in young grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2018, 77, 53–70. [Google Scholar] [CrossRef]
- Liu, L.; Liang, X.; Li, J.; Fang, J.; Alam, M. Effects of dietary selenium on growth performance and oxidative stress in juvenile grass carp (Ctenopharyngodon idellus). Aquacult. Nutr. 2018, 24, 1296–1303. [Google Scholar] [CrossRef]
- Su, C.F.; Luo, L.; Wen, H.; Chen, X.C.; Sheng, X.S.; Chen, Z. Effects of selenium on growth, nutritional composition and digestive enzyme activity of grass carp (Ctenopharyngodon idella). J. Shanghai Ocean Univ. 2007, 02, 124–129. [Google Scholar]
- Barwick, M.; Maher, W. Biotransference and biomagnification of selenium copper, cadmium, zinc, arsenic and lead in a temperate seagrass ecosystem from Lake Macquarie Estuary, NSW, Australia. Mar. Environ. Res. 2003, 56, 471–502. [Google Scholar] [CrossRef] [PubMed]
- Waisberg, M.; Joseph, P.; Hale, B.; Beyersmann, D. Molecular and cellular mechanisms of cadmium carcinogenesis. Toxicology 2003, 192, 95–117. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Qu, W.; Kadiiska, M.B. Role of oxidative stress in cadmium toxicity and carcinogenesis. Toxicol. Appl. Pharmacol. 2009, 238, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.Y.; Zhang, H.J.; Liu, X.Y. Low levels of cadmium exposure induce DNA damage and oxidative stress in the liver of Oujiang colored common carp (Cyprinus carpio) var. Color. Fish Physiol. Biochem. 2011, 37, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Wan, N.; Xu, Z.; Liu, T.Q.; Min, Y.H.; Li, S. Ameliorative Effects of Selenium on Cadmium-Induced Injury in the Chicken Ovary: Mechanisms of Oxidative Stress and Endoplasmic Reticulum Stress in Cadmium-Induced Apoptosis. Biol. Trace Elem. Res. 2018, 184, 463–473. [Google Scholar] [CrossRef]
- Liu, L.L.; Yang, B.Y.; Cheng, Y.P.; Lin, H.J. Ameliorative Effects of Selenium on Cadmium-Induced Oxidative Stress and Endoplasmic Reticulum Stress in the Chicken Kidney. Biol. Trace Elem. Res. 2015, 167, 308–319. [Google Scholar] [CrossRef]
- Zhang, J.Q.; Zheng, S.F.; Wang, S.C.; Liu, Q.Q.; Xu, S.W. Cadmium-induced oxidative stress promotes apoptosis and necrosis through the regulation of the miR-216a-PI3K/AKT axis in common carp lymphocytes and antagonized by selenium. Chemosphere 2020, 258, 127341. [Google Scholar] [CrossRef]
- Li, J.L.; Jiang, C.Y.; Li, S.; Xu, S.W. Cadmium induced hepatotoxicity in chickens (Gallus domesticus) and ameliorative effect by selenium. Ecotoxicol. Environ. Saf. 2013, 96, 103–109. [Google Scholar] [CrossRef]
- Li, K.; Feng, T.J.; Liu, L.Y.; Liu, H.M.; Huang, K.X.; Zhou, J. Hepatic Proteomic Analysis of Selenoprotein T Knockout Mice by TMT: Implications for the Role of Selenoprotein T in Glucose and Lipid Metabolism. Int. J. Mol. Sci. 2021, 22, 227–233. [Google Scholar] [CrossRef]
- Ghiselli, A.; Serafini, M.; Natella, F.; Scaccini, C. Total antioxidant capacity as a tool to assess redox status: Critical view and experimental data. Free Radical Biol. Med. 2000, 29, 1106–1114. [Google Scholar] [CrossRef] [PubMed]
- Qu, R.J.; Wang, X.H.; Wang, Z.Y.; Wei, Z.B.; Wang, L.S. Metal accumulation and antioxidant defenses in the freshwater fish (Carassius auratus) in response to single and combined exposure to cadmium and hydroxylated multi-walled carbon nanotubes. J. Hazard. Mater. 2014, 275, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.F.; Shen, H.; Wang, X.R.; Wu, J.C.; Xue, Y.Q. Effects of chronic exposure of 2,4-dichlorophenol on the antioxidant system in liver of freshwater fish (Carassius auratus). Chemosphere 2004, 55, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Pillet, M.; Castaldo, G.; De Weggheleire, S.; Bervoets, L.; Blust, R.; De Boeck, G. Limited oxidative stress in common carp (Cyprinus carpio) exposed to a sublethal tertiary (Cu, Cd and Zn) metal mixture. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2019, 218, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Hermesz, E.; Abrahám, M.; Nemcsók, J. Tissue-specific expression of two metallothionein genes in common carp during cadmium exposure and temperature shock. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2001, 128, 457–465. [Google Scholar] [CrossRef]
- Park, J.D.; Liu, Y.P.; Klaassen, C.D. Protective effect of metallothionein against the toxicity of cadmium and other metals. Toxicology 2001, 163, 93–100. [Google Scholar] [CrossRef]
- Knapen, D.; Reynders, H.; Bervoets, L.; Verheyen, E.; Blust, R. Metallothionein gene and protein expression as a biomarker for metal pollution in natural gudgeon populations. Aquat. Toxicol. 2007, 82, 163–172. [Google Scholar] [CrossRef]
- Tan, S.W.; Li, H.; Jin, Y.; Yu, H. In vitro and in vivo effects of sublethal cadmium on the expression of MT2 and ABCC2 genes in grass carp (Ctenopharyngodon idellus). Ecotoxicol. Environ. Saf. 2014, 108, 258–264. [Google Scholar] [CrossRef]
- Klaassen, C.D.; Liu, J.; Choudhuri, S. Metallothionein: An intracellular protein to protect against cadmium toxicity. Annu. Rev. Pharmacol. Toxicol. 1999, 39, 267–294. [Google Scholar] [CrossRef]
- Rotruck, J.T.; Pope, A.L.; Ganther, H.E.; Swanson, A.B.; Hafeman, D.G.; Hoekstra, W.G. Selenium: Biochemical role as a component of glutathione peroxidase. Science 1973, 179, 588–590. [Google Scholar] [CrossRef]
- Della, T.C.; Zaja, R.; Loncar, J.; Smital, T.; Focardi, S.; Corsi, I. Interaction of ABC transport proteins with toxic metals at the level of gene and transport activity in the PLHC-1 fish cell line. Chem.-Biol. Interact. 2012, 198, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.; Ahmad, I.; Maria, V.L.; Serafim, A.; Bebianno, M.J.; Pacheco, M.; Santos, M.A. Hepatic metallothionein concentrations in the golden grey mullet (Liza aurata)-Relationship with environmental metal concentrations in a metal-contaminated coastal system in Portugal. Mar. Environ. Res. 2010, 69, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Payen, L.; Sparfel, L.; Courtois, A.; Vernhet, L.; Guillouzo, A.; Fardel, O. The drug efflux pump MRP2: Regulation of expression in physiopathological situations and by endogenous and exogenous compounds. Cell Biol. Toxicol. 2002, 18, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.; Li, Q.; Li, J.; Cui, Z.B. Molecular analysis, developmental function and heavy metal-induced expression of ABCC5 in zebrafish. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2011, 158, 46–55. [Google Scholar] [CrossRef]
- Luzio, A.; Monteiro, S.M.; Fontaínhas-Fernandes, A.A.; Pinto-Carnide, O.; Matos, M.; Coimbra, A.M. Copper induced upregulation of apoptosis related genes in zebrafish (Danio rerio) gill. Aquat. Toxicol. 2013, 128–129, 183–189. [Google Scholar] [CrossRef]
- Pathak, N.; Mitra, S.; Khandelwal, S. Cadmium induces thymocyte apoptosis via caspase-dependent and caspase-independent pathways. J. Biochem. Mol. Toxicol. 2013, 27, 193–203. [Google Scholar] [CrossRef]
- Lu, Z.Y.; Miao, Y.S.; Muhammad, I.; Tian, E.; Hu, W.J.; Wang, J.; Wang, B.; Li, R.; Li, J.C. Colistin-induced autophagy and apoptosis involves the JNK-Bcl2-Bax signaling pathway and JNK-p53-ROS positive feedback loop in PC-12 cells. Chem.-Biol. Interact. 2017, 277, 62–73. [Google Scholar] [CrossRef]
- Dhanasekaran, D.N.; Reddy, E.P. JNK signaling in apoptosis. Oncogene 2008, 27, 6245–6251. [Google Scholar] [CrossRef]
- Hockenbery, D.M.; Oltvai, Z.N.; Yin, X.M.; Milliman, C.L.; Korsmeyer, S.J. Bcl-2 functions in an antioxidant pathway to prevent apoptosis. Cell 1993, 75, 241–251. [Google Scholar] [CrossRef]
- Li, D.; Ueta, E.; Kimura, T.; Yamamoto, T.; Osaki, T. Reactive oxygen species (ROS) control the expression of Bcl-2 family proteins by regulating their phosphorylation and ubiquitination. Cancer Sci. 2004, 95, 644–650. [Google Scholar] [CrossRef]
- Liu, X.M.; Shi, L.; Wang, W.; Jin, M.H.; Du, H.Y.; Liu, Y.; Sun, L.; Sun, Z.W. Mitochondrial damage induced by cadmium chloride in human hepatoma cell line SMMC-7721. Teratog. Carcinog. Mutagen. 2010, 22, 276–278. [Google Scholar]
- Duan, Y.J.; Duan, J.; Feng, Y.; Huang, X.L.; Fan, W.; Wang, K.Y.; Ouyang, P.; Deng, Y.Q.; Du, Z.J.; Chen, D.F.; et al. Hepatoprotective Activity of Vitamin E and Metallothionein in Cadmium-Induced Liver Injury in Ctenopharyngodon idellus. Oxid. Med. Cell. Longev. 2018, 2018, 9506543. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.Z.; Zhang, Y.; Yang, J.; Liu, Q.; Zhao, R.H.; Hamid, S.; Wang, H.; Xu, S.W.; Zhang, Z.W. Antagonistic effects of selenium against necroptosis injury via adiponectin-necrotic pathway induced by cadmium in heart of chicken. RSC Adv. 2017, 7, 44438–44446. [Google Scholar] [CrossRef]
- Miao, Z.R.; Miao, Z.Y.; Shi, X.; Wu, H.; Yao, Y.J.; Xu, S.W. The antagonistic effect of selenium on lead-induced apoptosis and necroptosis via P38/JNK/ERK pathway in chicken kidney. Ecotoxicol. Environ. Saf. 2022, 231, 113176. [Google Scholar] [CrossRef]
- Zhang, T.T.; Sun, S.Y.; Gavrilović, A.; Li, D.P.; Tang, R. Selenium alleviates cadmium-induced oxidative stress, endoplasmic reticulum stress, and apoptosis in L8824 cells. Ecotoxicol. Environ. Saf. 2023, 262, 115337. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Lin, J.; Ge, J.; Wang, L.L.; Li, N.; Sun, X.T.; Cao, H.B.; Li, J.L. Selenium triggers Nrf2-mediated protection against cadmium-induced chicken hepatocyte autophagy and apoptosis. Toxicol. In Vitro 2017, 44, 349–356. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).