Cyanobacterial Nitrogen Fixation Influences the Nitrogen Removal Efficiency in a Constructed Wetland





Abstract
:1. Introduction
2. Materials and Methods
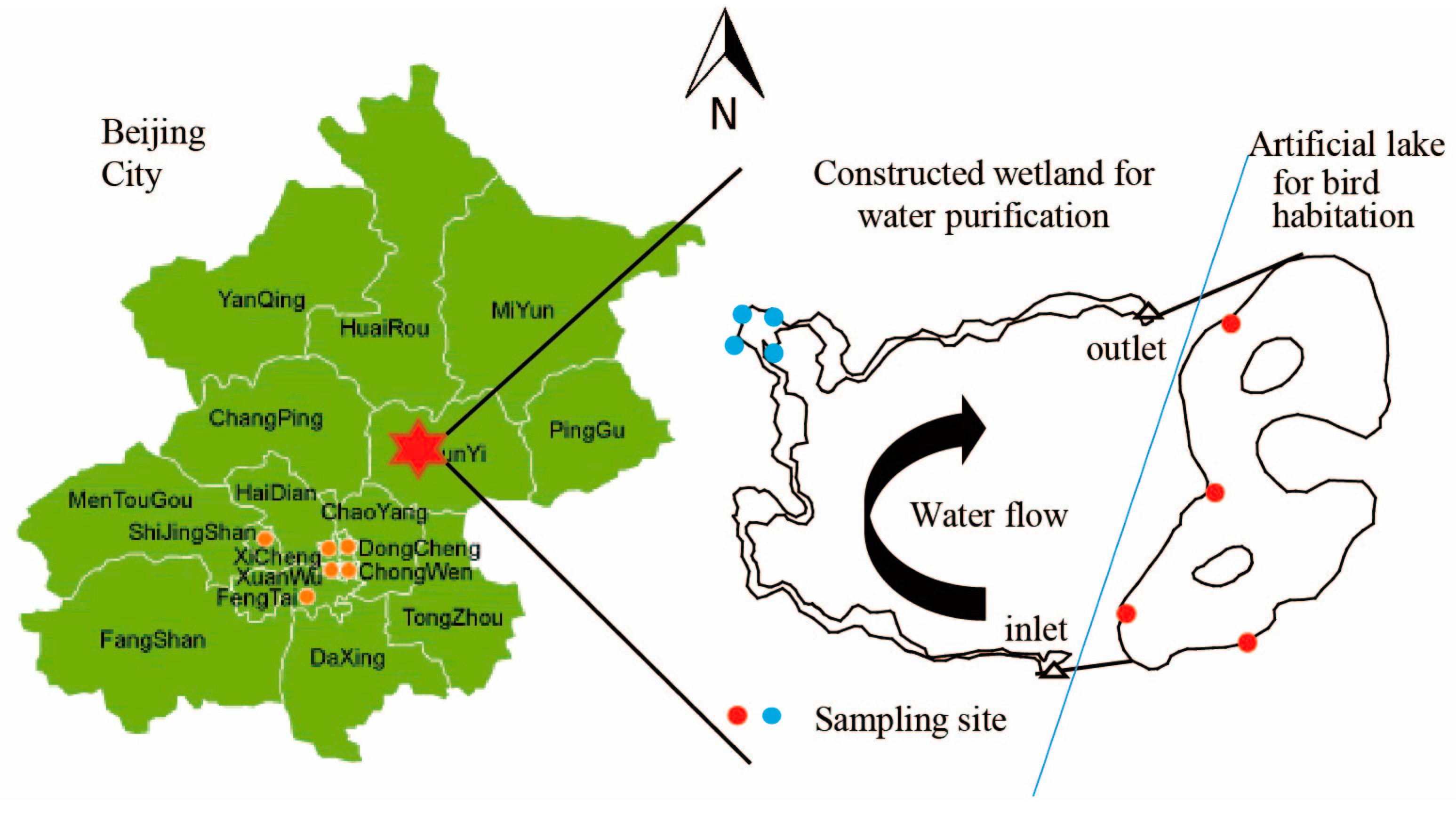
2.1. Study Site
2.2. Sampling
2.3. Concentration of Nitrogen Fractions
2.4. Nitrogen Fixation Rate
2.5. Statistical Analysis
3. Results
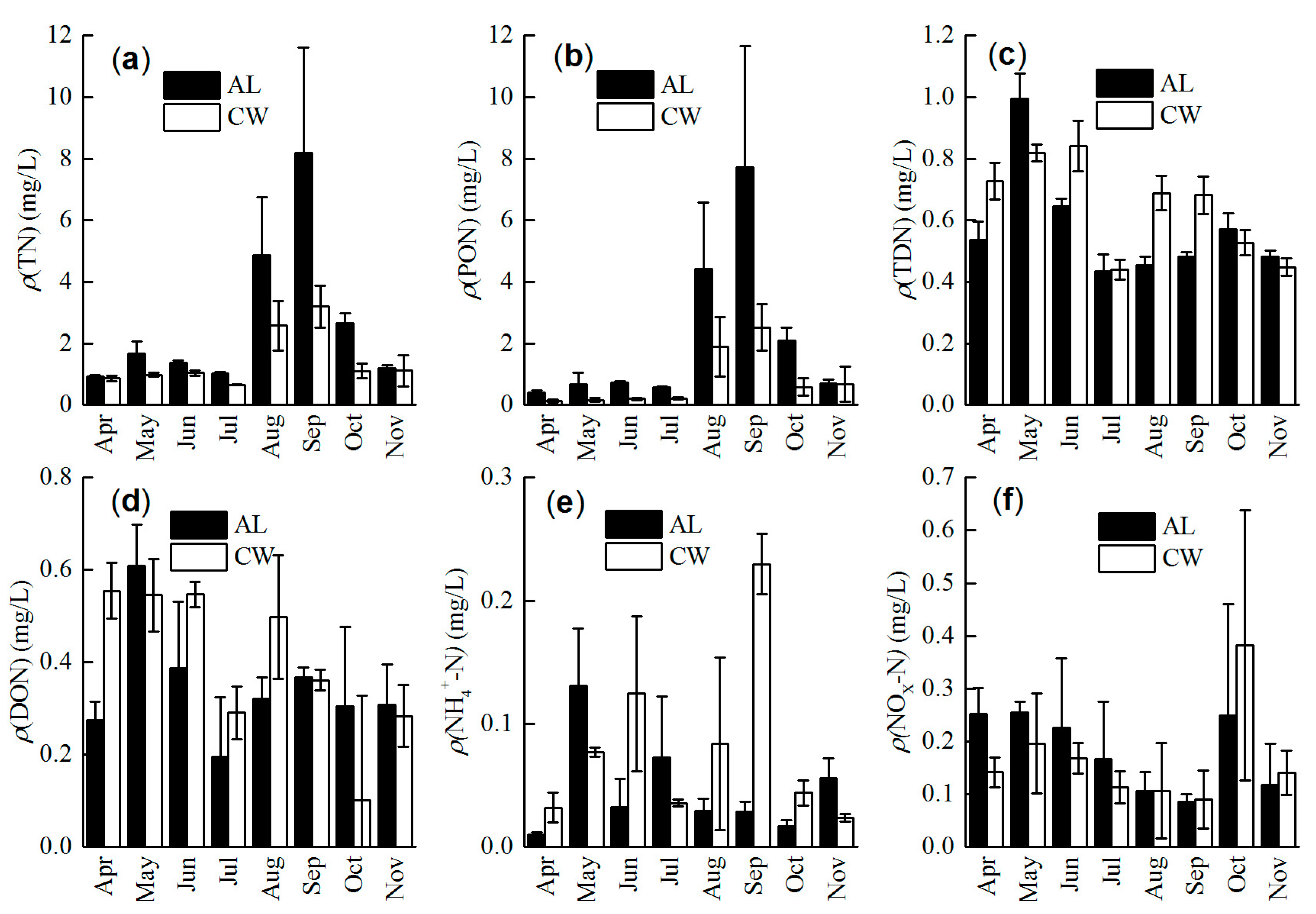
3.1. Concentration of Nitrogen Forms
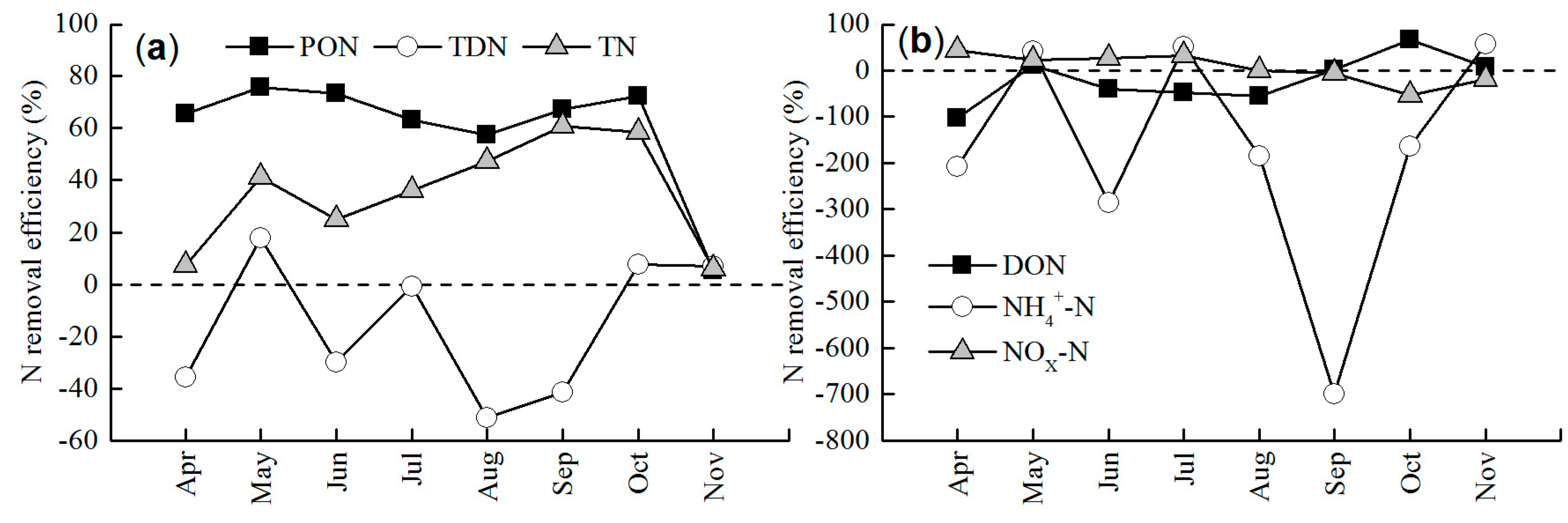
3.2. Nitrogen Removal Efficiency by CW
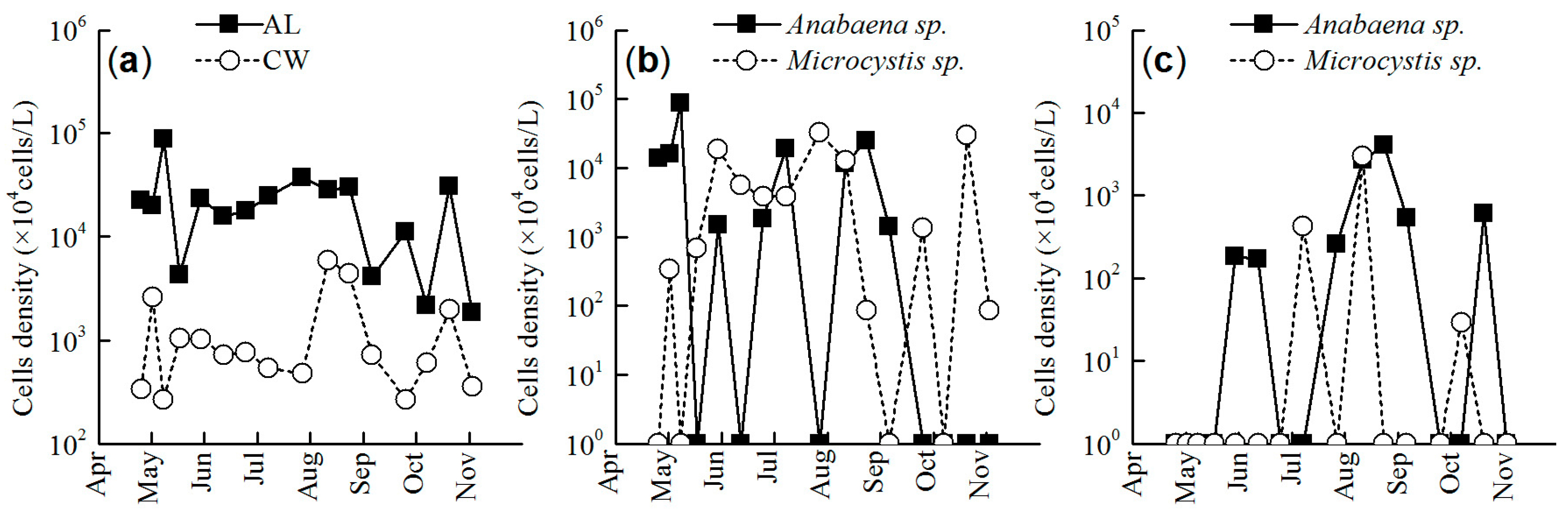
3.3. Cell Density of N-Fixing and Non-Nitrogen Fixing Cyanobacteria
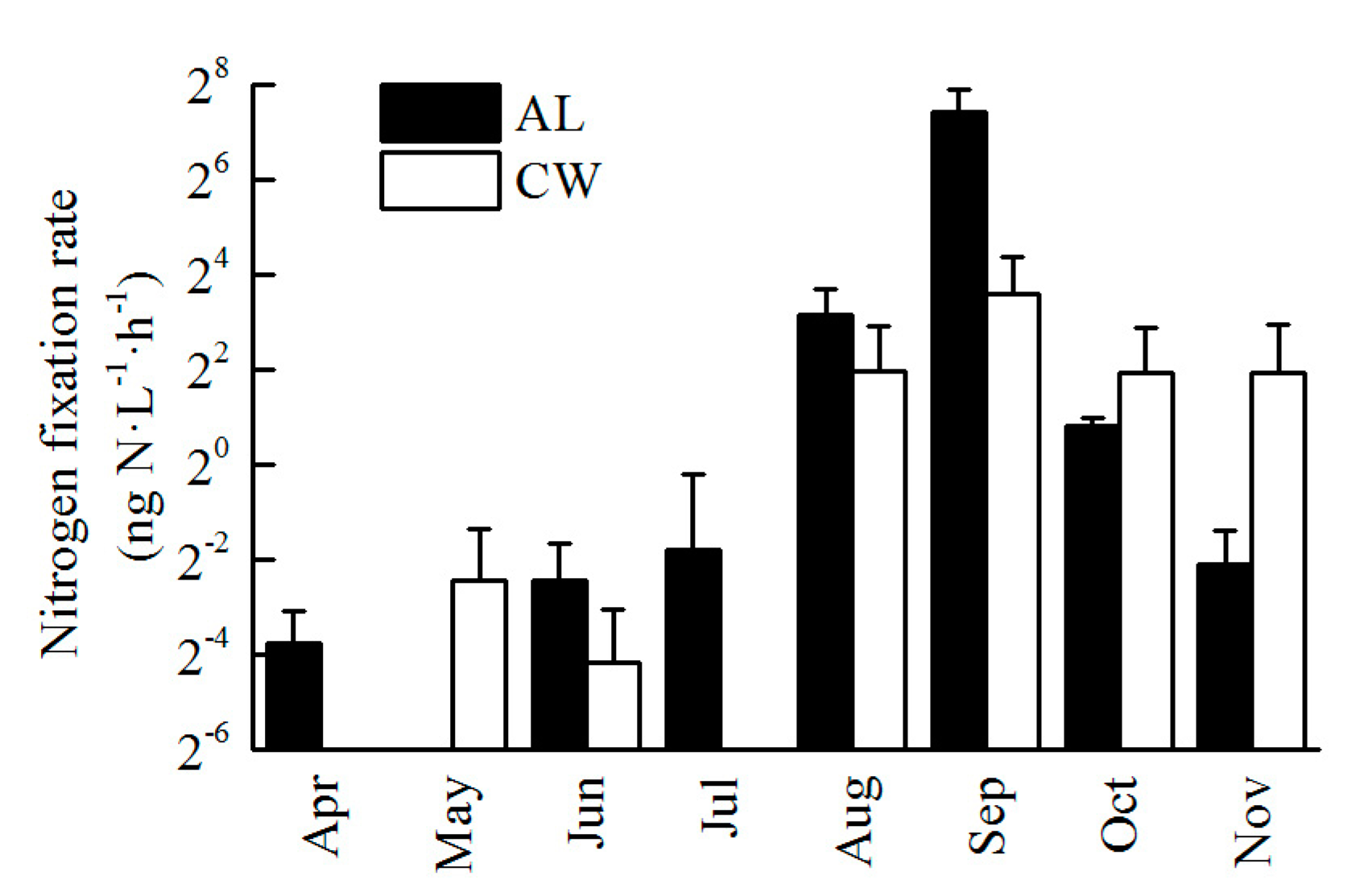
3.4. Nitrogen Fixation Rate
4. Discussion
4.1. Dynamics of Nitrogen Removal Efficiency
4.2. Dynamics of N-Fixing and Non-N-Fixing Cyanobacteria
4.3. Relationships among Nitrogen Fixation, N-Fixing Cyanobacteria, and PON Removal
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Vymazal, J. Removal of nutrients in various types of constructed wetlands. Sci. Total Environ. 2007, 380, 48–65. [Google Scholar] [CrossRef] [PubMed]
- Vergeles, Y.; Vystavna, Y.; Ishchenko, A.; Rybalka, I.; Marchand, L.; Stolberg, F. Assessment of treatment efficiency of constructed wetlands in East Ukraine. Ecol. Eng. 2015, 83, 159–168. [Google Scholar] [CrossRef]
- Saeed, T.; Sun, G. A review on nitrogen and organics removal mechanisms in subsurface flow constructed wetlands: Dependency on environmental parameters, operating conditions and supporting media. J. Environ. Manag. 2012, 112, 429–448. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhou, Q.; He, R. Mechanism of nitrogen and phosphorus removal in free-water surface constructed wetland. Ecol. Environ. 2004, 13, 98–101. [Google Scholar]
- Taylor, G.D.; Fletcher, T.D.; Wong, T.H.F.; Breen, P.F.; Duncan, H.P. Nitrogen composition in urban runoff-implications for stormwater management. Water Res. 2005, 39, 1982–1989. [Google Scholar] [CrossRef] [PubMed]
- Dunne, E.J.; Coveney, M.F.; Marzolf, E.R.; Hoge, V.R.; Conrow, R.; Naleway, R.; Lowe, E.F.; Battoe, L.E.; Inglett, P.W. Nitrogen dynamics of a large-scale constructed wetland used to remove excess nitrogen from eutrophic lake water. Ecol. Eng. 2013, 61, 224–234. [Google Scholar] [CrossRef]
- Taylor, P.G.; Wieder, W.R.; Weintraub, S.; Cohen, S.; Cleveland, C.C.; Townsend, A.R. Organic forms dominate hydrologic nitrogen export from a lowland tropical watershed. Ecology 2015, 96, 1229–1241. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Kuschk, P.; Brix, H.; Vymazal, J.; Dong, R. Development of constructed wetlands in performance intensifications for wastewater treatment: A nitrogen and organic matter targeted review. Water Res. 2014, 57, 40–55. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.G.; Fletcher, T.D.; Sun, G.Z. Nitrogen removal in constructed wetland systems. Eng. Life Sci. 2009, 9, 11–22. [Google Scholar] [CrossRef]
- Kalff, J. Limnology: Inland Water Eosystems; Prentice Hall: Upper Saddle River, NJ, USA, 2002. [Google Scholar]
- Ye, L.L.; Zhang, M.; Kong, F.X.; Yang, Z.; Shi, X.L.; Yan, D.Z.; Liu, B. Progress and prospect of research on cyanobacteria nitrogen fixing in aquatic ecosystem. J. Lake Sci. 2014, 26, 9–18. [Google Scholar] [CrossRef]
- Yadav, R.K.; Abraham, G.; Singh, Y.V.; Singh, P.K. Advancements in the Utilization of Azolla-anabaena System in Relation to Sustainable Agricultural Practices. Proc. Indian Natl. Sci. Acad. 2014, 80, 301–306. [Google Scholar] [CrossRef]
- Cui, L.J.; Li, W.; Zhang, M.Y.; Zhao, X.S.; Zhang, Y.; Wang, Y.F. Effects of waterfowl sewage treatment by integrated artificial wetland running for 2 years. Trans. CSAE 2011, 27, 234–240. [Google Scholar]
- Zhang, Y.; Cui, L.; Li, W.; Zhang, M.Y.; Zhao, X.S.; Wang, Y.F. Performance evaluation of an integrated constructed wetland used to treat a contaminated aquatic environment. Wetl. Ecol. Manag. 2014, 22, 493–507. [Google Scholar] [CrossRef]
- Hu, H.J.; Wei, Y.X. The Freshwater Algae of China: Systematics, Taxonomy and Ecology; Science Press: Beijing, China, 2006. [Google Scholar]
- China Meteorological Data Network. Available online: http://data.cma.cn/site/index.html (accessed on 17 September 2017).
- Montoya, J.P.; Voss, M.; Kahler, P.; Capone, D.G. A Simple, High-Precision, High-Sensitivity Tracer Assay for N2 Fixation. Appl. Environ. Microbiol. 1996, 62, 986–993. [Google Scholar] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016; Available online: https://www.R-project.org/ (accessed on 17 September 2017).
- Zhang, Y.; Huo, S.; Zan, F.; Xi, B.D.; Zhang, J.T. Dissolved organic nitrogen (DON) in seventeen shallow lakes of Eastern China. Environ. Earth Sci. 2015, 74, 4011–4021. [Google Scholar] [CrossRef]
- Moisander, P.H.; Ochiai, M.; Lincoff, A. Nutrient limitation of Microcystis aeruginosa in northern California Klamath River reservoirs. Harmful Algae 2009, 8, 889–897. [Google Scholar] [CrossRef]
- Beversdorf, L.J.; Miller, T.R.; McMahon, K.D. The role of nitrogen fixation in cyanobacterial bloom toxicity in a temperate, eutrophic lake. PLoS ONE 2013, 8, e56103. [Google Scholar] [CrossRef] [PubMed]
- Tsujimura, S.; Ishikawa, K.; Tsukada, H. Effect of temperature on growth of the cyanobacterium Aphanizomenon flos-aquae in Lake Biwa and Lake Yogo. Phycol. Res. 2001, 49, 275–280. [Google Scholar] [CrossRef]
- Qin, B.Q.; Xu, H.; Dong, B.L. The Principle and Practice of Eutrophic Lake Restoration and Management; Higher Education Press: Beijing, China, 2011. [Google Scholar]
- Liu, F.; Feng, M.; Shang, L.; He, Y.Z.; Li, W.C. Effects of temperature on the growth and generation of extracellular organic matter of Microcystis aeruginosa and Anabaena sp. J. Great Lakes Sci. 2014, 26, 780–788. [Google Scholar] [CrossRef]
- Yin, Z.K.; Li, Z.; Wang, S.; Guo, J.S.; Xiao, Y.; Liu, J.; Zhang, P. Effect of Light and Temperature on Growth Kinetics of Anabaena flosaquae Under Phosphorus Limitation. Environ. Sci. 2015, 36, 963–968. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, M.; Wang, R.N. The temporal and spation variation of the cyanobacteria which caused the water bloom in the Dianchi Lake, Kunming, China. J. Yunnan Univ. Nat. Sci. 2005, 27, 272–276. [Google Scholar]
- Lv, X.J.; Zhu, J.; Meng, L. Pilot study on diversity of cyanobacteria bloom in Erhai Lake. Envion. Sci. Surv. 2010, 29, 32–35. [Google Scholar]
- Paerl, H.W.; Xu, H.; Hall, N.S.; Zhu, G.W.; Qin, B.Q.; Wu, Y.L.; Rossignol, K.L.; Dong, L.H.; McCarthy, M.J.; Joyner, A.R. Controlling cyanobacterial blooms in hypertrophic lake Taihu, China: Will nitrogen reductions cause replacement of non-N2-fixing by N2-fixing taxa? PLoS ONE 2014, 9, e113123. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, S.W.; Farnsley, S.E.; LeCleir, G.R.; Layton, A.C.; Satchwell, M.F.; DeBruyn, J.M.; Boyer, G.L.; Zhu, G.W.; Paerl, H.W. The relationships between nutrients, cyanobacterial toxins and the microbial community in Taihu (Lake Tai), China. Harmful Algae 2011, 10, 207–215. [Google Scholar] [CrossRef]
- Patoine, A.; Graham, M.D.; Leavitt, P.R. Spatial variation of nitrogen fixation in lakes of the northern Great Plains. Limnol. Oceanogr. 2006, 51, 1665–1677. [Google Scholar] [CrossRef]
- Bonnet, S.I.C.; Biegala, I.C.; Dutrieux, P.; Slemons, L.O.; Capone, D.G. Nitrogen fixation in the western equatorial Pacific: Rates, diazotrophic cyanobacterial size class distribution, and biogeochemical significance. Glob. Biogeochem. Cycles 2009, 23, GB3012. [Google Scholar] [CrossRef]
- Ohlendieck, U.; Stuhr, A.; Siegmund, H. Nitrogen fixation by diazotrophic cyanobacteria in the Baltic Sea and transfer of the newly fixed nitrogen to picoplankton organisms. J. Mar. Syst. 2000, 25, 213–219. [Google Scholar] [CrossRef]
- Schindler, D.W.; Hecky, R.E.; Findlay, D.L.; Stainton, M.P.; Parker, B.R.; Paterson, M.J.; Beaty, K.G.; Lyng, M.; Kasian, S.E.M. Eutrophication of lakes cannot be controlled by reducing nitrogen input: Results of a 37-year whole-ecosystem experiment. Proc. Natl. Acad. Sci. USA 2008, 105, 11254–11258. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ρ(PON) | ρ(DON) | ρ(NH4+-N) | ρ(NOX-N) | Total Phytoplankton | Anabaena sp. | Microcystis sp. | |
|---|---|---|---|---|---|---|---|
| RNfix in AL | 0.648 *** | 0.662 *** | −0.078 | −0.244 | 0.309 | 0.467 ** | −0.259 |
| RNfix in CW | 0.739 *** | 0.618 *** | 0.417 * | −0.107 | 0.489 ** | 0.533 ** | −0.223 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Jia, X.; Yan, L.; Wang, J.; Kang, X.; Cui, L. Cyanobacterial Nitrogen Fixation Influences the Nitrogen Removal Efficiency in a Constructed Wetland. Water 2017, 9, 865. https://doi.org/10.3390/w9110865
Zhang X, Jia X, Yan L, Wang J, Kang X, Cui L. Cyanobacterial Nitrogen Fixation Influences the Nitrogen Removal Efficiency in a Constructed Wetland. Water. 2017; 9(11):865. https://doi.org/10.3390/w9110865
Chicago/Turabian StyleZhang, Xiaodong, Xin Jia, Liang Yan, Jinzhi Wang, Xiaoming Kang, and Lijuan Cui. 2017. "Cyanobacterial Nitrogen Fixation Influences the Nitrogen Removal Efficiency in a Constructed Wetland" Water 9, no. 11: 865. https://doi.org/10.3390/w9110865
APA StyleZhang, X., Jia, X., Yan, L., Wang, J., Kang, X., & Cui, L. (2017). Cyanobacterial Nitrogen Fixation Influences the Nitrogen Removal Efficiency in a Constructed Wetland. Water, 9(11), 865. https://doi.org/10.3390/w9110865