
Zeta Diversity Reveals the Small Wetland Complex Promotes Biodiversity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
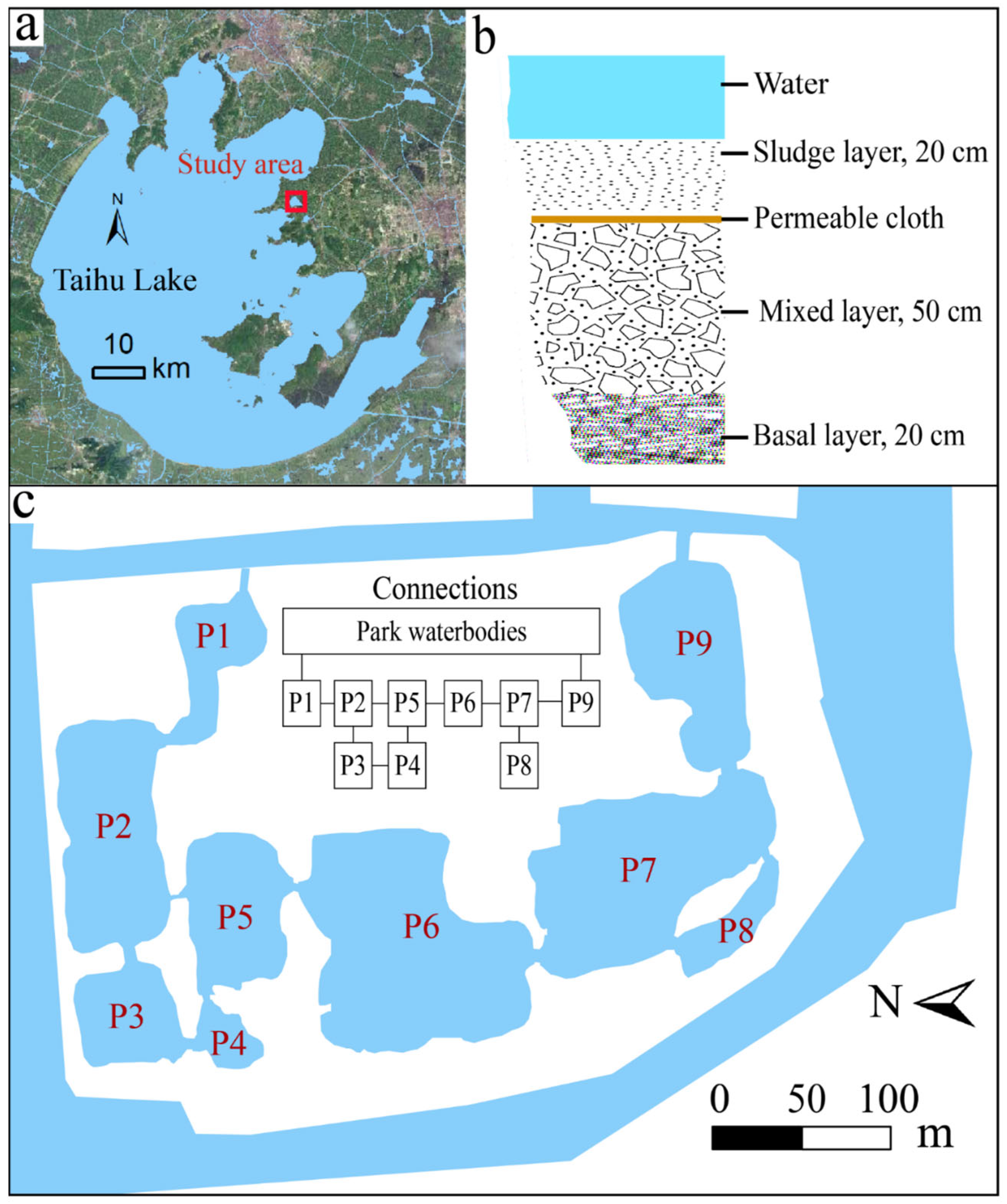
2.1. Study Area
2.2. Experimental Design and Sampling
2.3. Data Analysis
3. Results
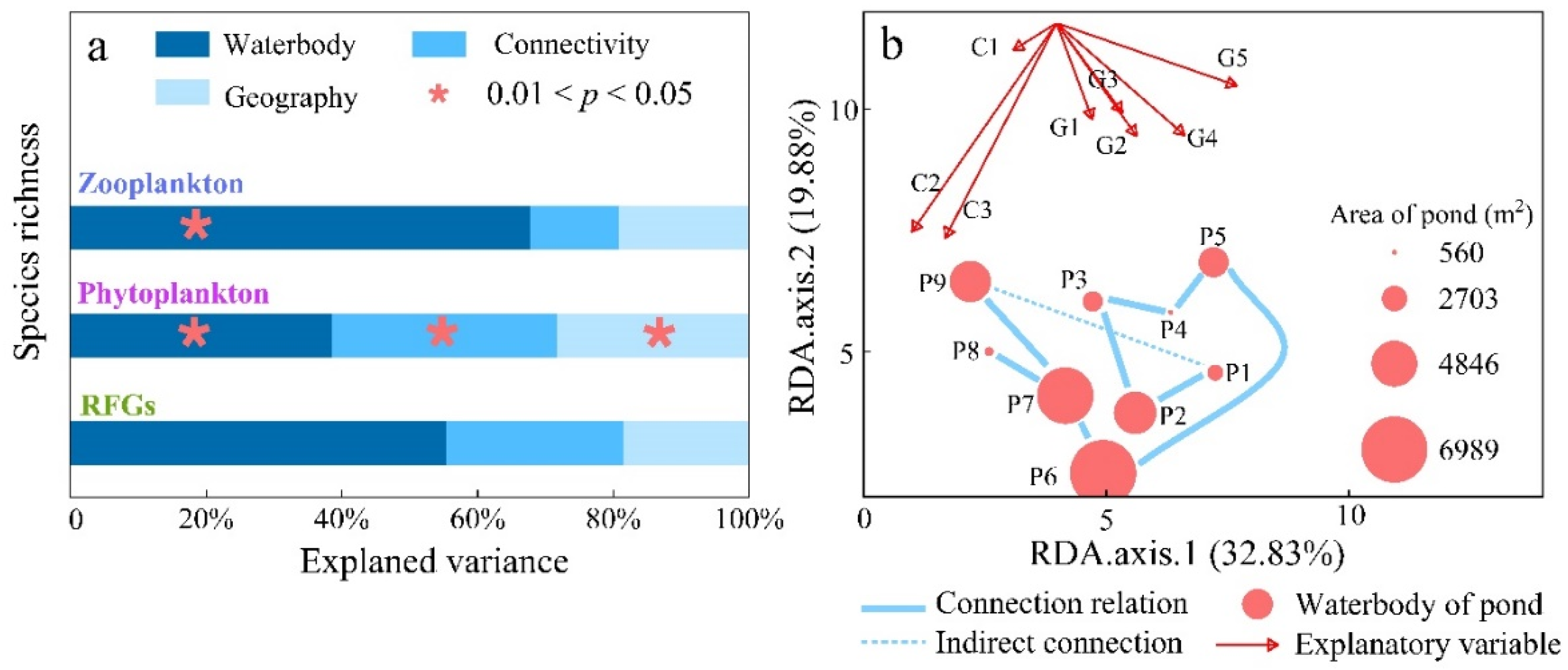
3.1. Connectivity Drives Habitat and Community Richness
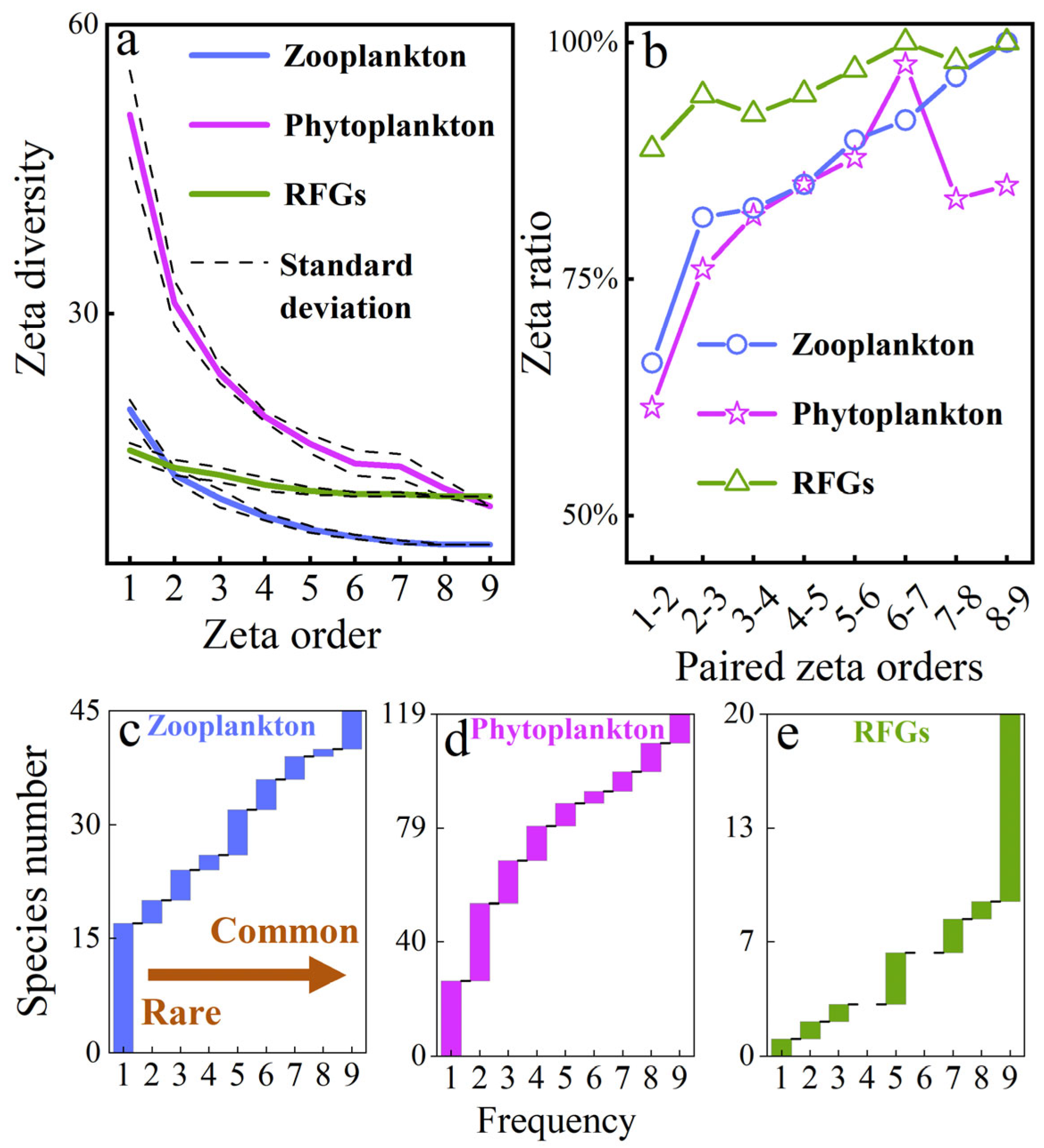
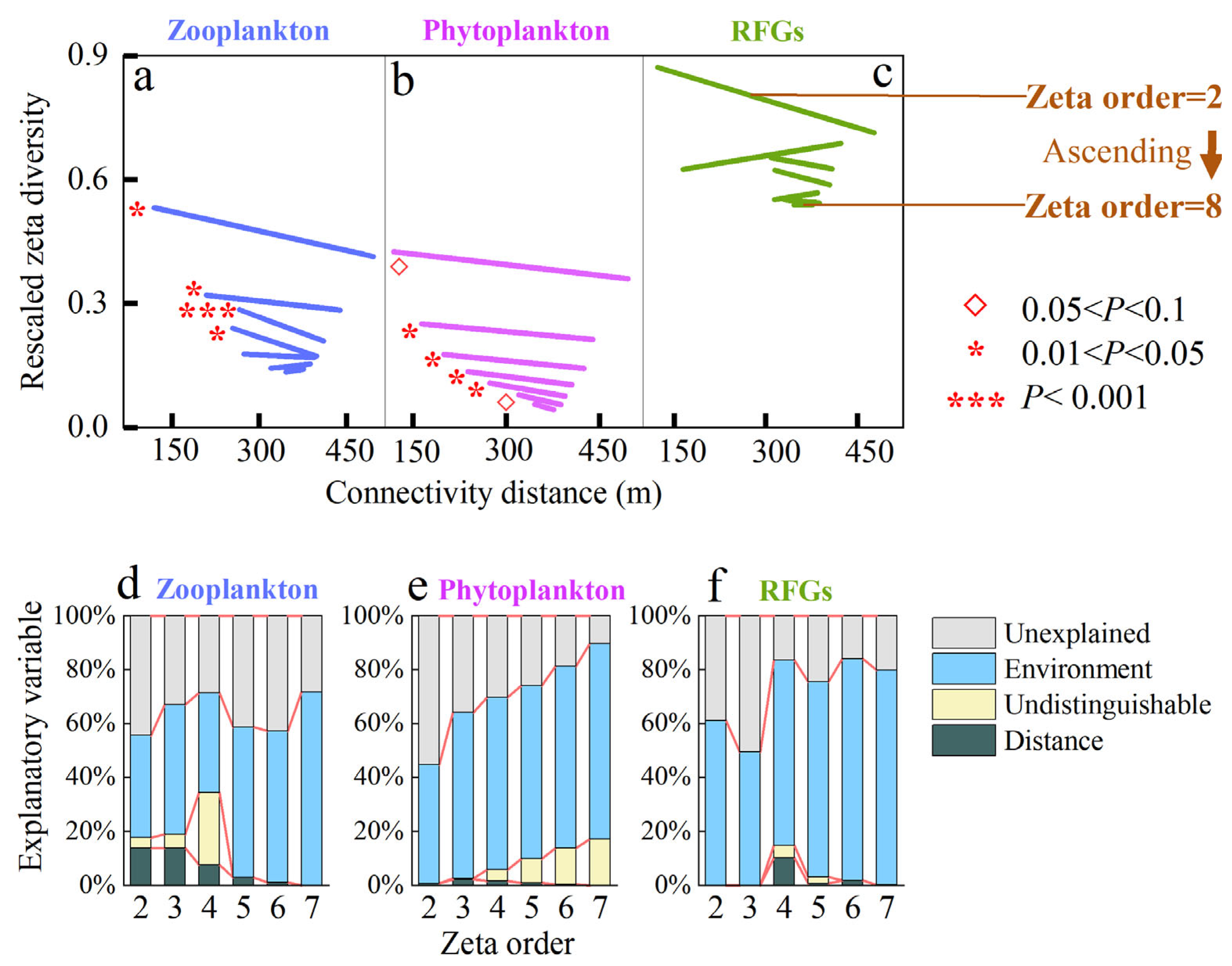
3.2. Connectivity Drives Species Turnover via Differential Mechanisms
4. Discussion
4.1. Zeta Diversity Quantified the Effect of Connectivity on Community Assembly
4.2. The Small Wetland Complex Could Efficiently Enhance Species Richness
4.3. The Ecological Effects of Connectivity May Benefit Rare Species Conservation
5. Conclusions
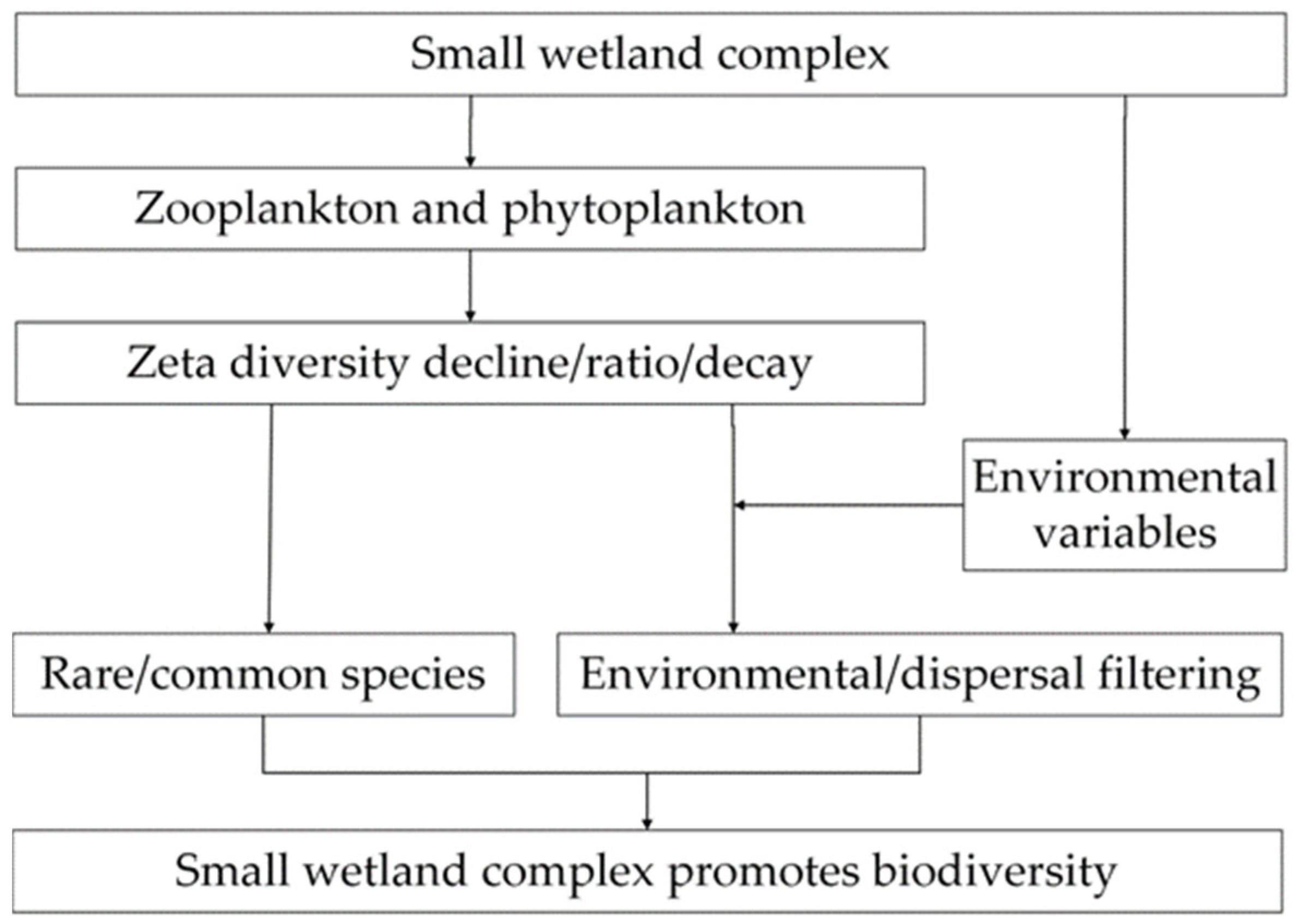
- The plankton community in the small wetland complex comprises 46 zooplankton and 119 phytoplankton species, with the species richness primarily influenced by the waterbody properties (p < 0.05). This suggests that the small wetland complex studied provides a diverse habitat capable of accommodating various types of plankton. Conversely, the impact of waterbody properties on plankton reflects an environmental filtering effect during community assembly.
- Zeta diversity decay describes the distance-dependent effect of zooplankton and phytoplankton (p < 0.05), indicating that hydrological connectivity distance influences plankton zeta diversity. This phenomenon of distance attenuation reflects a dispersal filtering effect during community assembly.
- Relatively rare species outnumber relatively common species in the small wetland complex, suggesting that increasing small wetlands facilitates capturing these relatively rare common species. Based on zeta diversity decline and ratio, rare species dominate community turnover, indicating their positive contribution to enhancing both species richness and zeta diversity within the small wetland complex.
- Furthermore, comparing environmental filtration and dispersal filtration reveals that environmental filtration has a stronger influence than dispersal filtration in this small wetland complex.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Richardson, S.J.; Clayton, R.; Rance, B.D.; Broadbent, H.; McGlone, M.S.; Wilmshurst, J.M. Small wetlands are critical for safeguarding rare and threatened plant species. Appl. Veg. Sci. 2015, 18, 230–241. [Google Scholar] [CrossRef]
- Zhao, H.; Chen, J.Q.; Chen, X.; Xiong, Y.Y.; An, S.Q. Conservation and Management of Small and Micro Wetlands. Wetl. Sci. Manag. 2018, 14, 24–28. [Google Scholar]
- Dahl, T.E. Status and Trends of Prairie Wetlands in the United States 1997 to 2009; US Fish and Wildlife Service: Washington, DC, USA, 2014. [Google Scholar]
- Jia, X.H.; Wu, M.; Shao, X.X.; Jiang, K.Y. Community characteristics of phytoplanktons in closed ponds of Xixi wetland and related affecting factors. Chin. J. Ecol. 2010, 29, 1743–1748. [Google Scholar]
- Biggs, J.; Williams, P.; Whitfield, M.; Nicolet, P.; Weatherby, A. 15 years of pond assessment in Britain: Results and lessons learned from the work of Pond Conservation. Aquat. Conserv. Mar. Freshw. Ecosyst. 2005, 15, 693–714. [Google Scholar] [CrossRef]
- Convention on Wetlands. Report No. XIII.21: Conservation and Management of Small Wetlands. 2018. Available online: https://www.ramsar.org/document/resolution-xiii21-conservation-and-management-small-wetlands (accessed on 29 October 2018).
- Convention on Wetlands. Draft Resolution on Guidance on Conservation and Management of Small Wetlands. 2022. Available online: https://www.ramsar.org/document/sc59-doc244-draft-resolution-guidance-conservation-management-small-wetlands (accessed on 23 April 2021).
- Luo, Y.; Wu, J.; Wang, X.; Wang, Z.; Zhao, Y. Can policy maintain habitat connectivity under landscape fragmentation? A case study of Shenzhen, China. Sci. Total Environ. 2020, 715, 136829. [Google Scholar] [CrossRef] [PubMed]
- Perkin, J.S.; Gido, K.B.; Cooper, A.R.; Turner, T.F.; Osborne, M.J.; Johnson, E.R.; Mayes, K.B. Fragmentation and dewatering transform Great Plains stream fish communities. Ecol. Monogr. 2015, 85, 73–92. [Google Scholar] [CrossRef]
- Ginders, M.A.; Collier, K.J.; Duggan, I.C.; Hamilton, D.P. Influence of Hydrological Connectivity on Plankton Communities in Natural and Reconstructed Side-Arms of a Large New Zealand River. River Res. Appl. 2016, 32, 1675–1686. [Google Scholar] [CrossRef]
- Si, X.; Cadotte, M.W.; Davies, T.J.; Antonelli, A.; Ding, P.; Svenning, J.C.; Faurby, S. Phylogenetic and functional clustering illustrate the roles of adaptive radiation and dispersal filtering in jointly shaping late-Quaternary mammal assemblages on oceanic islands. Ecol. Lett. 2022, 25, 1250–1262. [Google Scholar] [CrossRef] [PubMed]
- Chust, G.; Villarino, E.; Chenuil, A.; Irigoien, X.; Bizsel, N.; Bode, A.; Broms, C.; Claus, S.; Fernandez de Puelles, M.L.; Fonda-Umani, S.; et al. Dispersal similarly shapes both population genetics and community patterns in the marine realm. Sci. Rep. 2016, 6, 28730. [Google Scholar] [CrossRef]
- Xiao, R.; Wang, Q.; Zhang, M.X.; Pan, W.B.; Wang, J.J. Plankton distribution patterns and the relationship with environmental gradients and hydrological connectivity of wetlands in the Yellow River Delta. Ecohydrol. Hydrobiol. 2020, 20, 584–596. [Google Scholar] [CrossRef]
- Salinas-de-Leon, P.; Jones, T.; Bell, J.J. Successful determination of larval dispersal distances and subsequent settlement for long-lived pelagic larvae. PLoS ONE 2012, 7, e32788. [Google Scholar] [CrossRef]
- Jeliazkov, A.; Lorrillière, R.; Besnard, A.; Garnier, J.; Silvestre, M.; Chiron, F. Cross-scale effects of structural and functional connectivity in pond networks on amphibian distribution in agricultural landscapes. Freshw. Biol. 2019, 64, 997–1014. [Google Scholar] [CrossRef]
- Hui, C.; McGeoch, M.A. Zeta diversity as a concept and metric that unifies incidence-based biodiversity patterns. Am. Nat. 2014, 184, 684–694. [Google Scholar] [CrossRef] [PubMed]
- Latombe, G.; Hui, C.; McGeoch, M.A. Multi-site generalised dissimilarity modelling: Using zeta diversity to differentiate drivers of turnover in rare and widespread species. Methods Ecol. Evol. 2017, 8, 431–442. [Google Scholar] [CrossRef]
- Nekola, J.C.; McGill, B.J. Scale dependency in the functional form of the distance decay relationship. Ecography 2014, 37, 309–320. [Google Scholar] [CrossRef]
- Simons, A.L.; Mazor, R.; Stein, E.D.; Nuzhdin, S. Using alpha, beta, and zeta diversity in describing the health of stream-based benthic macroinvertebrate communities. Ecol. Appl. 2019, 29, e01896. [Google Scholar] [CrossRef]
- Hu, H.J.; Wei, Y.X. The Freshwater Algae of China—Systematics, Taxonomy and Ecology; Scientific & Technical Publishers: Beijing, China, 2006. [Google Scholar]
- Weng, J.Z.; Xu, H.S. Atlas of Freshwater Planktonic Alage in China; Science & Technology Press: Beijing, China, 2010. [Google Scholar]
- Reynolds, C.S.; Huszar, V.; Kruk, C.; Naselli-Flores, L.; Melo, S. Towards a functional classification of the freshwater phytoplankton. J. Plankton Res. 2002, 24, 417–428. [Google Scholar] [CrossRef]
- Blanchet, F.G.; Legendre, P.; Borcard, D. Modelling directional spatial processes in ecological data. Ecol. Model. 2008, 215, 325. [Google Scholar] [CrossRef]
- Zorzal-Almeida, S.; Soininen, J.; Bini, L.M.; Bicudo, D.C. Local environment and connectivity are the main drivers of diatom species composition and trait variation in a set of tropical reservoirs. Freshw. Biol. 2017, 62, 1551–1563. [Google Scholar] [CrossRef]
- McGeoch, M.A.; Latombe, G.; Andrew, N.R.; Nakagawa, S.; Nipperess, D.A.; Roige, M.; Marzinelli, E.M.; Campbell, A.H.; Verges, A.; Thomas, T.; et al. Measuring continuous compositional change using decline and decay in zeta diversity. Ecology 2019, 100, e02832. [Google Scholar] [CrossRef] [PubMed]
- Wilson, H.D. Natural History of Banks Peninsula; Canterbury University Press: Christchurch, New Zealand, 2009. [Google Scholar]
- Thompson, P.A.; Bonham, P.; Waite, A.M.; Clementson, L.A.; Cherukuru, N.; Hassler, C.; Doblin, M.A. Contrasting oceanographic conditions and phytoplankton communities on the east and west coasts of Australia. Deep-Sea Res. Part Ii-Top. Stud. Oceanogr. 2011, 58, 645–663. [Google Scholar] [CrossRef]
- Eros, T.; Czegledi, I.; Toth, R.; Schmera, D. Multiple stressor effects on alpha, beta and zeta diversity of riverine fish. Sci. Total Environ. 2020, 748, 141407. [Google Scholar] [CrossRef]
- Leibold, M.A.; Holyoak, M.; Mouquet, N.; Amarasekare, P.; Chase, J.M.; Hoopes, M.F.; Holt, R.D.; Shurin, J.B.; Law, R.; Tilman, D.; et al. The metacommunity concept: A framework for multi-scale community ecology. Ecol. Lett. 2004, 7, 601–613. [Google Scholar] [CrossRef]
- Limberger, R.; Birtel, J.; Farias, D.D.S.; Matthews, B. Ecosystem flux and biotic modification as drivers of metaecosystem dynamics. Ecology 2017, 98, 1082–1092. [Google Scholar] [CrossRef]
- Carwardine, J.; Klein, C.J.; Wilson, K.A.; Pressey, R.L.; Possingham, H.P. Hitting the target and missing the point: Target-based conservation planning in context. Conserv. Lett. 2009, 2, 3–10. [Google Scholar] [CrossRef]
- Fischer, J.; Lindenmayer, D.B. Small patches can be valuable for biodiversity conservation: Two case studies on birds in southeastern Australia. Biol. Conserv. 2002, 106, 129–136. [Google Scholar] [CrossRef]
- Williams, P.; Biggs, J.; Crowe, A.; Murphy, J.; Nicolet, P.; Weatherby, A.; Dunbar, M. Countryside Survey: Ponds Report from 2007. 2010. Available online: https://www.ceh.ac.uk/our-science/projects/countryside-survey (accessed on 1 January 2010).
- Deane, D.C.; Fordham, D.A.; He, F.L.; Bradshaw, C.J.A. Future extinction risk of wetland plants is higher from individual patch loss than total area reduction. Biol. Conserv. 2017, 209, 27–33. [Google Scholar] [CrossRef]
- Singh, M.; Sinha, R.; Mishra, A.; Babu, S. Wetlandscape (dis)connectivity and fragmentation in a large wetland (Haiderpur) in west Ganga plains, India. Earth Surf. Process. Landf. 2022, 47, 1872–1887. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Zheng, F.; Li, D.; Zhou, R.; Jv, W.; Leng, X.; Xia, L.; An, S. Zeta Diversity Reveals the Small Wetland Complex Promotes Biodiversity. Water 2024, 16, 476. https://doi.org/10.3390/w16030476
Chen J, Zheng F, Li D, Zhou R, Jv W, Leng X, Xia L, An S. Zeta Diversity Reveals the Small Wetland Complex Promotes Biodiversity. Water. 2024; 16(3):476. https://doi.org/10.3390/w16030476
Chicago/Turabian StyleChen, Jun, Fuchao Zheng, Dianpeng Li, Rixiu Zhou, Wenming Jv, Xin Leng, Lu Xia, and Shuqing An. 2024. "Zeta Diversity Reveals the Small Wetland Complex Promotes Biodiversity" Water 16, no. 3: 476. https://doi.org/10.3390/w16030476
APA StyleChen, J., Zheng, F., Li, D., Zhou, R., Jv, W., Leng, X., Xia, L., & An, S. (2024). Zeta Diversity Reveals the Small Wetland Complex Promotes Biodiversity. Water, 16(3), 476. https://doi.org/10.3390/w16030476